
Abstract
The EU Common Agricultural Policy (CAP) has tried to counteract negative impacts of intensive agriculture on biodiversity and associated ecosystem services mostly by means of voluntary agri-environment schemes (AES). More recently, direct payments to farmers have been linked to the application of greening measures derived from previous AES experiences. AES and greening measures (CAP greening hereafter) have become the main farmland conservation tools in Europe due to large budgets and extensive application. Effectiveness of greening measures (i.e. differences in biodiversity or ecosystem service measurements that can be attributed to the application of such measures) has not yet been evaluated thoroughly, whereas evaluations of AES effectiveness—although still not systematically incorporated into policy design—have lead to the conclusion that AES generally increase farmland biodiversity at the field scale with effect sizes that depend on the surrounding landscape. On the basis of knowledge gaps derived from available AES evaluations, we develop a five-stage hierarchical decision-making proposal to improve the effectiveness of CAP greening. Effectiveness is difficult to predict because non-linear relationships between diversity and land-use intensity at both field and landscape scales constrain and modulate it. Besides, relationships vary regionally and among target species, species groups and ecosystem services. Hence, regional targeting, landscape-scale thinking, and learning processes linked to systematic evaluations are key elements in any decision-making procedure aimed at improving this effectiveness. Ideas and guidelines developed here will help to develop regionally adapted measures aimed at overcoming constraints to CAP greening effectiveness and improving farmland conservation policies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Agriculture is a key driver of biodiversity change all over the world [1]. Conversion of natural land into cropland, intensification of existing farmland and abandonment of extensive agricultural uses have strong effects of biological diversity, which vary, however, with regional history of land use [2, 3]. In Europe, and also probably in other areas with a long history of agriculture, biodiversity protection depends more and more on maintaining biodiversity in human-dominated landscapes [4, 5••].
The main responses in Europe to the environmental problems associated with agriculture were the agri-environment schemes (AES hereafter) linked to Pillar II (aids to rural development) of the EU’s Common Agricultural Policy (CAP). AES are voluntary contracts with interested farmers that involve the implementation of a set of management practices that are expected to benefit biodiversity or the environment and are usually applied at field or farm scales. This latter fact may limit its effectiveness if regional uptake is too low to fulfill the requirements of target groups (see below and [5••]). The CAP reform in 2014 tried to counteract this limitation by increasing environmental compliance for Pillar I (direct payments to all farmers given as production subsidies allocated per unit area of productive land), incorporating some management practices originally designed as AES into the practices obligatory for farmers to qualify for subsidies. Compulsory greening measures have generally not been incorporated, however, because of massive use of exemptions contemplated in the reformed CAP [6, 7••]. In spite of these limitations, AES and AES-derived greening measures (CAP greening hereafter) are prime conservation tools in Europe as they are applied in large parts of the territory (9–95 % of agricultural land depending on countries for AES, and 50 % for compulsory greening measures) and have budgets orders of magnitude higher than other conservation measures [5••, 7••].
The conservation potential of CAP greening has not been linked, however, to a systematic evaluation to improve its performance. No systematic data on the effects of compulsory greening measures are yet available [7••], and evaluation of AES’ ecological effectiveness has generally not been routinely and rigorously incorporated into policy evaluations [5••]. Numerous local and regional evaluation studies [e.g. 8–10] emerged after Kleijn et al.’s [11] worrying results for Dutch farmland, and have now been summarized in distinct reviews and meta-analyses [5••, 12–14, 15•, 16]. These studies, however, are still strongly biased to a handful of countries, mostly in northern and central Europe. For instance, the 284 studies listed in the last meta-analysis available [5••] have been carried out in 12 countries, 84 % in Germany, UK, Sweden, the Netherlands and Switzerland (see also [12]). On this basis, general findings are that application of AES generally increases farmland biodiversity at field scales, but effect sizes depend on the structure and management of the surrounding landscape. The likely mechanistic causes of this landscape dependence of AES effectiveness were established in the seminal works by Tscharntke et al. [17] and Concepción et al. [18]. However, this knowledge, which is key to developing effective conservation measures, does not seem to have been properly incorporated in CAP reforms. In fact, AES effectiveness has not improved since 2007 as compared to previous assessments [5••].
