
Abstract
Background
Streptococcus iniae is an important fish pathogen that cause significant economic losses to the global aquaculture industry every year. Although there have some reports on the genotype of S.iniae and its relationship with virulence, no genome-scale comparative analysis has been performed so far. In our previous work, we characterized 17 isolates of S.iniae from Trachinotus ovatus and divided them into two genotypes using RAPD and rep-PCR methods. Among them, BH15-2 was classified as designated genotype A (in RAPD) and genotype 1 (in rep-PCR), while BH16-24 was classified as genotype B and genotype 2. Herein, we compared the differences in growth, drug resistance, virulence, and genome between BH15-2 and BH16-24.
Results
The results showed that the growth ability of BH16-24 was significantly faster than that of BH15-2 at the exponential stage. Antimicrobial tests revealed that BH15-2 was susceptible to most of the tested antibiotics except neomycin and gentamycin. In contrast, BH16-24 was resistant to 7 antibiotics including penicillin, sulfasomizole, compound sulfamethoxazole tablets, polymyxin B, spectinomycin, rifampin and ceftazidime. Intraperitoneal challenge of T.ovatus, showed that the LD50 value of BH15-2 was 4.0 × 102 CFU/g, while that of BH16-24 was 1.2 × 105 CFU/g. The genome of S.iniae BH15-2 was 2,175,659 bp with a GC content of 36.80%. Meanwhile, the genome of BH16-24 was 2,153,918 bp with a GC content of 36.83%. Comparative genome analysis indicated that compared with BH15-2, BH16-24 genome had a large-scale genomic inversion fragment, at the location from 502,513 bp to 1,788,813 bp, resulting in many of virulence and resistance genes differentially expression. In addition, there was a 46 kb length, intact phage sequence in BH15-2 genome, which was absent in BH16-24.
Conclusion
Comparative genomic studies of BH15-2 and BH16-24 showed that the main difference is a 1.28 Mbp inversion fragment. The inversion fragment may lead to abnormal expression of drug resistant and virulence genes, which is believed to be the main reason for the multiple resistance and weakened virulence of BH16-24. Our study revealed the potential mechanisms in underlying the differences of multidrug resistance and virulence among different genotypes of S.iniae.
Similar content being viewed by others


Background
Streptococcus iniae is a prominent warm-water pathogen and is known to infect a wide range of fish species. The infected fish exhibit varied clinical symptoms of erratic swimming, lethargy, meningoencephalitis, exophthalmia, enteritis and septicaemia [1, 2]. According to the previous reports, S.iniae can cause severe diseases in Trachinotus ovatus and result significant economic losses [3, 4]. Meanwhile, S.iniae is a zoonotic bacteria that poses a threat to public health [5].
Different from other streptococci of the same genus, the serotype of S.iniae cannot be classified by the traditional method of Lancefield [6]. Random amplified polymorphic DNA (RAPD) or repetitive primer polymerase chain reaction (rep-PCR) method can be used to distinguish the serotypes of S.iniae [7, 8]. Through analysis of rep-PCR and RAPD, 29 isolates of S.iniae were divided into two genotypes and a correlation between the genotype and the virulence had been identified [9]. Fuller et al. reported that S.iniae virulence is associated with distinct genetic profile and demonstrated differences between pathogenic and nonpathogenic isolates [10]. Rep-PCR analysis of 14 S.iniae strains from diseased O.niloticus presented genetic heterogeneity and were divided into six genotypes banding patterns [11]. The routine methods for preventing bacterial infections are the use of antibiotics and chemotherapeutants, but antibiotic usage in the aquaculture industry is largely uncontrolled. Abuse and/or excessive use of antibiotics can lead to the emergence of antimicrobial-resistant bacteria [12]. However, there is little information about the mechanisms behind antibiotic resistance and virulence differentiation among diverse S.iniae strains, with the lack of information hampering the effective treatment of the disease.
Comparative genomics analysis is a helpful way to identify genome-wide genetic variants of bacteria that may be associated with host and geographic origin but also to better understand their potential pathogenicity and antibiotic resistance. Comparative genomic analysis of S. agalactiae isolates with distinct clinical origins or host associations has provided insight into potential mechanisms of evolution, virulence, and host adaptation [13]. Fanelli et al. identified a large number of antibiotic, heavy metals and virulence determinants by reporting the whole genome sequencing and genomic characterization of two Arcobacter butzleri strains isolated from shellfish [14]. Similarly, the virulence and antimicrobial resistance-associated genomic determinants of two Salmonella Typhimurium strains were reported through comparative genomics approaches [15]. Although some complete genome sequences of S.iniae isolated have been sequenced, to our knowledge, there have been no reports on the comparative genomics analysis for understanding the genetic basis of pathogenicity and multidrug resistance. In our previous work, we isolated 17 strains of S.iniae from T.ovatus and differentiated them into two genotypes using RAPD and rep-PCR methods [16]. Among them, BH15-2 was divided as designated genotype A in RAPD analysis and genotype 1 in rep-PCR analysis, while BH16-24 was classified as genotype B and genotype 2. Here, we compared the phenotypes (growth, drug-resistance, and virulence variation) and genome of BH15-2 and BH16-24, which may explain the mechanism of their differences in biological properties.
Results
Clinical symptoms of infected fish and biochemical characterization of the strain
In this study, the natural infected fish were observed with symptoms of meningoencephalitis, enteritis, and hemorrhage of pterygiophore, operculum and liver (Fig. 1A-E). After 24 h incubation at 28 ℃ on BHI agar plates, the pathogenic bacteria formed circular, buff, smooth surface, intact edge, convex colonies with a diameter of approximately 0.5-1.0 mm (Fig. 1F). Gram staining showed that the strains are Gram-positive coccus appearing in short chains under an optical microscope. Biochemical analyses revealed complete homogeneity in BH15-2 and BH16-24. According to the growth curves, the growth rate of BH16-24 was significantly faster than that of BH15-2 after 6 h of culture (p < 0.05) (Fig. 2).

Clinical signs of diseased T.ovatus. (A) pterygiophore hemorrhage, (B) meningoencephalitis, (C) operculum hemorrhage (D) liver hemorrhage (E) enteritis (F) Bacterial colony on BHI agar

Growth curve of BH15-2 and BH16-24 strains cultured in BHI broth
Aliquots of cell culture were taken at various time points and measured for cell density at OD600, *indicates P < 0.05,**indicates P < 0.01
The results of virulence test
The infection results from BH15-2 and BH16-24 were shown in Table 1. Cumulative mortality rates of BH15-2 and BH16-24 after post infection were illustrated in Table 2. At 28 days post-infection (dpi), BH15-2 induced higher levels of mortality and revealed a greater virulence than BH16-24. Concretely, the LD50 value of BH15-2 was 4.0 × 102 CFU/g, while that of BH16-24 was 1.2 × 105 CFU/g (body weight). The virulence of BH15-2 was 1000 times than BH16-24. BH15-2 caused 75%, 75%, 87.5% and 100% cumulative mortality, while BH16-24 caused 6.25%, 6.25%, 12.5% and 62.5% at 1.5 × 105, 1.5 × 106, 1.5 × 107, 1.5 × 108 cfu/ml concentrations in T. ovatus respectively. In the control groups, there was no mortality during the observation period. Most deaths occurred within 48 h p.i. and did not show any clinical signs of disease prior to death. But bacterial isolated were retrieved from the brain and liver of the challenged fish and found to be the same species.
