
Abstract
Fungi respond and adapt to many environmental signals including light. The photobiology of fungi has been extensively investigated, but in recent years the identification of the first fungal photoreceptor, WC-1 in the ascomycete Neurospora crassa, and the discovery that similar photoreceptors are required for photoreception in other ascomycete, basidiomycete and zygomycete fungi has allowed the molecular characterization of light reception and the early steps of signal transduction in a number of model fungi. This contribution is based on presentations made at the Special Interest Group Meeting on “Fungal Photobiology” held during IMC9. The contributions summarize the current status of fungal photobiology in Aspergillus nidulans, Neurospora crassa, Mucor circinelloides, and Coprinopsis cinerea.
Similar content being viewed by others


Introduction
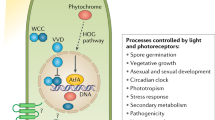
Fungi sense and interact with the environment by changing their pattern of growth and their metabolism, and light is one of the signals that regulates fungal biology (Corrochano & Avalos 2010). A prominent example of the effect of light in fungi is the regulation by light of fungal development and behaviour (Fig. 1) (Corrochano & Galland 2006). Blue light is the type of light most associated with fungal photomorphogenesis. In addition, blue light can activate fungal metabolic pathways or direct the growth of fungal structures (Idnurm et al. 2010).

Sporophores (sporangiophores) of Phycomyces blakesleeanus. Sporangiophores can grow several cm guided by environmental signals towards the open air for spore dispersal. The Phycomyces sporangiophores are guided by light, gravity, wind, touch, and the presence of nearby objects. At the tip of each sporangiophore there is a small ball, the sporangium, filled with vegetative spores. Photograph: Dolores Pérez de Camino, Luis M. Corrochano, and Javier Avalos (University of Seville).
Photoreceptors are molecules that receive photons through specialized light-absorbing chromophores and transduce the photon energy into the cell to promote a response, and several types of photoreceptors have been described in fungi (Corrochano 2007, Herrera-Estrella & Horwitz 2007). The isolation and characterization of fungal photoreceptors was initiated by the isolation of the Neurospora wc-1 and wc-2 genes (Ballario et al. 1996, Linden & Macino 1997) required for all the photoresponses in this fungus. This was followed by the characterization of the wc-1 gene product, WC-1, as a photoreceptor with DNA-binding capacity, that operates with its partner, WC-2 (Froehlich et al. 2002, He et al. 2002). Genes similar to wc-1 and wc-2 have been identified in the genomes of several ascomycete, basidiomycete, and zygomycete fungi, and many of these genes are required for fungal photoresponses, leading to the hypothesis that the WC complex arose early in fungal evolution to regulate fungal photoresponses as a photoreceptor and transcription factor (Idnurm & Heitman 2005). In addition, the completion of several fungal genomes has allowed the identification of additional fungal photoreceptor genes, many of them unexpected like red light absorbing photoreceptors, phytochromes, additional blue-light absorbing photoreceptors, cryptochromes, and rhodopsins (Galagan et al. 2003). In this contribution we provide a summary of the presentations made during the Special Interest Group meeting on “Fungal Photobiology” that took place during the 9th International Mycological Congress (IMC9) in Edinburgh during August 2010.
Contributions
The aim of the meeting was to discuss recent developments in the molecular mechanisms of fungal photoreception and to review the different responses to light observed in different classes of fungi. The meeting was organized by myself with the help of the organizing committee of IMC9 and took place during the morning of 1 August 2010. The five presentations covered various aspects of fungal photobiology in a wide array of fungi, including zygomycetes, basidiomycetes, and ascomycetes. Gerhard Braus (Georg-August-University Goettingen) and Julio Rodríguez-Romero (University of Karlsruhe) described different aspects of light reception in the ascomycete Aspergillus nidulans, and Gencer Sancar (University of Heidelberg) reported on recent developments in the molecular mechanisms that regulate the circadian clock in the ascomycete Neurospora crassa. The responses to light in the zygomycete Mucor circinelloides were discussed by Victoriano Garre (University of Murcia), and Takashi Kamada (Okayama University) reviewed recent results on the regulation by light of basidiome development in the basidiomycete Coprinopsis cinerea.