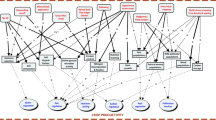
In this paper, we critically review current knowledge on the factors that condition AES- and AES-based greening measures of the CAP. In particular, we focus on regional- and landscape-scale effects on the effectiveness of field- and farm-scale conservation measures and their theoretical and mechanistic basis. On these grounds, we generate guidelines for developing regionally adapted measures aimed at overcoming constraints to AES effectiveness. These guidelines can be arranged into a hierarchical decision-making proposal (Table 1, Fig. 1), ultimately aimed at improving the ecological effectiveness of farmland conservation policies. We define effectiveness as the difference in biodiversity or ecosystem service target measurements between conventionally farmed areas and areas that incorporate conservation measures, regardless of initial levels of these measurements.

Flowchart for a five-stage decision-making proposal aimed at improving the ecological effectiveness of CAP greening measures. Greening: compulsory (Pillar I) wide-entry measures aimed at providing basic green landscape infrastructure; AES agri-environment schemes, HNVF high-nature value farmland, BACI before-after-control-impact experimental designs. See also Table 1
A Five-Stage Decision-Making Proposal for Improving CAP Greening
Targeting Conservation Measures
It is hard to evaluate the effectiveness—and even more so, the efficiency—of any proposal if it has no explicit goals [8, 19]. Nevertheless, many assessments are still based on indirect or vaguely defined targets [e.g. 22], and this is also despite the fact that targeting has been proven to influence both effectiveness itself [12, 23] and its assessments [19].
A key first stage is, thus, defining the specific aims of each AES within the general goal of biodiversity conservation (Table 1, Fig. 1). Kleijn et al. [19] proposed that conservation of intrinsic biodiversity values (i.e. rare and endangered species) should be restricted to the extensive systems still maintaining these species. In these systems—typically occurring in many Mediterranean, Alpine and Eastern regions of Europe—conservation measures should include analyses of the responses of target species, beside a general assessment of changes in overall abundance and species richness [e.g. 24, 25]. Very few evaluations are available for this kind of system, however [5••]. On the other hand, conservation measures in intensive systems devoid of rare species may rather be aimed at the conservation of functional biodiversity (i.e. the maintenance of ecosystem services provided by biodiversity such as pollination or pest control [19]). The restoration of populations of species common before intensification but decreasing nowadays, when known (e.g. farmland birds), can also be a valid aim [5••]. Most available assessments, and the general ideas derived from their meta-analyses, have been carried out in these intensive systems [8]. Here, evaluations of effectiveness can be based on overall species richness and abundance metrics, assuming that ecosystem services are related to these simple community variables, or on surrogates of such functional diversity. Under these assumptions, it is valid to use species characteristics linked to specific functions (e.g. pollination, pest control), the number of functional groups, or species within a given functional group, instead of species identity, to calculate diversity [e.g. 26, 27]. For instance, Garibaldi et al. [28••] showed that farms with high richness of wild insects have higher levels of pollination. However, more research is still necessary to assess whether effective AES in terms of taxonomic or functional diversity are in fact effective in promoting ecosystem services.
Considering Field- and Landscape-Scale Requirements of Targets
Batáry et al. [5••] concluded their meta-analysis of properly designed AES assessments by stating that the main factors affecting the effectiveness of field-scale measures are land-use intensity and the ecological contrast created by AES, modulated by local and regional effects of the surrounding landscape, thus supporting the model proposed by Kleijn et al. [19]. Field-scale requirements (availability of food and cover; Table 1, Fig. 1) are usually addressed by means of measures applied within productive areas (on-field practices [5••, 28••]), such as reductions in pesticides, fertilizers or mowing operations. These measures aim at reducing land-use intensity within fields, and thus increase the ecological contrast with those fields that are farmed conventionally.
