
Abstract
Background
Host-parasite interaction studies represent the dependency of the parasites on the host through various models. Gill infesting copepods, monogeneans and isopods served as excellent models to study the details of host-parasite interaction. In the present investigation, Wallago attu (Bloch & Schneider, 1801) was extensively studied for histopathological damage caused by ectoparasites like monogeneans (Thaparocleidus indicus, Thaparocleidus wallagonius and Mizelleus indicus), copepod (Ergasilus malnadensis) and an isopod (Alitropus typus) during August 2017 to February 2018. Both the control (least infested) and severely infested gills were examined through light microscope to approximate the intensity of damage caused by above ectoparasites.
Results
Pathological effects of ectoparasites displayed a fusion of secondary lamella, hyperplasia of the gill filaments and the epithelial cells, propagation of bronchial tips, thinning of the central axis, deshaping, shortening and fusion of secondary gill lamellae epithelium, severe degenerative and necrotic changes in gill filaments and secondary lamellae, curling of secondary lamellae and mucous cells proliferation.
Conclusions
Ectoparasites cause severe damage to gills of W. attu, thereby diminishing the gill breathing capacity of the fish. Timely pathological studies can reveal the extent of damage caused by the parasites and thus various diagnostic programs and best management practices can be implemented in aquaculture for the better yield.
Similar content being viewed by others
Background
Ectoparasitic infestation of wild and farmed freshwater and marine fishes caused by crustaceans, monogeneans, protozoans and isopods may severely affect the host health (Johnson et al., 2004; Boxshall, 2005; Shinn et al., 2015; Modi and Vankara, 2021). Branchiuran copepod parasites (Argulus) and ergasilids can cause severe impact on the freshwater fish production (Møller, 2009). Ergasilids are found mainly on the gills of fish which are specialized to perform various functions like gaseous-exchange, hormone production, circulation, ionic and acid–base balance and excretion (Pelster and Bagatto, 2010; Lima et al., 2013; Pádua et al., 2015). The infested gill tissues hinder the water flow and diminish the capability of the fish to sustain normal oxygen uptake (Ojha and Hughes, 2001). Monogeneans do not require any piercing mouthparts to feed on gill blood and need much effort to damage thin walls of the secondary lamellae, perhaps by suction or by applying of digestive secretions. Till date commendable work on gill histopathology of wild and farmed freshwater and marine fish was carried out by Roubal (1999), Campos et al. (2001), Dezfuli et al. (2003), Avenant-Oldewage et al. (2005), Vinobada (2010), Mohammadi et al. (2012), Kaur and Srivastava (2014), Singh and Kaur (2014), Weli et al. (2017), Mathews et al. (2018), and Suliman et al. (2021). Histopathological changes can be extensively used as biomarkers in the appraisal of health condition of fish (Roberts, 2001). The exploitation of these histopathological biomarkers in environmental monitoring permits the inspection of specific organs (lungs, gills, liver, kidney, etc.) accountable for vital functions such as respiration, excretion, accumulation and biotransformation of xenobiotics in the fish (Gernhofer et al., 2001). The thorough information of histology gives a crystal clear idea to differentiate the pathological changes induced by the parasites in infested and control tissues. The type and intensity of mechanical activity exerted by the ectoparasites are the main causes of the tissue damage and severity of the effect, but the secondary infections through bacteria, virus and fungi in the wounds produced by crustacean parasites are considered to be more dangerous. The freshwater shark, Wallago attu (Bloch & Schneider, 1801) is one among the 10 largest catfish species on earth and are commercially important species harvested across tropical Asia owing to its high nutritional quality and consumer demand (Thella et al., 2018). In the present study, the control gill tissue (least infested) and the infested gill tissue (with severe infection) of the W. attu were considered to note the pathological changes induced by the ectoparasites.