In Aspergillus nidulans red and blue light regulate the balance between sexual and asexual development, and many proteins, including photoreceptors, participate in this regulation. Interestingly, many of these regulatory proteins interact to form protein complexes, and the formation of these complexes, and their localization within the cell are key features for their roles in the regulation of development (Bayram et al. 2010, Rodriguez-Romero et al. 2010). Braus discussed how a protein complex composed of proteins VelB, VeA, and LaeA is required for the proper regulation by light of sexual development (Bayram et al. 2008). In addition, VelB forms a complex with VosA, which is required for spore viability and resistance to various stresses. The LaeA protein was initially identified as a global regulator of secondary metabolism and plays a key role in the crosstalk between both complexes (Sarikaya Bayram et al. 2010). In A. nidulans red light is perceived by a phytochrome, FphA, and blue light is mostly perceived by the WC-1 homolog LreA. The phytochrome is required for the inhibition by red light of sexual development (Blumenstein et al. 2005) and both, FphA and LreA, interact in a protein complex that includes VeA (Purschwitz et al. 2008). Light activates gene transcription, and the talk by Rodríguez-Romero summarized the results obtained by microarray hybridizations to identify genes differentially regulated by light and how blue light and red light is required for this regulation. Both the red light photoreceptor phytochrome (FphA) and the blue light receptor LreA were required for light-dependent gene activation. The presence of photoreceptor proteins in the promoter of light-regulated genes suggested a direct role for these proteins in photoreception and gene transcription.
The molecular mechanisms of photoreception have been extensively investigated in Neurospora crassa. In Neurospora light promotes the accumulation of orange pigments (carotenoids) in vegetative mycelia, promote asexual (conidiation) and sexual development, and resets the circadian clock (Chen et al. 2010b). All these responses require WC-1 and WC-2, two proteins that form a photoreceptor and transcription factor complex (WCC) that binds the promoters of light-regulated genes to activate transcription. In addition, a small blue-light photoreceptor (VVD) interacts with the WCC to modulate its activity and to allow Neurospora to react to changes in light intensities (Chen et al. 2010a, Hunt et al. 2010, Malzahn et al. 2010). Sancar summarized recent work that helped to identify WCC binding sites along the entire Neurospora genome by ChIP-seq technology after a brief illumination (Smith et al. 2010). The WCC was found in about 400 regions in the genome, and half of them where located in promoters of genes. The WCC was found to directly control the expression of 24 transcription factor genes, including the clock-controlled gene adv-1. The presence of the WCC in the promoters of transcription factor genes confirmed previous observation of a hierarchical regulation in the response to light with the WCC activating a set of transcription factor genes that eventually would activate additional genes responsible for the cellular response (Chen et al. 2009).
The responses to light have been extensively investigated in the zygomycete Phycomyces blakesleeanus, in particular since it was promoted as a model organism for sensory perception by Nobel laureate Max Delbrück in the mid-1950s (Bergman et al. 1969, Cerdá-Olmedo 2001, Cerdá-Olmedo & Lipson 1987). A less known but very promising zygomycete with several light responses is Mucor circinelloides, a fungus that can be easily transformed with exogenous DNA, unlike Phycomyces (Gutiérrez et al. 2011). Light promotes the accumulation of beta-carotene and the phototropism of the fruiting body of Mucor (Silva et al. 2006). Garre reviewed the identification of multiple wc genes in the Mucor genome, a trait observed in other zygomycetes including Phycomyces (Corrochano & Garre 2010). The genome of Mucor contains three wc-1 genes and four wc-2 genes, but only the role of the wc-1 genes has been investigated in detail (Silva et al. 2006). Thus, mcwc-1a and mcwc-1c control phototropism and photocarotenogenesis respectively (Silva et al. 2006). In addition, the product of mcwc-1b is an activator of photocarotenogenesis, but the activation is blocked by ubiquitylation mediated by the repressor CrgA (Silva et al. 2008). In Phycomyces, however, one wc-1 gene, madA, and one wc-2 gene, madB, are required for all the responses of this fungus to light (Idnurm et al. 2006, Sanz et al. 2009). The corresponding proteins, MadA and MadB, interact to form a protein complex, presumably similar to the WCC in Neurospora, that allows phototropism and other responses to light (Sanz et al. 2009). The function of the other wc genes is currently unknown, but they are induced by light and may provide additional light-sensing components to the basic Mad complex for extended sensitivity to light (Sanz et al. 2009).
