
Abstract
Background
Most of long-distance migratory raptors suspend moult during migration but detailed information is patchy for most of the Palearctic species. The aim of this research is to verify if active moulting in migrating Western Marsh Harriers occurs and to quantify the extent of moulting along the season focusing on primary feathers.
Methods
During a whole post-breeding migration at the Strait of Messina in Southern Italy, we gathered information about symmetrical flight feather moult from 221 adults by taking pictures of raptors passing at close range.
Results
We found active moulting primaries during autumn migration in 48.4% of our samples. Slight differences on the extension and timing among sex classes were recorded during the season, with adult females showing a more advanced moult stage than adult males.
Conclusion
The finding that the extension of the suspended moult was already defined in migratory individuals might be explained as an adaptation to minimize the energy required for moulting during migration.
Similar content being viewed by others

Background
Breeding, migration, and moulting are considered the three-main energy-demanding events in the yearly cycle of birds (Newton 2008) and usually studied separately due to their isolated occurrence (Newton 2009). The importance of the moult in the life cycle of birds of prey for advancing our understanding of the ecology of each species has been recently highlighted by Zuberogoitia et al. (2018). In several migrating species moulting leads to age-dependent migration strategies (Cristol et al. 1999; Zenzal and Moore 2016) and these differences are especially pronounced among long-distance migrants. Among passerines, first-year birds migrate earlier than adults. The opposite occurs in species in which adults suspend or postpone moult until they reach their wintering grounds (Schifferli 1965; Payevski 1994; Woodrey and Chandler 1997; Woodrey and Moore 1997; Jakubas and Wojczulanis-Jakubas 2010). Long distance migrant raptors typically avoid moulting during migration (Berthold 1975; Newton 1979) such as in the case of Osprey (Pandion haliaaetus; Ferguson-Lees and Christie 2001). In small raptors the time required to grow a new feather is 2‒3 weeks while it is much longer in large raptors such as vulture species (Zuberogoitia et al. 2018). In the case of damaged feathers, raptors can replace them much faster than in the usual time of moulting (Ellis et al. 2016). However, there are species that do not stop moulting during long-distance movements (Kjellén 1992). For example, it has been found that Palearctic Circus sp. moult during migration and eventually slow the moulting process (Arroyo and King 1996). Moreover, we can find the “suspended moult” strategy which is defined as a moult interrupted temporarily and subsequently resumed (Baker 2016). This strategy frequently involves individuals in late breeding attempts having to interrupt their normal moult cycle in order to migrate.
In Western Marsh Harriers (WMHs), females started moulting during incubation (May‒June) while males start up to 6 weeks later (Forsman 1999). During chicks rearing period WMH may suspend moult for better hunting efficiency while they end moulting in late October‒early November (Forsman 1999). During spring migration Gorney and Yom (2001) showed that adult WMHs perform their journey with non-active moult in their flight feathers.
Unfortunately, moulting patterns in relation to migration strategies are poorly investigated in raptor species despite it is well known that long-distance migrant Accipitriformes need a large amount of energy to overcome barriers during migration where they must use flapping powered flight (Agostini et al. 2015). The aim of this work is to confirm and to describe the active moult of WMH’s flight feathers during the spring migration across the Strait of Messina.
Methods
Fieldwork
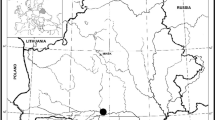
From August 13th to September 30th 2016 we collected data on moulting Western Marsh Harriers migrating along the Mid-Mediterranean Flyway (Fig. 1). Fieldwork was carried out on the continental side of the Strait of Messina, a hotspot for soaring birds (Panuccio et al. 2005; Agostini et al. 2017; Pastorino et al. 2017). We used two watch points both located on the Aspromonte Plains, a flat highland a few kilometers inland of the Strait of Messina, on the western side of the Apennines ridge. The highland lies between 1000 and 1200 m a.s.l. with a landscape altered by human agricultural activity. One of the watch points was located in the middle of the highland while the other one was located close the mountain slopes. Observations were carried out daily from sunrise to sunset.