Methods
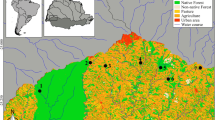
About 95 freshwater shark (W. attu) (11.52 ± 1.95 cm, mean total length ± standard deviation; total length ranged from 7 to 15 cm and 293.15 ± 100.9gms mean total weight ± standard deviation; total weight ranged from 150 to 500 g) were obtained from the two different sampling stations of YSR Kadapa District (Lat. 14°28′N 78°49′E, 137 m altitude), located in Andhra Pradesh State during August 2017 to February 2018. The two sampling sites include Adinimmayapalli Dam across the river Penna in Chennur Village (Lat.14″34′0.12″N, 78″48′0″E longitude), YSR Kadapa District (Site-I, n = 34) and backwaters of Somasila reservoir across the river Penna in Somasila Village (14°29′22″ N 79°18′19″E) Nellore District that reach near Vontimitta Village in YSR Kadapa District (Site-II, n = 61) (Fig. 1). Fish were examined for parasitic diagnosis and gross pathology under the stereo zoom microscope (LM-52-3621 Elegant) and light microscope Lynx trinocular microscope (N-800M) at parasitology laboratory, Department of Zoology, Yogi Vemana University, India. The temporary slides of monogenetic trematodes were prepared using neutral red (C15H17N4Cl) and ammonium picrate-glycerine mixture by the method of Malmberg (1970) and were identified using the taxonomic volumes of Gusev (1976a, b) and Pandey and Agrawal (2008). The copepod and isopod parasites were fixed in 10% formalin solution and processed according to the method of Pillai (1985). The fragments of both infested and uninfested gill arches were carefully removed and processed according to usual histopathological techniques, embedded in paraffin at 58 °C, sectioned at 4–8 µm with Yorco YSI 115 rotary microtome, and stained in hematoxylin-eosin. The slides were investigated and photomicrographs were obtained using drawing tube. Ocular micrometer measurements were depicted in micrometers (µm) using Image J software (https://imagej.nih.gov/ij/download.html) unless indicated.

Geographical location of the two fish sampling sites of YSR Kadapa District, Andhra Pradesh. Site-1: Adinimmayapalli Dam across the Penna River in Chennur Village, YSR Kadapa District. Site-2: Backwaters of Somasila reservoir across the river Penna in Somasila Village, Nellore District, Andhra Pradesh reach near Vontimitta Village, Kadapa
Results
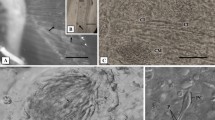
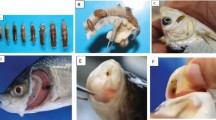
Of the 95 examined fish, 94 fish (P = 98.6%) were found be infected with one or more parasites. The diseased fish showed distinct alterations such as focal and multi-focal paleness and necrosis, hemorrhages, and congestion signs on the gill filaments. In the current work, gills of W. attu were found infested with three monogeneans, i.e. Thaparocleidus indicus (Kulkarni, 1969) Lim, 1996 (n = 688, P = 55.8%, MI = 13.0), Thaparocleidus wallagonius Jain, 1952, (n = 405, P = 56.8%, MI = 7.5) and Mizelleus indicus Jain, 1957 (n = 3, P = 2.1%, MI = 1.5), one parasitic copepod Ergasilus malnadensis Venkateshappa, Seenappa & Manohar, 1998 (n = 2096, P = 98.6%, MI = 22.8) and an isopod, Alitropus typus Milne-Edwards, 1840 (n = 10, P = 9.5%, MI = 1.11) was also found in W. attu (Figs. 2a–d, 3a–e). These parasites attach to all the parts of gill and lead to the damage of the tissue. Normal gill histology revealed the intact nature of both primary and secondary lamellae whereas diseased fish had marked pathological changes in its gills. Histological section of the normal gills of W. attu revealed an intact structure with primary gill lamellae (PGL), secondary gill lamellae (SGL), interlamellar region (ILR) and intact water channels (WC) (Fig. 4a). Histopathological observation revealed the severe damage to primary gill lamellae and secondary gill lamellae, due to attachment and feeding by the parasite. In the present study, all infested W. attu showed color spots. Pathological effects of ectoparasites on the gills include curling of secondary gill lamellae (CSGL), a fusion of secondary lamellae (FSL), damaged central core (DCC) and aneurysm in secondary gill lamellae (ANSGL) (Fig. 4b, c). In a severe infestation, epithelial hyperplasia (EH) of different cell types was observed in both primary and secondary lamellae (Fig. 4d). Also there is proliferated tips of secondary gill lamellae (PTSGL) (Fig. 5a), curling of secondary gill lamellae (CSGL) (Fig. 5b), secretion of mucus (SM), vacuole formation (VF), hyperplasia of epithelial cells (HE) (Fig. 5c), necrosis of gill filaments (NGF) (Fig. 5d), secretions of mucus from gill arch (SM) (Fig. 6a), mucus secretions from epithelial cells (Fig. 6b) and copepod attachment to the epithelial cells (Fig. 6c) resulted thinning of a central core (TCC) (Fig. 6d). The section also exhibited the attachment of the copepod parasite near the gill arches.