Drug resistance
The antibiogram study results are shown in Table 2. Both strains were sensitive to tetracycline, erythromycin, norfloxacin, azithromycin, ciprofloxacin, chloramphenicol, deoxytetracycline, and intermediate resistant toward to neomycin, gentamycin. BH16-24 was resistant or intermediate resistant toward all the β-lactam antibiotics tested in this study, i.e., penicillin, ampicilin, cefazolin, ceftriaxone Sodium, and ceftazidime, while BH15-2 was sensitive to these antibiotics. In addition, BH16-24 was resistant to sulfonamide antibiotics (sulfasomizole, compound sulfamethoxazole), polymyxin B, aminoglycoside antibiotic (spectinomycin), rifampin, whereas BH15-2 was susceptibility to these antibiotics. Therefore, BH16-24 is a multidrug-resistant (MDR) bacterium. The calculated MAR index of BH15-2 was 0, while BH16-24 was 0.37.
Comparative Genomics
General features of whole genome sequencing results
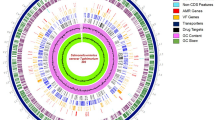
The general features of BH15-2 and BH16-24 genomes are summarized in Table 3. The genome size of BH15-2 was 2,175,659 bp with a GC content of 36.80%, while BH16-24 is 2,153,918 bp long with a GC content of 36.83%. Both genomes were optimized to assemble a circular genome with 0 gap. The genome of BH15-2 contained 2090 genes, 60 tRNAs and 15 rRNAs, while the genome of BH16-24 had 2039 genes 68 tRNAs and 18 rRNAs. Figure 3 shows the assembling results of BH15-2 and BH16-24 strains. The circular genomes of the two strains exhibited the coding sequence (CDS), repetitive sequences, GC content, number of RNA and GC skew (Fig. 3), where the outer 2 and 3 circles represented the CDS on the positive strand and negative stands. The genome sequences of BH15-2 and BH16-24 have been summited to the National Center for Biotechnology Information database with the accession number CP132229 and CP132230, respectively. Bioinformatics analysis indicated that BH15-2 and BH16-24 contains 71 and 21 unique genes, respectively, with a total of 2018 common genes.

Schematic circular diagrams of the S.iniae BH15-2 (A) and BH16-24 (B) genomes
Circle 1 (from outside to inside): scale (in kb); circles 2 and 3: genes in positive strand and negative strand; circle 4: repetitive sequences; circle 5: tRNA (in blue) and rRNA (in purple); circles 6: GC content; circles 7: GC skew
GIs analysis
The genomes of BH15-2 and BH16-24 were screened for horizontally acquired DNA using IslandPath-DIMOB, SIGI-HMM, IslandPick, and Islander methods integrated with the IslandViewer server (Fig. 4). In the genome of BH15-2, 17 presumed genome islands (GIs) ranging from 4912 to 50,177 bp were detected. The largest GI consisted of 50,177 bp with 79 predicted gene coding regions, of which 71 genes were unique in BH15-2 genome. And the shortest GI consisted of only 5 predicted gene. A total of 275 genes were predicted into GIs. For strain BH16-24, 21 presumed GIs ranging from 4716 to 38,833 bp were detected. The largest GI consisted of 38,833 bp and predicted to encode 45 genes. A total of 263 genes were predicted into GIs. It was found that most of the GIs of the two strains are the same by comparing the GIs of BH15-2 and BH16-24. However, based on the large inversion fragment between BH15-2 and BH16-24, the positions of several GIs are very different. The two strains each have a unique gene island, composed of unique genes (circled in red in Fig. 4). Both strains have no unique genes encoding virulence or drug resistance related genes.

Circular visualization of the predicted Genomic Islands (GIs) on BH15-2 (A) and BH16-24 (B) strains. The analysis was conducted in IslandViewer 4. The interactive visualization of the distinct islands across the genomes is shown with blocks colored according to the predictor tool as described: red represents the predicted by at least one method, blue represents that the results of IslandPath-DIMOB predicted, yellow represents the predicted results of SIGI-HMM
Prophage analysis
Prophage analysis of BH15-2 and BH16-24 showed that the two strains harbored 11 identical incomplete and questionable prophages (Supplementary Figure S1). But due to the inversion fragment, several of them are in different positions. Further, BH15-2 harbored an intact prophage and is also an active prophage predicted by Prophage Hunger (Fig. 5), which was absent in BH16-24. The intact prophage is 46 kb size located at 897,486 bp – 943,491 bp, contained a putative attL site and an attR site. And it encoded 60 prophage-related proteins and 17 hypothetical proteins, including phage tail protein, phage protein, siphovirus Gp157 family protein, ImmA/IrrE family metallo-endopeptidase, XRE family transcriptional regulator and endolysin.

Genomic organization of coding sequences (CDS) of prophage in BH15-2 (show annotated only)
Att: Attachment site; PLP: Phage-like protein; Ter: Terminase; Por: Portal protein; Sha: Tail shaft; Coa: Coat protein; Tra: Transposase
Large-scale genomic inversion fragment
The genomic synteny analysis of the whole S.iniae genome of BH15-2, BH16-24, and other three strains SF1, YSFST01-82 and ISET0901 which separately isolated from Oreochromis niloticus, and Paralichthys olivaceus was performed using Mauve software (The strain information is listed in Table 4). The result showed that all strains except BH16-24, were quite similar with respect to genome structure, with the exception of some small inversion. Thus, all strains except BH16-24 shared a similar synteny with each other. The BH16-24 genome displayed a large-scale inversion including 1.28 Mbp occurred across the origins/terminus axis (ori/ter axis), which located in the region from 502,513 bp-1,788,813 bp (Fig. 6).

Mauve comparison diagrams of the BH15-2, BH16-24, SF1, YSFST01-82 and ISET0901. The scale represents the coordinates of each genome. Different color blocks represent local collinear blocks (LCBs), which are conserved segments in five genomes. Within LCBs, the white area represents low similarity regions or regions unique to one genome but absent in another. LCBs above the black horizontal central line are in forwarding orientation and below this are in reverse orientation. Colored lines show the rearrangement of LCBs among the genomes
Antibiotic resistance genes in the inversion fragment
The large inversion fragment encodes 1242 genes. Among them, many antibiotic resistance genes including efflux system genes, ABC transporter and β-lactamase-encoding were screened out, such as penicillin-binding protein 2B (pbp2B), penicillin-binding protein 1 A (pbp1A), penicillin-binding protein 2X (pbp2X), aminoacyltransferase (femA), DNA gyrase subunit A (gyrA), oligopeptide-binding protein (oppA), aminoglycoside phosphotransferase (aph), dihydrofolate reductase (dfrA), macrolide export ATP-binding/permease protein (macB) (Table 5).
