
Abstract
This paper investigates an SIS epidemic model with variable population size including a vaccination program. Dynamics of the endemic equilibrium of the model are obtained, and it will be shown that this equilibrium exists and is locally asymptotically stable when \({\mathcal {R}}_0 > 1\). In this case, the disease uniformly persists, and moreover, using a geometric approach we conclude that the model is globally asymptotically stable under some conditions. Also, a numerical discussion is given to verify the theoretical results.
Similar content being viewed by others


Introduction
The susceptible–infected–susceptible (SIS) model is one of the most well-known type of epidemic models. These models are appropriate for some infections, for instance, common cold and influenza, or bacterial diseases such as meningitis and cholera, or sexually transmitted diseases, that do not cause permanent immunity after recovery. To immunize individuals from infection and control the infectious diseases, vaccination is usually preferred because of its efficiency compared with other drug and non-drug interventions. There are many epidemic models [1, 3, 8, 13, 14, 16, 19] and also some SIS epidemic models in which temporary or permanent vaccination has been included [6, 9,10,11, 17, 22]. These models may be deterministic [10, 11] or stochastic [4, 6, 22], with constant [10, 13] or variable [9] population size, and with general incidence [7, 21] or a particular incidence such as standard, bilinear, and saturated [2, 5, 13]. In this paper, we introduce an SIS epidemic model with vaccination and standard incidence. In the next section, we first describe the model and then some basic properties of it will be given. Sections “Local asymptotical stability” and “Global asymptotical stability” are devoted to investigating the local and global stability of endemic equilibrium of the model, respectively. Finally, we summarize the conclusions in section “Conclusions”, after a numerical consideration in section “Numerical examples”.
Model description
Consider the following system:
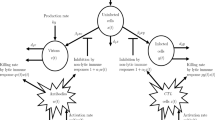
Here, the population has been divided into three subpopulations as susceptible, infected, and vaccinated individuals. The size of population in each class at time t is denoted by S, I, and V, respectively, whereas the number of all individuals in this time is given by \(N=S+I+V\). The recruiting is done by entering a number of \(\varLambda\) individuals into population per unit time that may be either immigrants or newborns. The vaccination program is applied on the new members by a proportion \(\vartheta\). A proportion \(\sigma\) of susceptible individuals are also vaccinated, and the rate of losing immunity of vaccination is \(\xi\). The vaccine is supposed to be completely effective, and no vaccinated individual becomes infected. Susceptible individuals become infected at standard incidence rate \(\frac{\beta S I}{N},\) where \(\beta\) is transmission coefficient, whereas the rate of recovery is \(\eta\). The rate of natural death in population is \(\delta\) , and the mortality due to disease is also included in the model with a rate \(\alpha\). All values in the model are nonnegative except \(\varLambda\) and \(\delta\) that are assumed positive.
From system (1), it can be seen that the total number of members N is not fixed in general and it is expressed by the following equation:
We have \(\frac{{\text{d}}N}{{\text{d}}t} \le \varLambda - \delta N\), and thus, \(\mathop {\limsup N(t)}\limits _{t\rightarrow \infty } \le \frac{\varLambda }{\delta }\). This also states that I, S, and V are bounded. We can see that the feasible region
is a positively invariant set for system (1), and the system is well posed for mathematical and epidemiological considerations in \(\varGamma\).
The equilibria of the model (1) are obtained by solving the following equations:
The system has two solutions: When \(\bar{I}=0\) , the disease-free equilibrium (DFE) \(E^0\)
and when \(\bar{I}>0\), the endemic equilibrium \(E^*=(I^*,S^*,V^*)\), where
Here,
and thus, we have
The quantity \({\mathcal {R}}_0\) is called the basic reproduction number of the model. Furthermore, the total population sizes at two equilibria \(E^0\) and \(E^*\) are obtained, respectively, as follows:
and
In the following sections, we investigate dynamics of the model (1) at the endemic equilibrium \(E^*\) and its asymptotic stability will be obtained. The next section is devoted to consider the local asymptotic stability.
