
Abstract
In selection tasks, target and distractor features can be encoded together with the response into the same short-lived memory trace, or event file (see Hommel, 2004), leading to bindings between stimulus and response features. The repetition of a stored target or distractor feature can lead to the retrieval of the entire episode, including the response—so-called “binding effects.” Binding effects due to distractor repetition are stronger for grouped than for nongrouped target and distractor stimulus configurations. Modulation of either of two mechanisms that lead to the observed binding effects might be responsible here: Grouping may influence either stimulus–response integration or stimulus–response retrieval. In the present study we investigated the influences of grouping on both mechanisms independently. In two experiments, target and distractor letters were grouped (or nongrouped) via color (dis)similarity separately during integration and retrieval. Grouping by color similarity affected integration and retrieval mechanisms independently and in different ways. Color dissimilarity enhanced distractor-based retrieval, whereas color similarity enhanced distractor integration. We concluded that stimulus grouping is relevant for binding effects, but that the mechanisms that contribute to binding effects should be carefully separated.
Similar content being viewed by others

Humans act in an environment that provides them with a continuous stream of complex stimuli requiring an arsenal of different responses. Fortunately, several mechanisms regarding stimulus selection, encoding, and processing enable the efficient control of behavior. One of these mechanisms is the binding between stimuli and responses. According to the theory of event coding (TEC; Hommel, Müsseler, Aschersleben, & Prinz, 2001) perceived stimulus features and executed responses are integrated in loose networks of temporary associations that have been called event files. An event file is created at the time of responding and includes binary bindings between individual stimulus and response features of the event. A reencounter with one or more of the stored features causes the automatic retrieval of the previously constructed event file (Colzato, Raffone, & Hommel, 2006; Hommel, 2004). Repetition of a stimulus feature can thus retrieve the previously integrated response. Therefore stimulus–response (S–R) binding effects (i.e., integration of features and their retrieval upon repetition) have become an important and ubiquitously researched phenomenon in action control and priming (Henson, Eckstein, Waszak, Frings, & Horner, 2014). S–R binding effects are usually investigated using a sequential priming paradigm, which consists of two events: the prime (or trial n–1) and the probe (or trial n). Every time a response is executed to a stimulus in the prime, the stimulus and the executed response are integrated (Henson et al., 2014). Subsequently reencountering the stimulus in the probe triggers retrieval of the prime response and thereby influences probe responding. If the same response is required again probe responding is facilitated, and if a different response is required in the probe, prime response retrieval impairs probe responding. Notably, the measured binding effects in these sequential tasks always reflect two separate processes (Frings et al., 2017; Moeller & Frings, 2014a). In a first step the integration of stimulus and response features in the prime is necessary as a prerequisite for the second step—stimulus–response retrieval—to occur in the probe.
In addition, not only target but also response irrelevant distractor stimuli that accompany the target can be integrated with the response and start response retrieval later on (an effect that has been termed distractor–response binding; Frings, Rothermund, & Wentura, 2007). If the prime response is repeated in the probe, the repetition of the prime distractor facilitates performance because the repeated distractor retrieves the previous integrated response, which matches the demanded response in the probe. If the response is changed from prime to probe, the repetition of the distractor is disadvantageous, because the distractor retrieves the incompatible prime response. The finding of integration of distractor stimuli with a response and the later retrieval thus poses the question of what determines the integration of a distractor in an event file. Hommel (1998, 2005) emphasized that only little or no attention is needed for the integration of (distractor) stimuli and responses. However, by now there is consensus on a few factors that have been shown to reliably modulate binding effects in general (Frings & Rothermund, 2011; Ihrke, Behrendt, Schrobsdorff, Hermann, & Hasselhorn, 2011; Moeller & Frings, 2014a). For instance, grouping or Gestalt principles modulate distractor-based binding effects (e.g., Frings & Moeller, 2012; Frings & Rothermund, 2011; Moeller & Frings, 2017a; Moeller, Rothermund, & Frings, 2012). This modulating influence of stimulus grouping on S–R binding has already been demonstrated for different kinds of grouping, including grouping via common area (Moeller & Frings, 2015; Moeller et al., 2012; van Dam & Hommel, 2010), grouping by the principle of belongingness (Frings & Rothermund, 2011), and grouping via similarity (Baylis & Driver, 1992). Frings and Rothermund (2011) postulated that the principle of belongingness plays a role in the integration of target and distractor stimuli. The authors assumed that an association between two stimuli and a response is only formed if the two elements are perceived as belonging together or as belonging to the same object. In one of their experiments two distractor letters and one target letter were either presented in a horizontal row (grouped condition) or in a vertical column (nongrouped condition). The authors found that binding effects between distractor stimuli and responses were stronger if targets were horizontally flanked by distractors resembling a short word, than if distractors flanked a target on the vertical axis. It thus stands to reason, that grouping of target and distractor stimuli modulates the strength of distractor-based binding effects. According to Frings and Rothermund (2011), effects are stronger if distractors are perceived as belonging to the target stimulus, than if distractors and targets are perceived as parts of separate objects.
Yet it is still unclear which particular process that led to the observed binding effects—integration of features or response retrieval due to feature repetition—was influenced by grouping. One reason for this lack of insight is the design that has typically been used in the past. In virtually all previous studies the stimuli either were grouped both at integration in the prime and at retrieval in the probe or were nongrouped at both times. Although both processes can be assumed to have led to the observed binding effects (as in the study by Frings & Rothermund, 2011), a lack of binding effects can in principle be due to either only the retrieval or only the integration process being influenced by the grouping modulation.
Taking a closer look at integration and retrieval processes, it becomes clear that grouping might influence these separate processes in different ways. To be integrated into a given event file, a stimulus must be perceived as part of this event. This becomes even more apparent if we consider that event files have been interpreted as recent action plans (e.g., Moeller & Frings, 2014a, 2017a; Stoet & Hommel, 1999). Grouping of a to-be-integrated stimulus with other relevant stimuli can signal belongingness to the same event and should hence increase the probability of its integration. Therefore, one would expect integration to be more likely or stronger if the distractor stimulus is grouped with the target stimulus. In contrast, a stimulus that triggers retrieval of a previously integrated response does not necessarily need to be grouped with other relevant stimuli (at least if one assumes automatic episodic retrieval; Logan, 1980). As long as a stimulus is processed to a sufficient degree (e.g., by being salient, presented before the onset of the target, etc.), a repeated stimulus would start the retrieval process (e.g., Dutzi & Hommel, 2009; Moeller, Pfister, Kunde, & Frings, 2016), which then competes with any additional response selection process in a metaphoric horserace (Frings & Moeller, 2012; Neill, 1997; Tipper, 2001). A nongrouped distractor should be more salient (i.e., stands out more) than one that is perceived as belonging to the target stimulus, and accordingly might have an advantage in the metaphoric horserace with a grouped distractor.
Taken together, two separate processes—S–R integration and S–R retrieval—are necessary for S–R binding effects to manifest. Even though these processes are likely to be influenced differently by factors that have been shown to modulate binding effects, past studies have ignored the different influences that factors like grouping might have on integration and retrieval. The present study was designed to disentangle the possibly separate influences that stimulus grouping has on integration and retrieval mechanisms in S–R binding.