Virulence related genes in the inversion fragment
According to the annotation results of the VFDB (Virulence Factors of Pathogenic Bacteria) database, the two strains contain 336 putative virulence related genes, of which 221 virulence genes are in the inversion fragment. Among them, several important virulence factor such as capsule (cpsA, cpsB, cpsC, cpsD), CAMP factor (cfa/cfb), C5a peptidase (scpA, scpB, scpI), phosphoglucomutase (pgmA), beta-hemolysin/cytolysin (cylG, cylA, cylA, cyll), hemolysin A/B (hlyA, hlyB), laminin-binding protein (lmb), arein the inversion fragment (Table 6).
Discussion
There have been many studies to distinguish the genotypes of S.iniae using molecular methods. However, the multidrug resistance and virulence differentiation characters and the underlying mechanisms with different genotypes have been poorly explored to date. The investigation on the molecular mechanisms of drug resistance and virulence variation is essential to the prevention of further spreading of these multidrug resistance strains or the occurrence of new resistant strains. The results showed great significance to explain the multidrug resistant and virulence differentiation of S.iniae strains. As reflected by our results, BH16-24 is a native multidrug resistant and low virulence strain. Further genomic comparisons between BH15-2 and BH16-24 revealed valuable information on the possible multidrug resistance and virulence differentiation among S.iniae strains. To our knowledge, this is the first report on the differences in biological characteristics and genomes among different genotypes of S.iniae.
Fish infected of S.iniae often show a variety of clinical signs, such as anorexia, lethargy, erratic swimming, and visceral hemorrhage [19]. Similarly in the present study, clinical signs including meningoencephalitis, enteritis, hemorrhage of pterygiophore, operculum, liver hemorrhage were also observed. In our previous study, the two genotypes strains of S.iniae showed identical phenotypic features [16]. However, the growth ability of the two strains was different, where the growth rate of BH16-24 was significantly faster than that of BH15-2 at the exponential stage according to the growth curve.
S.iniae recovered from farmed fish were genetically distinct from wild reef fish and exhibited a trend toward higher minimal inhibitory concentrations against several antibiotics [20]. Consistently, our results suggest that BH15-2 was susceptible to most of the tested antibiotics except neomycin and gentamycin. In contrast, BH16-24 exhibited multidrug resistance, including high resistance against penicillin, ceftazidime, sulfasomizole, compound sulfamethoxazole, spectinomycin, polymyxin B, rifampin and intermediate against ceftriaxone sodium, ampicillin, cefazolin, neomycin, and gentamycin. In the previous study, S.iniae obtained from fish were susceptible to most of the antibiotics [4, 21]. However, according to recent reports, more and more resistant strains of S.iniae was found to be resistant to many important antibiotics including amoxicillin, penicillin, ampicillin, gentamycin,spectinomycin, amikacin, neomycin, enrofloxacin, lincomycin, and sulfamethoxazole [17, 22, 23]. The emergence of multidrug resistance strain is worrisome. In addition, both BH15-2 and BH16-24 remained highly susceptible to tetracycline, erythromycin, norfloxacin, azithromycin, ciprofloxacin, chloramphenicol and deoxytetracycline, which is indicative that these antibiotics might be useful in controlling the disease in the future.
The relationship between genotype and virulence of S.iniae isolates was investigated. Fuller et al. reported that S.iniae virulence is associated with distinct genetic profile and demonstrated differences between pathogenic and nonpathogenic isolates [10]. The isolates belong to genotype 1 in rep-PCR analysis showed a high virulence in the flounder, while the isolated belonging to genotype 2 were relatively low in virulence [9]. Similarly, among 14 isolates of S.iniae with six clonal patterns, two clones have one fold lower in pathogenicity challenge than others [11]. 11 S.iniae isolates from diseased wild and farmed fish showed significant differences in virulence and persistence, with a certain correlation to genogroup [24]. In this study, the pathogenicity of the two strains were compared by intraperitoneal injection in T.ovatus. Similar results were obtained in our analyses, where the virulence of genotype 1 strain BH15-2 showed much greater virulence than that of genotype 2 strain BH16-24.
Genetic variability not only depends on point mutations but also largely on horizontal genes transfer and intra-genomic rearrangements, which may disrupt chromosome organization [25]. During cell division, the symmetry of the origin and terminus loci play a role in the precise choreography of replicated chromosome separation [26]. Genome arrangement may affect gene expression and is thought to be related to the diversity of phenotypes seen in organisms [27]. Studies have shown that after the common ancestor branch of Streptococcus, chromosome reversal across the replication axis often occurs in a single streptococcal species [28]. A previously study demonstrated that unbalanced genome is prone to generate DNA rearrangements in the M3 strain of S.pyogenes, which is caused by the loss or acquisition of phages [29]. Moreover, results from a previous study suggest that a large-scale genomic rearrangement may resulted in biological discrepancies between a native avirulent and highly virulent S.suis strains [30]. Compared with its parental strain GX005, YM011 had a 0.4 M large inversion fragment which may result in abnormal expression of some genes including drug resistance genes and virulence factors, eventually leads to virulence attenuation [31]. By comparing the whole genomes of the two strains with other S.iniae which were also isolated from diseases fish, a large-scale inverted fragment about 1.28 Mbp was found as the major difference in BH16-24. The genomic recombination in BH16-24 leads to abnormal expression of some resistance and virulence genes in the fragment, eventually leads to multidrug resistance and virulence attenuation. Thus, our findings are consistent with the notion that inversion events of intergenic regions correlate to phenotypic variation [32].