Local asymptotical stability
Letting \(F = \frac{\beta S I}{N}\), the Jacobian matrix of the model (1) at \(E^*\) has the following form:
in which
The characteristic equation of matrix \(J^*\) is
where
We have
Obviously \(a_3 < 0\) and \(a_1 = (\delta +\eta +\alpha )\left( \frac{(\delta +\alpha )(\widetilde{{\mathcal {R}}}_0-{\mathcal {R}}_0)+\delta }{\delta \widetilde{{\mathcal {R}}}_0+\alpha (\widetilde{{\mathcal {R}}}_0-{\mathcal {R}}_0)}\right) >0\). Moreover, notice that the equilibrium \(E^*\) exists if \({\mathcal {R}}_0 > 1\) and this implies
and thus, \(a_2>0\).
Using (8), we obtain
-
(I)
\(-a_1+a_2 = -(\delta +\eta +\alpha )+\delta \beta \left( \frac{{\mathcal {R}}_0-1}{\delta \widetilde{{\mathcal {R}}}_0 +\alpha (\widetilde{{\mathcal {R}}}_0-{\mathcal {R}}_0)}\right);\) thus,
$$\begin{aligned} b_1=\delta \beta \left( \frac{{\mathcal {R}}_0-1}{\delta \widetilde{{\mathcal {R}}}_0 +\alpha (\widetilde{{\mathcal {R}}}_0-{\mathcal {R}}_0)}\right) +(2\delta +\sigma +\xi )>0. \end{aligned}$$ -
(II)
We can see that
$$\begin{aligned}&-a_1(2\delta +\sigma +\xi ) + a_3\sigma +(\delta +\eta +\alpha )(2\delta +\sigma +\xi )\\&\quad = (\delta +\eta +\alpha ) \left( \frac{\delta ({\mathcal {R}}_0-1)}{\delta \widetilde{{\mathcal {R}}}_0+\alpha (\widetilde{{\mathcal {R}}}_0-{\mathcal {R}}_0)}\right) (2\delta +\xi ), \end{aligned}$$and therefore,
$$\begin{aligned} b_2&= (\delta +\eta +\alpha ) \left( \frac{\delta ({\mathcal {R}}_0-1)}{\delta \widetilde{{\mathcal {R}}}_0+\alpha (\widetilde{{\mathcal {R}}}_0-{\mathcal {R}}_0)}\right) (2\delta +\xi )\\&+ a_2(2\delta +\alpha +\xi ) + \delta (\delta +\sigma +\xi )>0. \end{aligned}$$ -
(III)
We have
$$\begin{aligned} b_3= & {} \delta (\delta +\sigma +\xi )(\delta +\eta +\alpha ) \left( \frac{\delta ({\mathcal {R}}_0-1)}{\delta \widetilde{{\mathcal {R}}}_0+\alpha (\widetilde{{\mathcal {R}}}_0-{\mathcal {R}}_0)}\right) \\&+(\beta (\delta +\xi )\\&-(\delta +\sigma +\xi )(\delta +\eta +\alpha ))\delta (\delta +\alpha ) \left( \frac{{\mathcal {R}}_0-1}{\delta \widetilde{{\mathcal {R}}}_0+\alpha (\widetilde{{\mathcal {R}}}_0-{\mathcal {R}}_0)}\right) \\&+\delta (\delta +\sigma +\xi )(\delta +\eta +\alpha ). \end{aligned}$$Besides, \(\widetilde{{\mathcal {R}}}_0>{\mathcal {R}}_0\) and \({\mathcal {R}}_0>1\) implies \(\beta (\delta +\xi )>(\delta +\sigma +\xi )(\delta +\eta +\alpha )\). Therefore,
$$\begin{aligned} b_3>\delta (\delta +\sigma +\xi )(\delta +\eta +\alpha )\left( 1+\frac{\delta ({\mathcal {R}}_0-1)}{\delta \widetilde{{\mathcal {R}}}_0+\alpha (\widetilde{{\mathcal {R}}}_0-{\mathcal {R}}_0)}\right) >0. \end{aligned}$$ -
(IV)
$$\begin{aligned} b_1b_2-b_3&= (\delta +\xi )(b_2-a_2(\delta +\alpha )) \\&\quad+(\delta +\eta +\alpha )(b_2-\delta (\delta +\sigma +\xi ))\\&\quad+ (-a_1+a_2+\delta +\sigma )b_2\\&\quad+a_1\delta (\delta +\sigma +\xi )-a_3\sigma (\delta +\alpha )\\&= (\delta +\xi )(b_2-a_2(\delta +\alpha ))\\&\quad+((\delta +\eta +\alpha )-a_1)(b_2-\delta (\delta +\sigma +\xi ))\\&\quad+(a_2+\delta +\sigma )b_2-a_3\sigma (\delta +\alpha ). \end{aligned}$$