a Wallago attu infested with ectoparasites, b Gill rakers of W. attu showing adult and larval stages of Ergasilus malnadensis and monogeneans Thaparocleidus indicus and T. wallagonius, c adult copepod, E. malnadensis attached to the gill rakers of W. attu, d adult copepod, E. malnadensis in embryo cup

a–e Microphotographs of ectoparasites of W. attu, a Thaparocleidus indicus; b Thaparocleidus wallagonius; c Mizelleus indicus, d Ergasilus malnadensis, e Alitropus typus

a Normal gill with inter lamellar spaces (ILS), secondary gill lamella (SGL), water channels (WC). b Infested gill with curling of secondary gill lamella (CSGL), fusion of secondary gill lamella (FSGL), damaged central core (DCC). c Infested gill with fusion of secondary gill lamella (FSGL), damaged central core (DCC) and aneurysm in secondary gill lamella (ANSGL). d Infested gill with hyperplasia (H)

a Infested gill showing secretions of mucus (SM), fusion of secondary gill lamella (FSGL), degeneration of primary gill lamella (DPGL), proliferated tips of secondary gill lamella (PTGSL). b Infested gill with curling of secondary gill lamella (CSGL), fusion of secondary gill lamella (FSGL), damaged central core (DCC), proliferated tips of secondary gill lamella (PTGSL). c Infested gill showing degeneration of central axis (DCA), secretion of mucus (SM), vacuole formation (VF), hyperplasia of epithelial cells (HE). d Infested gill showing necrosis of gill filaments (NGF)

a Infested gill with damaged gill arch (DGA), necrosis of secondary gill filaments (NSGF), secretion of mucus (SM). b Infested gill with aneurysm in secondary gill lamella (ANSGL), curling of secondary gill lamella (CSGL), thinning of central core (TCC). c Gills showed damaged epithelial cells due to parasite attachment (DECPA). d Infested gill with aneurysm in secondary gill lamella (ANSGL), curling of secondary gill lamella (CSGL), thinning of central core (TCC), degeneration of primary gill lamella (DPGL)
Discussion
W. attu infested with ectoparasites (copepods, isopods and monogeneans) demonstrated serious lesions in the gill filaments such as necrosis, hemorrhages, congestion, epithelial and mucus cells proliferation. These alterations may impact the hydromineral balance, gaseous exchange, hormonal production, circulation, nitrogenous waste secretion and making the fish hypoxic or anoxic and finally lead to death and similar pathological results were also observed by Mohammadi et al., (2012) who worked on parasitic infestation of skin and gill on Oscar (Astronotus ocellatus) and discus (Symphysodon discus) and Dias et al. (2021) who worked on Colossoma macropomum (Serrasalmidae), respectively. The severity of respiratory damage is directly related to the number of parasites on the gills. The higher intensity, the more serious the gill damage which can decrease the body weight and condition factor, severe changes in osmoregulation or respiratory dysfunction and ultimately lead to death. Ectoparasites such as monogeneans and copepods append to the primary lamellae with their anchors and legs resulted in cell damage and hemorrhages at the site of adherence. The oozed blood from the hemorrhagic site is fed by the parasite while some parasites secrete exogenous enzymes to digest the gill filaments of the host fish which can be witnessed in the gill histological sections with a high number of cell nuclei within the gut of gill parasites (Vinobada, 2010; Noga, 2010; Seenappa and Venkateshappa, 2000; Pádua et al., 2015). Though the gill epithelial cells are mechanically injured by the hooks of monogenean parasites and copepods (Purivirojkul, 2012), the greatest histopathological damage is induced by scraping and sucking activities of the parasites on host tissues as per the works of Derwa (1995), Endrawes (2001) and Hanna (2001).
The histological sections of the infested gills by the parasitic infection revealed the functional impairment of the gill lamellae, attachment of parasite at the tips of the gill filament, and swelling of the host tissue at the site of attachment. Pathological alterations observed in this study were similar to the works of Campos et al. (2001), Fadaei et al. (2001), Barzegar and Jalali (2004), Shamsi et al. (2009), Kaur and Shrivastav (2014), Pádua et al. (2015) and Arya and Singh (2020) who also recorded hyperplasia of the epithelial cells and succeeding lamellar fusion to be the distinctive feature of the parasitic infection. Hyperplasia of epithelial cells and the fusion of some secondary lamella are the patterns of the defense mechanisms of gills. Severe stress in fish may result in some alterations in blood vessels and damaged cells cause blood congestion or even an aneurysm (Rosety-Rodriguez et al., 2002). The impact of the diseases caused by ectoparasites especially monogeneans and copepods are not only linked to health status of fish but also to economic and societal impacts in case of culture fish, due to diminished respiratory capacity obstructing the growth leading to high mortality rates (Dezfuli et al., 2011; Shinn et al., 2015). The imbalance in the environment can be detected through host-parasite relationships and the consequences caused by the parasites to its hosts. The present findings stress on the fact that diseases are serious threats to cultivable and wild fishes which can be visualized with naked eyes. Hence, regular diagnostic programs together with exceptional management practices need to be executed from time to time to avoid sudden parasitic outbreaks and economic losses in aquaculture.