β-lactam antibiotics are commonly being used in aquaculture in some parts of the world to treat bacterial infections [33]. Penicillin-binding proteins (PBPs) are membrane proteins involved in the biosynthesis of peptidoglycans in bacterial cell walls. The β-lactam antibiotics participate in the synthesis of peptidoglycans by binding to the active site of PBPs, thereby disrupting the formation of normal cell walls and inducing cell death through bacteriolysis [34]. The major PBPs causing for the β-lactam antibiotics resistance of S. pneumoniae are PBP2x, PBP2b and PBP1a [35]. In our study, BH16-24 was resistant to the two β-lactam antibiotics i.e., penicillin, ceftazidime, and intermediate resistant toward the other three β-lactam antibiotics used in this study, i.e., ampicillin, cefazolin, ceftriaxone sodium, whereas BH15-2 was susceptibility to these antibiotics. Pbp1a, pbp2x, and pbp2b gene of BH16-24 is located on an inverted fragment, and its abnormal expression changes the amount of PBP protein, which may lead to BH16-24 resistance to β-lactam antibiotics. In addition, the present of the RNA polymerase beta subunit (rpoB) with mutations have been reported in rifamycin resistance in Brucella melitensis, Mycobacterium tuberculosis, and other microorganisms [36, 37]. The DNA gyrase (gyrA) have been suggested to be involved in fluoroquinolone resistant isolates of S.agalatiae [38]. The dihydrofolate reductase enzyme encoded by the dfr gene promotes bacterial resistance to trimethoprim [39, 40]. Aminoglycoside antibiotics are transported by the oligopeptide transport system, thus when the gene for oppA protein was deleted, sensitivity to aminoglycoside antibiotics was greatly decreased [41]. Aminoglycoside phosphotransferases which encoded by aph gene are bacterial enzymes responsible for the inactivation of aminoglycoside antibiotics by O-phosphorylation [42]. In this study, genes such as gryA, gryB, rpoB, dfrA, oppA, aph were in the inverted fragment, which may be related to the resistance of BH16-24 to sulfonamide antibiotics, aminoglycoside antibiotic, rifampin. Moreover, the efflux pumps play an important role in conferring resistance by actively excreting the harmful antibiotic drugs from the bacteria [43]. Efflux pumps exists in almost all bacterial species. It can not only excrete a wide range of antibiotics, but also reduce intracellular antibiotic concentration and promote mutation accumulation [43, 44]. Here, we identified several efflux-pump related genes from different efflux-pump families among the genomes. The abnormal expression of these resistance genes on the inverted fragment may lead to multiple drug resistance of BH16-24.
To date, several virulence-associated factors (VAFs) have been characterized which are closely related to the pathogenesis of S.iniae infection. The most critical VAF validated in S.iniae including polysaccharide capsular (cps), phosphoglucomutase (pgmA), M-like protein (simA), beta-hemolysin/cytolysin (cyl), C5a peptidase (scp). CPS of S.iniae is a major virulence factor that provides resistance to the bactericidal activity of phagocytes and stimulates prolonged inflammatory responses, which including cpsA, cpsB, cpsC, cpsD, cpsE, cpsG, cpsJ [45, 46]. The streptococcal CpsA protein was reported to associated with important virulence determinants, including cell wall processing [47], polysaccharide synthesis [48], and reaction to antimicrobial stress [49]. The absence of cpsD could reduce the ability of S.iniae to survive phagocytosis and escape the immune system [50]. CpsJ influences the synthesis of CPS and loss of this protein showed lower virulence in a channel catfish infection model [51]. The pgm gene play an pivotal role in normal cell wall morphology, surface capsule expression, resistance to innate immune clearance mechanisms, therefore it is necessary for the virulence in S.iniae [52]. M-like protein contributes to cellular adherence and invasion and provides resistance to phagocytic killing based in vitro cell analysis [53]. The virulence factor Beta-hemolysin/cytolysin is encoded by cyl gene, capable of exerting cytolytic, proapoptotic, proinvasive, proinflammatory, or antiphagocytic effects on a variety of target cells [54]. In our study, both genomes harbor 336 putative genes involved in virulence, among which 221 virulence genes are located on the inversion fragment. Among the 221 virulence genes, we observed genes encoding CPS, phosphoglucomutase, M-like protein, beta-hemolysin/cytolysin, C5a peptidase. The abnormal expression of these virulence-related genes on the inverted fragments contributed to the reduced virulence of BH16-24.
Prophage can enhance bacterial adherence to animal cells, encode a series of bacterial toxins, and affect bacterial biofilm formation, which is closely related to bacterial virulence [55,56,57]. As reported by Wang et al., the phage was the major reason of causing different levels of virulence between S.agalactiae strains [58]. Prophage analysis of BH15-2 and BH16-24 showed that both strains harbored 11 identical incomplete and questionable prophages, although several of them are in different positions due to the inversion fragment. Moreover, there is a 46 kb length, intact phage sequence which only existed in the BH15-2 genome. The GC contents of the prophage fragment (35.46%) deviate from the host genomes (36.80%). The prophage encoded 77 proteins, and 17 genes encoded hypothetical proteins, the other 60 genes encoded phage hit proteins such as phage lysin, phage tail protein, and phage integrase. However, the function of the prophage in BH15-2 is still unknown, and whether it is related to its virulence needs further experiments to verify.
Conclusions
In summary, we compared the biological characteristics such as growth, virulence, drug resistance, and whole genome sequence of two different genotypes of S.iniae. Comparative genomic studies of BH15-2 and BH16-24 showed that the main difference is a 1.28 Mbp inversion fragment. The inversion fragment may lead to abnormal expression of drug resistant and virulence genes, which is believed to be the main reason for the multiple resistance and weakened virulence of BH16-24. Aside from the differences in genomic rearrangement, BH15-2 harbored a novel intact prophage which is absented in BH16-24. There was finished concordance between genotypic evidence and biological characteristics. Further research is needed on how the genomic rearrangements affect the gene expression, drug resistance, and pathogenicity of S.iniae.
Materials and methods
Bacterial strains
The S.iniae strain BH15-2 and BH16-24 were originally isolated from the livers of moribund cultured golden pompano on two separate farms of China in 2015 ang 2016, respectively [16]. The fish from the two outbreaks displayed similar clinical signals, and the cumulative mortality rate was approximately 20–30%. Briefly, the stored strains were removed from − 80 ℃ refrigerator and streaked onto the BHI plate, cultured at 30 ℃ for 24–48 h. Then picked up a single colony and inoculated into 10 ml of BHI medium, cultivated at 30 ℃ by shaking.
Growth analysis
To measure the growth level of bacteria in BHI broth, overnight cultures of BH15-2 and BH16-24 were inoculated into BHI with an initial OD600 of 0.01 in a ratio of 1:50, respectively. The cultures were collected every 2 h of intervals and the optical density was measured at 600 nm from 0 to 18 h of growth at 30 ℃ with shaking in 180 r/min. Data were expressed as mean ± standard deviation (SD). Statistical analyses were performed using Student’s t-test using SPSS 21.0 and p < 0.05 was considered significant.
Comparison of the virulence between BH15-2 and BH16-24
Adult golden pompano with a mean weight of 50 g, were purchased from a local fish farm and maintained in a 16 m3 tank with aeration and sand-filtered seawater supply. The fish (n = 150) were acclimated for 2 weeks at 28–30 ℃ and checked randomly to confirm that no bacterial infected. Fish were fed twice daily with commercial fish expanded pellets (Guangdong Yuehai Feed Group), and waste was removed daily. The bacterial concentration determined by plating 10-fold serial dilutions onto BHI agar plates. Suspensions from 1.5 × 108 to 1.5 × 105 CFU/ml were prepared by serial 10-fold dilution. Fish were divided into nine groups with 16 fish per group. Before experimental treatment or organ extraction, fish were euthanized in 100 mg/L MS-222 (Sigma, USA). Eight groups were injected intraperitoneally (i. p.) with 0.1 ml of diluted bacterial cell suspension of the strain BH15-2 and BH16-24 at the final concentration of 1.5 × 108, 1.5 × 107, 1.5 × 106 and 1.5 × 105 CFU/ml, respectively. The control group were i. p. with the same amount of sterilized saline. The mortalities were recorded every 24 h interval for 28 days post-infection. The bacteria were reisolated from the brain, kidney and spleen tissues of all dead fishes at the end of the experiment and identified.