The present study
Different predictions about the impacts of grouping on the integration and retrieval processes can be made (see Fig. 1). With regard to the integration process, we would expect benefits due to grouping. As has been suggested previously (Frings & Rothermund, 2011; Hommel, 1998), presumably not every feature in every situation is integrated into an S–R episode—uncontrolled binding of every feature in every situation would result in many incompatible S–R episodes (Wiswede, Rothermund, & Frings, 2013). Several mechanisms besides grouping have been discussed as hedges against “unwanted” bindings—for example, feedback (Waszak & Pholulamdeth, 2009), errors due to binding (Wiswede et al., 2013), or figure–ground asymmetries (Frings & Rothermund, 2017). In this regard, it seems likely that grouping is one mechanism controlling feature integration: Only features belonging to the configuration to which a response has been given are integrated (Frings & Rothermund, 2011). Thus, grouped prime stimulus configurations that suggest belongingness of the distractor and the target to the same event should elicit stronger binding effects. Nevertheless, it has been shown that the process that integrates a stimulus with an executed response is a highly automatic process and that not much is needed to integrate a stimulus with a response (Hommel, 1998). For example, differences in neither free cognitive capacity (Hommel, 2005) nor the task relevance of additional stimuli (Giesen, Frings, & Rothermund, 2012) have affected distractor-based binding effects. In line with these findings, it could also be assumed that the integration process is not further influenced by stimulus grouping.

Predicted impacts of stimulus configurations (grouped vs. nongrouped) on the size of binding effects, due to stimulus–response integration (at the prime) and due to stimulus–response retrieval (at probe)
Concerning retrieval due to irrelevant features, one might come to a different conclusion about the kind of influence that grouping can have on the retrieval process. In stimulus configurations with separable target and distractor stimuli, it is often assumed that the targets and distractors compete (in a metaphoric horserace) for access to the response selection stage (Frings & Moeller, 2012; Neill, 1997; Tipper, 2001). A simple rule is that the “stronger” the distractor (i.e., in terms of being salient, presented before the onset of the target, etc.), the more the horserace tilts in its favor. For instance, Ihrke et al. (2011; see also Moeller & Frings, 2014a) observed stronger distractor-based retrieval if the distractor was attended at retrieval, as compared to a condition in which it was unattended at retrieval. Notably, if a stimulus is nongrouped at retrieval, it is more salient—that is, it sticks out from surrounding stimuli that it might be grouped with. Hence, irrelevant features or objects that are grouped with the target can be less salient than ungrouped features or objects. For instance, in the case of flanker-like stimulus configurations (with distractors flanking the target stimulus; e.g., Eriksen, 1995), dissimilar flanker stimuli (e.g., AbA) would be less grouped with the target than would similar ones (e.g., aba). Therefore, dissimilar (i.e., ungrouped) distractors might be more salient and hence lead to stronger retrieval. Thus, nongrouped probe stimulus configurations that would possibly enhance the processing of the distractor should elicit stronger binding effects.
Overview
In two experiments we manipulated the grouping of distractor and target stimuli via color similarity during the integration process and during the retrieval process independently. A prime–probe design, requiring responses to both the prime and the probe, was chosen for both experiments. This design allows for the orthogonal variation of response relation and distractor relation between prime and probe (see Frings & Rothermund, 2011). An interaction of distractor relation and response relation indicates distractor-based binding effects: Distractor repetitions facilitate performance more if the response is repeated than if the response has to be changed between prime and probe (Frings et al., 2007).
Orthogonally to response and distractor relation, the color (dis)similarity between the distractor and target was varied in both the prime and the probe. This design offers the possibility to differentiate between the separate influences of color similarity (i.e., grouping) on integration and retrieval processes in stimulus–response binding effects.
In Experiment 1 we analyzed the influence of grouping on retrieval. We assumed that probe retrieval would be stronger in a nongrouped stimulus condition; if the distractor were not grouped with the target in the probe, it would stand out from the background (i.e., be salient), capture more attention, and hence have more potential to start the retrieval process and win or modulate the metaphoric horserace. Thus, we expected stronger distractor-based retrieval effects with color-dissimilar probes, independent of the color (dis)similarity in the prime. Statistically, this would be indicated by a three-way interaction between color (dis)similarity in the probe, response relation, and distractor relation. Note that we assumed that integration and retrieval would be independently influenced by grouping—statistically, we should therefore not observe a four-way interaction of color (dis)similarity in the prime, color (dis)similarity in the probe, response relation, and distractor relation.
In Experiment 2 we analyzed the influence of grouping on prime integration. Stimulus–response integration should be more likely, or stronger, if the to-be-integrated distractor stimuli are similar to the target and hence perceived as belonging to the same object as the target (as was suggested by Frings & Rothermund, 2011), as compared to a condition in which the stimuli are dissimilar. Thus, the distractor-based binding effect should be stronger for prime displays with color-similar stimuli, independent of the color (dis)similarity in the probe. Statistically, this pattern would be indicated by a three-way interaction between color (dis)similarity in the prime, response relation, and distractor relation. Again, we assumed that integration and retrieval would be independently influenced by grouping—statistically, we should therefore not observe a four-way interaction of color (dis)similarity in the prime, color (dis)similarity in the probe, response relation, and distractor relation.
To anticipate the results, with two slightly different stimulus configurations we found evidence for separate influences of grouping on probe retrieval (Exp. 1) and prime integration (Exp. 2). In particular, S–R binding effects were larger with nongrouped distractors in the probe in Experiment 1, whereas they were larger with grouped distractors in the prime in Experiment 2.
Experiment 1
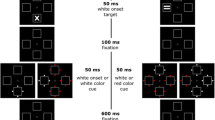
The design of the first experiment closely corresponded to that used by Frings and Rothermund (2011). Stimuli were presented in a flanker configuration, and participants’ task was always to identify the shape of the central target letter and to ignore the flanking distractor letters. To manipulate grouping, we systematically varied the color (dis)similarity between the target and distractor stimuli. Importantly, color (dis)similarity was varied independently in the prime (at distractor–response integration) and the probe (at response retrieval due to distractor repetition).
Of main interest was the influence of color (dis)similarity between the distractor and target on the two processes that generate the distractor–response binding effect. If the distractor–response binding effect is modulated by prime color (dis)similarity, this would indicate that the magnitude of the binding effect can be influenced via the integration process. We would then expect larger binding effects for similar (i.e., grouped) than for dissimilar (nongrouped) prime stimuli. If the distractor–response binding effect is modulated by probe color (dis)similarity, this would evidence that differences in the retrieval process can modulate binding effects. We would then expect larger binding effects for dissimilar (i.e., salient) than for similar (nonsalient) probe stimuli.
Method
Participants
Thirty-one students (19 female, 12 male) from the University of Trier took part in the experiment. Their median age was 24 years, with a range from 20 to 29 years. None of the participants reported deficiencies in color vision. All participants took part in exchange for partial course credit.
Design
The design of the experiment included four within-subjects factors: response relation (response repetition vs. response change), distractor relation (distractor repetition vs. distractor change), color similarity between distractor and target in the prime (similar prime distractor vs. dissimilar prime distractor), and color similarity between distractor and target in the probe (similar probe distractor vs. dissimilar probe distractor).