Geographical context of the study area, the Strait of Messina (SM). The asterisk indicates the Aspromonte plain
We used high-quality photos taken in the field from which moult patterns can be inferred (Snyder et al. 1987; Zuberogoitia et al. 2016; Vieira et al. 2017). Digital cameras (i.e. Panasonic Lumix DMC-FZ72, FZ300) were equipped with lens with 8‒60 × magnification. Photography data collection was supported by visual observations that were made using binoculars (i.e. Leica 10 × 42) and scopes (i.e. Swarowski 20‒60 ×) by at least two experienced birders operating simultaneously. Visual observations were used only to detect if birds were moulting or not. Pictures and data were compared daily with those from the other watch points to delete possible double-counting of the same individuals according to the time and location of the birds passage as well as the individual moult features and sex. Only birds observed or photographed at closer range (< 100 m) were considered in the analysis, providing a random sample of the whole number of WMHs detected. We determined: (1) if harriers were moulting or not; (2) for a subsample of individuals, we were able to describe the moulting stage according to Ginn and Melville (1983). In this last subset of data, we recorded the number of primaries that the bird was moulting, if any. Therefore, we assigned a value from 1 to 10 according to the primary feathers moulting pattern of the species, being 1 = P1, 2 = P2 and so on (Baker 2016).
Data analysis
We used two different analyses. A Binary Logistic Regression Analysis (hereafter BLRA) (logit link function) (Boyce et al. 2002; Keating and Cherry 2004; Rushton et al. 2004) was used to test the factors influencing numbers of moulting individuals. For this we compared moulting and non-moulting birds (using both, data from visual observations and from pictures inspection), that in the BLRA was our dependent binary variable (1/0). The covariates were:
-
1.
The Julian date calculated as the number of the day from the beginning of the year (January 1st being day number 1).
-
2.
The squared Julian date was used as covariate as well to account for non-linear effects of the Julian date on bird migration (Knudsen et al. 2007; Lindén 2011; Lindén and Mäntyniemi 2011; Panuccio et al. 2016).
-
3.
Sex of the bird (male, female).
-
4.
Age (2nd CY, adult).
-
5.
Watch point.
We tested the fitness of the model by means of the area under the curve of the Receiver Operating Characteristic (ROC) plot (Pearce and Ferrier 2000; Boyce et al. 2002; Fawcett 2006). This area provides a measure of discrimination ability, varying from 0.5 for a model with a discrimination ability no better than random, to 1.0 for a model with perfect discriminatory ability.
To verify the variation of moulting stage among the photographed individuals, we used a Linear Model (LM) using the number of moulting primaries as dependent variable (from 1 to 10). As covariates we used the same of the BLRA with the exclusion of the watchpoint. We tested the fitness of the model checking the R2 value.
For both, LM and BLRA, we made a stepwise model selection comparing the different models by the AICc value and choosing the ones with the lowest value. All statistical analyses were made with R open source software (R Development Core Team 2015).
Results
We observed a total of 3522 Western Marsh Harriers between the 13th August to 30th September. Among those, the individuals that were aged as adult birds were 1125 while juveniles were 413. From adult birds, 802 were positively identified as males and 323 as females, of which 19 were 2nd CY birds. Other 673 individuals were recorded under the category female/juvenile type. The remaining 1311 observed WMHs were undetermined. We took close pictures that allowed us to identify clear signs of moult from 107 individuals, while other 114 harriers did not show moult signs. From all these WMHs we obtained active moult schemes extracted from photos of 35 WMHs, 21 males and 14 females respectively. Another seven WMHs that recently suspended the moult of their primaries were confirmed but not included in the analyses because they weren’t active moulting birds. We did not record any individuals with old feathers missing or with new pin feathers, neither individuals with new feathers at one-third of its whole length. The timing of the primary feathers moult varies slightly between males and females (Fig. 2). Females showed a more advanced moult stage on average than males, with 100% of the females with the inner 3rd primary already moulted unlike the 79% of the males. Similarly, different proportions between sexes have been recorded for the 4th, 5th, 6th and 7th primaries. However, the three outermost primaries, the 8th, the 9th and the 10th, remained unmoulted in both sexes (Fig. 3).

Proportions of moulting WMH during autumn migration at the Strait of Messina. Moulting (Yes = light grey) and non-moulting (No = dark grey) birds across the season (11th August‒30th September). (a) Males, (b) Females

Proportion of moulted primaries for male and female WMHs recorded in the present study (SS = secondary feathers)
The results of both, the BLRA and the LM (Table 1), indicate that the Julian date is the only relevant variable explaining the passage of moulting harriers in the study area, with higher numbers observed early in the season and with a more advanced moult later in the season (Table 1). Females were observed with more moulted primaries than males (Fig. 4). Moreover, 2nd CY harriers have a more advanced moult comparing to adults (Table 1).

Moult of male (a) and female (b) Western Marsh Harriers in relation to Julian date
Discussion
Our results suggest that adult WMHs start their autumn migration once the last primary to be moulted (maximum until the P7) was already expelled, and the previous primaries are already growing. Growing stages of those feathers could be visible during the lapse of migration that it takes for feathers to grow until their complete development. Therefore, during post-breeding migration only growing new flight feathers and moulted new ones were recorded, but no lacking feathers. We detected the presence of primary feathers unmoulted, already moulted, and growing at medium/late moulting stages. Not a single individual showed the lack of a primary or an immediate growing stage such as 1 or 2 (Ginn and Melville 1983). This indicates that replacing the next old feathers in the sequence is unlikely to occur during migration but in the winter quarters.