Comparison of the antibiogram between BH15-2 and BH16-24
The antibiogram study of the bacterium was determined on BHI plates according to the disc diffusion method, and the diameters of the inhibition zones were measured using Vernier calipers. The tested antibiotic impregnated discs were summarized in Table 2. Resistant, intermediate, and susceptible phenotype determinations were based on manufacturer guidelines (Hangzhou Binhe Microorganism Reagent Co., Ltd., China). Multiple antibiotic resistance (MAR) index of the two strains against the tested antibiotics was calculated by following the procedure described by Krumperman [59].
Genome sequencing and annotation
The genomes of BH15-2 and BH16-24 were sequenced by PacBio sequencing at the Beijing Biomarker Bioinformatics Technology Co., Ltd. For genome assembly, the filtered subreads were assembled by Canu v1.5 software [60], and then circlator v1.5.5 was taken to cyclizing assembly genome [61].
Genome component prediction
For genome component prediction, coding genes prediction was performed by Prodigal v2.6.3 [62]. The GenBlastA v1.0.4 program was used to scan the whole genomes after masking predicted functional genes [63]. Putative candidates were then analyzed by searching for non-mature mutations and frame-shift mutations using GeneWise v2.2.0 [64]. Transfer RNA (tRNA) genes were predicted with tRNAscan-SE v2.0 [65], Ribosome RNA (rRNA) genes were predicted with Infernal v1.1.3 [66]. Repetitive sequences were predicted using RepeatMasker v4.0.5 [67]. CRT v1.2 was used for CRISPR identification [68]. Circos v0.66 was used to draw genomic circles [69].
Gene functions
For functional annotation, the predicted proteins were blast (e-value: 1e− 5) against Nr (Non-Redundant Protein Database databases), Swiss-Prot, TrEMBL, KEGG (Kyoto Encyclopedia of Genes and Genomes), eggNOG, GO (Gene ontology). The pathogenicity and drug resistance of pathogenic bacteria were analyzed using VFDB and ARDB (Antibiotic Resistance Genes Database).
Comparative genomics analysis
Genomic synteny was analyzed using Mauve v2.3.1 [70]. GIs of BH15-2 and BH16-24 were determined with IslandViewer 4 [71]. PHASTER was used to identify prophage sequences [72]. Prophage Hunger was used to predict the intact prophage [73].
Data Availability
The whole-genome sequence data of BH15-2 and BH16-24 have been deposited at the National Center for Biotechnology Information database with the accession number CP132229 and CP132230.
Abbreviations
- RAPD:
-
Random amplified polymorphic DNA
- rep-PCR:
-
Repetitive primer polymerase chain reaction
- BHI:
-
Brain heart infusion
- i. p.:
-
Injected intraperitoneally
- Dpi:
-
Days post-infection
- MDR:
-
Multidrug-resistant
- CDS:
-
Coding sequence
- LCBs:
-
Local collinear blocks
- VFDB:
-
Virulence Factors of Pathogenic Bacteria
- ARDB:
-
Antibiotic Resistance Genes Database
- Nr:
-
Non-Redundant Protein Database databases
- KEGG:
-
Kyoto Encyclopedia of Genes and Genome
- GO:
-
Gene ontology
- ori/ter axis:
-
Origins/terminus axis
- GIs:
-
Gene Islands
- PBPs:
-
Penicillin-binding proteins
- SD:
-
Standard deviation
References
Zhang BC, Zhang J, Sun L. Streptococcus iniae SF1: complete genome sequence, Proteomic Profile, and Immunoprotective Antigens. PLoS ONE. 2014;9(3):e91324. https://doi.org/10.1371/journal.pone.0091324
Chen DF, Peng S, Chen DY, Yang F, Liu JX, Wang J, Liu QW, Huang XL, Ouyang P, Wang KY, Li ZQ, Geng Y. Low lethal doses of Streptococcus iniae caused enteritis in Siberian sturgeon (Acipenser baerii). Fish Shellfish Immunol. 2020;104(1):020. https://doi.org/10.1016/j. fsi.2020.06.
Cai XH, Peng YH, Wang ZC, Huang T, Xiong XY, Huang YC, Wang B, Xu LW, Wu ZH. Characterization and identification of Streptococci from golden pompano in China. Dis Aqut Org. 2016;119:207–17. https://doi.org/10.3354/dao02998
Guo S, Mo ZQ, Wang Z, Xu J, Li YW, Dan XM, Li AX. Isolation and pathogenicity of Streptococcus iniae in offshore cage-cultured Trachinotus ovatus in China. Aquaculture. 2018;492:247–52. 10.1016/j. aquaculture. 2018.04. 015.
Sun JR, Yan JC, Yeh CY, Lee SY, Lu JJ. Invasive Infection with Streptococcus iniae in Taiwan. J Med Microbiol. 2007;56:1246–9. https://doi.org/10.1099/jmm.0.47180-0
Pier GB, Madin SH. Streptococcus iniae sp. nov., a beta-hemolytic Streptococcus isolated from an Amazon freshwater dolphin, Inia geoffrensis. Int J Syst Bacteriol. 1976;26:545–53. https://doi.org/10.1099/00207713-26-4-545
Bachrach G, Zlotkin A, Hurvitz A, Evans DL, Eldar A. Recovery of Streptococcus iniae from diseased fish previously vaccinated with a Streptococcus vaccine. Appl Env Microbiol. 2001;67(8):3756–8. https://doi.org/10.1128/AEM.67.8.3756-3758.2001
Malathum K, Singh KV, Weinstock GM, Murray BE. Repetitive sequence-based PCR versus pulsed-field gel electrophoresis for typing of Enterococcus faecalis at the subspecies level. J Clin Microbiol. 1998;36(1):211–5. https://doi.org/10.1128/JCM.36.1.211-215.1998
Kim MS, Jin JW, Han HJ, Choi HS, Hong S, Cho JY. Genotype and virulence of Streptococcus iniae isolated from diseased olive flounder Paralichthys olivaceus in Korea. Fish Sci. 2014;80(6):1277–84. https://doi.org/10.1007/s12562-014-0804-0
Fuller JD, Bast DJ, Nizet V, Low DE, Azavedo JCS. Streptococcus iniae virulence is associated with a distinct genetic profile. Infect Immun. 2001;69(4):1994–2000. https://doi.org/10.1128/IAI.69.4.1994-2000.2001
Younes AM, Gaafar AY, El-Din A, Abu-Bryka Z, Bayoumy EM. Genotyping and pathogenicity of Streptococcus iniae strains recovered from cultured Oreochromis niloticus at Kafr El-Shiekh Governorate, Egypt. Egypt J Aquat Biology Fisheries. 2019;23(3). https://doi.org/10.21608/ejabf.2019.51630
Higuera-Llanten S, Vasquez-Ponce F, Barrientos-Espinoza B, Mardones FO, Marshall SH, Olivares-Pacheco J. Extended antibiotic treatment in salmon farms select multiresistant gut bacteria with a high prevalence of antibiotic resistance genes. PLoS ONE. 2018;13(9):e0203641. https://doi.org/10.1371/journal.pone.0203641
Delannoy CMJ, Zadoks RN, Crumlish M, Rodgers D, Lainson FA, Ferguson HW, Turnbull J, Fontaine MC. Genomic comparison of virulent and non-virulent Streptococcus agalactiae in fish. J Fish Dis. 2014;39(1):13–29. https://doi.org/10.1111/jfd.12319
Fanelli F, Di Pinto A, Mottola A, Mule G, Chieffi D, Baruzzi F, Tantillo G, Fusco V. Genomic characterization of Arcobacter butzleri isolated from Shellfish: Novel Insight into Anbiotic Resistance and Virulence determinants. Front Microbiol. 2019;10:670. 10.3389 /fmicb.2019.00670.