In addition, numerous studies emphasize the need for halting agricultural intensification impacts at both local (field) and landscape scales [10, 14, 16, 19, 21, 29, 30•]. For instance, richness of wild insects and pollination levels have been found to increase most with off-field practices aimed at increasing landscape heterogeneity, such as wildflower planting or restoring semi-natural habitats [28••]. These measures improved other ecosystem services as well (e.g. pest control, soil erosion mitigation, as well as scenic and cultural values). Scheper et al. [15•] showed that sown flower strips were the most effective conservation measure for pollinators, and that their effectiveness was mostly driven by the ecological contrast in floral resources created by their application. On-field practices like reduced pesticide application and mechanical operations, however, have also been found to increase wildflower plant richness [28••]. The results of Clough et al. [31] in arable-dominated landscapes surrounding semi-natural grasslands also emphasize that landscape management measures tailored to pollinators and the wild plants that they pollinate are both required. Hence, AES design should include measures to cope with the ecological needs of target groups at landscape scales as well (e.g. [32–34]; Table 1, Fig. 1), especially for groups or species that require different resources over their life cycles, require mosaic-like rather than continuous habitats, or have large home ranges [19, 21, 24]. In fact, many AES incorporate management prescriptions potentially useful to create mosaics or uncultivated refuges for species that perceive landscapes at higher scales than fields or farms, such as conservation headlands, pulse crops or buffer strips [5••, 32]. Reaching landscape-scale goals may be difficult, however, as it would require some coordination between AES management on multiple different farms, unless average farms where measures are applied were large enough to influence the landscape structure. Likewise, some of these off-field practices have been incorporated as compulsory greening measures in the last CAP reform. Insufficient uptake levels, even for supposedly compulsory measures, may, however, severely limit their effectiveness [6, 7••].
Considering Landscape-Scale Constraints Under a Functional Approach
Even if properly designed to enhance both within- and around-field requirements of target species, conservation measures may still fail to deliver significant ecological benefits due to landscape-scale and regional processes limiting effectiveness [10, 17–19]. Effectiveness (i.e. the difference in the of biodiversity or ecosystem service target measurements between conventionally farmed areas and areas that incorporate conservation measures) is expected to reach its maximum at intermediate landscape complexity levels, a pattern known as the ‘landscape-moderated effectiveness hypothesis’ [19] or the ‘intermediate landscape complexity hypothesis’ [29]. This hypothesis has been derived from the theoretical nonlinear relationships between landscape complexity and field-scale species richness, which interact with local management effects [10, 18]. Species richness at field scales generally increases from simple to complex landscapes because more diverse resources and dispersal corridors allow more species to coexist. Nevertheless, a minimum amount of unfarmed habitat and corridors would be necessary to maintain the presence of many species inhabiting fields. At the other extreme, a saturation point of landscape complexity would be reached, from where no further increases in species richness are expected (Fig. 2a). Such saturation may be due to full occupancy of fields by species coming from the species-rich surrounding landscape, or to potential negative effects of unfarmed habitats on open-country organisms inhabiting farmland patches [17, 18]. In any case, effectiveness (measured by some metrics, such as species richness) is expected to become null in both the simpler and the more complex landscapes, so that landscape complexity constrains, rather than only moderates, AES effectiveness (Fig. 2b). This non-linear model has been proven to be true in European agricultural land, both by means of experimental studies [10] and through meta-analyses [5••, 16]. It is based on the need for a minimum amount of semi-natural habitat around fields to support species within fields (lower threshold) and on an equilibrium distribution of the whole regional species pool over fields in the other extreme of the gradient (upper threshold at high complexity levels [17, 18]). The model, although developed and tested for species richness, may be potentially applied to any ecological variable showing a sigmoidal relationship with landscape traits (i.e. minimum requirements and saturation points where no further increases are possible due to space or resource constraints). Obvious relevant candidates are abundance of groups or target species, and most ecosystem services provided by farmland, although no evidence that such variables follow such a sigmoidal pattern has yet been found. This lack of evidence seems to be due to the large amounts of high-quality data required to test these non-linear models (e.g. [10]), although precise constraints may differ in practice for measures that are more sensitive to environmental variation than species richness. Explicit modeling approaches rather than meta-analyses are required to test the general applicability of models including thresholds [35].