Mushroom development requires an appropriate light/dark cycle, and this light-dependent developmental regulation has been investigated in detail in the basidiomycete Coprinopsis cinerea (Kamada et al. 2010). Kamada summarized the effect of light on Coprinopsis development and the identification of blind mutants that produce etiolated “dark stipes” under light/dark cycles that would result in full mushroom development in the wild type. The product of dst1 has been identified as a homologue of the Neurospora wc-1 gene (Terashima et al. 2005), while dst2 encodes a protein with a putative flavin adenine dinucleotide (FAD) domain (Kuratani et al. 2010). The presence of this domain suggests that the product of dst2 could be a novel fungal photoreceptor, but further work is still necessary to confirm this hypothesis.
These are very exciting times forfungal photobiology. Proteins similarto WC-1 and WC-2, have been identified in most fungi, and many of them are required for their responses to light. In addition, most fungal genomes contain genes for other photoreceptors, but their roles in the fungal cell are largely unknown. The SIG meeting on “Fungal Photobiology” helped to review the current status of fungal photoreceptors and photoresponses and to propose new avenues of research and collaborations.
References
Ballario P, Vittorioso P, Magrelli A, Talora C, Cabibbo A, Macino G (1996) White collar-1, a central regulator of blue light responses in Neurospora, is a zinc finger protein. EMBO Journal 15: 1650–1657.
Bayram O, Braus GH, Fischer R, Rodriguez-Romero J (2010) Spotlight on Aspergillus nidulans photosensory systems. Fungal Genetics and Biology 47: 900–908.
Bayram O, Krappmann S, Ni M, Bok JW, Helmstaedt K, Valerius O, Braus-Stromeyer S, Kwon NJ, Keller NP, Yu JH, Braus GH (2008) VelB/VeA/LaeA complex coordinates light signal with fungal development and secondary metabolism. Science 320: 1504–1506.
Bergman K, Burke PV, Cerdá-Olmedo E, David CN, Delbrück M, Foster KW, Goodell EW, Heisenberg M, Meissner G, Zalokar M, Dennison DS, Shropshire Wjr (1969) Phycomyces. Bacteriological Reviews 33: 99–157.
Blumenstein A, Vienken K, Tasler R, Purschwitz J, Veith D, Frankenberg-Dinkel N, Fischer R (2005) The Aspergillus nidulans phytochrome FphA represses sexual development in red light. Current Biology 15: 1833–1838.
Cerdá-Olmedo E (2001) Phycomyces and the biology of light and color. FEMS Microbiological Reviews 25: 503–512.
Cerdá-Olmedo E, Lipson ED (eds) (1987) Phycomyces. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press.
Chen CH, Demay BS, Gladfelter AS, Dunlap JC, Loras JJ (2010a) Physical interaction between VIVID and white collar complex regulates photoadaptation in Neurospora. Proceedings of the National Academy of Sciences, USA 107: 16715–16720.
Chen CH, Dunlap JC, Loros JJ (2010b) Neurospora illuminates fungal photoreception. Fungal Genetics and Biology 47: 922–929.