Eunsuk K, Soyeon P, Seongbeom C, Tae-Wook H, Hyunjin Y. Comparative Genomics Approaches to understanding virulence and Antimicrobial Resistance of Salmonella Typhimurium ST1539 isolated from a Poultry Slaughterhouse in Korea. J Microbiol Biotechn. 2019;29(6):962–72. https://doi.org/10.4014/jmb.1904.04028
Xiong XY, Huang GQ, Wang ZC, Wen X. Molecular typing, antibiogram type and detection of virulence genes of Streptococcus iniae strains isolated from golden pompano (Tranchinotus ovatus) in Guangxi Province. J Fisheries China. 2018;42(4):586–95. https://doi.org/10.11964/jfc.20170410791
Pridgeon JW, Zhang D. Complete genome sequence of a virulent strain, Streptococcus iniae ISET0901, isolated from diseased tilapia. Genome Announc. 2014;2(2):e00553–14. https://doi.org/10.1128/genomeA.00553-14
Rajoo S, Jeon W, Park K, Yoo S, Yoon I, Lee H, Ahn J. Complete genome sequence of Streptococcus iniae YSFST01-82, isolated from olive flounder in Jeju, South Korea. Genome Announc. 2015;3(2):e00319–15. https://doi.org/10.1128/genomeA.00319-15
Rahmatullah M, Ariff M, Kahieshesfandiari M, Daud HM, Zamri-Saad M, Sabri MY, Amal MNA, Ina-Salwany MY. Isolation and pathogenicity of Streptococcus iniae in cultured red hybrid tilapia (Oreochromis sp.) in Malaysia. J Aquat Anim Health. 2017;29:208–13. https://doi.org/10.1080/08997659.2017.1360411
Chou L, Griffin MJ, Fraites T, Ware C, Ferguson H, Keirstead N, Brake J, Wiles J, Hawke JP, Kearney MT, Getchell RG, Gaunt P, Soto E. Phenotypic and genotypic heterogeneity among Streptococcus iniae isolates recovered from cultured and wild fish in North America, Central America and the Caribbean Islands. J Aquat Anim Health. 2014;26(4):263–71. https://doi.org/10.1080/08997659.2014.945048
Park YK, Nho SW, Shin GW, Park SB, Jang HB, Cha MA, Kim YR, Dalvi RS, Kang BJ. Antibiotic susceptibility and resistance of Streptococcus iniae and Streptococcus parauberis isolated from olive flounder (Paralichthys olivaceus). Vet Microbiol. 2009;136(1–2):76–81. https://doi.org/10.1016/j.vetmic.2008.10.002
Muhammad M, Zhang T, Gong SY, Bai J, Ju JS, Zhao BH. Streptococcus iniae: a growing threat and causative Agent of Disease Outbreak in Farmed Chinese Sturgeon (Acipenser sinensis). Pakistan J Zool. 2020;52(5):1–9. https://doi.org/10.17582/journal.pjz/20190209200236
Feng Y, Bai MH, Geng Y, Chen DF, Huang XL, Ouyang P, Guo HR, Zuo ZC, Huang C, Lai WM. The potential risk of antibiotic resistance of Streptococcus iniae in sturgeon cultivation in Sichuan, China. Environ Sci Pollut Res. 2021;28:69171–80. https://doi.org/10.1007/s11356-021-15501-1
Heckman TI, Griffin MJ, Camus AC, LaFrenta BR, Morick D, Smirnov R, Ofek T, Soto E. Multilocus sequence analysis of diverse Streptococcus iniae isolates indicates an underlying genetic basis for phenotypic heterogeneity. Dis Aquat Org. 2020;141:53–69. https://doi.org/10.3354/dao03521
Savic DJ, Nguyen SV, McCullor K, McShan WM. Biological Impact of a large-scale genomic inversion that grossly disrupts the relative positions of the origin and terminus loci of the Streptococcus pyogenes chromosome. J Bacteriol. 2019;201(17):e00090–19. https://doi.org/10.1128/JB.00090-19
Rocha EP. The replication-related organization of bacterial genomes. Microbiology. 2004;150:1609–27. https://doi.org/10.1099/mic.0.26974-0
Couturier E, Rocha EP. Replication-associated gene dosage effects shape the genomes of fast-growing bacteria but only for transcription and translation genes. Mol Microbiol. 2006;59:1506–18. https://doi.org/10.1111/j.1365-2958.2006.05046.x
Bentley RW, Leigh JA, Collins MD. Intrageneric structure of Streptococcus based on comparative analysis of small-subunit rRNA sequences. Int J Syst Evol Micr. 1991;41(4):487–94. https://doi.org/10.1099/00207713-41-4-487
Nakagawa I, Kurokawa K, Yamashita A, Nakata M, Hamada S. Genome sequence of an M3 strain of Streptococcus pyogenes reveals a large-scale genomic rearrangement in invasive strains and New insights into Phage Evolution. Genome Res. 2003;13(6A):1042–55. 10.1101/gr. 1096703.
