
Abstract
Administration of therapeutic genes to human osteoarthritic (OA) cartilage is a potential approach to generate effective, durable treatments against this slow, progressive disorder. Here, we tested the ability of recombinant adeno-associated virus (rAAV)-mediated overexpression of human insulinlike growth factor (hIGF)-I to reproduce an original surface in human OA cartilage in light of the pleiotropic activities of the factor. We examined the proliferative, survival and anabolic effects of the rAAV-hIGF-I treatment in primary human normal and OA chondrocytes in vitro and in explant cultures in situ compared with control (reporter) vector delivery. Efficient, prolonged IGF-I secretion via rAAV stimulated the biological activities of OA chondrocytes in all the systems evaluated over extended periods of time, especially in situ, where it allowed for the long-term reconstruction of OA cartilage (at least for 90 d). Remarkably, production of high, stable amounts of IGF-I in OA cartilage using rAAV advantageously modulated the expression of central effectors of the IGF-I axis by downregulating IGF-I inhibitors (IGF binding protein (IGFBP)-3 and IGFBP4) while upregulating key potentiators (IGFBP5, the IGF-I receptor and downstream mitogen-activated protein kinase/extracellular signal-regulated kinase 1/2 (MAPK/ERK-1/2) and phosphatidylinisitol-3/Akt (PI3K/Akt) signal transduction pathways), probably explaining the enhanced responsiveness of OA cartilage to IGF-I treatment. These findings show the benefits of directly providing an IGF-I sequence to articular cartilage via rAAV for the future treatment of human osteoarthritis.
Similar content being viewed by others
Introduction
Osteoarthritis (OA) is a progressive disease of the entire joint that affects millions of people worldwide. OA is mainly characterized by a gradual, irreversible degeneration of the articular cartilage with a loss of its major matrix components (proteoglycans, type II collagen) (1), with concomitant changes in the subchondral bone and synovium. Although several pharmacological treatment options and surgical interventions are available to manage the progression of OA, regeneration of the articular cartilage remains an unresolved problem, in particular for patients who are too young to undergo partial or total joint replacement.
Disturbances in cartilage homeostasis are believed to play determining roles in the pathogenesis and progression of OA. Proinflammatory cytokines (interleukin [IL]-1 and tumor necrosis factor [TNF]-α) and adipokines (leptin, adiponectin and resistin) locally produced by the inflamed synovium, infrapatellar fat pad, osteophytes or the chondrocytes themselves may all contribute to the pathophysiology of OA (2–5). The putative implication of articular chondrocytes during OA progression has received particular attention because of pathological changes in their gene expression patterns, their incapacity to restore natural levels of extracellular matrix components and an ability to produce increased amounts of matrix-degrading enzymes. Interestingly, several investigations have focused on these cells, the unique cellular components in adult articular cartilage, as targets to develop new therapeutic interventions.
Delivery of candidate genes to articular chondrocytes is an attractive strategy that has the potential to allow for a durable reestablishment of the structural integrity in OA cartilage. Gene transfer approaches might be better suited to treat a slow and irreversible disorder such as OA over time instead of systems on the basis of the application of recombinant factors with relatively short pharmacological half-lives. Current approaches that aim at re-equilibrating the metabolic balance in OA cartilage are on the basis of the transfer of sequences coding for agents that either counteract the processes of matrix degradation or enhance the synthesis of matrix components. Protective effects against cartilage breakdown have been documented in experimental models of OA using sequences coding for inhibitors of inflammatory pathways (an IL-1 receptor antagonist [IL-1Ra], soluble TNF receptor [sTNFR], tissue inhibitor of metalloproteinases [TIMP-1]) (6–14) or factors such as IL-10 (12), heat shock protein 70 (15), glutamine:fructose-6-phosphate amidotransferase (16), thrombospontin-1 (17), kallistatin (18) and inhibitors of nuclear factor κB (19). Even though application of these stimuli was capable of containing cartilage degradation, it was not sufficient to compensate for the loss of matrix elements and cells and to reestablish an original cartilage surface. Growth, transcription and enzymatic factors are potent candidates to achieve these goals because of their anabolic or mitogenic properties such as fibroblast growth factor 2 (FGF-2) (20,21), bone morphogenetic proteins 2 and 4 (BMP-2 and BMP-4) (22,23), transforming growth factor β (TGF-β) (22,24,25), transcription factor sex-determining region Y-type high mobility group box 9 (SOX9) (20,26,27), glucuronosyltransferase-I (28), bcl-2 (29) and telomerase (30). A critical agent that may have the strongest value to readjust the disturbed homeostasis in OA cartilage is insulinlike growth factor (IGF)-I, since it has the ability to influence concomitantly metabolic and proliferative processes, affording protection against extracellular matrix degradation in horse and rabbit articular cartilage explant cultures experimentally treated with proinflammatory cytokines (13,14,22).
Importantly for the treatment of OA, the development of effective gene treatments will necessitate that the gene vehicle allows for high and sustained levels of expression of the candidate sequence due to the slow and irreversible progression of this disorder. In contrast with vectors derived from adenoviruses (6),10,13,14,16–19,21,22,25) and retro-viruses (7,11,12,23,26,30) or with nonviral compounds (8,15,28,29), systems based on the replication-defective, nonpathogenic human adeno-associated virus (AAV) may provide better tools for OA, since recombinant AAV (rAAV) can deliver genes in nondividing cells such as chondrocytes both in vitro and in situ in their dense extracellular matrix at high efficiencies and for extended periods of time (20,24,27,31). Also, removal of the viral protein coding sequences in rAAV make them less immunogenic than adenoviral vectors characterized by short-term transgene expression levels (32).
In the present study, we tested the hypothesis that efficient and prolonged IGF-I overexpression can be achieved via rAAV in primary human normal and OA chondrocytes in monolayer and three-dimensional hydrogel cultures in vitro and, most importantly, within their extracellular matrix in articular cartilage explants in situ, leading simultaneously to enhanced levels of cell proliferation and survival and of matrix synthesis vis-à-vis control (reporter gene vector) treatment. We also analyzed the extent by which the candidate treatment with the rAAV-human IGF-I (rAAV-hIGF-I) vector was capable of durably restoring the structure of human OA cartilage vis-à-vis normal cartilage. Finally, we investigated the influence of rAAV-mediated IGF-I production upon the expression of key effectors of the IGF-I axis, specifically the IGF-binding proteins 3, 4 and 5 (IGFBP-3 to IGFBP-5) as well as the IGF-I receptor (IGF-IR) and downstream mitogen-activated protein kinase/extracellular signal-regulated kinase 1/2 (MAPK/ERK-1/2) and phosphatidylinisitol-3/Akt (PI3K/Akt) signal transduction pathways (33–39), reported as being inhibitors (IGFBP3 and IGFBP4) (40–42) or potentiators (IGFBP5 and IGF-IR) (42,43) of the IGF-I actions during OA pathogenesis.