We see that \(a_1 = (\delta +\eta +\alpha ) \left( 1-\frac{\delta ({\mathcal {R}}_0-1)}{\delta \widetilde{{\mathcal {R}}}_0+\alpha (\widetilde{{\mathcal {R}}}_0-{\mathcal {R}}_0)}\right) < (\delta +\eta +\alpha ),\) and as a result,
$$\begin{aligned} b_1b_2-b_3> (\delta +\xi )(b_2-a_2(\delta +\alpha ))+(a_2+\delta +\sigma )b_2-a_3\sigma (\delta +\alpha ) >0. \end{aligned}$$
In preceding relations, we got \(b_1>0\), \(b_2>0\), \(b_3>0\) and \(b_1b_2-b_3>0\); thus, by using Routh–Hurwitz criterion the real part of all eigenvalues of the Jacobian matrix \(J^*\) must be negative. Therefore, the following theorem has been proven:
Theorem 1
For epidemic model (1), the endemic state \(E^*\) exists and is stable if \({\mathcal {R}}_0>1\).
Corollary 1
The endemic state \(E^*\) is stable if parameter values \(\vartheta\) and \(\sigma\) lie under the following line in the \(\vartheta\)-\(\sigma\) plane:
Corollary 1 gives minimum amount of vaccination proportions that is needed to the vaccine be effective. Indeed, those values of \(\vartheta\) and \(\sigma\) that lie above the mentioned line in (9) are sufficient to the disease eradicated from population. When the proportion of vaccination in new members and in susceptibles is equal, i.e., \(\tau =\vartheta =\sigma\), this optimum amount of vaccination will be as
Global asymptotical stability
The global asymptotical stability of the endemic state \(E^*\) is discussed in this section. The analysis of the global stability of epidemic models is generally a difficult task, and the methods introduced for carrying this out are scant. To analyze the global asymptotic stability of the endemic state, we employ a geometric method developed by Li and Muldowney [12] and used by many authors [2, 15, 18,19,20, 23]. The following result has been proved in [12].
Theorem 2
Assume that f is a \(C^1\) function on a simply connected open set \(D\subset \mathbb {R}^n\), the system \(x'=f(x)\) has a unique equilibrium \(\bar{x}\) in D, and there exists a compact absorbing set \(\varTheta \subset D\). Then, \(\bar{x}\) is globally stable in D if
in which \(\varUpsilon =\varSigma _f\varSigma ^{-1}+\varSigma J^{[2]}\varSigma ^{-1}\). Here, \(\varSigma\) is an \(\left( {\begin{array}{c}n\\ 2\end{array}}\right) \times \left( {\begin{array}{c}n\\ 2\end{array}}\right)\) matrix-valued function, \(J^{[2]}\) is the second additive compound matrix of the Jacobian matrix J, \(\varSigma _f\) is obtained by \((\sigma _{ij})_f=\left( \frac{\partial \sigma _{ij}}{\partial x}\right) ^\top . f(x)\), and \(\varPsi (\varUpsilon )\) is the Lozinski\(\breve{\imath }\) measure of \(\varUpsilon\) with respect to a vector norm |.| in \(\mathbb {R}^m\), with \(m=\left( {\begin{array}{c}n\\ 2\end{array}}\right)\), defined by \(\varPsi (\varUpsilon )= \mathop {\lim }\limits _{h \rightarrow 0^+} \frac{|I+h\varUpsilon |-1}{h}\).