Yao XY, Li M, Wang J, Wang CJ, Hu D, Zheng F, Pan XZ, Tan YL, Zhao T, Hu LW, Tang JQ, Hu FQ. Isolation and characterization of a native avirulent strain of Streptococcus suis serotype 2: a perspective for vaccine development. Sci Rep. 2015;5:9835. https://doi.org/10.1038/srep09835
Liu Y, Li LP, Yu FZ, Luo YJ, Liang WW, Yang Q, Wang R, Li M, Tang JY, Gu QH, Luo ZP, Chen M. Genome-wide analysis revealed the virulence attenuation mechanism of the fish-derived oral attenuated Streptococcus iniae vaccine strain YM011. Fish Shellfish Immunol. 2020;106(5):546–54. https://doi.org/10.1016/j.fsi.2020.07.046
Uwe G, Elzbieta B, Katrin G, Jessica S, Thomas R, Boyke B, Cathrin S, Daniela W, Anja P, Cynthia C, Wolfgang B, Jörg O, Ortrud Z, Rolf D, Heiko L. Comparative genome and phenotypic analysis of three Clostridioides difficile strains isolated from a single patient provide insight into multiple Infection of C. Difficile. BMC Genomics. 2018;19(1). https://doi.org/10.1186/s12864-017-4368-0
Park YH, Hwang SY, Hong MK, Kwon KH. Use of antimicrobials agents in aquaculture. Sci Tech Rev. 2012;31:189–97. https://doi.org/10.1186/1751-0147-54-22
Jacobs MR. Antimicrobial-resistant Streptococcus pneumoniae: trends and management. Expert Rev Anti Infect Ther. 2008;6(5):619–35. https://doi.org/10.1586/14787210.6.5.619
Zapun A, Contreras-Martel C, Vernet T. Penicillin-binding proteins and beta-lactam resistanc. FEMS Microbiol Rev. 2008;32(2):361–85. https://doi.org/10.1111/j.1574-6976.2007.00095.x
Dadar M, Bazrgari N, Garosi GA, Hassan S. Investigation of mutations in the rifampin-resistance-determining region of the rpoB Gene of Brucella melitensis by Gene Analysis. Jundishapur J Microbiol. 2021;14(2). https://doi.org/10.5812/jjm.115526
Zeng MC, Jia QJ, Tang LM. rpoB gene mutations in rifampin-resistant Mycobacterium tuberculosis isolates from rural areas of Zhejiang. China J Int Med Res. 2021;49(3):030006052199759. https://doi.org/10.1177/0300060521997596
Kawamura Y, Fujiwara H, Mishima N, Tanaka Y, Tanimoto A, Ikawa S, Itoh Y, Ezaki T. First Streptococcus agalactiae isolates highly resistant to quinolones, with point mutations in gyrA and parC. Antimicrob Agents Chemother. 2003;47:3605–9. https://doi.org/10.1128/AAC.47.11.3605-3609.2003
Ho PL, Wong RC, Chow KH, Que TL. Distribution of integron-associated trimethoprim–sulfamethoxazole resistance determinants among Escherichia coli from humans and food-producing animals. Lett Appl Microbiol. 2009;49:627–34. https://doi.org/10.1111/j.1472-765X.2009.02717.x
Hu LF, Chang X, Ye Y, Wang ZX, Shao YB, Shi W, Li X, Li JB. Stenotrophomonas maltophilia resistance to trimethoprim/sulfamethoxazole mediated by acquisition of Sul and dfrA genes in a plasmid-mediated class 1 integron. Int J Antimicrob Ag. 2011;37(3):230–4. https://doi.org/10.1016/j.ijantimicag.2010.10.025
Kashiwagi K, Miyaji A, Ikeda S, Tobe T, Igarashi K. Increase of sensitivity to aminoglycoside antibiotics by polyamine-induced protein (oligopeptide-binding protein) in Escherichia coli. J Bacteriol. 1992;174(13):4331–7. https://doi.org/10.1111/j.1365-2672.1992
Lallemand P, Leban N, Kunzelmann S, Chaloin L, Serpersu EH, Webb MR, Barman T, Lionne C. Transient kinetics of aminoglycoside phosphotransferase (3′)-IIIa reveals a potential drug target in the antibiotic resistance mechanism. FEBS Lett. 2012;586(23):4223–7. https://doi.org/10.1016/j.febslet.2012.10.027
Sun JJ, Deng ZQ, Yan AX. Bacterial multidrug efflux pumps: mechanisms, physiology and pharmacological exploitations. Biochem Biophys Res Commun. 2014;453:254–67. https://doi.org/10.1016/j.bbrc.2014.05.090
Balganesh M, Dinesh N, Sharma S, Kuruppath S, Nair AV, Sharma U. Efflux pumps of Mycobacterium tuberculosis play a significant role in antituberculosis activity of potential drug candidates. Antimicrob Agents Chemother. 2012;56:2643–51. https://doi.org/10.1128/AAC.06003-11
Griffin AM, Morris VJ, Gasson MJ. The cpsABCDE genes involved in polysaccharide production in Streptococcus salivarius ssp. thermophilus strain NCBF 2393. Gene. 1997;183(1):23–7. https://doi.org/10.1016/S0378-1119(96)00405-2
Pirollo T, Perolo A, Mantegari S, Barbieri I, Scali F, Alborali GL, Salogni C. Mortality in farmed European eel (Anguilla anguilla) in Italy due to Streptococcus iniae. Acta Vet Scand. 2023;65(1):5–13. https://doi.org/10.1186/s13028-023-00669-y
Chatfield CH, Koo H, Quivey RGJ. The putative autolysin regulator LytR in Streptococcus mutans plays a role in cell division and is growth-phase regulated. Microbiology. 2005;151:625–31. https://doi.org/10.1099/mic.0.27604-0
Cieslewicz MJ, Kasper DL, Wang Y, Wessels MR. Functional analysis in type Ia group B Streptococcus of a cluster of genes involved in extracellular polysaccharide production by diverse species of Streptococci. J Biol Chem. 2001;276:139–46. https://doi.org/10.1074/jbc.M005702200
Utaida S, Dunman PM, Macapagal D, Murphy E, Wilkinson BJ. Genome-wide transcriptional profiling of the response of Staphylococcus aureus to cell-wall-active antibiotics reveals a cell-wall-stress stimulon. Microbiology. 2003;149:2719–32. https://doi.org/10.1099/mic.0.26426-0
Colussi S, Pastorino P, Mugetti D, Antuofermo E, Sciuto S, Esposito G, Polinas M, Tomasoni M, Burrai GP, Fernández-Garayzábal JF, Acutis PL, Pedron C, Prearo M. Isolation and genetic characterization of Streptococcus iniae virulence factors in Adriatic sturgeon (Acipenser naccarii). Microorganisms. 2022;10(5):883–94. https://doi.org/10.3390/microorganisms10050883
Zeng Y, He Y, Wang KY, Wang J, Zeng YK, Chen YX, Chen DF, Geng Y, OuYang P. cpsJ gene of Streptococcus iniae is involved in capsular polysaccharide synthesis and virulence. Antonie Van Leeuwenhoek. 2016;109(11):1–10. https://doi.org/10.1007/s10482-016-0750-1
Buchanan JT, Stannard JA, Lauth X, Ostland VE, Nizet V. Streptococcus iniae phosphoglucomutase is a virulence factor and a target for vaccine development. Infect Immun. 2005;73(10):6935–44. https://doi.org/10.1128/IAI.73.10.6935-6944.2005
Locke JB, Aziz RK, Vicknair MR, Nizet V, Buchanan JT, Ahmed N. Streptococcus iniae M-like protein contributes to virulence in fish and is a target for live attenuated vaccine development. PLoS ONE. 2008;3(7):e2824. https://doi.org/10.1371/journal.pone.0002824
Liu GY, Nizet V. The group B streptococcal β-hemolysin/cytolysin - ScienceDirect. Compr Sourceb Bacterial Protein Toxins (Third Edition). 2006;737–47.