Materials and Methods
Reagents
Reagents were from Sigma (Munich, Germany) unless indicated. The dimethylmethylene blue dye was from Serva (Heidelberg, Germany). The anti-β-gal (GAL-13) and anti-bromodeoxyuridine (BrdU) (BU-33) antibodies were from Sigma; the anti-IGF-I antibody (AF-291-NA) was from R&D Systems (Wiesbaden-Nordenstadt, Germany); the anti-type I collagen (AF-5610) and anti-type II collagen (AF-5710) antibodies were from Acris (Hiddenhausen, Germany); the anti-IGF-I receptor (IGF-IR) antibody (Ab-4) was from Oncogene Research (Merck, Darmstadt, Germany); the anti-IGF binding proteins 3, 4 and 5 (IGFBP3, IGFBP4 and IGFBP5) (YY07, C-20 and E87, respectively) and anti-SOX9 (C-20) antibodies were from Santa Cruz Biotechnology (Heidelberg, Germany); and the anti-phospho-ERK-1/2 (D13.14.4E), anti-phospho-Akt (D9E) and anti-phospho-Elk-1 (2B1) antibodies were from Cell Signaling Technology (New England Biolabs, Frankfurt am Main, Germany). Biotinylated secondary antibodies and ABC reagent were from Vector Laboratories (Alexis Deutschland, Grünberg, Germany). Production of IGF-I, IGFBP3, IGFBP4 and IGFBP5 was measured with Quantikine IGF-I and Quantikine IGFBP3 enzyme-linked immunosorbent assays (ELISAs) (DG100 and DGB300, respectively) and the Duo-Set IGFBP-4 and IGFBP5 ELISAs (DY804 and DY875, respectively) (all from R&D Systems). The type I and type II collagen ELISAs (Arthrogen-CIA Capture ELISAs) were from Chondrex (Redmond, WA, USA), the cell proliferation reagent WST-1 was from Roche Applied Science (Mannheim, Germany), and apoptosis was determined using the Cell Death Detection ELISAPLUS (Roche Applied Science) and ApopTag® Plus Peroxidase In Situ Apoptosis Detection Kit (Chemicon-Millipore, Schwalbach/Ts., Germany).
Cartilage and Cells
Human normal articular cartilage was obtained from unaffected areas in knee joints removed during tumor surgery (nine patients, 68–72 years of age). OA was excluded on safranin O-stained sections according to the Mankin scale (Mankin score 1–2) (44). Human OA articular cartilage was obtained from joints undergoing total knee arthroplasty (14 patients, 67–77 years of age) (Mankin score 7–9). The study was approved by the Ethics Committee of the Saarland Physicians Council. Cartilage explant cultures (6.2-mm diameter, 2-mm thick) and articular chondrocytes were prepared as previously described (20,27,31,45). Human anterior cruciate ligament (ACL) was obtained in patients undergoing total knee arthroplasty (three patients, 70–76 years of age), and primary human ACL fibroblasts were prepared as previously described (46).
Plasmids and rAAV Vectors
The constructs and pACP were derived from pSSV9, an AAV-2 genomic clone (47,48). pAd8 contains the AAV-2 replication and encapsidation functions (48). rAAV-lacZ carries the lacZ gene for β-galactosidase (β-gal) under the control of the cytomegalovirus immediate-early (CMV-IE) promoter (20,27,31,45,49,50). rAAV-red fluorescent protein (rAAV-RFP) carries a 776-bp Discosoma sp. RFP cDNA fragment (45,49,50). A hIGF-I cDNA (51) was generated by polymerase chain reaction (PCR) using the primers 5′-I-A (A5ctgcag[Pst I]G17CTTCA GAAGC A) and 3′-I-A (A5aagctt[Hind III]TGCGG TGGCA TGTCA CTCTT CAC) with pCMVhIGF-I (52) as a template for amplification. The resulting hIGF-I sequence (536 bp) was cloned in pACP to generate rAAV-hIGF-I, where the hIGF-I fragment was confirmed by sequencing. rAAVs were packaged as conventional (not self-complementary) vectors in the 293 cell line, an adenovirus-transformed human embryonic kidney cell line, using adenovirus 5 and pAd8 for helper functions. The preparations were purified by dialysis and titered by real-time PCR (20,27,31,45,49,50), averaging 1010 transgene copies/mL (ratio virus particles to functional vectors = 500/1) (49).
rAAV Gene Transfer
Chondrocytes (passages 2–3, 10 d of culture) in monolayer culture (4 × 104 cells) were transduced with rAAV (multiplicity of infection [MOI] = 20) and kept in Dulbecco’s modified Eagle’s medium, 100 U/mL penicillin G, 100 µL/mL streptomycin and 10% fetal bovine serum (growth medium) in a humidified atmosphere of air with 5% CO2 at 37°C for up to 20 d (45). Chondrocytes (106) were also transduced with rAAV (MOI = 6) for 2 d and encapsulated in alginate spheres in growth medium for up to 26 d (27,31,45). For comparison, human ACL fibroblasts in monolayer culture (4 × 104 cells) were transduced with increasing doses of rAAV (MOI = 20, 200 or 2,000) and kept in growth medium in a humidified atmosphere of air with 5% CO2 at 37°C for up to 20 d.
Cartilage explant cultures were transduced by direct application of rAAV (4 × 108 functional vectors) to the surface of the samples downwards during 1–2 min of contact (27,31,45). Growth medium was then added to the cultures without removal of the vector solution to allow for further in-depth penetration of the viral particles. The explants were then maintained in growth medium for up to 90 d with regular medium change every 2–3 d, starting on d 2 after vector administration.
Transgene Expression
RFP was detected by live fluorescence using a fluorescent microscope with a 568-nm filter (Olympus CKX41, Hamburg, Germany) (20,27,31,45,50). To assess IGF-I secretion, samples were washed twice and placed for 24 h in serum-free medium. Supernatants were collected and centrifuged, and IGF-I production was measured by ELISA on a GENios spectrophotometer/fluorometer (Tecan, Crailsheim, Germany). Expression was also monitored by immunocytochemical and immunohistochemical analyses with specific primary antibodies, biotinylated secondary antibodies and the ABC method using diaminobenzidine (DAB) as the chromogen (20,27,31,50). To control for secondary immunoglobulins, samples were processed with omission of primary antibodies. Samples were examined under light microscopy (Olympus BX 45). Western blotting analyses were performed using 15-µg proteins prepared from transduced cells. Signals were revealed with specific antibodies, horseradish peroxidase-labeled secondary antibodies (Vector Laboratories) and the ECL Advance Western blotting detection kit (Amersham Biosciences, Freiburg, Germany) (20,27).
Histological, Immunocytochemical and Immunohistochemical Analyses
Monolayer, alginate and explant cultures were harvested and fixed in 10% buffered formalin. Spheres and explants were dehydrated in graded alcohols and embedded in paraffin. Paraffin-embedded sections (5 µm) were stained with safranin O (proteoglycans) and hematoxylin and eosin (H&E) (cells) (20,27,31,45). Expression of type I and type II collagen, IGF-IR, IGFBP3, IGFBP4, IGFBP5, phospho-ERK-1/2, phospho-Akt, phospho-Elk-1 and SOX9 was detected using specific antibodies, biotinylated secondary antibodies, the ABC method and DAB, as described above.
Cell Proliferation
Proliferative activities were assessed by immunolabeling after BrdU incorporation (50). BrdU was introduced at a final concentration of 3 µg/mL in the culture medium 24 h after rAAV transduction. Samples were immunochemically processed as described above to monitor proliferation rates with a specific anti-BrdU antibody and a biotinylated secondary antibody and by revelation using the ABC method and DAB. Proliferation was also assessed using the cell proliferation reagent WST-1, with optical density (OD) being proportional to the cell numbers (50).
Apoptosis Assays
In vitro, the specific enrichment of cytoplasmic histone-associated DNA fragments (mono- and oligonucleosomes) released from monolayer cultures was estimated using the Cell Death Detection ELISAPLUS according to the manufacturer’s protocol. In situ, nuclear DNA fragmentation consistent with apoptosis was determined by the terminal deoxynucleotidyl transferase-mediated dUTP nickend labeling (TUNEL) method (20).