Materials and apparatus
The experiment was conducted using the E-Prime software (version 2.0). Instructions and stimuli were shown on a standard color monitor (1,680 × 1,050 pixels). Reponses were collected via a serial response box (Psychology Software Tools Inc., Sharpsburg, USA). The stimuli were the letters S, L, X, and C presented on a black background. Each letter had a horizontal visual angle of 1.1° and a vertical visual angle of 1.0° at an approximate viewing distance of 50 cm. The targets were presented in the colors red, green, blue, or yellow. Distractors were presented in yellowish red (according to the RGB color system: red = 255, green = 120, and blue = 29) or bluish green (according to the RGB color system: red = 0, green = 196, and blue = 182). Hence, the yellowish-red distractor color was mainly a compound of the colors yellow and red, whereas bluish green was mainly a compound of the colors blue and green (see Ansorge & Heumann, 2004).Consequently, the distractor color yellowish red was similar to the target colors yellow and red and dissimilar to the target colors blue and green, whereas bluish green was similar to the target colors blue and green and dissimilar to the target colors red and yellow. It can be assumed that a distractor that is more similar to a target is perceived as belonging to the target or, respectively, as belonging to the same object. This conforms to studies by Ansorge and Heumann (2004) and Tipper, Weaver, and Houghton (1994). The target stimulus was always flanked by two identical distractor stimuli. The two distractor stimuli on one display always had the same color.
Procedure
Each participant was tested individually in a soundproof chamber. Instructions were given on the computer screen. The participants were instructed to place their index fingers of both hands on the leftmost and rightmost keys of the response box, which were marked with an X or a C. Their task was always to classify the identity of the target letter by pressing the corresponding key with the respective index finger. It was emphasized that participants should respond as quickly and accurately as possible. A single trial consisted of the following events: A plus sign was presented for 500 ms as a fixation mark at the center of the screen. After that, the prime target flanked by the distractor letters appeared at the center of the screen. The stimuli were presented until a response was given by pressing one of the keys. In the case of an error, a feedback display appeared for 1,000 ms, reminding the participant to respond as quickly as possible without making errors. A fixation mark was then presented for 500 ms, followed by the probe target and distractor letters, which were again presented at the center of the screen until the participant responded by pressing one of the two keys. The distractor and target identities and the target color could change between the prime and probe displays. After the probe display, a black screen appeared for 500 ms, and then the next trial started.
In response repetition trials, the same response was required in the prime and in the probe. In response change trials, different responses had to be given to the target in the prime and to the target in the probe. Orthogonally to the response relation, the distractor relation was varied. In distractor repetition trials, the prime distractor identity was repeated in the probe display, whereas in distractor change trials, the distractor identity changed from prime to probe.
This manipulation of response relation and distractor relation leads to four different conditions: in the response repetition and distractor repetition condition (RRDR), the prime response and the prime distractor identity were repeated in the probe; in the response repetition and distractor change condition (RRDC), the prime response was repeated in the probe, while the distractor identity was different in the prime and the probe; in the response change and distractor repetition condition (RCDR), the prime target response differed from that in the probe, while the prime distractor identity was repeated in the probe; and in the response change and distractor change condition (RCDC), both the response and distractor identities changed between prime and probe.
Orthogonally to response and distractor relation, the color (dis)similarity between distractor and target was varied within the prime and within the probe. From now on, the factor color (dis)similarity between distractor and target in the prime (similar prime distractor vs. dissimilar prime distractor) will be labeled “prime color similarity.” The factor color (dis)similarity between distractor and target in the probe (similar probe distractor vs. dissimilar probe distractor) will be labeled “probe color similarity.”
The experiment was subdivided in two blocks depending on the distractor’s color. In one block the distractor letters were always presented in bluish green, and in the other block they were always presented in yellowish red. Depending on the current color similarity condition, the target letters could be blue, green, red, or yellow. For example, in a trial with similar target and distractor color in the prime and dissimilar target and distractor color in the probe in the yellowish-red block, the prime target was presented in red or yellow and the probe target in blue or green, and vice versa in the bluish-green block. Half of the participants saw the bluish-green block first, the other half saw the yellowish-red block first.
At the beginning of each block and the corresponding training, the participants were informed about the distractor color in the following block. Additionally, they saw all colors that would be presented and were informed, respectively, that the distractor color bluish green was a combination of the colors blue and green or that the color yellowish red was a combination of the colors yellow and red. The participants were also informed that none of the colors would be relevant for their response.
All participants worked through two experimental blocks (bluish-green block and yellowish-red block). Each trial condition (response repetition/distractor repetition, response repetition/distractor change, response change/distractor repetition, and response change/distractor change) was realized 64 times in each block. That is, each experimental block consisted of 256 trials. Every 90 trials, participants had the possibility to take a break. Prior to each experimental block, all participants had to work through a practice block consisting of 20 trials. The sole difference in the experimental block was that participants received feedback for both incorrect and correct responses. Participants were randomly assigned a block order, and the sequence of trials was randomized for each participant.
Results
Only the probe reaction times (RTs) in trials with correct answers to both the prime and the probe target were considered. Moreover, for each person, only RTs slower than 200 ms and faster than 1.5 interquartile ranges from the third quartile of each person’s RT distribution were analyzed (Tukey, 1977). According to these constraints, 9.00% of the trials were discarded: 2.76% of the trials were excluded because of erroneous responses in the prime, 2.21% of the trials were excluded because of erroneous responses in the probe, and 4.03% due to the RT outlier criteria. The mean RTs and error rates for the probe are depicted in Table 1. The distractor–response binding effect was computed as the distractor repetition benefit in response repetition trials minus the distractor repetition benefit in response change trials.
A 2 Response Relation (response repetition vs. response change) × 2 Distractor Relation (distractor repetition vs. distractor change) × 2 Prime Color Similarity (similar prime distractors vs. dissimilar prime distractors) × 2 Probe Color Similarity (similar probe distractors vs. dissimilar probe distractors) multivariate analysis of variance (MANOVA) on probe RTs yielded a significant main effect of response relation, F(1, 30) = 33.36, p < .001, ηp2 = .53. Participants responded more quickly if the response had to be repeated (M = 417 ms, SD = 42 ms) than if the response had to be changed (M = 444 ms, SD = 44 ms) from prime to probe. The main effect of distractor relation was significant, too, F(1, 30) = 4.86, p = .035, ηp2 = .14. Participants responded more quickly if the distractor was repeated (M = 429 ms, SD = 43 ms) than if the distractor changed (M = 431 ms, SD = 44 ms) from prime to probe. Additionally, the main effect of probe color similarity was significant, F(1, 30) = 16.38, p < .001, ηp2 = .35; that is, participants responded faster if the probe color was dissimilar. The main effect of prime color similarity did not reach significance, F(1, 30) = 1.52, p = .227, ηp2 = .05.