Li DZ, Chen Y, Qian XJ, Liu Y, Ren JL, Xue F, Sun JH, Tang F, Dai JJ. orf20 in prophage phiv142-3 contributes to the adhesion and colonization ability of avian pathogenic Escherichia coli strain DE142 by affecting the formation of flagella and I fimbriae. Vet Microbiol. 2019;235:301–9. https://doi.org/10.1016/j.vetmic.2019.07.020
Li Y, Liu X, Tang K, Wang X. Prophage encoding toxin/antitoxin system PfiT/PfiA inhibits Pf4 production in Pseudomonas aeruginosa. Microb Biotechnol. 2020;13. 10.1111/ 1751-7915.13570.
Smirnova NI, Agafonov DA, Kul’shan’ TA, Yu E, Ya S, Krasnov M, Lozovsky V, Kutyrev VV. Effect of CTXφ prophage deletion in Cholera agent on expression of regulatory genes controlling virulence and biofilm formation. Russian J Genet. 2017;53:302–13. https://doi.org/10.1134/s1022795417020119
Wang R, Li LP, Huang T, Lei AY, Huang Y, Luo FG, Wang DY, Huang WY, Chen M, Huang J. Genomic comparison of virulent and non-virulent serotype V ST1 Streptococcus agalactiae in fish. Vet Microbio. 2017;207:164–9. https://doi.org/10.1016/j.vetmic.2017.06.007
Krumperman PH. Multiple antibiotic indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods[J]. Appl Environ Microb. 1983;46(1):165–70. https://doi.org/10.1128/AEM.46.1.165-170.1983
Koren S, Walenz BP, Berlin K, Miller JR, Bergman NH, Phillippy AM. Canu: scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017;27(5):722–36. https://doi.org/10.1101/gr.215087.116
Hunt M, De Silva N, Otto TD, Parkhill J, Keane JA, Harris SR. Circlator: automated circularization of genome assemblies using long sequencing reads. Genome Biol. 2015;16:294. https://doi.org/10.1186/s13059-015-0849-0
Hyatt D, Chen GL, LoCascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics. 2010;11. https://doi.org/10.1186/1471-2105-11-119. ,119.
She R, Chu JSC, Wang K, Pei J, Chen NS. GenBlastA: enabling BLAST to identify homologous gene sequences. Genome Res. 2009;19(1):143–9. https://doi.org/10.1101/gr.082081.108
Birney E, Clamp M, Durbin R, GeneWise. And Genomewise Genome Research. 2004;14(5):988–95. https://doi.org/10.1101/gr.1865504
Chan PP, Lowe TM. tRNAscan-SE: searching for tRNA genes in genomic sequences. Methods Mol Biol. 2019;1062:1–14. https://doi.org/10.1007/978-1-4939-9173-0_1
Nawrocki EP, Eddy SR. Infernal 1.1: 100-fold faster RNA homology searches. Bioinformatics. 2013;222933–5. https://doi.org/10.1093/bioinformatics/btt509
Tarailo-Graovac M, Chen NS. Using RepeatMasker to identify repetitive elements in genomic sequences. Curr Protocols Bioinf. 2009. 4.10.1–4.10.14.
Bland C, Ramsey TL, Sabree F, Lowe M, Brown K, Kyrpides NC, Hugenholta P. CRISPR recognition tool (CRT): a tool for automatic detection of clustered regularly interspaced palindromic repeats. BMC Bioinformatics. 2007;8(1):209. https://doi.org/10.1186/1471-2105-8-209
Krzywinski M, Schein J, Birol I, Connors J, Gascoyne R, Horsman D, Jones SJ, Marra MA. Circos: an information aesthetic for comparative genomics. Genome Res. 2009;19(9):1639–45. https://doi.org/10.1101/gr.092759.109
Darling ACE, Mau B, Blattner FR, Perna NT, Mauve. Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004;14(7):1394–403. https://doi.org/10.1101/gr.2289704
Bertelli C, Laird MR, Williams KP, Simon Fraser University Research Computing Group, Lau BY, Hoad G, Winsor G, Brinkman FSL. IslandViewer 4: expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017;45(W1):W30–5. https://doi.org/10.1093/nar/gkx343
Arndt D, Grant JR, Marcu A, Sajed T, Pon A, Liang Y, Wishart DS. PHASTER: a better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016;44:W16–W21. https://doi.org/10.1093/nar/gkw387
Song WC, Sun HX, Zhang C, Cheng L, Peng Y, Deng Z, Wang D, Wang Y, Hu M, Liu W, Yang H, Shen Y, Li J, You L, Xiao M. Prophage Hunter: an integrative hunting tool for active prophages. Nucleic Acids Res. 2019;47(W1):W74–W80. https://doi.org/10.1093/nar/gkz380
Acknowledgements
Not applicable.
Funding
This work was supported by Natural Science Foundation of Guangxi Province [Grant no. 2020GXNSFBA159011], Scientific Research and Technology Development Program of Guangxi Province [Grant no. AB18221112].
Author information
Authors and Affiliations
Contributions
X.X. preformed the experiments and wrote the paper. R.C. contributed to collecting the data. J.L. contributed to data analysis.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All animal experiments were performed according to the Regulations for the Administration of Affairs Concerning Experimental Animals (Ministry of Science and Technology, China,2004). Ethical approval of the respective Animal Research and Ethics Committees of Guangxi Academy of Sciences. All methods are reported in accordance with ARRIVE guidelines.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
12864_2023_9882_MOESM1_ESM.docx
Supplementary Figure 1: Circular visualization of the predicted prophage on BH15-2 (A) and BH16-24 (B) strains. The analysis was conducted in PHASTER. The interactive visualization of the distinct prophage across the genomes is shown with blocks colored according to the predictor tool as described: red represents incomplete prophages, blue represents questionable prophages, green represents intact prophages
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Xiong, X., Chen, R. & Lai, J. Comparative genomics analysis of Streptococcus iniae isolated from Trachinotus ovatus: novel insight into antimicrobial resistance and virulence differentiation. BMC Genomics 24, 775 (2023). https://doi.org/10.1186/s12864-023-09882-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12864-023-09882-5