Morphometric Analyses
The cell numbers and viability were monitored by trypan blue exclusion and by counting cells on H&E-stained histological sections (20,27,31,45,49). The transduction efficiencies; the percentage of cells positive for BrdU uptake; apoptotic events; IGF-IR, IGFBP3, IGFBP4, IGFBP5, phospho-ERK-1/2, phospho-Akt, phospho-Elk-1 and SOX9 immunoreactivity; the cell densities; and the intensities of safranin O staining and of type I and type II collagen immunostaining were measured at three standardized sites or by using 10 serial histological and immunohistochemical sections for each parameter, test and replicate condition. Analysis programs included SIS AnalySIS (Olympus), Adobe Photoshop (Adobe Systems, Unterschleissheim, Germany) and Scion Image (Scion Corporation, Frederick, MD, USA) (20,27,31,45,49,50). The safranin O staining intensities were in percentages as the ratios of positively stained tissue surface to the total surface of the site evaluated. The type I and type II collagen immunostaining intensities were in pixels per standardized area (intensity units).
Biochemical Assays
Solubilized spheres and explant cultures were digested to monitor the DNA contents, proteoglycan production and type I and type II collagen contents by Hoechst 33258 assay, binding to the dimethylmethylene blue (DMMB) dye and ELISA, respectively (20,27,31,50). Data were normalized to total cellular proteins using a protein assay (Pierce Thermo Scientific, Fisher Scientific, Schwerte, Germany). To assess IGFBP secretion, samples were washed twice and placed for 24 h in serum-free medium. Supernatants were collected and centrifuged, and production of IGFBP3, IGFBP4 and IGFBP5 was measured by ELISA. All measurements were performed with a GENios spectrophotometer/fluorometer.
Statistical Analysis
Each condition was performed in triplicate in three independent experiments with monolayer, alginate and explant cultures. Data were obtained by three individuals blinded with respect to the groups. The t test and Mann-Whitney rank sum test were used where appropriate. P values <0.05 were considered statistically significant.
Results
Overexpression of IGF-I via rAAV in Human Normal and OA Chondrocytes In Vitro
The candidate (rAAV-hIGF-I) and control (rAAV-RFP or rAAV-lacZ) vectors were first applied to primary human normal and OA chondrocytes in monolayer culture to define transgene expression in vitro. A strong fluorescent signal restricted to cells transduced with rAAV-RFP was already detected on d 5 (not shown) and for at least 20 d compared with rAAV-hIGF-I (Figure 1A), with transduction efficiencies ranging between 80% and 95% when applying an MOI of 20. For comparison, a similar signal was noted in rAAV-RFP-transduced primary human fibroblasts only when an MOI of at least 2,000 was provided to the cells by d 20 (transduction efficiencies of 60–71%) (Figure 1B). Conversely, strong IGF-I immunoreactivity was noted in cells where rAAV-hIGF-I was provided vis-α-vis control treatment (Figure 1C). Western blotting analyses revealed an IGF-I immunoreactive band of approximately 7.6 kDa that was about four-fold more intense in rAAV-hIGF-I-treated OA cells than in control cells (Figure 1E). Similar results were noted in normal chondrocytes. Significant, prolonged synthesis of IGF-I was achieved with rAAV-hIGF-I compared with control conditions (at least an 8.6-fold increase in normal cells and a 7.3-fold increase in OA cells; P ≤ 0.001) (Table 1), with well-maintained levels in both types of cells over time (P ≥ 0.200 between d 5 and d 20) (see Table 1).

Detection of transgene expression in monolayer and alginate cultures of rAAV-transduced human articular chondrocytes and human ACL fibroblasts. Primary human normal and OA chondrocytes were transduced with either rAAV-hIGF-I or a control vector (rAAV-RFP or rAAV-lacZ) for maintenance in monolayer (A, C, E: MOI = 20) or alginate culture (D: MOI = 6). The vectors were also applied to monolayer cultures of primary human ACL fibroblasts using increasing vector doses (B: MOI = 20, 200 or 2,000). Transgene expression was detected by live fluorescence (A: magnification 10×; B: magnification 20×; insets: same fields under transmitted light; d 20 after transduction), anti-IGF-I immunocyto- and immunohistochemistry (C: d 20 after transduction; D: d 26 after transduction; both: magnification 20×) and Western blotting analysis (E: lane 1, rAAV-lacZ-transduced OA chondrocytes; lane 2, rAAV-hIGF-I-transduced OA chondrocytes; d 5 after transduction).
Transduced cells were next encapsulated in alginate spheres to characterize transgene expression in a three-dimensional environment supportive of the chondrocyte phenotype. β-Gal activity, restricted to control cells, was already detected on the day of encapsulation and for at least 26 d (not shown). Significant, prolonged IGF-I expression and synthesis was seen when rAAV-hIGF-I was provided compared with control treatment (at least a 9.3-fold increase in normal cells and a 6.6-fold increase in OA cells; P ≤ 0.001) (Figure 1D and Table 1), with well-maintained levels over time (P ≥ 0.165 between d 5 and d 26) (Table 2). Transduction efficiencies were again between 80% and 92%.
Effects of IGF-I Overexpression on the Proliferative and Synthetic Activities of Human Normal and OA Chondrocytes In Vitro
rAAV-hIGF-I was next administered to chondrocytes in monolayer and alginate cultures to examine the effects of IGF-I overexpression on cell proliferation and survival in vitro over time compared with rAAV-lacZ treatment. Immunodetection of BrdU incorporation in monolayer cultures revealed significantly increased levels of proliferation mediated by rAAV-hIGF-I vis-à-vis rAAV-lacZ (75–80% versus <10% of BrdU+ normal or OA cells on d 20 [that is, up to eightfold difference]; P ≤ 0.001) (Figure 2A). These results were corroborated by a WST-1 assay performed at the same time point (0.628 versus 0.312 OD450nm in normal cells or 1.175 versus 0.716 OD450nm in OA cells using rAAV-hIGF-I versus rAAV-lacZ [that is, a 2- or 1.6-fold increase]; P ≤ 0.001). Also, the apoptotic events significantly decreased in the presence of rAAV-hIGF-I on d 20 compared with rAAV-lacZ (normal cells: a 1.15 versus 1.50 factor [that is, a 1.3-fold decrease]; OA cells: a 0.89 versus 1.62 factor [that is, a 1.8-fold decrease]; P ≤ 0.001). The levels of proliferation in encapsulated cells were also significantly enhanced by treatment with rAAV-hIGF-I compared with the controls (70–82% versus <12% of BrdU+ normal or OA cells on d 26 [that is, up to seven-fold difference]; P ≤ 0.001) (Figure 2B), a result confirmed when estimating the viable cell numbers in rAAV-hIGF-I-transduced spheres vis-à-vis control spheres (up to 38.8-fold difference; P ≤ 0.001) (see Table 2), the cell densities on histological sections (up to 9.8-fold difference; P ≤ 0.001) (Figure 2C and Table 2), and the DNA contents (up to 27.3-fold difference; P ≤ 0.001) (see Table 2). Interestingly, a significant decrease in the viable cell numbers, cell densities and DNA contents was noted over time in both types of control spheres (up to 35-fold; P ≤ 0.001), whereas these parameters were well maintained in the rAAV-hIGF-I-treated spheres (P ≥ 0.598), probably due to the steady expression levels of the IGF-I transgene in cells (see Table 1).