The global stability of the endemic state is expressed in the next theorem.
Theorem 3
When \({\mathcal {R}}_0>1\) and \(\delta +\sigma >\xi\), the endemic equilibrium \(E^*\) is globally asymptotic stable if \(\delta +\xi > \max \{\sigma , \alpha \}+ \eta\).
Proof
Considering the feasible region \(\varGamma\), we see that interior and boundary of \(\varGamma\) are \(\mathop {\varGamma }\limits ^{\circ } = \{(I, S, V)\in \varGamma : I>0 \}\) and \(\partial \varGamma = \varGamma \setminus \mathop {\varGamma }\limits ^{\circ } = \{(I, S, V)\in \varGamma : I=0 \}\), respectively. When \({\mathcal {R}}_0>1\), there exists an unique endemic state \(E^*\) , and moreover, the disease-free equilibrium (DFE) \(E^0 \in \partial \varGamma\) is unstable. Now, using Acyclicity Theorem similar to the proof of Theorem 3.2 in [18] we find that when \({\mathcal {R}}_0>1\), system (1) is uniformly persistent. On the other hand, we know that the solutions are bounded because the total population size N is bounded. Considering this with the uniform persistence of the system, it can be concluded that there exists a compact absorbing set \(\varTheta \subset \varGamma\).
The Jacobian matrix of the system at \(E^*\) was given by (6), and thus, its second additive compound matrix is
where
Matrix \(\varSigma\) in Theorem 2 acts as a Lyapunov function. So, for the method to be applicable and efficient, this matrix must be chosen suitable. Now, we choose matrix \(\varSigma\) as \(\varSigma =\frac{S}{I}\mathbb {I}_3\), where \(\mathbb {I}_3\) is the \(3\times 3\) identity matrix. Then, \(\varSigma ^{-1}=\frac{I}{S}\mathbb {I}_3\), \(\varSigma _f=\left( \frac{\frac{{\text{d}}S}{{\text{d}}t}}{I}-\frac{S\left( \frac{{\text{d}}I}{{\text{d}}t}\right) }{I^2}\right) \mathbb {I}_3\) and \(\varSigma _f\varSigma ^{-1}=\left( \frac{\frac{{\text{d}}S}{{\text{d}}t}}{S}-\frac{\frac{{\text{d}}I}{{\text{d}}t}}{I}\right) \mathbb {I}_3\).