Conclusions
In present study we have observed with the histopathological alterations induced by the ectoparasites on the gill tissues. The greatest histopathological damage in the gills of the W. attu was induced by scraping and sucking activities of the parasites on host tissues. Histopathological observations disclose the health status of the host fish, i.e., stunted growth, susceptibility to various diseases, resulting in hypoxia or anoxia and thereby leading to the death. Thus, the findings of the present work accomplished that ectoparasitic infestations are serious threats for fish and can cause extensive damage and yield.
Availability of data and materials
The raw data used to support the findings of this study are available from the corresponding author upon reasonable request.
Abbreviations
- PGL:
-
Primary gill lamellae
- SGL:
-
Secondary gill lamellae
- ILR:
-
Interlamellar region
- WC:
-
Water channels
- CSGL:
-
Curling of secondary gill lamellae
- FSL:
-
Fusion of secondary lamellae
- DCC:
-
Damaged central core
- ANSGL:
-
Aneurysm in secondary gill lamellae
- HE:
-
Epithelial hyperplasia
- PTSGL:
-
Proliferated tips of secondary gill lamellae
- CSGL:
-
Curling of secondary gill lamellae
- SM:
-
Secretion of mucus
- VF:
-
Vacuole formation
- HE:
-
Hyperplasia of epithelial cells
- NGF:
-
Necrosis of gill filaments
- DECPA:
-
Degeneration of epithelial cells due to copepod parasite attachment
- TCC:
-
Thinning of central core
References
Arya, P., & Singh, H. S. (2020). Wallago attu (Bl.) and its parasitic monogenea Mizelleus indicus (Jain, 1957), Pandey et al., 2003: A model towards histopathological studies for host parasite interaction. Asian Journal of Biological and Life Sciences, 9, 25. https://doi.org/10.5530/ajbls.2020.9.48
Avenant-Oldewage, A., Tsotetsi, A. M., & Mashego, S. (2005). Aspects of the pathology of Lamproglena clariae (Copepoda: Lernaeidae) on gills of Clarias gariepinus from the Vaal River system South Africa. African Zoology, 40, 169–178.
Barzegar, M., & Jalali, B. (2004). Helminthes, Acanthocephala and crustacean parasites of fishes in Vahdat reservoir. Iranian Journal of Veterinary Science, 2, 229–234.
Boxshall, G. A. (2005). Crustacean parasites (Copepoda). In Rohde, K. (Ed.), Marine Parasitology (CABI) (pp. 123–138). Oxon.
Campos, C. M., Moraes, J. R. E., & Moraes, F. R. (2001). Histopathology of gills of Piaractus mesopotamicus (Holmberg, 1887) and Prochilodus lineatus (Valenciennes, 1836) infested by monogenean and myxosporea, caugth in Aquidauana River State of Mato Grosso do Sul, Brazil. Rev. Bras. Parasitol. Vet. Jaboticabal, 20(1), 67–70.
Derwa, H. I. M. (1995). Some studies on gill affections of some freshwater fishes M.Sc. thesis Faculty of Veterinary Medicine Suez Canal University.
Dezfuli, B. S., Giari, L., Lui, A., Lorenzoni, M., & Noga, E. J. (2011). Mast cell responses to Ergasilus (Copepoda), a gill ectoparasite of sea bream. Fish Shellfish Immun, 30, 1087–1094.
Dezfuli, B., Sayya, F., Luisa, G., Robert, K., Paul, J., & Maurizio, M. (2003). Immunohistochemistry, ultrastructure and pathology of gills of Abramis brama from Lake Mondsee, Austria, infested with Ergasilus sieboldi (Copepoda). Dis. Aqua. Organ, 53, 257–262.