Conceptual models depicting the consequences of non-linear relationships between landscape complexity (a and b) and land-use intensity (c and d) and field-scale biodiversity for the potential ecological effectiveness of agri-environment schemes (AES). Arrows near axes in c indicate the decrease in intensity achieved by the application of AES (X-axis) and the consequences of these intensity changes on field-scale biodiversity (Y-axis), in systems with low (light grey) and high (dark grey) overall levels of land-use intensity. Note that in the case of landscape complexity (a and b), the X-axes are the same, whereas for land-use intensity (c and d), the X-axes are different and in d indicate the intensity contrast (i.e. the reduction in land-use intensity accomplished by the application of AES) along hypothetical situations of increasing overall levels of land-use intensity, indicated by decreasing line width and dashing. After [9] and [18]
Constraints on biodiversity responses due to non-linear effects of landscape complexity may interact with non-linear effects of land-use intensity [9], also reflecting from ecological theory (‘intermediate disturbance hypothesis’, which predicts peak diversity at intermediate rather than at undisturbed or heavily disturbed fields [9, 12] (Fig. 2c). Because of these non-linear relationships between field-scale intensity and biodiversity, the ecological contrast between conventionally farmed fields and fields that incorporate conservation measures will have much larger effects in extensive than in intensive systems (Fig. 2d).
Kleijn et al. [19] considered that the combination of both non-linear effects will still produce landscape moderation, i.e., higher effectiveness at intermediate landscapes and lower, but not zero effectiveness at the simpler and more complex landscapes. Effectiveness at simpler landscapes will, hence, be enhanced by means of measures providing high ecological contrasts at field scales, somewhat compensating for the negative effects of landscape-scale intensity on local diversity [15•, 16, 19]. Landscape moderation would, however, turn into landscape constraint in the simpler and more complex landscapes if upper and lower thresholds of landscape complexity on field-scale diversity are taken into account (Fig. 2). In both simpler and complex landscapes, complexity will constrain AES effectiveness to zero irrespective of ecological contrast. Increasing contrast will only increase maximum effectiveness between complexity thresholds at intermediate landscape complexity levels, either symmetrically, if there is no interaction between landscape complexity and field-scale intensity (Fig. 3a, b), or biased towards more complex landscapes, if increased landscape complexity is correlated with decreased land-use intensity at the field scale (Fig. 3c, d; [36]). Consequently, increased landscape complexity, besides favoring biodiversity [10, 21, 30•], will also enhance the effectiveness of measures aimed at reducing within-field land-use intensity, but only within landscape complexity thresholds [10, 18]. Overall, the combination of two key non-linear relationships between land-intensity at landscape and field scales generates highly variable expectations for AES effectiveness, a fact that may explain the highly unpredictable, mixed results of most effectiveness evaluations [5••, 8]. A further consequence of non-linearity is that the range of intensities of the system under analysis may uncover only part of the full intensity gradient, and this may bias results of intensity effectiveness evaluations [10, 18]. This fact requires explicit modeling approaches to test the general applicability of the model, as meta-analysis results may be biased by this property of non-linear relationships.