Therefore,
The matrix \(\varUpsilon\) can be written in the block form as \(\varUpsilon =\left( \begin{array}{cc} \varUpsilon _{11} &{} \varUpsilon _{12} \\ \varUpsilon _{21} &{} \varUpsilon _{22} \\ \end{array} \right) ,\) in which
Considering the vector norm \(| (\mu _1,\mu _2,\mu _2) | = \max \{ | \mu _1 |,| \mu _2 | + | \mu _3 |\}\) for all \((\mu _1, \mu _2, \mu _3) \in \mathbb {R}^3\) as in [12], we have
where
Thus, we obtain
and
From the second equation in system (1)
Therefore, we have
and
Hence, assuming
yields to
Thus, for each solution \((I(0), S(0), V(0))\in \varTheta\), we have
which implies
Therefore, the conditions of Theorem 2 are hold and this concludes that \(E^*\) is globally asymptotically stable. \(\square\)
Numerical examples
Suppose the parameters in model (1) as \(\varLambda =30\), \(\beta =0.8\), \(\delta =0.2\), \(\alpha =0.1\), \(\eta =0.1\) , and \(\xi =0.2\). For these values, the mentioned line in (9) is shown in Fig. 1. The ordered pair \((\vartheta , \sigma )=(0.2, 0.1)\) lies below the line and by corollary 1 the endemic state \(E^*=(I^*, S^*, V^*)=(34.74, 66.32, 31.58)\) is stable. The point \((\vartheta , \sigma )=(0.4, 0.5)\) lies above the line. In this case, the disease-free equilibrium \(E^0=(I^0, S^0, V^0)=(0, 53.33, 96.67)\) is stable and the infection will be wiped out from population. Besides, for these values for \((\vartheta , \sigma )\) we have \({\mathcal {R}}_0= 1.44>1\) and \({\mathcal {R}}_0=0.71<1\), respectively. Thus, these results can be confirmed also by Theorem 1. Now, if it is supposed that the vaccination proportions in new members and in susceptibles are equal, then the optimal value of such quantity is given by relation (10) as \(\tau ^*=0.286\). For \(\tau =0.05<\tau ^*\) , the infection will remain in the population and the endemic state is stable, while for \(\tau =0.4>\tau ^*\) the number of infected individuals vanishes and the disease-free state is stable (see Fig. 2). Figures 3 and 4 describe the behavior of the model for various initial values and show the solutions are stable. Figure 3 shows the number of infected individuals (sub-figure (a)) and the phase diagram of I(t) and S(t) (sub-figure (b)) when \(\tau =0.4\), and it can be seen that \(E^0\) is stable. Figure 4 shows that I(t) does not vanish when \(\tau =0.05\), and in this case, \(E^*\) is stable .

The line of optimal proportions of vaccine. For those values of \(\vartheta\) and \(\sigma\) that lie below the line, \(E^*\) is stable (red point), and for such values that lie above the line, \(E^0\) is stable (blue point)

Solutions of model (1) for \(\tau =0.05\) (left) and \(\tau =0.4\) (right)

Solutions of I(t) (a) and the phase diagram of I(t) and S(t) (b) for \(\tau =0.4\) with different initial values in each subpopulation. In this case, \({\mathcal {R}}_0=0.80<1\) and disease will be extinct with \(I^0=0, S^0=53.33, V^0=96.67\)

Diagrams of I(t) (a) and the phase diagram of I(t) and S(t) (b) for \(\tau =0.05\) with different initial values. This gives \({\mathcal {R}}_0=1.73>1\) , and disease will persist with \(I^*=34.74, S^*=66.32, V^*=31.58\)
Conclusions
In this paper, a deterministic SIS epidemic model with temporary vaccination was studied. Vaccination includes both susceptible and new members, and disease transmission takes place at standard incidence rate. The number of individuals which are added to the population per unit time is constant and differs from the number of individuals that die and leave the population per unit time. Thus, the total population size is variable. Dynamics of the model at the endemic equilibrium were determined by the basic reproduction number \({\mathcal {R}}_0\); when \({\mathcal {R}}_0 >1\) , the unique endemic equilibrium \(E^*\) is locally asymptotically stable and uniformly persists. In addition, it was shown that in this case \(E^*\) is globally asymptotically stable under some conditions by employing a geometric approach and second additive compound matrix method. A numerical discussion was also performed to support the theoretical results.