Dias, M., Ferreira, G., & Videira, M. (2021). Histopathological alterations caused by monogenean parasites the gills of tambaqui Colossoma macropomum (Serrasalmidae). Semina Ciências Agrárias. https://doi.org/10.5433/1679-0359.2021v42n3Supl1p2057
Endrawes, M. N. (2001). Observations on some external and internal parasitic diseases in Nile catfishes. A Master thesis submitted to Department of Fish Diseases and Management. Faculty of Veterinary Medicine, Zagazig University.
Fadaei, F., Mokhayer, B., & Ghorbani, H. (2001). Identification of fishes and their parasites in Choghakhor Lagoon. Journal of Faculty of Veterinary Medicine, 56, 109–113.
Gernhofer, M., Pawet, M., Schramm, M., Müller, E., & Triebskorn, R. (2001). Ultrastructural biomarkers as tools to characterize the health status of fish in contaminated streams. Journal of Aquatic Ecosystem Stress and Recovery, 8, 241–260.
Gusev, A. V. (1976). The systematic composition of the Indian fauna, Zoogeography and evolution of freshwater monogeneans. Novaya Seriva, 35, 5–32.
Gusev, A. V. (1976a). Freshwater Indian Monogeneanea. Principles of systematics, analysis of the world faunas and their evolution. Indian Journal of Helminth, 25&26, 1–241.
Hanna, M. I. (2001). Epizootiological studies on parasitic infections in fishes cultured under different fish cultural systems in Egypt. A Master thesis submitted to Department of Fish Diseases and Management. Faculty of Veterinary Medicine, Zagazig University.
Johnson, S. C., Treasurer, J. W., Bravo, S., Nagasawa, K., & Kabata, Z. (2004). A review of the impact of parasitic copepods on marine aquaculture. Zoological Studies, 43(2), 229–243.
Kaur, P., & Shrivastav, R. (2014). Histological effect of monogenean parasites on gills of freshwater carps. European Journal of Biotechnology and Bioscience, 2(2), 50–53.
Lima, F. S., Casali, G. P., & Takemoto, R. M. (2013). Crustacea. In G. C. Pavanelli, R. M. Takemoto, & J. C. Eiras (Eds.), Parasitologia de peixes de água doce do Brasil (1st ed., pp. 371–397). Maringá: Eduem.
Malmberg, G. (1970). The excretory systems and the marginal hooks as a basis for the systematics of Gyrodactylus (Trematoda, Monogenea). Ark. Zool, 23(1), 1–235.
Mathews, P. D., Patta, A. C. M. F., Gama, G. S., & Mertins, O. (2018). Infestation by Ergasilus coatiarus (Copepoda: Ergasilidae) in two Amazonian cichlids with new host record from Peru: An ectoparasites natural control approach. Comp. Rend. Biol, 341(1), 16–19.
Modi, A. K., & Vankara, A. P. (2021). Prevalence and spatial distribution of the ectoparasites on the gills of Mystus vittatus from river Penna flowing through YSR Kadapa District, Andhra Pradesh, India. J. Paras. Dis, 45(1), 43–49.
Mohammadi, F., Mousavi, S. M., & Rezaie, A. (2012). Histopathological study of parasitic infestation of skin and gill on Oscar (Astronotus ocellatus) and discus (Symphysodon discus). Int. J. Bioflux Soc, 5(1), 88–93.
Møller, O. S. (2009). Branchiura (Crustacea)—survey of historical literature and taxonomy. Arthropod Syst. Phylo, 67(1), 41–55.
Noga, E. J. (2010). Fish disease: Diagnosis and treatment (2nd ed.). New York: Wiley.
Ojha, J., & Hughes, G. M. (2001). Effect of branchial parasites on the efficiency of the gills of a freshwater catfish Wallago attu. Journal of Zoology, 255, 125–129.
Pádua, S. B., Jerônimo, G. T., Menezes-Filho, R. N., Taboga, S. R., Martins, M. L., & Belo, M. A. A. (2015). Pathological assessment of farmed yellowtail tetra Astyanax altiparanae infested by Acusicola sp. (Ergasilidae). Aquaculture Reports, 2, 63–66.
Pandey, K. C., & Agrawal, N. (2008). An encyclopedia of Indian Monogeneanea (pp. 1–522). Lucknow.
Pelster, B., & Bagatto, B. (2010). Respiration. Fish Physiol, 29, 289–309.
Pillai, N. K. (1985). Parasitic copepods of marine fishes. In The Fauna of India. Calcutta: Zoological Survey of India.