Analysis of cell proliferation and extracellular matrix synthesis in rAAV-transduced human articular chondrocytes in monolayer and alginate cultures. Primary human normal and OA chondrocytes were transduced with either rAAV-lacZ or rAAV-hIGF-I, as described in Figure 1, and cell encapsulation was performed where indicated, as also denoted in Figure 1. Monolayer cultures were processed for the evaluations on d 20 after transduction and alginate cultures on d 26. (A, B) BrdU immunodetection in monolayer (A: magnification 10×) and alginate (B: magnification 20×) cultures of transduced chondrocytes after BrdU incorporation. (C) H&E staining of histological sections from transduced alginate cultures (magnification 20×). (D) Immunodetection of type II collagen on sections from transduced alginate cultures (magnification 20×).
rAAV-hIGF-I was next provided in the alginate system to determine the effects of IGF-I on the synthesis of extracellular matrix components in three-dimensional culture conditions in vitro over time vis-à-vis rAAV-lacZ. The proteoglycan and type II collagen contents were significantly enhanced in spheres from both types of cells using rAAV-hIGF-I versus rAAV-lacZ (up to 6.8-fold difference; P ≤ 0.001) (see Table 2). The results related to the type II collagen were corroborated by an analysis of the staining intensities for this marker on sections from spheres (up to 5.4-fold difference; P ≤ 0.001) (Figure 2D and Table 2). Notably, a significant decrease in the amounts of proteoglycans and type II collagen and in the intensities of type II collagen immunoreactivity was observed over time in both types of control spheres (up to 1.3-fold; P ≤ 0.001), whereas these parameters significantly increased in the rAAV-hIGF-I-treated spheres (up to 2.4-fold; P ≤ 0.001), probably because of the steady expression levels of the IGF-I transgene in cells (see Table 1). Little or no expression of type I collagen was detected in any of the treatment groups when the samples were processed either by immunohistochemistry or by ELISA to monitor the type I collagen contents.
Expression of the IGF-I Transgene in Human Normal and OA Articular Cartilage After Direct rAAV Application In Situ
The candidate rAAV-hIGF-I vector was then directly applied to articular cartilage explant cultures to monitor IGF-I expression over time in a native environment compared with rAAV-lacZ treatment. Immunoreactivity to the growth factor was preferentially detected when rAAV-hIGF-I was provided to the explants (normal and OA cartilage), both in the superficial and middle zones, as early as 5 d after vector application (not shown) and for at least 90 d (Figure 3A). About 72–81% of the cells expressed the transgene compared with the controls (below 4%; up to 20.3-fold difference; P ≤ 0.001). Significant, prolonged IGF-I synthesis was achieved in rAAV-hIGF-I-transduced explants compared with control treatment (at least a 294.6-fold increase in normal explants and a 413.3fold increase in OA explants; P ≤ 0.001) (see Table 1), with increasing levels detected in both types of rAAV-hIGF-I-treated cultures over time (1.9- and 2.6-fold increase between d 5 and d 90 in normal and OA explants, respectively; P ≤ 0.001) (see Table 1).

Immunohistochemical analyses in rAAV-transduced human articular cartilage. Primary human normal and OA cartilage explant cultures were transduced with either rAAV-hIGF-I or the control rAAV-lacZ vector (4 × 108 each functional vector) and processed after 90 d to detect transgene expression by anti-IGF-I immunohistochemistry (A: magnification 20×), the expression of major extracellular matrix components (B: safranin O at magnification 10× and type II collagen in the insets at magnification 10×), the rates of cell proliferation after BrdU incorporation and immunodetection (C: magnification 20×) and apoptotic events by TUNEL assay (D: magnification 4×). All are views of the middle zone.
Effects of IGF-I Overexpression on the Proliferative, Synthetic and Restorative Activities in Human Normal and OA Articular Cartilage In Situ
rAAV-hIGF-I was next provided to cartilage explant cultures to evaluate the effects of IGF-I overexpression on the proliferative, survival and metabolic activities of normal and OA chondrocytes in a native environment compared with rAAV-lacZ treatment. An histomorphometric analysis revealed significant increases in the cell densities of the explants transduced with rAAV-hIGF-I vis-à-vis rAAV-lacZ for at least 90 d, the longest time point examined, both in the superficial and middle zones of the cartilage (a 2.8-fold increase in either normal or OA cartilage; P ≤ 0.001) (Figure 3B and Table 3). These results were confirmed when evaluating the percentage of BrdU+ cells (28- and 19.3-fold increases in normal and OA cartilage, respectively; P ≤ 0.001) (Figure 3C and Table 3) and the DNA contents (a 1.9-fold increase in either normal or OA cartilage; P ≤ 0.001) (see Table 3), overall reflecting the long-term stimulatory effects of the IGF-I treatment on cell proliferation in situ, probably due to the steady expression of the factor (see Table 1). Of note, these parameters were higher in rAAV-hIGF-I-treated OA cartilage than in control normal cartilage (P ≤ 0.001). A TUNEL analysis further showed that application of the IGF-I vector promoted a significant decrease in the percentage of apoptotic cells in OA cartilage vis-à-vis rAAV-lacZ treatment (9.8-fold; P ≤ 0.001), bringing back the levels to those observed in control normal cartilage (almost undetectable levels) (Figure 3D and Table 3).
Remarkably, transduction with rAAV-hIGF-I compared with control conditions led to significant increases in safranin O staining intensity for at least 90 d, both in the superficial and middle zones of the cartilage (2- and 8.5-fold increases in normal and OA cartilage, respectively; P ≤ 0.001) (Figure 3B and Table 3), in the proteoglycan contents (3.7- and 8.7-fold increases, respectively; P ≤ 0.001) (see Table 3), in type II collagen immunostaining intensity (2.3- and 4.3-fold increases, respectively; P ≤ 0.001) (insets of Figure 3B and Table 3) and in the type II collagen contents (2.8- and 4.7-fold increases, respectively; P ≤ 0.001) (see Table 3). Interestingly, expression of these components was higher in rAAV-hIGF-I-treated OA cartilage than in control normal cartilage (P ≤ 0.001). In good agreement with the findings in vitro, little or no type I collagen expression was detected in any of the treatment groups when the samples were processed either by immunohistochemistry (superficial and middle zones) or by ELISA to monitor the type I collagen contents (53).
Regulation of Central Effectors of the IGF-I Axis by rAAV-Mediated IGF-I Overexpression
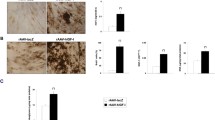
rAAV-hIGF-I was next provided to human normal and OA articular cartilage explant cultures to determine the possible effects of rAAV-mediated IGF-I production on the expression of components of the IGF-I axis known to play key roles in the pathogenesis of OA, precisely the IGF-binding proteins 3, 4 and 5 (IGFBP-3 to IGFBP-5) as well as IGF-IR and downstream MAPK/ERK-1/2 and PI3K/Akt signal transduction pathways. Remarkably, treatment with the rAAV IGF-I vector promoted a significant decrease in IGFBP3 and IGFBP4 expression, reaching almost undetectable levels both in normal and OA cartilage vis-à-vis control vector application after 90 d, the longest time point evaluated (up to 35- and 67-fold in normal and OA cartilage, respectively; P ≤ 0.001) (Figures 4A, B; Table 3). In good agreement with previous observations (54–58), expression of these markers was higher in control OA versus control normal cartilage.

Expression of IGFBPs and IGF-IR in rAAV-transduced human articular cartilage. Primary human normal and OA cartilage explant cultures were transduced with either rAAV-hIGF-I or the control rAAV-lacZ vector, as described in Figure 3, and processed after 90 d to detect the expression of IGFBP3 (A), IGFBP4 (B), IGFBP5 (C) and IGF-IR (D). Magnification 20×. All are views of the middle zone.