Hypothetical combination of the effects of landscape complexity and AES-driven intensity contrast on the effectiveness of AES along gradients of landscape complexity, based on models shown in Fig. 2. The models a and b assume constant effects of ecological contrasts along gradients (i.e. no interaction among effects of landscape complexity and land-use intensity on field-scale biodiversity), whereas the models c and d try to incorporate non-linear effects of land-use intensity by assuming decreasing intensity, and, hence, increasing effectiveness for the same contrast, with increasing landscape complexity. Subsequently, maximum AES effectiveness would be reached at higher levels of landscape complexity when considering such potential interactions between landscape complexity and field-scale land-use intensity (c and d). Dashing in a and c and the thin line in b and d correspond to panels a and b in Fig. 2, and increased line widths indicate increased intensity contrast (reduction in land-use intensity accomplished by the application of AES)
Field-scale land-use intensity can be estimated by means of several alternative mechanisms [e.g. 10, 37], all clearly linked to ecological mechanisms influencing local abundance or diversity (e.g. resource availability or disturbance regime) and usually tightly correlated [9]. Landscape complexity, however, is much more elusive to measure [19]. Complexity, as defined by Concepción et al. [18], refers to the amount and distribution of distinct landscape elements in relation to three main landscape gradients of functional significance for organisms inhabiting agricultural landscapes: field traits (size, shape, boundaries), landscape composition [of land uses, including crop types and (semi-)natural habitats], and landscape connectivity [boundaries of (semi-)natural vegetation]. This functional approach implies that complexity will depend on how organisms perceive their immediate surroundings and how large are the spatial and temporal scales at which they perceive the landscape [38]. Most likely, organisms of different size, dispersal ability and life-span will perceive the same landscape at different scales, so that it may appear complex for large organisms with a long life-span and high dispersal abilities (e.g. large vertebrates) and simple for small and/or sessile organisms (e.g. plants or ground-dwelling arthropods [18]).
Landscape perception may also vary within groups according to habitat requirements or geographical origin [e.g. 24, 34]. In fact, recent reviews on how landscape traits may influence organisms’ distributions in agricultural landscapes emphasize that landscape variables used to measure landscape composition, structure, connectivity and, ultimately, complexity [18] should be selected according to species’ requirements and scales of perception, to reflect the functional consequences of landscapes for organisms [20]. Following this approach, characteristics of regional pools of target species are expected to influence threshold values of landscape complexity on the effectiveness of CAP greening measures. Consideration of regional influences on effectiveness becomes even more compelling when considering multiple taxa, farmland and crop types [10, 14, 30•], as well as response variables other than species richness and abundance, such as species composition, population trends, and functional and ecological traits [16, 19, 25, 30•, 31]. If prescriptions are not matched to perception and requirements of local species pools, conservation measures may be ineffective even if they have proven effective in other sites or landscapes (Table 1, Fig. 1).
A way of dealing with non-linear relationships is to divide them into categories based on threshold points. For instance, Batáry et al. [14] and Kleijn et al. [19] differentiated cleared (i.e. extremely simple) from simple and complex landscapes in their reviews and meta-analyses of the landscape moderation hypothesis using a threshold value of 20 % cover of natural vegetation (mostly woody) in the landscape. This value was inspired by Andrén’s [39] highly influential paper on the effects of habitat fragmentation on animal communities, which was, in turn, derived from percolation theory as the threshold value for cover of the original habitat under which isolation of remaining habitat patches increases exponentially [39]. This theoretical setting will be useful, and, in fact, it was, for analyzing responses of forest and woodland organisms to changes in landscape complexity, assuming that complexity is correlated with the amount of woody vegetation in the landscape. This assumption may, however, fail to predict equivalent threshold values differentiating the simple and complex landscapes of Concepción et al.’s [18] model (Fig. 2). Such a complexity threshold will not be predicted to arise for forest organisms, since an increased proportion of woody habitat in the landscape will only favor them (i.e. no evident saturation point would be reached). However, following the same approach, a threshold for organisms linked to the open matrix will arise when cover of the original uncultivated woody habitat will reach 80 % of the landscape. This will mean that the proportion of open habitats, either cultivated or not, will be below a 20 % threshold of connectivity. At least for birds, farmland communities are dominated by ecotonic and forest species in most of Europe [e.g. 40], but by open-land, steppic species in the southwest and southeast [24, 41]. Measures aimed at increasing complexity in north and central European farmland usually are (rightly) based on the recovery of natural woody vegetation [e.g. 42], but these same measures are known to be harmful in other regions [43–45], since they do not contemplate the landscape-scale requirements of their regional species pools. These results have not yet influenced meta-analyses, probably because data for these regions are still scarce as compared to the rest of Europe [5••], but must be taken into account for further improvement of the effectiveness of CAP greening in terms of increased target diversity and/or ecosystem services.