The analyses further revealed an interaction between response relation and distractor relation, F(1, 30) = 54.28, p < .001, ηp2 = .64, indicating a general distractor–response binding effect: Repeating a distractor leads to facilitation if the response is repeated too, but leads instead to impairment if the response changes. Most importantly, the three-way interaction between probe color similarity, response relation, and distractor relation was significant, F(1, 30) = 5.47, p = .026, ηp2 = .15, whereas the interaction of prime color similarity, response relation, and distractor relation was not, F(1, 30) = 0.59, p = .449, ηp2 = .02. That is, distractor–response binding was modulated by the color similarity in the probe, but not by color similarity in the prime. Further analyses indicated a larger binding effect if the probe target and distractor colors were dissimilar, F(1, 30) = 63.63, p < .001, ηp2 = .68, than if the probe target and distractor colors were similar, F(1, 30) = 9.06, p = .005, ηp2 = .23 (see Fig. 2, upper panels). Figure 3 visualizes the result pattern for all four color-similarity conditions. Numerically, distractor repetition always leads to an advantage (i.e., shorter RTs than for distractor change) in response repetition trials, but to a disadvantage in response change trials. Yet this distractor–response binding pattern is more pronounced in dissimilar-probe conditions than in similar-probe conditions. The four-way interaction between prime color similarity, probe color similarity, response relation, and distractor relation was not significant, F(1, 30) = 1.01, p = .323, ηp2 = .03. Hence, the modulating effect of grouping (grouped vs. nongrouped) during probe retrieval was independent of the modulation of grouping (grouped vs. nongrouped) during prime integration (see also Fig. 3).Footnote 1

Distractor–response binding effects, in milliseconds, in Experiment 1 in the upper panels and in Experiment 2 in the lower panels, as a function of color similarity during prime integration (collapsed over color similarity during the probe; left-hand side) and color similarity during probe retrieval (collapsed over color similarity during the prime; right-hand side). Distractor–response binding effects were calculated as the difference between the distractor repetition effects in response repetition and response change trials. Error bars depict standard errors of the means

Reaction times (RTs) in milliseconds in Experiment 1, as a function of response relation, distractor relation, prime color similarity, and probe color similarity. Error bars depict standard deviations of the means
Additionally, the interaction of prime color similarity, probe color similarity, and response relation was also significant, F(1, 30) = 158.72, p < .001, ηp2 = .84. The RT benefit due to response repetition (as compared to response change) was modulated by both prime color similarity and probe color similarity. The benefit was larger if color similarity was identical in the prime and probe (i.e., the condition with similar distractor in the prime and similar distractor in the probe, and the condition with dissimilar distractor in the prime and dissimilar distractor in the probe) than in those conditions in which the similarity between distractor and target changed between prime and probe. For the sake of completeness, the interaction of prime color similarity and probe color similarity, F(1, 30) = 10.46, p = .003, ηp2 = .26, was also significant. None of the other effects reached significance, all Fs(1, 30) < 1.52, ps > .227.
The same analysis on error rates yielded a significant interaction of response relation and distractor relation (distractor–response binding effect), F(1, 30) = 24.36, p = .001, ηp2 = .45. In addition, the interaction between prime color similarity, probe color similarity, and response relation reached significance, F(1, 30) = 43.65, p = .001, ηp2 = .59.
Discussion
In Experiment 1 we investigated the influence of grouping via color similarity on the distractor–response binding effect. Due to the experimental design, this influence could be examined independently for the integration process (during the prime) and for the retrieval process (during the probe). Participants responded to a target letter and ignored distractor letters that flanked the target on both sides.
We found an interaction of response relation and distractor relation, which indicated the standard distractor–response binding effect (e.g., Frings et al., 2007): If the response was repeated from prime to probe, the repetition of the prime distractor facilitated performance, whereas its repetition impaired performance if the response had to be changed from prime to probe.
Most importantly, we found a modulating influence of probe color similarity on the distractor–response binding effect, indicated by the three-way interaction of response relation, distractor relation, and probe color similarity. The distractor–response binding effect was larger when probe target and probe distractor were dissimilar than when they were similar (see Fig. 3). That is, grouping in the probe reduced the S–R binding effect.
In this setup a dissimilar distractor was more salient and was easier to separate from the target. It thus produced an advantage to start a retrieval process (as compared to a similar distractor), leading to a benefit in the metaphoric horserace of target responding and distractor–response retrieval. This interpretation is in line with the suggestion that stimulus saliency boosts binding effects of the salient stimulus (Hommel et al., 2001). Given the present results, it is possible to more specifically pinpoint one mechanism (i.e., response retrieval) that is modulated by grouping (i.e., stimulus saliency of the distractor in the nongrouped condition) and can thus account for differences in the binding effect. This finding corresponds to those in previous studies (e.g., Ihrke et al., 2011, and Moeller & Frings., 2014a), showing modulations of binding effects via differences in the retrieval process. Hence, there is now ample evidence that S–R binding effects can be influenced via the feasibility of triggering the retrieval process.
Finally, this effect of probe retrieval is independent of prime integration processes, as indicated by the nonsignificant four-way interaction, and hence is independent of whether the dissimilar distractor in the probe is following a similar or a dissimilar prime display. Particularly, since response repetition effects were influenced by prime and probe similarity (i.e., the significant three-way Response Repetition × Prime Color Similarity × Probe Color Similarity interaction), it is important to note that binding effects were independent of prime–probe context effects here. We will turn to this issue in detail in the General Discussion.
In contrast, the three-way interaction between response relation, distractor relation, and prime color similarity was not significant. Hence, the color similarity between target and distractors only had an influence on the retrieval process, whereas the integration process was not further affected by grouping via color similarity. This might be explained by the fact that the flanker configuration by itself already represents a form of grouping that influences integration (see Frings & Rothermund, 2011). Hence, it is possible that grouping via the flanker configuration (i.e., both temporal and spatial grouping) already ensured distractor–response integration, independent of additional grouping by color similarity between the target and distractor. Due to the temporal and spatial grouping in the flanker configuration, the dissimilarity between distractor and target was simply too weak to cancel the integration due to temporal and spatial proximity (probably the two strongest influences for feature integration; Holmes & Spence, 2005; Spence, 2013; Treisman & Gelade, 1980). To increase the likelihood that grouping by color similarity would influence the binding process, we slightly changed the stimulus configuration in the second experiment. These changes prevented integration by other variables such as spatial grouping or simultaneous onset, and also ensured similar chances of retrieval in all conditions.
Experiment 2
Experiment 2 was designed to examine the influence of target–distractor grouping via color similarity on the distractor–response binding effect, while grouping by other factors was minimized. Distractor and target letters were no longer presented in a flanker configuration, but one after the other in a temporal sequence. Hence, binding of the distractor stimulus and the response would have to be enabled by a grouping effect other than temporal co-occurrence. In addition, due to this stimulus configuration, the influence of grouping on stimulus–response integration could be investigated while influences of stimulus similarity on retrieval were minimized. Participants’ task was always to identify the shape of the target letter (the second letter) and to ignore the distracting letter that had been presented prior to the target letter at the same position. As in Experiment 1, the color similarity between distractor and target was varied independently in the prime (the integration process) and in the probe (the retrieval process). However, presenting the potentially retrieval-triggering stimulus before target onset during the probe ensured that at least some retrieval would take place.
Method
Participants
Thirty students (20 female, 10 male) from the University of Trier took part in Experiment 2. Their median age was 21 years, with a range from 20 to 31 years. None of the participants reported deficiencies in color vision, and all took part in exchange for partial course credit. The average RTs of three participants constituted outliers according to Tukey (1977) when compared to the sample distribution of the average RTs of all participants. The data of these three participants were therefore excluded from the analyses.
Design
The design of Experiment 2 included four within-subjects factors: response relation (response repetition vs. response change), distractor relation (distractor repetition vs. distractor change), prime color similarity (similar prime distractor vs. dissimilar prime distractor), and probe color similarity (similar probe distractor vs. dissimilar probe distractor).