Strikingly, and in marked contrast with the observations for IGFBP3 and IGFBP4, application of rAAV-hIGF-I significantly increased IGFBP5 and IGF-IR expression both in normal and OA cartilage vis-à-vis control vector application after 90 d (up to 3.3- and to 1.7-fold in normal and OA cartilage, respectively; P ≤ 0.001) (Figures 4C, D; Table 3). This effect mediated by IGF-I overexpression was more substantial in normal cartilage than in OA cartilage, since the expression levels of these markers were lower in control normal versus control OA cartilage, consistent with previous findings (54,56,59). Remarkably, these effects of IGF-I via rAAV gene transfer in normal and OA cartilage were associated with significant increases in the expression levels of downstream components of the IGF-IR signal transduction pathways, that is, MAPK/ERK-1/2 (Figure 5A and Table 3) and PI3K/Akt (Figure 5B and Table 3) compared with control conditions (up to 28.9- and 1.5-fold increase in normal and OA cartilage, respectively; P ≤ 0.001) and with an activation of the specific transcription factors Ets-like gene-1 (Elk-1) (Figure 5C and Table 3) and SOX9 (Figure 5D and Table 3) (up to 18.8- and 1.6-fold increase in normal and OA cartilage, respectively; P ≤ 0.001).

Expression of phospho-ERK-1/2, phospho-Akt, phospho-Elk-1 and SOX9 in rAAV-transduced human articular cartilage. Primary human normal and OA cartilage explant cultures were transduced with either rAAV-hIGF-I or the control rAAV-lacZ vector, as described in Figure 3, and processed after 90 d to detect the expression of phospho-ERK-1/2 (A), phospho-Akt (B), phospho-Elk-1 (C) and SOX9 (D). Magnification 20×. All are views of the middle zone.
Discussion
Application of sequence coding for factors that might successfully and durably influence the metabolic and proliferative pathways in OA articular cartilage is an attractive approach to readjust the disturbed balance in this disorder and to counteract its long-term progression. Among the candidates tested for their reparative properties, IGF-I was described for its ability to modulate cell proliferation and extracellular matrix synthesis in horse and rabbit experimental models of cytokine-induced cartilage matrix degradation in situ (13,14,22). Yet, to the best of our knowledge, the effects of IGF-I gene transfer upon the long-term remodeling of human OA cartilage have not been evaluated to date. Notably, in these previous reports, delivery of IGF-I was performed using adenoviral vectors, a class of vectors that promote high but only very transient levels of gene expression, an important issue for the treatment of a disorder with a slow and irreversible progression such as OA (32). Vectors on the basis of the nonpathogenic adeno-associated virus might be thus better suited to treat this disease, since rAAVs are capable of transducing OA cells with high efficiency and for persistent periods of time and also when the cells are still surrounded by extracellular matrix components in situ (20,24,27,31). In the present study, we therefore examined the effects of rAAV-mediated overexpression of IGF-I on the long-term reconstruction of human OA cartilage and on the expression of the major components of its own axis.
The data first indicate that highly effective, significant and prolonged expression of IGF-I could be achieved via rAAV in primary human normal and OA chondrocytes in monolayer and alginate cultures in vitro (maintenance for up to 20 d and 26 d, respectively) and most remarkably in cartilage explants in situ (regular increase until d 90). Transduction efficiencies reached up to 95%, even when using relatively low MOI and vector doses (MOI = 6–20 in vitro; 4 × 108 functional recombinant particles in situ), probably because of the high permissivity of the cells to rAAV transduction and to their ability to rapidly process the viral particles compared with fibroblasts, which require much higher amounts of rAAV (MOI of at least 2,000) to allow for an efficient viral trafficking to the nucleus and to detectable transgene expression, in good agreement with previous findings (20,27,31,45,60–62). Interestingly, transgene expression was seen throughout the thickness of the cartilage, probably because of the ability of the small rAAV particles to penetrate the dense cartilage matrix and in agreement with previous findings using this class of vector (20,27,45). In monolayer cultures, high concentrations of IGF-I were produced in transduced normal and OA cells (at least 18.87 ± 0.92 ng/107 cells/24 h on d 5), remaining steady until d 20 (above 16.68 ± 0.64 ng/107 cells/24 h; P ≥ 0.200). The levels produced early on via rAAVs were lower than those achieved at similar time points with nonviral vectors (up to 83 ng/107 cells/24 h with 2 µg DNA [that is, 2.8- to 4.4-fold higher than here using a relatively low MOI of 20]) (52,63) or with plasmid vectors carrying AAV sequences (up to 900 ng/107 cells/24 h with 2 µg DNA [that is, 30.7- to 47.7-fold more than with our recombinant particles]) (64). Nevertheless, such levels decreased rapidly (230 ng/107 cells/24 h on d 6) (64) and were probably undetectable by d 20, as is commonly observed with nonviral compounds (65). A rapid decline in IGF-I expression was also reported when using adenoviral vectors (22,66,67). On d 3, IGF-I production levels were of 200–600 ng/107 cells/24 h at high adenoviral vector doses (MOI = 100–500 [that is, 5- to 25-fold higher than here]), which relates well with the concentrations achieved here with an MOI of 20 (that is, 6.8- to 31.8-fold difference), followed by a dramatic decrease on d 20 (67 ng/107 cells/24 h at an MOI of 500 [that is, 25-fold more adenoviral vector than here, for a difference of only 3.1- to 4.1-fold in concentration]) (67). Next, we measured long-term IGF-I secretion in encapsulated cells at vector doses that were even lower than those applied in the monolayer cultures (from 25.02 ± 0.22 to 24.60 ± 0.16 and from 24.73 ± 0.09 to 22.79 ± 0.13 ng/107 cells/24 h between d 5 and d 26 in normal and OA cells, respectively, at an MOI of 6; P ≥ 0.165). This result was also reported when using nonviral vectors, but again with high amounts of genetic material (28.4 ng/107 cells/24 h on d 36 with 2 µg DNA) (68). Finally (and for the first time to the best of our knowledge), we demonstrate that rAAV allows for a significant (P ≤ 0.001 compared with each respective control) and increasing production of IGF-I in human normal and OA cartilage over a sustained period of time that might be relevant for successful treatment OA (from 16.97 ± 1.22 to 32.41 ± 1.84 and from 14.14 ± 1.02 to 37.20 ± 2.12 pg/mg dry weight/24 h between d 5 and d 90, respectively; P ≤ 0.001).
Most remarkably, the levels of IGF-I achieved here via rAAV stimulated the proliferative, survival and biosynthetic activities of human normal and OA chondrocytes in vitro and in situ vis-à-vis control treatment, even at prolonged time points, with well-maintained cell proliferation and enhanced matrix synthesis in vitro over time, consistent with the properties of the growth factor (64,69–71). The levels of activation attained in normal and OA cells in vitro and in situ after rAAV-hIGF-I treatment vis-à-vis control treatment were superior to those reported when using nonviral (52,64) or adenoviral vectors (13,14,22,66,67). Whereas increases of up to 2.9-fold (proliferation) and 3.2-fold (anabolism) were evidenced with these other classes of vectors, we noted enhanced responses of up to 38.8- and 8.7-fold, respectively, with rAAV compared with the controls, probably due to the elevated, well-maintained (in vitro) or increasing (in situ) levels of IGF-I expression that may reflect the persistence of the rAAV transgenes in their targets (32). Interestingly, application of the candidate rAAV-hIGF-I vector had no detectable effect on the expression of type I collagen, an observation in good agreement with previous findings when applying an IGF-I gene sequence to chondrocytes via adenoviral gene transfer (22,67). Quite strikingly, OA chondrocytes were particularly well responsive to the treatment by rAAV IGF-I here, in marked contrast with their otherwise established hyporesponsiveness to the growth factor (59,72). Of note, we also observed that IGF-I overexpression in OA cartilage in situ was capable of significantly decreasing the expression of IGFBP3 and IGFBP4, two inhibitors of the IGF-I actions (40–42), while increasing the levels of two of its potentiators (IGFBP5 and its own receptor [IGF-IR]) (42,43), an effect noted at prolonged time points and probably the result of the steady levels of IGF-I production from the rAAV transgene sequence. In this regard, the effects of IGF-I on these components of the IGF-I axis via rAAV in OA cartilage are in good agreement with previous findings when providing IGF-I as a recombinant molecule to articular chondrocytes in vitro (73–76). Also noteworthy, the elevated, sustained levels of IGF-I produced in OA cartilage via rAAV might have been capable of counteracting the well-known effects of pathogenic adipokines on the destruction of articular cartilage (3,4). Of further note, the activating effects of IGF-I via rAAV on the expression of its receptor in normal and OA cartilage were associated with significant increases in the expression of effectors of the IGF-IR signal transduction pathways (MAPK/ERK-1/2 and PI3K/Akt) and downstream transcription factors (Elk-1, SOX9) described for their impact on cell proliferation and survival and on the synthesis of extracellular cartilage matrix components (proteoglycans, type II collagen) (38,39,77–79). These results are in good agreement with previous findings using recombinant IGF-I (35–37).