Adapting Policy Instruments to Regional Landscape Constraints
Non-linearity of key factors influencing farmland biodiversity and particularities of regional species pools and target traits make effectiveness of CAP greening measures hard to predict. Differences in landscape complexity perception among target groups further increase both unpredictability and the design of AES aimed at enhancing multi-group diversity. For this reason, ex ante evaluations of potential effectiveness [e.g. 22, 32] are unlikely to be reliable except at a quite crude level. Guidelines for adapting policy instruments to this reality should then consider the key factors driving major changes in expected effectiveness: target-defined requirements and landscape constraints (Table 1, Fig. 1).
Once the regional stage is set, criteria for the application of available, or newly developed, policy instruments should be guided by target-defined landscape constraints. In simpler/cleared agricultural landscapes, compulsory measures aimed at recovering complexity levels should be the top priority. Note that ‘cleared’ would mean ‘with low cover of natural vegetation’ if targets are woodland or ecotonic organisms, but ‘with low cover of open fields’ if targets are linked to open land or steppes. Landscape extensification measures could be implemented in these landscapes by means of compulsory greening measures [7••, 46] or wide entry-level schemes (i.e. measures accessible to many farmers with a few simple prescriptions [47]). Landscapes with intermediate levels of complexity, either current or achieved by means of compulsory measures, are the ones in which to apply AES-based extensification measures, designed according to the targeted species, communities or ecosystem services on the basis of their requirements in terms of minimum ecological contrasts needed. In complex landscapes with high biodiversity levels, extensification measures would be ineffective for further increasing field-scale diversity ([10, 18]; Figs. 2, 3). Here, measures that support farmers to maintain already existing extensive land-use practices should be implemented through, for example, subsidized high–nature value farmland (HNVF) programs [48], or regional-scale conservation agreements [e.g. 49]. The goal here should be to prevent abandonment or change to financially more attractive land uses, such as afforestation or perennial crops, which may reduce landscape complexity at least for open-land farmland organisms [e.g. 43, 50].
Systematic Evaluation and Learning Process
Interactions between complex and varying organism responses to changes in land use, and the policies developed to preserve them by influencing land-use change, add further levels of uncertainty to the task of improving the conservation goals of CAP reforms. This goal may even approach the category of wicked problems, that is, problems difficult or impossible to solve because of incomplete, contradictory and changing requirements that are often difficult to recognize [51, 52]. Apart from trying to disentangle problem complexity by indentifying the main goals and key points generating wickedness [52], a way of improving effectiveness is to systematically incorporate feeding-up results of evaluations into the successive reform rounds of CAP greening following principles of adaptive learning processes (Table 1, Fig. 1; [53, 54]). Systematic, direct evaluations have been claimed from the first seminal papers that showed large variance in AES effectiveness [8, 12], even proposing robust evaluation designs based on well-known paired or BACI (before-after-control-impact) experimental designs [8, 19]. Unfortunately, knowledge accumulated through non-systematic evaluations carried out in a handful of countries does not seem to have generated more effective measures yet [5••], a fact that may be partly due to the long duration of schemes (5–10 years; [5••]). Feedback from these studies, as well as systematic evaluations for the systems, regions and targets still barely covered in the literature, are urgently needed to ensure that common European agriculture and the policies supporting it meet their goal to be wildlife-friendly.
Conclusions
Measures aiming at enhancing and maintaining the complexity of agricultural landscapes will not only favor biodiversity, but also enhance the effectiveness of measures aimed at reducing within-field land-use intensity over most of the length of many landscape gradients ([10, 21, 30•]; Fig. 3). Landscape-scale thinking is, thus, essential to improve conservation in farmland, and specific management recommendations should be formulated regionally due to non-linear effects of landscape- and field-scale land-use intensity that vary among target species, groups and ecosystem services.