Chen CH, Ringelberg CS, Gross RH, Dunlap JC, Loros JJ (2009) Genome-wide analysis of light-inducible responses reveals hierarchical light signalling in Neurospora. EMBO Journal 28: 1029–1042.
Corrochano LM (2007) Fungal photoreceptors: sensory molecules for fungal development and behaviour. Photochemichal and Photobiological Sciences 6: 725–736.
Corrochano LM, Avalos J (2010) Light sensing. In: Cellular and Molecular Biology of Filamentous Fungi (KA Borkovich, DJ Ebbole, eds): 417–441. Washington DC: American Society for Microbiology Press.
Corrochano LM, Galland P (2006) Photomorphogenesis and gravitropism in fungi. In: The Mycota. Vol. 1. Growth, Differentiation and Sexuality (U Kües, R Fischer, eds): 233–259. Berlin: Springer-Verlag.
Corrochano LM, Garre V (2010) Photobiology in the Zygomycota: multiple photoreceptor genes for complex responses to light. Fungal Genetics and Biology 47: 893–899.
Froehlich AC, Liu Y, Loros JJ, Dunlap JC (2002) White Collar-1, a circadian blue light photoreceptor, binding to the frequency promoter. Science 297: 815–819.
Galagan JE, Calvo SE, Borkovich KA, Selker EU, Read ND, Jaffe D, FitzHugh W, Ma LJ, Smirnov S, Purcell S, Rehman B, Elkins T, Engels R, Wang S, Nielsen CB, Butler J, Endrizzi M, Qui D, Ianakiev P, Bell-Pedersen D, Nelson MA, Werner-Washburne M, Selitrennikoff CP, Kinsey JA, Braun EL, Zelter A, Schulte U, Kothe GO, Jedd G, Mewes W, Staben C, Marcotte E, Greenberg D, Roy A, Foley K, Naylor J, Stange-Thomann N, Barrett R, Gnerre S, Kamal M, Kamvysselis M, Mauceli E, Bielke C, Rudd S, Frishman D, Krystofova S, Rasmussen C, Metzenberg RL, Perkins DD, Kroken S, Cogoni C, Macino G, Catcheside D, Li W, Pratt RJ, Osmani SA, DeSouza CP, Glass L, Orbach MJ, Berglund JA, Voelker R, Yarden O, Plamann M, Seiler S, Dunlap J, Radford A, Aramayo R, Natvig DO, Alex LA, Mannhaupt G, Ebbole DJ, Freitag M, Paulsen I, Sachs MS, Lander ES, Nusbaum C, Birren B (2003) The genome sequence of the filamentous fungus Neurospora crassa. Nature 422: 859–868.
Gutiérrez A, López-García S, Garre V (2011) High reliability transformation of the basal fungus Mucor circinelloides by electroporation. Journal of Microbiological Methods 84: 442–446.
He Q, Cheng P, Yang Y, Wang L, Gardner KH, Liu Y (2002) White collar-1, a DNA binding transcription factor and a light sensor. Science 297: 840–843.
Herrera-Estrella A, Horwitz BA (2007) Looking through the eyes of fungi: molecular genetics of photoreception. Molecular Microbiology 64: 5–15.
Hunt SM, Thompson S, Elvin M, Heintzen C (2010) VIVID interacts with the WHITE COLLAR complex and FREQUENCY-interacting RNA helicase to alter light and clock responses in Neurospora. Proceedings of the National Academy of Sciences, USA 107: 16709–16714.
Idnurm A, Heitman J (2005) Light controls growth and development via a conserved pathway in the fungal kingdom. PLoS Biology 3: e95.
Idnurm A, Rodriguez-Romero J, Corrochano LM, Sanz C, Iturriaga EA, Eslava AP, Heitman J (2006) The Phycomyces madA gene encodes a blue-light photoreceptor for phototropism and other light responses. Proceedings of the National Academy of Sciences, USA 103: 4546–4551.