Materials and procedure
The materials and procedure were the same as those in Experiment 1, with the following exceptions. The stimuli were the letters S, L, X, and O, and a trial in Experiment 2 consisted of the following events: A plus sign was presented as a fixation mark for 500 ms at the center of the screen. After that, the fixation mark disappeared and the prime distractor letter (L or S) appeared for 300 ms on the left or the right side of the fixation mark’s position. The prime distractor was followed by the prime target letter (X or O), which was presented until a response was given by pressing one of the particular keys. The target was always presented at the same location as the preceding distractor and was followed by a fixation mark at the center of the screen for 500 ms. Then the probe distractor letter (L or S) appeared for 300 ms, followed by the probe target letter (X or O) at the same position as the probe distractor letter until a response had been made by pressing a key. Finally, the screen went black for 500 ms. The position of the distractor and the target was orthogonally varied with both response and distractor relation and could differ between prime and probe. The prime and probe positions were randomly assigned in each trial.
Results
As in Experiment 1, only probe RTs for trials with correct answers to both the prime and the probe target were considered. Moreover, only RTs slower than 200 ms and faster than 1.5 interquartile ranges over the third quartile of each person’s RT distribution were analyzed (Tukey, 1977). According to these constraints, 10.68% of the trials were discarded: 5.27% of the trials were excluded because of erroneous responses in the prime, 2.82% of the trials were excluded because of erroneous responses in the probe, and 2.59% due to the RT outlier criteria. The mean RTs and error rates on the probe are depicted in Table 2. Again, the distractor–response binding effect is computed as the distractor repetition benefit in response repetition minus the distractor repetition benefit in response change trials.
In a 2 Response Relation (response repetition vs. response change) × 2 Distractor Relation (distractor repetition vs. distractor change) × 2 Prime Color Similarity (similar prime distractor vs. dissimilar prime distractor) × 2 Probe Color Similarity (similar probe distractor vs. dissimilar probe distractor) × 2 Position Relation Between Prime and Probe (position repetition vs. position change) MANOVA on the probe RTs, with Pillai’s trace as the criterion, the main effect of position relation, F(1, 26) = 3.56, p = .069, ηp2 = .11, was not significant. Additionally, the position relation factor did not interact with any of the following analyses, and hence was discarded.
A 2 Response Relation (response repetition vs. response change) × 2 Distractor Relation (distractor repetition vs. distractor change) × 2 Prime Color Similarity (similar prime distractor vs. dissimilar prime distractor) × 2 Probe Color Similarity (similar probe distractor vs. dissimilar probe distractor) MANOVA yielded a significant main effect of response relation, F(1, 26) = 17.20, p < .001, ηp2 = .40. Participants responded more quickly if the response had to be repeated (M = 388 ms, SD = 35 ms) than if the response had to be changed (M = 407 ms, SD = 36 ms) from prime to probe. Neither the main effect of distractor relation, F(1, 26) = 0.47, p = .500, ηp2 = .02, nor the main effect of prime color similarity, F(1, 26) = 0.11, p = .749, ηp2 = .01), nor the main effect of probe color similarity, F(1, 26) = 2.35, p = .137, ηp2 = .08, reached significance. The interaction between response relation and distractor relation was significant, F(1, 26) = 14.14, p = .001, ηp2 = .35, indicating a general distractor–response binding effect.
Most importantly, in contrast to Experiment 1, the three-way interaction between prime color similarity, response relation, and distractor relation was significant, F(1, 26) = 4.47, p = .044, ηp2 = .15, but the interaction of probe color similarity, response relation, and distractor relation was not, F(1, 26) = 0.23, p = .635, ηp2 = .01. That is, the distractor–response binding effect was modulated by color similarity in the prime, but not by color similarity in the probe (see Fig. 2, lower panels). Follow-up analyses indicated a significant binding effect only if the prime target and distractor colors were similar, F(1, 26) = 20.48, p < .001, ηp2 = .44, but no binding effect if the prime target and distractor colors were dissimilar, F(1, 26) = 3.74, p = .064, ηp2 = .13. As can be seen in Fig. 4, the result pattern indicating distractor–response binding effects is found in both conditions with similar primes (i.e., with a similar probe as well as with a dissimilar probe): RTs in the response repetition conditions were faster in the case of distractor repetition than in the case of distractor change, whereas RTs in the response change conditions were slower in the case of distractor repetition than in the case of distractor change. The same pattern was less pronounced for both conditions with dissimilar primes. The four-way interaction between prime color similarity, probe color similarity, response relation, and distractor relation was not significant, F(1, 26) = 0.18, p = .676, ηp2 = .01. Hence, the modulating effect of (non)grouping during prime integration was independent of the modulation of (non)grouping during probe retrieval.Footnote 2

RTs in milliseconds in Experiment 2, as a function of response relation, distractor relation, prime color similarity, and probe color similarity. Error bars depict standard deviations of the means
Again, the interaction between prime color similarity, probe color similarity, and response relation was significant, F(1, 26) = 26.40, p < .001, ηp2 = .50. The RT benefit for response repetitions in comparison to response changes was modulated by an interaction of prime color similarity and probe color similarity. The benefit was larger if the prime and the probe color similarities were the same (i.e., the condition with similar distractors in the prime and similar distractors in the probe, and the condition with dissimilar distractors in the prime and dissimilar distractors in the probe) than in those conditions in which the color similarity of the distractor changed between prime and probe. For the sake of completeness, the interactions between prime color similarity and distractor relation, F(1, 26) = 5.96, p = .022, ηp2 = .19; between prime color similarity and response relation, F(1, 26) = 12.21, p = .002, ηp2 = .32; and the interaction between prime color similarity, probe color similarity, and distractor relation, F(1, 26) = 8.26, p = .008, ηp2 = .24, all reached significance. None of the other effects was significant, all Fs(1, 26) < 2.35, ps > .137.
In the same analysis on error rates, only the interaction between prime color similarity, probe color similarity, and response relation reached significance, F(1, 26) = 5.66, p = .025, ηp2 = .18.
Discussion
The aim of Experiment 2 was to further investigate the modulating effect of color similarity on stimulus–response integration, while excluding a possible influence due to spatial and temporal grouping of the stimuli. To this end, the target and distractor letters were presented sequentially.
The interaction of response relation and distractor relation was again significant, indicating a general effect of distractor–response binding across all conditions, whereas the main effect of distractor relation was not significant (in contrast to Exp. 1). Note that the main effect of distractor relation can be interpreted as reflecting distractor inhibition (Houghton & Tipper, 1994; Tipper, 1985). That is, ignoring the distractor in the prime leads to an inhibited representation of that stimulus. The inhibition of the distractor is assumed to last until presentation of the probe display, leading to better performance if the distractor is repeated than if the distractor is changed, because it is easier to select the target against an already inhibited distractor. This process is independent of distractor-based retrieval effects (Giesen et al., 2012; Singh, Laub, Burgard, & Frings, 2017), and as such is of no relevance to the main results concerning distractor–response binding.
The distractor–response binding effect was not further modulated by probe color similarity. Due to the sequential stimulus presentation, stimulus–response retrieval could be initiated as soon as the distractor was presented in the probe, and hence grouping had no further influence on the already proceeding retrieval process (see Frings & Moeller, 2012, for this logic concerning asynchronous distractors). In contrast, the interaction between response relation, distractor relation, and prime color similarity was significant; that is, the distractor–response binding effect was stronger in conditions with a prime distractor that was similar to the prime target (see Fig. 4). Apparently, the binding process benefited from grouping via color similarity. This might be due to the similarity resolving the temporal sequence between distractor and target and thus generating stimulus grouping over time. To the best of our knowledge, this is the first evidence that has pinpointed the effects of grouping in the prime, during which integration takes place.