Importantly, we also report that overexpression of IGF-I in human OA cartilage led to higher levels of proliferation and biosynthesis than noted in control normal cartilage. It remains to be seen whether elevated levels of activities will influence the cartilage structure over time. Smith et al. (22) and Brower-Toland et al. (67) reported that production of IGF-I via adenoviral vectors led to decreased amounts of matrix synthesis in chondrocytes at high vector doses (MOI = 200–500). Equally important, sustained overexpression of IGF-I via gene transfer may have undesirable effects on bone formation by stimulating the replication and metabolic activities of osteoblasts that might be targeted after growth factor release in an in vivo setting (72,80,81). Thus, regulation of IGF-I expression may be critical when developing an appropriate treatment for OA that does not further alter the cartilage and bone integrity. Instead of CMV-IE, expression might be controlled by regulated (tetracycline-sensitive), cartilage tissue-specific (SOX9, type II collagen) or disease-induced transcription elements (82,83).
Despite significant effects of IGF-I, application of the candidate rAAV vector did not fully restore the native architecture in OA cartilage, and additional treatments might be necessary to refine the processes of regeneration. Combined delivery with other factors, such as those that prevent cartilage degradation (IL-1Ra, sTNFR, IL-10) (12–14,21) or correct the OA phenotype in the cells (SOX9) (20,26,27), might be useful to further attenuate pathological events in OA. Again, rAAVs are strong tools, since they can be used for cotransduction strategies in OA chondrocytes and cartilage (20). Yet, recovery from cartilage degeneration by direct gene transfer will be practicable only with some remaining cartilage surface and cells (early OA), whereas transplantation approaches will be desirable for the advanced cases of the disease.
Conclusion
In summary, the present results demonstrate the ability of rAAV-mediated IGF-I overexpression to stimulate cellular activities in human OA chondrocytes in vitro and in situ over extended periods of time, making rAAV an advantageous vector to develop long-term effective therapies against human OA. Additional future studies will be required to test the effects of the construct in vivo, taking care of selecting an experimental model of OA that is the most adequate to reproduce the clinical situation among the many systems available to date (8,12,21,84) and of first reproducing the current findings using human cells by translation to such animal cells. The present findings provide motivation to further elaborate the current approach for human OA.
Disclosure
The authors declare that they have no competing interests as defined by Molecular Medicine, or other interests that might be perceived to influence the results and discussion reported in this paper.
References
Poole AR. (1999) An introduction to the pathophysiology of osteoarthritis. Front. Biosci. 4:D662–70.
Pelletier JP, DiBattista JA, Roughley P, McCollum R, Martel-Pelletier J. (1993) Cytokines and inflammation in cartilage degradation. Rheum. Dis. Clin. North Am. 19:545–68.
Conde J, et al. (2011) Adipokines and osteoarthritis: novel molecules involved in the pathogenesis and progression of disease. Arthritis. 2011:203901.
Goldring MB, Otero M. (2011) Inflammation in osteoarthritis. Curr. Opin. Rheumatol. 23:471–8.
Dumond H, et al. (2003) Evidence for a key role of leptin in osteoarthritis. Arthritis Rheum. 48:3118–29.
Baragi VM, et al. (1995) Transplantation of transduced chondrocytes protects articular cartilage from interleukin 1-induced extracellular matrix degradation. J. Clin. Invest. 96:2454–60.
Pelletier JP, et al. (1997) In vivo suppression of early experimental osteoarthritis by interleukin-1 receptor antagonist using gene therapy. Arthritis Rheum. 40:1012–9.
Fernandes J, et al. (1999) In vivo transfer of interleukin-1 receptor antagonist gene in osteoarthritic rabbit knee joints: prevention of osteoarthritis progression. Am. J. Pathol. 154:1159–69.
Frisbie DD, McIlwraith CW. (2000) Evaluation of gene therapy as a treatment for equine traumatic arthritis and osteoarthritis. Clin. Orthop. Relat. Res. 379 (Suppl.):S273–87.
Frisbie DD, Ghivizzani SC, Robbins PD, Evans CH, McIlwraith CW. (2002) Treatment of experimental equine osteoarthritis by in vivo delivery of the equine interleukin-1 receptor antagonist gene. Gene Ther. 9:12–20.
Kafienah W, Al-Fayez F, Hollander AP, Barker MD. (2003) Inhibition of cartilage degradation: a combined tissue engineering and gene therapy approach. Arthritis Rheum. 48:709–18.
Zhang X, Mao Z, Yu C. (2004) Suppression of early experimental osteoarthritis by gene transfer of interleukin-1 receptor antagonist and interleukin-10. J. Orthop. Res. 22:742–50.
Haupt JL, et al. (2005) Dual transduction of insulin-like growth factor-I and interleukin-1 receptor antagonist protein controls cartilage degradation in an osteoarthritic culture model. J. Orthop. Res. 23:118–26.
Nixon AJ, et al. (2005) Gene-mediated restoration of cartilage matrix by combination insulin-like growth factor-I/interleukin-1 receptor antagonist therapy. Gene Ther. 12:177–86.
Grossin L, et al. (2006) Gene transfer with HSP 70 in rat chondrocytes confers cytoprotection in vitro and during experimental osteoarthritis. FASEB J. 20:65–75.
Gouze JN, et al. (2004) Adenovirus-mediated gene transfer of glutamine: fructose-6-phosphate amidotransferase antagonizes the effects of interleukin-1beta on rat chondrocytes. Osteoarthritis Cartilage. 12:217–24.
Hsieh JL, et al. (2010) Intraarticular gene transfer of thrombospondin-1 suppresses the disease progression of experimental osteoarthritis. J. Orthop. Res. 28:1300–6.
Hsieh JL, et al. (2009) Adenovirus-mediated kallistatin gene transfer ameliorates disease progression in a rat model of osteoarthritis induced by anterior cruciate ligament transection. Hum. Gene Ther. 20:147–58.
Chen LX, et al. (2008) Suppression of early experimental osteoarthritis by in vivo delivery of the adenoviral vector-mediated NF-kappaBp65-specific siRNA. Osteoarthritis Cartilage. 16 174–84.
Cucchiarini M, Terwilliger EF, Kohn D, Madry H. (2009) Remodelling of human osteoarthritic cartilage by FGF-2, alone or combined with Sox9 via rAAV gene transfer. J. Cell. Mol. Med. 13:2476–88.
Chen B, Qin J, Wang H, Magdalou J, Chen L. (2010) Effects of adenovirus-mediated bFGF, IL-1Ra and IGF-1 gene transfer on human osteoarthritic chondrocytes and osteoarthritis in rabbits. Exp. Mol. Med. 42:684–95.