It is urgent to incorporate all these ideas arising from recent research into CAP greening efforts, since effectiveness has not been improved after the 2007 CAP reforms [5••] and lack of improvement was not due to relevant gaps in knowledge, as the relevant research and subsequent recommendations were available well before. Moreover, the reformed CAP for 2014–2020 is unlikely to perform better, as potential effects of the incorporation of compulsory AES-like measures to compensate for low take-up of voluntary AES is counteracted by low regional targeting and a high number of exemptions and reduced requirements that are permitted for the application of many environmental prescriptions [7••]. Accordingly, the current CAP is likely to continue driving agricultural intensification, as well as abandonment of less productive land. Guidelines developed here may help to explicitly incorporate current knowledge on how and why agricultural land-use intensity gradients modulate and constrain biodiversity and ecosystem services at field and landscape scales. Now, it is just time to apply this knowledge to policy design and evaluation.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance •• Of major importance
Foley JA, Ramankutty N, Brauman KA, Cassidy ES, Gerber JS, Johnston M, et al. Solutions for a cultivated planet. Nature. 2001;478:337–42.
Chaplin-Kramer R, Sharp RP, Mandle L, Sim S, Johnson J, Butnar I, et al. Spatial patterns of agricultural expansion determine impacts on biodiversity and carbon storage. PNAS. 2015;112:7402–7.
Queiroz C, Beilin R, Folke C, Lindborg R. Farmland abandonment: threat or opportunity for biodiversity conservation? A global review. Front Ecol Environ. 2014;12:288–96.
Mendenhall CD, Karp DS, Meyer CFJ, Hadly EA, Daily GC. Predicting biodiversity change and averting collapse in agricultural landscapes. Nature. 2014;509:213–7.
•• Batáry P, Dicks LV, Kleijn D, Sutherland WJ. The role of agri-environment schemes in conservation and environmental management. Conserv Biol. 2015;29:1006–16. - updated review on AES and available studies on AES effectiveness in Europe, with a meta-analysis of results, - updated information on history, goals, budget and prospects of AES-based policy.
Dicks LV, Hodge I, Randall N, Scharlemann JPW, Siriwardena GM, Smith HG, et al. A transparent process for ‘evidence-informed’ policy making. Conserv Lett. 2013;7:119–25.
•• Pe’er G, Dicks LV, Visconti P, Arlettaz R, Báldi A, Benton TG, et al. EU agricultural reform fails on biodiversity. Science. 2014;344:1090–2. - brief but at the same time exhaustive review of CAP shortcomings for biodiversity conservation.
Kleijn D, Baquero RA, Clough Y, Díaz M, De Esteban J, Fernández F, et al. Mixed biodiversity benefits of agri-environment schemes in five European countries. Ecol Lett. 2006;9:243–54.
Kleijn D, Kohler F, Báldi A, Batáry P, Concepción ED, Clough Y, et al. On the relationship between farmland biodiversity and land-use intensity in Europe. Proc R Soc B. 2009;276:903–9.
Concepción ED, Díaz M, Kleijn D, Báldi A, Batáry P, Clough Y, et al. Interactive effects of landscape context constrains the effectiveness of local agri-environmental management. J Appl Ecol. 2012;49:695–705.
Kleijn D, Berendse F, Smit R, Gilissen N. Agri-environment schemes do not effectively protect biodiversity in Dutch agricultural landscapes. Nature. 2001;413:723–5.
Kleijn D, Sutherland WJ. How effective are European agri-environment schemes in conserving and promoting biodiversity? J Appl Ecol. 2003;40:947–69.
Bengtsson J, Ahnström J, Weibull C. The effects of organic agriculture on biodiversity and abundance: a meta-analysis. J Appl Ecol. 2005;42:261–9.
Batáry P, Báldi A, Kleijn D, Tscharntke T. Landscape-moderated biodiversity effects of agri-environmental management: a meta-analysis. Proc R Soc B. 2011;278:1894–902.