The lack of feathers might imply a less efficient flight with a disproportional increase of the energy required for flapping (Pennycuick 2008). Therefore, it is possible that adult WMHs which belong to the same sex group with a higher number of primary feathers involved in their moult should come from breeding grounds farther away than individuals showing a less extended ongoing moult. Moreover, our data suggests that there is a relationship between the extension and timing of the suspended moult and the timing of the migration itself with a difference between males and females. We found adult females showed an averaged more extended moult than the adult males and a later timing of migration as well (Agostini and Panuccio 2010; Agostini et al. 2017). This confirms that adult females start moulting in the breeding season before the males. However, they suspend that moult later on average as well (Cramp and Simmons 1980; Kjellén 1992).
Conclusion
If we considered “active moult” as the lapse of time during which a bird replaces feathers rather than the time spent by a bird on feathers growth, we might consider that active moult of WMHs ends at the breeding ground immediately after shedding the last primary to be replaced and just before starting the post-breeding migration. So, during migration, only growing feathers take place, being the extension of the suspended moult already defined at the breeding ground just before leaving. This strategy might be evolved to minimize the effect of moulting during migration. Such movements are energetically demanding and harriers are known to be raptors that largely use powered-flapping flight over long distances (Spaar and Bruderer 1997; Panuccio et al. 2013, 2016; Agostini et al. 2015, 2017).
References
Agostini N, Panuccio M. Western Marsh harrier (Circus aeruginosus) migration through the Mediterranean sea: a review. J Raptor Res. 2010;44:136–42.
Agostini N, Panuccio M, Pastorino A, Sapir N, Dell’Omo G. Migration of the Western Marsh Harrier to the African wintering quarters along the Central Mediterranean flyway: a 5-year study. Avian Res. 2017;8:24.
Agostini N, Panuccio M, Pasquaretta C. Morphology, flight performance, and water crossing tendencies of Afro-Palearctic raptors during migration. Curr Zool. 2015;61:951–8.
Arroyo BE, King JR. Age and sex differences in molt of the Montagu’s Harrier. J Raptor Res. 1996;30:224–33.
Baker K. Identification guide to European Non-Passerines. BTO Guide 24. 2016.
Berthold P. Migration: control and metabolic physiology. In: Farner DS, King JR, editors. Avian biology, vol. 5. New York: Academic; 1975. p. 77–128.
Boyce MS, Vernier PR, Nielsen SE, Schmiegelow FKA. Evaluating resource selection functions. Ecol Model. 2002;157:281–300.
Cramp S, Simmons KEL. The birds of the Western Palearctic, vol. II. Oxford: Oxford University Press; 1980.
Cristol DA, Baker MB, Carbone C. Differential migration revisited. In: Nolan V, Ketterson ED, Thompson CF, editors. Current ornithology, vol. 15. Boston: Springer; 1999.
Ellis DH, Rohwer VG, Rohwer S. Experimental evidence that a large raptor can detect and replace heavily damaged flight feathers long before their scheduled moult dates. Ibis. 2016;159:217–20.
Fawcett T. An introduction to ROC analysis. Pattern Recogn Lett. 2006;27:861–74.
Ferguson-Lees J, Christie D. Raptors of the world. London: Helm; 2001.
Forsman D. The raptors of Europe and the Middle East. London: T & A D Poyser; 1999.
Ginn HB, Melville DS. Moult in birds (BTO Guides No. 19). Norfolk, UK: British Trust for Ornithology; 1983.
Gorney E, Yom Tov Y. Molt during spring migration: a comparison of four species of raptors. J Field Ornithol. 2001;72:96–105.
Lindén A. Using first arrival dates to infer bird migration phenology. Boreal Environ Res. 2011;16:49–60.
Lindén A, Mäntyniemi S. Using the negative binomial distribution to model overdispersion in ecological count data. Ecology. 2011;92:1414–21.
Jakubas D, Wojczulanis-Jakubas K. Sex- and age-related differences in the timing and body condition of migrating Reed Warblers Acrocephalus scirpaceus and Sedge Warblers Acrocephalus schoenobaenus. Naturwissenschaften. 2010;97:505–11.
Keating KA, Cherry S. Use and interpretation of logistic regression in habitat-selection studies. J Wildl Manag. 2004;68:774–89.
Kjellén N. Differential timing of autumn migration between sex and age groups in raptors at Falsterbo, Sweden. Ornis Scand. 1992;23:420–34.