References
Allen, L.J.: Introduction to Mathematical Biology. Pearson/Prentice Hall, Upper Saddle River (2007)
Arino, J., McCluskey, C.C., van den Driessche, P.: Global results for an epidemic model with vaccination that exhibits backward bifurcation. SIAM J. Appl. Math. 64(1), 260–276 (2003)
Brauer, F., Castillo-Chavez, C.: Mathematical Models in Population Biology and Epidemiology, vol. 1. Springer, Berlin (2001)
Chalub, F.A., Souza, M.O.: Discrete and continuous SIS epidemic models: a unifying approach. Ecol. Complex. 18, 83–95 (2014)
Farnoosh, R., Parsamanesh, M.: Disease extinction and persistence in a discrete-time SIS epidemic model with vaccination and varying population size. Filomat 31(15), 4735–4747 (2017)
Farnoosh, R., Parsamanesh, M.: Stochastic differential equation systems for an SIS epidemic model with vaccination and immigration. Commun. Stat. Theory Methods 46(17), 8723–8736 (2017)
Hu, Z., Teng, Z., Jiang, H.: Stability analysis in a class of discrete SIRS epidemic models. Nonlinear Anal. Real World Appl. 13(5), 2017–2033 (2012)
Jami, P., Khodabin, M., Hashemizadeh, E.: Numerical solution of stochastic SIR model via split-step forward Milstein method. J. Interpolat. Approx. Sci. Comput. 2016(1), 38–45 (2016)
Jianquan, L., Zhien, M.: Global analysis of SIS epidemic models with variable total population size. Math. Comput. Model. 39(11), 1231–1242 (2004)
Kribs-Zaleta, C.M., Velasco-Hernandez, J.X.: A simple vaccination model with multiple endemic states. Math. Biosci. 164(2), 183–201 (2000)
Li, J., Ma, Z.: Qualitative analyses of SIS epidemic model with vaccination and varying total population size. Math. Comput. Model. 35(11), 1235–1243 (2002)
Li, M.Y., Muldowney, J.S.: A geometric approach to global-stability problems. SIAM J. Math. Anal. 27(4), 1070–1083 (1996)
Liu, X., Takeuchi, Y., Iwami, S.: SVIR epidemic models with vaccination strategies. J. Theoret. Biol. 253(1), 1–11 (2008)
Parsamanesh, M.: Global stability analysis of a VEISV model for network worm attack. Univ. Politehnica Bucharest Sci. Bull. Ser. A Appl. Math. Phys. 79(4), 179–188 (2017)
Parsamanesh, M.: Global dynamics of an SIVS epidemic model with bilinear incidence rate. Ital. J. Pure Appl. Math. 40, 544–557 (2018)
Rahmani, N., Khodabin, M., Hashemizadeh, E.: Numerical solution of stochastic SIR model by Bernstein polynomials. J. Interpol. Approx. Sci. Comput. 2016(1), 19–25 (2016)
Safan, M., Rihan, F.A.: Mathematical analysis of an SIS model with imperfect vaccination and backward bifurcation. Math. Comput. Simul. 96, 195–206 (2014)
Sun, C., Hsieh, Y.H.: Global analysis of an SEIR model with varying population size and vaccination. Appl. Math. Model. 34(10), 2685–2697 (2010)
Yang, W., Sun, C., Arino, J.: Global analysis for a general epidemiological model with vaccination and varying population. J. Math. Anal. Appl. 372(1), 208–223 (2010)
Yang, Y.: Global stability of VEISV propagation modeling for network worm attack. Appl. Math. Model. 39(2), 776–780 (2015)
Zhang, X., Liu, X.: Backward bifurcation and global dynamics of an SIS epidemic model with general incidence rate and treatment. Nonlinear Anal. Real World Appl. 10(2), 565–575 (2009)
Zhao, Y., Jiang, D.: The threshold of a stochastic SIS epidemic model with vaccination. Appl. Math. Comput. 243, 718–727 (2014)
Zhou, X., Cui, J.: Modeling and stability analysis for a cholera model with vaccination. Math. Methods Appl. Sci. 34(14), 1711–1724 (2011)
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Parsamanesh, M., Farnoosh, R. On the global stability of the endemic state in an epidemic model with vaccination. Math Sci 12, 313–320 (2018). https://doi.org/10.1007/s40096-018-0271-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40096-018-0271-3