Smith P, et al. (2000) Genetic enhancement of matrix synthesis by articular chondrocytes: comparison of different growth factor genes in the presence and absence of interleukin-1. Arthritis Rheum. 43:1156–64.
Matsumoto T, et al. (2009) Cartilage repair in a rat model of osteoarthritis through intraarticular transplantation of muscle-derived stem cells expressing bone morphogenetic protein 4 and soluble Flt-1. Arthritis Rheum. 60:1390–405.
Ulrich-Vinther M, Stengaard C, Schwarz EM, Goldring MB, Soballe K. (2005) Adeno-associated vector mediated gene transfer of transforming growth factor-beta1 to normal and osteoarthritic human chondrocytes stimulates cartilage anabolism. Eur. Cell. Mater. 10:40–50.
Blaney Davidson EN, Vitters EL, van den Berg WB, van der Kraan PM. (2006) TGF beta-induced cartilage repair is maintained but fibrosis is blocked in the presence of Smad7. Arthritis Res. Ther. 8:R65–72.
Tew SR, et al. (2005) Retroviral transduction with SOX9 enhances re-expression of the chondrocyte phenotype in passaged osteoarthritic human articular chondrocytes. Osteoarthritis Cartilage. 13:80–9.
Cucchiarini M, et al. (2007) Restoration of the extracellular matrix in human osteoarthritic articular cartilage by overexpression of the transcription factor SOX9. Arthritis Rheum. 56:158–67.
Venkatesan N, et al. (2004) Stimulation of proteoglycan synthesis by glucuronosyltransferase-I gene delivery: a strategy to promote cartilage repair. Proc. Natl. Acad. Sci. U. S. A. 101:18087–92.
Surendran S, et al. (2006) Anti-apoptotic Bcl-2 gene transfection of human articular chondrocytes protects against nitric oxide-induced apoptosis. J. Bone Joint Surg. Br. 88:1660–5.
Piera-Velazquez S, Jimenez SA, Stokes D. (2002) Increased life span of human osteoarthritic chondrocytes by exogenous expression of telomerase. Arthritis Rheum. 46:683–93.
Cucchiarini M, et al. (2005) Improved tissue repair in articular cartilage defects in vivo by rAAV-mediated overexpression of human fibroblast growth factor 2. Mol. Ther. 12:229–38.
Evans CH, Gouze JN, Gouze E, Robbins PD, Ghivizzani SC. (2004) Osteoarthritis gene therapy. Gene Ther. 11 379–89.
Boileau C, et al. (2006) PD-0200347, an alpha2delta ligand of the voltage gated calcium channel, inhibits in vivo activation of the Erk1/2 pathway in osteoarthritic chondrocytes: a PKCalpha dependent effect. Ann. Rheum. Dis. 65:573–80.
Pelletier JP, et al. (2003) In vivo selective inhibition of mitogen-activated protein kinase kinase 1/2 in rabbit experimental osteoarthritis is associated with a reduction in the development of structural changes. Arthritis Rheum. 48:1582–93.
Starkman BG, Cravero JD, Delcarlo M, Loeser RF. (2005) IGF-I stimulation of proteoglycan synthesis by chondrocytes requires activation of the PI 3-kinase pathway but not ERK MAPK. Biochem. J. 389:723–9.
Yin W, Park JI, Loeser RF. (2009) Oxidative stress inhibits insulin-like growth factor-I induction of chondrocyte proteoglycan synthesis through differential regulation of phosphatidylinositol 3-kinase-Akt and MEK-ERK MAPK signaling pathways. J. Biol. Chem. 284:31972–81.
Shakibaei M, Seifarth C, John T, Rahmanzadeh M, Mobasheri A. (2006) Igf-I extends the chondrogenic potential of human articular chondrocytes in vitro: molecular association between Sox9 and Erk1/2. Biochem. Pharmacol. 72:1382–95.
Loeser RF, Erickson EA, Long DL. (2008) Mitogen-activated protein kinases as therapeutic targets in osteoarthritis. Curr. Opin. Rheumatol. 20:581–6.
Beier F, Loeser RF. (2010) Biology and pathology of Rho GTPase, PI-3 kinase-Akt, and MAP kinase signaling pathways in chondrocytes. J. Cell. Biochem. 110:573–80.
Martin JA, Ellerbroek SM, Buckwalter JA. (1997) Age-related decline in chondrocyte response to insulin-like growth factor-I: the role of growth factor binding proteins. J. Orthop. Res. 15:491–8.
Neidel J, et al. (1997) Elevated levels of insulinlike growth factor (IGF) binding protein-3 in rheumatoid arthritis synovial fluid inhibit stimulation by IGF-I of articular chondrocyte proteoglycan synthesis. Rheumatol. Int. 17:29–37.
Kiepe D, et al. (2001) Intact IGF-binding protein-4 and -5 and their respective fragments isolated from chronic renal failure serum differentially modulate IGF-I actions in cultured growth plate chondrocytes. J. Am. Soc. Nephrol. 12:2400–10.
Kiepe D, Ciarmatori S, Haarmann A, Tonshoff B. (2006) Differential expression of IGF system components in proliferating vs. differentiating growth plate chondrocytes: the functional role of IGFBP-5. Am. J. Physiol. Endocrinol. Metab. 290:E363–71.
Mankin HJ, Dorfman H, Lippiello L, Zarins A. (1971) Biochemical and metabolic abnormalities in articular cartilage from osteo-arthritic human hips. II. Correlation of morphology with biochemical and metabolic data. J. Bone Joint Surg. Am. 53:523–37.
Madry H, Cucchiarini M, Terwilliger EF, Trippel SB. (2003) Recombinant adeno-associated virus vectors efficiently and persistently transduce chondrocytes in normal and osteoarthritic human articular cartilage. Hum. Gene Ther. 14:393–402.
Madry, et al. (2003) Recombinant adeno-associated virus vectors efficiently transduce ligaments and tendons in vivo. In: 49th Annual Meeting of the Orthopaedic Research Society; 2003 Feb 2–5; New Orleans, LA. Poster nr 0918. Available from: http://www.ors.org/abstracts/
Samulski RJ, Chang LS, Shenk T. (1987) A recombinant plasmid from which an infectious adenoassociated virus genome can be excised in vitro and its use to study viral replication. J. Virol. 61:3096–101.
Samulski RJ, Chang LS, Shenk T. (1989) Helper-free stocks of recombinant adeno-associated viruses: normal integration does not require viral gene expression. J. Virol. 63:3822–8.
Cucchiarini M, Ren XL, Perides G, Terwilliger EF. (2003) Selective gene expression in brain microglia mediated via adeno-associated virus type 2 and type 5 vectors. Gene Ther. 10:657–67.
Cucchiarini M, Ekici M, Schetting S, Kohn D, Madry H. (2011) Metabolic activities and chondrogenic differentiation of human mesenchymal stem cells following rAAV-mediated gene transfer and overexpression of fibroblast growth factor 2. Tissue Eng. Part A. 17:1921–33.
Jansen M, et al. (1983) Sequence of cDNA encoding human insulin-like growth factor I precursor. Nature. 306:609–11.
Madry H, Zurakowski D, Trippel SB. (2001) Overexpression of human insulin-like growth factor-I promotes new tissue formation in an ex vivo model of articular chondrocyte transplantation. Gene Ther. 8:1443–9.
Aigner T, Bertling W, Stoss H, Weseloh G, von der Mark K. (1993) Independent expression of fibril-forming collagens I, II, and III in chondrocytes of human osteoarthritic cartilage. J. Clin. Invest. 91:829–37.
Olney RC, et al. (1996) Chondrocytes from osteoarthritic cartilage have increased expression of insulin-like growth factor I (IGF-I) and IGF-binding protein-3 (IGFBP-3) and -5, but not IGF-II or IGFBP-4. J. Clin. Endocrinol. Metab. 81:1096–103.