Idnurm A, Verma S, Corrochano LM (2010) A glimpse into the basis of vision in the kingdom Mycota. Fungal Genetics and Biology 47: 881–892.
Kamada T, Sano H, Nakazawa T, Nakahori K (2010) Regulation of fruiting body photomorphogenesis in Coprinopsis cinerea. Fungal Genetics and Biology 47: 917–921.
Kuratani M, Tanaka K, Terashima K, Muraguchi H, Nakazawa T, Nakahori K, Kamada T (2010) The dst2 gene essential for photomorphogenesis of Coprinopsis cinerea encodes a protein with a putative FAD-binding-4 domain. Fungal Genetics and Biology 47: 152–158.
Linden H, Macino G (1997) White collar 2, a partner in blue-light signal transduction, controlling expression of light-regulated genes in Neurospora crassa. EMBO Journal 16: 98–109.
Malzahn E, Ciprianidis S, Káldi K, Schafmeier T, Brunner M (2010) Photoadaptation in Neurospora by competitive interaction of activating and inhibitory LOV domains. Cell 142: 762–772.
Purschwitz J, Müller S, Kastner C, Schoser M, Haas H, Espeso EA, Atoui A, Calvo AM, Fischer R (2008) Functional and physical interaction of blue- and red-light sensors in Aspergillus nidulans. Current Biology 18: 255–259.
Rodriguez-Romero J, Hedtke M, Kastner C, Muller S, Fischer R (2010) Fungi, hidden in soil or up in the air: light makes a difference. Annual Review of Microbiology 64: 585–610.
Sanz C, Rodriguez-Romero J, Idnurm A, Christie JM, Heitman J, Corrochano LM, Eslava AP (2009) Phycomyces MADB interacts with MADA to form the primary photoreceptor complex for fungal phototropism. Proceedings of the National Academy of Sciences, USA 106: 7095–7100.
Sarikaya Bayram O, Bayram O, Valerius O, Park HS, Irniger S, Gerke J, Ni M, Han KH, Yu JH, Braus GH (2010) LaeA control of velvet family regulatory proteins for light-dependent development and fungal cell-type specificity. PLoS Genetics 6. e1001226.
Silva F, Navarro E, Peñaranda A, Murcia-Flores L, Torres-Martinez S, Garre V (2008) A RING-finger protein regulates carotenogenesis via proteolysis-independent ubiquitylation of a white collar-1-like activator. Molecular Microbiology 70: 1026–1036.
Silva F, Torres-Martinez S, Garre V (2006) Distinct white collar-1 genes control specific light responses in Mucor circinelloides. Molecular Microbiology 61: 1023–1037.
Smith KM, Sancar G, Dekhang R, Sullivan CM, Li S, Tag AG, Sancar C, Bredeweg EL, Priest HD, McCormick RF, Thomas TL, Carrington JC, Stajich JE, Bell-Pedersen D, Brunner M, Freitag M (2010) Transcription factors in light and circadian clock signaling networks revealed by genomewide mapping of direct targets for neurospora white collar complex. Eukaryotic Cell 9: 1549–1556.
Terashima K, Yuki K, Muraguchi H, Akiyama M, Kamada T (2005) The dst1 gene involved in mushroom photomorphogenesis of Coprinus cinereus encodes a putative photoreceptor for blue light. Genetics 171: 101–108.
Acknowledgements
I would like to thank all the attendants of the SIG meeting on fungal photobiology and, in particular, the speakers for their contributions that helped to promote lively discussions on this topic. I would like to thank the organizers of ICM9 for their help and support during the organization of the meeting.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by-nc/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article
Cite this article
Corrochano, L.M. Fungal photobiology: a synopsis. IMA Fungus 2, 25–28 (2011). https://doi.org/10.5598/imafungus.2011.02.01.04
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.5598/imafungus.2011.02.01.04