As in the previous experiment, this effect was not further modulated by probe processes (i.e., the four-way interaction was again nonsignificant)—suggesting the independence of prime integration and probe retrieval. This is again important to note, as in Experiment 1, since response repetition effects were modulated by prime–probe similarity (see the General Discussion).
Note that as it was displayed 300 ms before target onset, the distractor was apparently presented within a time window that allowed for that integration of distractor and response in the prime. This is in line with the suggestion that the co-occurrence of two events within a particular time window of ± 0.5 s around the response determines whether these events are integrated (Hommel, 2005). In addition, the present results indicate that presentation within the proposed time window is not the only prerequisite for stimulus–response integration to occur. If the target and distractor stimuli are presented in a sequence, grouping over time seems to be necessary for distractor–response integration to occur.
General discussion
The aim of the present study was to investigate the modulating effect of grouping due to color similarity on the two processes contributing to binding effects: namely, stimulus–response integration and stimulus–response retrieval. In two experiments the color similarity between distractor and target was varied independently during the prime (the integration process) and during the probe (the retrieval process). The main difference between Experiment 1 and Experiment 2 was the stimulus configuration. In Experiment 1 the spatial and temporal configuration, as well as the color similarity, suggested grouping of the targets and distractors, as the target letter was presented synchronously with two flanking distractor letters. In Experiment 2 the stimulus configuration, aside from the factor of color similarity, suggested separate distractor and target objects, as the target letter and the distractor letter were presented sequentially, with the distractor letter appearing prior to the target at the same position. In both experiments we found distractor-based binding effects, as indicated by the two-way interaction of distractor relation and response relation. More importantly, the binding effects were modulated by grouping via color in opposite ways.
In Experiment 1, grouping via color similarity had an influence on the retrieval process, but the integration process was not further affected. The distractor–response binding effect was stronger if target and distractor color were dissimilar during the probe (when retrieval started), than if distractor and target were grouped by color similarity. If target and distractor stimuli were already grouped by spatial information as in the flanker configuration, a high saliency of the distractors (due to color dissimilarity) apparently facilitated retrieval, as compared to lower saliency (due to color similarity). Hence, it could be concluded that the distractor’s salience via color dissimilarity can boost the retrieval process in settings with spatially and temporally grouped probe distractor and target stimuli, and consequently can strengthen the distractor–response binding effect. If distractor and target are already grouped during stimulus–response integration and retrieval, further grouping (due to color similarity) seems to be unprofitable for the distractor-based retrieval. This is in line with former findings regarding the role of attention (Moeller & Frings, 2014a) and the role of stimulus function (Ihrke et al., 2011) for the retrieval process. Using similar flanker paradigms, these studies revealed larger binding effects for attended stimuli and for stimuli that were processed more strongly in the probe. Together, the results of Experiment 1 and past research suggest that the stimulus configuration in the probe display affects retrieval: Stimulus configurations that prioritize the retrieval-starting stimulus should enhance the observed binding effects (as was suggested by Hommel et al., 2001).
By contrast, in Experiment 2 the integration process was affected by the variation of color similarity. Moreover, the binding effect was boosted by similar distractor and target colors in the prime, indicating a different mechanism than for the modulation in Experiment 1. In Experiment 2 the distractor letter always preceded the target letter at the same position. This temporal sequence minimized grouping via a spatial and temporal arrangement and hence impeded the integration process. Thus, the sequential presentation prevented sufficient target–distractor grouping for distractor–response integration. Seemingly, in such a case sufficient target–distractor grouping can be ensured over time via color similarity. Hence, the two important findings of Experiment 2 were that (1) the integration process in S–R binding can be influenced and (2) the sensitivity regarding grouping influences differs largely for the two processes. For stimulus–response integration, increased grouping only has an influence if binding is impeded (as in Exp. 2), due to a lack of spatial and temporal target–distractor grouping. Apparently, the binding process needs only a minimum of stimulus grouping in order to integrate the distractor with the response to the target. Any additional grouping or distractor salience does not further modulate integration (as in Exp. 1). These prerequisites and characteristics of grouping differ for the retrieval process. There, if the distractor and target are spatially grouped in the probe, the retrieval process additionally benefits from distractor salience (Exp. 1).
Taken together, our results provide further evidence for the need to distinguish binding effects as resulting from processes of stimulus–response integration and stimulus–response retrieval. Both processes can be independently influenced by factors like salience and grouping and hence need to be considered separately for the investigation of sequential binding effects. This has important implications both for the interpretation of past findings and also for the design of future studies. In particular, the majority of previous research investigated the influence of factors like grouping, salience, or attention on sequential binding effects without being able to differentiate between influences on integration and influences on retrieval processes (e.g., Frings & Rothermund, 2011; Giesen et al., 2012; Hommel, 2005). Nevertheless, conclusions are oftentimes drawn regarding specifically one of the two processes. The present findings call for a much more cautious new interpretation of these studies. For example, if stimuli were spatially grouped during both integration and retrieval, it is likely that any modulation was due to differences in the retrieval and not in the integration process, as a spatially grouped stimulus configuration seems to be sufficient for integration, irrespective of further influences. However, if the two processes are not separately investigated but are influenced in diametrically opposite ways by additional factors, it is possible that the effect on one process could override the effects on the other process. In this case, it is not possible to draw distinct conclusions about which particular process is influenced in what way. For the design of future studies, it should always be considered whether the differentiation of effects on integration and retrieval is of interest. If this is the case, any influencing factor of interest then needs to be manipulated independently during the prime and the probe.
The present findings suggest that a nongrouped probe distractor has an advantage to start a retrieval process, whereas a grouped distractor in the prime enhances the integration process. However, we do not have any relevant data regarding what our participants experienced; that is, we do not have an independent measure of whether the stimulus configuration suggests grouping in the way we interpreted the grouping manipulation. In fact, it is possible that participants were not even aware of the grouping manipulation in the different color similarity conditions. Hence, the effects of our manipulation must not necessarily be mediated by differences in subjective experience, but might affect processing implicitly. Note that previous research has repeatedly shown that grouping does not require attention and is not necessarily perceived consciously (Kimchi & Razpurker-Apfeld, 2004; Russell & Driver, 2005; Shomstein, Kimchi, Hammer, & Behrmann, 2010). Furthermore, the aim of the present study was not to investigate the different subjective perceptions of grouped versus nongrouped stimulus configurations, but to show that the integration and the retrieval process are two independent processes that can be affected in different ways by stimulus configurations that suggest grouping. Even if our interpretation of the color similarity is not independently tested (e.g., by a manipulation check of the subjective impression participants experienced while looking at the stimulus displays; see Frings & Rothermund, 2017), the main point of our data does not hinge on the particular interpretation of the grouping variable.