Purivirojkul, W. (2012). Histological change of aquatic animals by parasitic infection, histopathology—Reviews and Recent Advances, Enrique Poblet Martinez, Intech Open. https://doi.org/10.5772/52769. https://www.intechopen.com/books/histopathology-reviews-and-recent-advances/histological-change-of-aquatic-animals-by-parasitic-infection.
Roberts, R. J. (2001). Fish Pathology, 3rd edn. R. J. Roberts and W. B. Saunders (Eds.).
Rosety- Rodriguez, M., Ordoez, F. J., Rosety, M., Rosety, J. M., Ribelles, A., & Carrasco, C. (2002). Morpho- histochemical changes in the gills of turbot, Scophthalmus maximus L., induced by sodium dodecyl sulphates. Ecotoxicology and Environmental Safety, 51, 223–228.
Roubal, F. R. (1999). Extent of gill pathology in the toad fish Tetractenos hamiltoni by Naobranchia variabilis (Copepoda: Naobranchiidae). Diseases of Aquatic Organisms, 35, 203–211.
Seenappa, D., & Venkateshappa, T. (2000). Multiple parasitic infestation on freshwater shark, Wallago attu (Schneider) and associated gross pathology. Journal of Agricultural Science, 34(2), 153–156.
Shamsi, S., Jalali, B., & Aghazady, M. M. (2009). Infection with Dactylogyrus spp. among introduced cyprinid fishes and their geographical distribution in Iran. Iranian Journal of Veterinary Research, 10(1), 70–74.
Shinn, A., Pratoomyot, J., Bron, J., Paladini, G., Brroker, E., & Brooker, A. (2015). Economic costs of protistan and metazoan parasites to global mariculture. Parasitol, 142(1), 196–270.
Singh, S., & Kaur, P. (2014). Histology of gills of Labeo rohita and Hypophthalmichthys molitrix infested by monogenean and copepod parasites. International Journal of Fisheries and Aquatic Studies, 1(6), 01–06.
Suliman, E. M., Osman, H. A., & Al-Deghayem, W. A. (2021). Histopathological changes induced by ectoparasites on gills and skin of Oreochromis niloticus (Burchell 1822) in fish ponds. Journal of Applied Biology and Biotechnology, 9(1), 68–74.
Thella, R., Dahanukar, N., Eldho, P. S., Ali, A., & Raghavan, R. (2018). Population dynamics of Wallago attu (Bloch and Schneider 1801) (Osteichthyes, Siluridae) in three small rivers of South India. Fisher Scientific, 31, 172–178.
Vinobada, P. (2010). Histopathological changes induced by ergasilid copepod infections on the gills of food fish from Batticaloa Lagoon Sri Lanka. Sri Lanka Journal of Aquatic Sciences, 12, 77–87.
Weli, S. C., Dale, O. B., & Hansen, H. (2017). A case study of Desmozoon lepeophtherii infection in farmed Atlantic salmon associated with gill disease, peritonitis, intestinal infection, stunted growth, and increased mortality. Parasites and Vectors, 10, 370.
Acknowledgements
The corresponding author commemorates this work to late Dr. Asha Kiran Modi, who worked under the financial assistance of UGC Faculty improvement programme (FIP)-Award No.APSC021/001(TF)ZOOLOGY/PH.DXII PLAN/2016-17 dt. July 2016.
Funding
The work was funded to Late, Mrs. Asha Kiran Modi under UGC Faculty improvement programme (FIP)- Award No.APSC021/001(TF)ZOOLOGY/PH.DXII PLAN/2016-17 dt. July 2016.
Author information
Authors and Affiliations
Contributions
APV designed the work and RAP assisted late Dr. Asha Kiran Modi for the fish sample collection to carry out the pathological work. The second and third authors CST and RK helped in framing the manuscript and correction part. All the authors have read and approved the manuscript.
Corresponding author
Ethics declarations
Ethical approval and consent to participate
All procedures contributing to this work comply with the ethical standards of the relevant national guides on the care and use of laboratory animals and have been approved and authorized by IAEC (Institution of Animal Ethics Committee-Regd. No.1460/PO/a/11/CPCSEA, dt. 20.05.2011), Zoology Department in Faculty of Life Sciences, Yogi Vemana University, Andhra Pradesh.
Consent for publication
Not applicable.
Competing interests
The authors declare that no competing interests exists.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Vankara, A.p., Thummala, C., Khateef, R. et al. Histopathological evaluation of Wallago attu (Bloch & Schneider, 1801) infested by ectoparasites. JoBAZ 83, 32 (2022). https://doi.org/10.1186/s41936-022-00294-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41936-022-00294-7