Eviatar T, Kauffman H, Maroudas A. (2003) Synthesis of insulin-like growth factor binding protein 3 in vitro in human articular cartilage cultures. Arthritis Rheum. 48:410–7.
Iwanaga H, et al. (2005) Enhanced expression of insulin-like growth factor-binding proteins in human osteoarthritic cartilage detected by immunohistochemistry and in situ hybridization. Osteoarthritis Cartilage. 13:439–48.
Hunziker EB, Kapfinger E, Martin J, Buckwalter J, Morales TI. (2008) Insulin-like growth factor (IGF)-binding protein-3 (IGFBP-3) is closely associated with the chondrocyte nucleus in human articular cartilage. Osteoarthritis Cartilage. 16:185–94.
Morales TI. (2008) The quantitative and functional relation between insulin-like growth factor-I (IGF) and IGF-binding proteins during human osteoarthritis. J. Orthop. Res. 26:465–74.
Dore S, et al. (1994) Human osteoarthritic chondrocytes possess an increased number of insulinlike growth factor 1 binding sites but are unresponsive to its stimulation: possible role of IGF-1-binding proteins. Arthritis Rheum. 37:253–63.
Hansen J, Qing K, Kwon HJ, Mah C, Srivastava A. (2000) Impaired intracellular trafficking of adeno-associated virus type 2 vectors limits efficient transduction of murine fibroblasts. J. Virol. 74:992–6.
Adriaansen J, et al. (2005) Enhanced gene transfer to arthritic joints using adeno-associated virus type 5: implications for intra-articular gene therapy. Ann. Rheum. Dis. 64:1677–84.
Boissier MC, et al. (2007) Synoviocyte infection with adeno-associated virus (AAV) is neutralized by human synovial fluid from arthritis patients and depends on AAV serotype. Hum. Gene Ther. 18:525–35.
Madry H, et al. (2002) Gene transfer of a human insulin-like growth factor I cDNA enhances tissue engineering of cartilage. Hum. Gene Ther. 13:1621–30.
Shi S, Mercer S, Trippel SB. (2010) Effect of transfection strategy on growth factor overexpression by articular chondrocytes. J.Orthop. Res. 28:103–9.
Madry H, Trippel SB. (2000) Efficient lipid-mediated gene transfer to articular chondrocytes. Gene Ther. 7:286–91.
Mi Z, et al. (2000) Adenovirus-mediated gene transfer of insulin-like growth factor 1 stimulates proteoglycan synthesis in rabbit joints. Arthritis Rheum. 43:2563–70.
Brower-Toland BD, et al. (2001) Direct adenovirus-mediated insulin-like growth factor I gene transfer enhances transplant chondrocyte function. Hum. Gene Ther. 12:117–29.
Madry H, et al. (2005) Enhanced repair of articular cartilage defects in vivo by transplanted chondrocytes overexpressing insulin-like growth factor I (IGF-I). Gene Ther. 12:1171–9.
Osborn KD, Trippel SB, Mankin HJ. (1989) Growth factor stimulation of adult articular cartilage. J. Orthop. Res. 7:35–42.
Goldring MB, Goldring SR. (1990) Skeletal tissue response to cytokines. Clin. Orthop. Relat. Res. 258:245–78.
Trippel SB. (1995) Growth factor actions on articular cartilage. J. Rheumatol. Suppl. 43:129–32.
Martel-Pelletier J, Di Battista JA, Lajeunesse D, Pelletier JP. (1998) IGF/IGFBP axis in cartilage and bone in osteoarthritis pathogenesis. Inflamm. Res. 47:90–100.
Matsumoto T, Gargosky SE, Oh Y, Rosenfeld RG. (1996) Transcriptional and post-translational regulation of insulin-like growth factor-binding protein-5 in rat articular chondrocytes. J. Endocrinol. 148:355–69.
Sunic D, McNeil JD, Rayner TE, Andress DL, Belford DA. (1998) Regulation of insulin-like growth factor-binding protein-5 by insulin-like growth factor I and interleukin-1alpha in ovine articular chondrocytes. Endocrinology. 139:2356–62.
Porter RM, Akers RM, Howard RD, Forsten-Williams K. (2007) Alginate encapsulation impacts the insulin-like growth factor-I system of monolayer-expanded equine articular chondrocytes and cell response to interleukin-1beta. Tissue Eng. 13:1333–45.
Yoon DM, Fisher JP. (2008) Effects of exogenous IGF-1 delivery on the early expression of IGF-1 signaling molecules by alginate embedded chondrocytes. Tissue Eng. Part A. 14:1263–73.
Bell DM, et al. (1997) SOX9 directly regulates the type-II collagen gene. Nat. Genet. 16:174–8.
Lefebvre V, Huang W, Harley VR, Goodfellow PN, de Crombrugghe B. (1997) SOX9 is a potent activator of the chondrocyte-specific enhancer of the pro alpha1(II) collagen gene. Mol. Cell. Biol. 17:2336–46.
Vincent AM, Feldman EL. (2002) Control of cell survival by IGF signaling pathways. Growth Horm. IGF Res. 12:193–7.
Canalis E. (1980) Effect of insulinlike growth factor I on DNA and protein synthesis in cultured rat calvaria. J. Clin. Invest. 66:709–19.
Massicotte F, et al. (2006) Abnormal insulin-like growth factor 1 signaling in human osteoarthritic subchondral bone osteoblasts. Arthritis Res. Ther. 8:R177–88.
Goldring MB, Fukuo K, Birkhead JR, Dudek E, Sandell LJ. (1994) Transcriptional suppression by interleukin-1 and interferon-gamma of type II collagen gene expression in human chondrocytes. J. Cell. Biochem. 54:85–99.
Apparailly F, et al. (2002) Tetracycline-inducible interleukin-10 gene transfer mediated by an adeno-associated virus: application to experimental arthritis. Hum. Gene Ther. 13:1179–88.
Aigner T, et al. (2010) Histopathology atlas of animal model systems: overview of guiding principles. Osteoarthritis Cartilage. 18 (Suppl. 3):S2–6.
Acknowledgments
We thank RJ Samulski (The Gene Therapy Center, University of North Carolina, Chapel Hill, NC, USA) and X Xiao (The Gene Therapy Center, University of Pittsburgh, Pittsburgh, PA, USA) for providing genomic AAV-2 plasmid clones and the 293 cell line. We also thank AJ D’Ercole and B Moats-Staats (Department of Pediatrics, University of North Carolina, Chapel Hill) for the human IGF-I cDNA.
This work was supported by grants from the German Research Society (Deutsche Forschungsgemeinschaft; grants DFG CU 55/1-1 and CU 55/1-2) and from the German Osteoarthritis Foundation (Deutsche Arthrose-Hilfe; grants DAH P65-A234-Kohn-EP3-cucc3-sonder-ko-38k-2004-07 and P66-A234-Kohn-EP4-cucc4-sonder-ko-30k-2006-08).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, and provide a link to the Creative Commons license. You do not have permission under this license to share adapted material derived from this article or parts of it.
The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this license, visit (http://creativecommons.org/licenses/by-nc-nd/4.0/)
About this article
Cite this article
Weimer, A., Madry, H., Venkatesan, J.K. et al. Benefits of Recombinant Adeno-Associated Virus (rAAV)-Mediated Insulinlike Growth Factor I (IGF-I) Overexpression for the Long-Term Reconstruction of Human Osteoarthritic Cartilage by Modulation of the IGF-I Axis. Mol Med 18, 346–358 (2012). https://doi.org/10.2119/molmed.2011.00371
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2119/molmed.2011.00371