Furthermore, the presented findings should be separated from findings regarding encoding specificity. According to Neill (1997), episodic retrieval in prime–probe tasks depends on the similarity of the encoding context and the retrieval context. This encoding-specific effect was thus expected and found in the present experiments, as an interaction of response relation, prime color similarity, and probe color similarity. The RT benefit due to response repetition (as compared to response change) from prime to probe was modulated by the similarity between the prime display (encoding context) and the probe display (retrieval context). The benefit was significantly larger if the (dis)similarity between distractor and target was identical in the prime and probe. This finding is also in line with research by Wong (2000), who found evidence for a modulation of repetition priming by prime–probe display similarity, or research by Fox and de Fockert (1998), who presented evidence that the negative-priming effect depended on the extent to which the probe display serves as a retrieval cue for the prime display. However, the modulation of the retrieval process in Experiment 1 and the modulation of the integration process in Experiment 2 by grouping of the distractor and target were independent of encoding-specific effects (indicated by the nonsignificant four-way interactions). The retrieval process in Experiment 1 was stronger if the distractor and target were nongrouped at retrieval in the probe, independent of the encoding context (i.e., whether the distractor and target were grouped or nongrouped at integration in the prime). Similarly, the integration process in Experiment 2 benefited from distractor–target grouping at integration, independent of the retrieval context (i.e., whether distractor and target were grouped or nongrouped at retrieval).
In addition, the present results, together with earlier studies that had targeted integration and retrieval individually, suggest that it is high time for a further specification of the TEC (Hommel et al., 2001). In its original version, the theory did not explicitly differentiate that two processes (i.e., integration and retrieval) are necessary in order to measure binding effects. Instead, that theory mostly focused on feature integration and implicitly attributed modulations due to such factors as intentional weighting or salience to differences in this first process. In an updated version, integration and retrieval processes and their individual characteristics would need to be discussed separately, as indicated by the present study as well as by various other studies from the last few years of research (e.g., Frings & Moeller, 2012; Hommel, Memelink, Zmigrod, & Colzato, 2014; Ihrke et al., 2011; Moeller & Frings, 2014a).
Finally, we would mention that the results of Experiment 2 might also contribute to a better understanding of the relation between binding processes and longer-term learning effects. It is not yet clear whether binding contributes to learning (e.g., Dutzi & Hommel, 2009; Elsner, & Hommel, 2001, 2004; Moeller & Frings, 2014b) or whether binding and learning effects rely on different processes (e.g., Colzato et al., 2006; Herwig & Waszak, 2012; Moeller & Frings, 2017b). Past binding studies focused on integration and retrieval effects with stimuli that occurred simultaneously with or shortly after response completion. Yet, for sequential learning, associations between responses and stimuli that are presented in a temporal sequence seem to be even more relevant. With the findings of Experiment 2, we reported evidence of binding in sequential setups for the first time, which is a prerequisite for the possibility that binding might indeed play a role in sequence learning. Moreover, we identified a factor that ensured binding over time, namely grouping via color similarity. This can be the basis for analyzing the possible role of binding in sequence learning in the future.
In summary, this study reports first evidence for the separate influences of (non)grouping via color (dis)similarity on stimulus–response integration and on stimulus–response retrieval processes in distractor–response binding. Target–distractor color dissimilarity enhanced retrieval, due to distractor repetition in a flanker paradigm. If integration had to be ensured over time (due to asynchronous target and distractor presentation), color similarity enhanced distractor–response integration. On a more general note, binding effects in sequential tasks always reflect a mixture of integration and retrieval processes. Thus, in sequential tasks researchers should carefully consider which process might be responsible for a particular data pattern, because stimulus integration and retrieval processes might be differently affected by experimental conditions.
Author note
The research reported in this article was supported by a grant of the Deutsche Forschungsgemeinschaft to B.M. (MO 2839/2-1).
Notes
To provide further support for this conclusion, we calculated Bayes factors using the Bayes ANOVA module of JASP (Love et al., 2015). The Bayes factor for the four-way interaction of prime color similarity, probe color similarity, response relation, and distractor relation in favor of the null model was BF01 = 4.17. According to the rules of thumb given by Raftery (1995), this value is considered “positive” evidence for the null hypothesis.
To provide further support for this conclusion, we calculated Bayes factors using the Bayes ANOVA module of JASP (Love et al., 2015). The Bayes factor for the four-way interaction of prime color similarity, probe color similarity, response relation, and distractor relation in favor of the null model was BF01 = 3.70. According to the rules of thumb given by Raftery (1995), this value is considered “positive” evidence for the null hypothesis.
References
Ansorge, U., & Heumann, M. (2004). Peripheral cuing by abrupt-onset cues: The influence of color in S–R corresponding conditions. Acta Psychologica, 116, 115–143. https://doi.org/10.1016/j.actpsy.2004.01.001
Baylis, G. C., & Driver, J. (1992). Visual parsing and response competition: The effect of grouping factors. Perception & Psychophysics, 51, 145–162. https://doi.org/10.3758/BF03212239
Colzato, L. S., Raffone, A., & Hommel, B. (2006). What do we learn from binding features? Evidence for multilevel feature integration. Journal of Experimental Psychology: Human Perception and Performance, 32, 705–716. https://doi.org/10.1037/0096-1523.32.3.705
Dutzi, I. B., & Hommel, B. (2009). The microgenesis of action-effect binding. Psychological Research PRPF, 73, 425–435.
Elsner, B., & Hommel, B. (2001). Effect anticipation and action control. Journal of Experimental Psychology: Human Perception and Performance, 27, 229–240. https://doi.org/10.1037/0096-1523.27.1.229
Elsner, B., & Hommel, B. (2004). Contiguity and contingency in action–effect learning. Psychological Research, 68, 138–154.
Eriksen, C. W. (1995). The flankers task and response competition: A useful tool for investigating a variety of cognitive problems. Visual Cognition, 2, 101–118. https://doi.org/10.1080/13506289508401726
Fox, E., & de Fockert, J. W. (1998). Negative priming depends on prime-probe similarity: Evidence for episodic retrieval. Psychonomic Bulletin & Review, 5, 107–113.
Frings, C., Koch, I., Rothermund, K., Dignath, D., Giesen, C., Hommel, B., … Philipp, A. (2017). Merkmalsintegration und Abruf als zentrale Prozesse der Handlungssteuerung—eine Paradigmen-übergreifende Perspektive. Manuscript submitted for publication.
Frings, C., & Moeller, B. (2012). The horserace between distractors and targets: Retrieval-based probe responding depends on distractor–target asynchrony. Journal of Cognitive Psychology, 24, 582–590. https://doi.org/10.1080/20445911.2012.666852
Frings, C., & Rothermund, K. (2011). To be or not to be included in an event file: Integration and retrieval of distractors in stimulus–response episodes is influenced by perceptual grouping. Journal of Experimental Psychology: Learning, Memory, and Cognition, 37, 1209–1227.
Frings, C., & Rothermund, K. (2017). How perception guides action: Figure–ground segmentation modulates integration of context features into S–R episodes. Journal of Experimental Psychology: Learning, Memory, and Cognition, 43, 1720–1729. https://doi.org/10.1037/xlm0000403
Frings, C., Rothermund, K., & Wentura, D. (2007). Distractor repetitions retrieve previous response to targets. Quarterly Journal of Experimental Psychology, 60, 1367–1377. https://doi.org/10.1080/17470210600955645
Giesen, C., Frings, C., & Rothermund, K. (2012). Differences in the strength of distractor inhibition do not affect distractor–response bindings. Memory & Cognition, 40, 373–387. https://doi.org/10.3758/s13421-011-0157-1
Henson, R. N., Eckstein, D., Waszak, F., Frings, C., & Horner, A. J. (2014). Stimulus–response bindings in priming. Trends in Cognitive Sciences, 18, 376–384.
Herwig, A., & Waszak, F. (2012). Action–effect bindings and ideomotor learning in intention- and stimulus-based actions. Frontiers in Psychology, 3, 444:1–18. https://doi.org/10.3389/fpsyg.2012.00444
Holmes, N. P., & Spence, C. (2005). Multisensory integration: Space, time and superadditivity. Current Biology, 15, R762–R764.