Knudsen E, Lindén A, Ergon T, Jonzén N, Vik JO, Knape J, Røer JE, Stenseth NC. Characterizing bird migration phenology using data from standardized monitoring at bird observatories. Clim Res. 2007;35:59–77.
Newton I. Population ecology of raptors. London: T & A D Poyser; 1979.
Newton I. Moult and plumage. Ringing Migr. 2009;24:220–6.
Newton I. Migration ecology of birds. London: Academic Press; 2008.
Panuccio M, Agostini N, Mellone U. Autumn migration strategies of honey buzzards, black kites, marsh and Montagu’s harriers over land and over water in the central Mediterranean. Avocetta. 2005;29:27–32.
Panuccio M, Chiatante G, Tarini D. Two different migration strategies in response to an ecological barrier: Western Marsh Harriers and juvenile European Honey Buzzards crossing the central-eastern Mediterranean in autumn. J Biol Res Thessalon. 2013;19:10–8.
Panuccio M, Barboutis C, Chiatante G, Evangelidis A, Agostini N. Pushed by increasing air temperature and tailwind speed: weather selectivity of raptors migrating across the Aegean Sea. Ornis Fenn. 2016;93:159–71.
Pastorino A, Ramirez J, Agostini N, Dell’Omo G, Panuccio M. Fog and rain lead migrating White storks Ciconia ciconia to perform reverse migration and to land. Avocetta. 2017;41:5–12.
Pennycuick CJ. Modelling the flying bird. Cambridge: Academic Press; 2008.
Payevski VA. Age and sex structure, mortality and spatial winter distribution of Siskins (Carduelis spinus) migrating through eastern Baltic area. Vogelwarte. 1994;37:190–8.
Pearce J, Ferrier S. Evaluating the predictive performance of habitat models developed using logistic regression. Ecol Model. 2000;133:225–45.
R Development Core Team. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing; 2015.
Rushton SP, Ormerod SJ, Kerby G. New paradigms for modelling species distributions? J Appl Ecol. 2004;41:193–200.
Schifferli A. Vom Zug verhalten in der Schweitz brutenden Turmfalken, Falco tinnunculus, nach den Ringfunden. Orn Beob. 1965;62:1–13.
Snyder NFR, Johnson EV, Clendenen DA. Primary molt of California Condors. Condor. 1987;89:468–85.
Spaar R, Bruderer B. Migration by flapping or soaring: flight strategies of marsh, Montagu’s and pallid harriers in southern Israel. Condor. 1997;99:458–69.
Vieira BP, Furness RW, Nager RG. Using field photography to study avian moult. Ibis. 2017;159:443–8.
Woodrey MS, Chandler CR. Age-related timing of migration: geographic and interspecific patterns. Wilson Bull. 1997;107:52–67.
Woodrey MS, Moore FR. Age-related differences in the stopover of fall landbird migrants on the coast of Alabama. Auk. 1997;114:695–707.
Zenzal TJ Jr, Moore FR. Stopover biology of ruby throated humming birds (Archilochus colubris) during autumn migration. Auk. 2016;133:237–50.
Zuberogoitia I, Gil JA, Martinez JE, Erni B, Aniz B, López-López P. The flight feather moult pattern of the bearded vulture (Gypaetus barbatus). J Ornithol. 2016;157:209–17.
Zuberogoitia I, Zabala J, Martínez JE. Moult in birds of prey: a review of current knowledge and future challenges for research. Ardeola. 2018;65:183–207.
Authors’ contributions
JR made the study design, collected data on moulting, prepared the dataset and wrote the first draft of the manuscript. MP coordinated the fieldwork, made the analysis and wrote the paper. Both authors read and approved the final manuscript.
Acknowledgements
We thank Giacomo Dell’Omo and Ornis italica for his support during the fieldwork. We are grateful to all the people who collected data at the Strait of Messina in autumn 2016 and in particular: Alberto Pastorino, Michele Cento, Gianpasquale Chiatante, Nicolantonio Agostini, Martina Zambon, Stiven Kocijančič, Giuseppe Cicero. We would also like to acknowledge Kylynn Clare for her contributions to the editing of this paper. We wish to thank MEDRAPTORS (www.raptormigration.org), a network of ornithologists involved in the research and the protection of migrating birds of prey through specific projects and observation camps.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets used in the present study are available from the corresponding author on request.
Consent for publication
Not applicable.
Ethical approval and consent to participate
This study did not imply manipulation of birds.
Funding
The fieldwork was supported by Ornis italica.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Ramírez, J., Panuccio, M. Flight feather moult in Western Marsh Harriers during autumn migration. Avian Res 10, 7 (2019). https://doi.org/10.1186/s40657-019-0146-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40657-019-0146-9