Hommel, B. (1998). Event files: Evidence for automatic integration of stimulus–response episodes. Visual Cognition, 5, 183–216. https://doi.org/10.1080/713756773
Hommel, B. (2004). Event files: Feature binding in and across perception and action. Trends in Cognitive Sciences, 8, 494–500. https://doi.org/10.1016/j.tics.2004.08.007
Hommel, B. (2005). How much attention does an event file need? Journal of Experimental Psychology: Human Perception and Performance, 31, 1067–1082. https://doi.org/10.1037/0096-1523.31.5.1067
Hommel, B., Memelink, J., Zmigrod, S., & Colzato, L. S. (2014). Attentional control of the creation and retrieval of stimulus–response bindings. Psychological Research, 78, 520–538.
Hommel, B., Müsseler, J., Aschersleben, G., & Prinz, W. (2001). The Theory of Event Coding (TEC): A framework for perception and action planning. Behavioral and Brian Sciences, 24, 849–878, disc. 878–937. https://doi.org/10.1017/S0140525X01000103
Houghton, G., & Tipper, S. P. (1994). A model of inhibitory mechanisms in selective attention. In D. Dagenbach & T. H. Carr (Eds.), Inhibitory processes in attention, memory, and language (pp. 53–112). San Diego: Academic Press.
Ihrke, M., Behrendt, J., Schrobsdorff, H., Herrmann, M., & Hasselhorn, M. (2011). Response-retrieval and negative priming. Experimental Psychology, 58, 154–161.
Kimchi, R., & Razpurker-Apfeld, I. (2004). Perceptual grouping and attention: Not all groupings are equal. Psychonomic Bulletin & Review, 11, 687–696. https://doi.org/10.3758/BF03196621
Logan, G. D. (1980). Attention and automaticity in Stroop and priming tasks: Theory and data. Cognitive Psychology, 12, 523–553. https://doi.org/10.1016/0010-0285(80)90019-5
Love, J., Selker, R., Marsman, M., Jamil, T., Dropmann, D., Verhagen, A. J., & Wagenmakers, E.-J. (2015). JASP (Version 0.7) [Computer software]. Amsterdam: JASP Project.
Moeller, B., & Frings, C. (2014a). Attention meets binding: Only attended distractors are used for the retrieval of event files. Attention, Perception, & Psychophysics, 76, 959–978. https://doi.org/10.3758/s13414-014-0648-9
Moeller, B., & Frings, C. (2014b). Long-term response–stimulus associations can influence distractor–response bindings. Advances in Cognitive Psychology, 10, 68–80.
Moeller, B., & Frings, C. (2015). Distractor–response binding in dual task scenarios. Visual Cognition, 23, 516–531. https://doi.org/10.1080/13506285.2015.1041437
Moeller, B., Pfister, R., Kunde, W., & Frings, C. (2016). A common mechanism behind distractor-response and response-effect binding?. Attention, Perception, & Psychophysics, 78, 1074–1086.
Moeller, B., & Frings, C. (2017a). Dissociation of binding and learning processes. Attention, Perception, & Psychophysics, 79, 2590–2605. https://doi.org/10.3758/s13414-017-1393-7
Moeller, B., & Frings, C. (2017b). Overlearned responses hinder S–R binding. Journal of Experimental Psychology: Human Perception and Performance, 43, 1–5. https://doi.org/10.1037/xhp0000341
Moeller, B., Rothermund, K., & Frings, C. (2012). Integrating the irrelevant sound. Grouping modulates the integration of irrelevant auditory stimuli into event files. Experimental Psychology, 59, 258–264. https://doi.org/10.1027/1618-3169/a000151
Neill, W. T. (1997). Episodic retrieval in negative priming and repetition priming. Journal of Experimental Psychology: Human Perception and Performance, 32, 1291–1305. https://doi.org/10.1037/0278-7393.23.6.1291
Raftery, A. E. (1995). Bayesian model selection in social research. Sociological Methodology, 25, 111–163, disc. 165–195.
Russell, C., & Driver, J. (2005). New indirect measures of “inattentive” visual grouping in a change-detection task. Perception & Psychophysics, 67, 606–623. https://doi.org/10.3758/BF03193518
Shomstein, S., Kimchi, R., Hammer, M., & Behrmann, M. (2010). Perceptual grouping operates independently of attentional selection: Evidence from hemispatial neglect. Attention, Perception, & Psychophysics, 72, 607–618. https://doi.org/10.3758/APP.72.3.607
Singh, T., Laub, R., Burgard, J. P., & Frings, C. (2017). Disentangling inhibition-based and retrieval-based aftereffects of distractors: Cognitive versus motor processes. Journal of Experimental Psychology: Human Perception and Performance. Advance online publication. https://doi.org/10.1037/xhp0000496
Spence, C. (2013). Just how important is spatial coincidence to multisensory integration? Evaluating the spatial rule. Annals of the New York Academy of Sciences, 1296, 31–49.
Stoet, G., & Hommel, B. (1999). Action planning and the temporal binding of response codes. Journal of Experimental Psychology: Human Perception and Performance, 25, 1625–1640. https://doi.org/10.1037/0096-1523.25.6.1625
Tipper, S. P. (1985). The negative priming effect: Inhibitory priming by ignored objects. Quarterly Journal of Experimental Psychology, 37, 571–590.
Tipper, S. P. (2001). Does negative priming reflect inhibitory mechanisms? A review and integration of conflicting views. Quarterly Journal of Experimental Psychology, 54A, 321–343. https://doi.org/10.1080/713755969
Tipper, S. P., Weaver, B., & Houghton, G. (1994). Behavioural goals determine inhibitory mechanisms of selective attention. Quarterly Journal of Experimental Psychology, 47A, 809–840. https://doi.org/10.1080/14640749408401098
Treisman, A. M., & Gelade, G. (1980). A feature-integration theory of attention. Cognitive Psychology, 12, 97–136. https://doi.org/10.1016/0010-0285(80)90005-5
Tukey, J. W. (1977). Exploratory data analysis. Reading: Addison-Wesley.
van Dam, W. O., & Hommel, B. (2010). How object-specific are object files? Evidence for integration by location. Journal of Experimental Psychology: Human Perception and Performance, 36, 1184–1192.
Waszak, F., & Pholulamdeth, V. (2009). Episodic S–R bindings and emotion: About the influence of positive and negative action effects on stimulus–response associations. Experimental Brain Research, 194, 489–494.
Wiswede, D., Rothermund, K., & Frings, C. (2013). Not all errors are created equally: Specific ERN responses for errors originating from distractor-based response retrieval. European Journal of Neuroscience, 38, 3496–3506. https://doi.org/10.1111/ejn.12340
Wong, K. F. E. (2000). Dissociative prime–probe contextual similarity effects on negative priming and repetition priming: A challenge to episodic retrieval as a unified account of negative priming. Journal of Experimental Psychology: Learning, Memory, and Cognition, 26, 1411–1422. https://doi.org/10.1037/0278-7393.26.6.1411
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Laub, R., Frings, C. & Moeller, B. Dissecting stimulus–response binding effects: Grouping by color separately impacts integration and retrieval processes. Atten Percept Psychophys 80, 1474–1488 (2018). https://doi.org/10.3758/s13414-018-1526-7
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-018-1526-7