• Scheper J, Holzschuh A, Kuussaari M, Potts SG, Rundlöf M, Smith HG, et al. Environmental factors driving the effectiveness of European agri-environmental measures in mitigating pollinator loss – a meta-analysis. Ecol Lett. 2013;16:912–20. - meta-analysis focused on the effectiveness of AES for halting the loss of pollinators, - emphasizes the need of aiming differentiation and subsequent AES design for the conservation of intrinsic (e.g., endangered species) and extrinsic (i.e., ecosystem services) biodiversity values.
Tuck SL, Winqvist C, Mota F, Ahnström J, Turnbull LA, Bengtsson J. Land-use intensity and the effects of organic farming on biodiversity: a hierarchical meta-analysis. J Appl Ecol. 2014;51:746–55.
Tscharntke T, Klein AM, Kruess A, Steffan‐Dewenter I, Thies C. Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecol Lett. 2005;8:857–74.
Concepción ED, Díaz M, Baquero R. Effects of landscape complexity on the ecological effectiveness of agri-environment schemes. Landsc Ecol. 2008;23:135–48.
Kleijn D, Rundlöf M, Scheper J, Smith HG, Tscharntke T. Does conservation on farmland contribute to halting the biodiversity decline? Trends Ecol Evol. 2011;26:474–81.
Fahrig L, Baudry J, Brotons L, Burel FG, Crist TO, Fuller RJ, et al. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett. 2011;14:101–12.
Gabriel D, Sait SM, Hodgson JA, Schmutz U, Kunin WE, Benton TG. Scale matters: the impact of organic farming on biodiversity at different spatial scales. Ecol Lett. 2010;13:858–69.
Primdahl J, Vesterager JP, Finn JA, Vlahos G, Kristensen L, Vejre H. Current use of impact models for agri-environment schemes and potential for improvements of policy design and assessment. J Environ Manag. 2010;91:1245–54.
Pywell RF, Heard MS, Bradbury RB, Hinsley S, Nowakowski M, Walker KJ, et al. Wildlife-friendly farming benefits rare birds, bees and plants. Biol Lett. 2012;8:772–5.
Concepción ED, Díaz M. Field, landscape and regional effects of farmland management on specialist open-land birds: does body size matter? Agric Ecosyst Environ. 2011;142:303–10.
Filippi-Codaccioni O, Devictor V, Bas Y, Julliard R. Toward more concern for specialisation and less for species diversity in conserving farmland biodiversity. Biol Conserv. 2010;143:1493–500.
Guerrero I, Carmona CP, Morales MB, Oñate JJ, Peco B. Non-linear responses of functional diversity and redundancy to agricultural intensification at the field scale in Mediterranean arable plant communities. Agric Ecosyst Environ. 2014;195:36–43.
Concepción ED, Moretti M, Altermatt F, Nobis MP, Obrist MK. Impacts of urbanisation on biodiversity: the role of species mobility, degree of specialisation and spatial scale. Oikos. 2015. doi:10.1111/oik.02166.
•• Garibaldi LA, Carvalheiro LG, Leonhardt SD, Aizen MA, Blaauw BR, Isaacs R, et al. From research to action: practices to enhance crop yield through wild pollinators. Front Ecol Environ. 2014;12:439–47. - exhaustive review on conservation measures that have proven effective for enhancing wild pollinators, also considering other ecosystem services.
Tscharntke T, Tylianakis JM, Rand T, Didham RK, Fahrig L, Batáry P, et al. Landscape moderation of biodiversity patterns and processes - eight hypotheses. Biol Rev. 2012;87:661–85.
• Gonthier DJ, Ennis KK, Farinas S, Hsieh H-Y, Iverson AL, Batáry P, et al. Biodiversity conservation in agriculture requires a multi-scale approach. Proc R Soc B. 2014;281:20141358. - review of studies comparing local management vs. landscape effects on biodiversity for a variety of taxonomic groups. It highlights the need of adopting conservation initiatives at local and landscape scales, especially in cases of multi-species targets.
Clough Y, Ekroos J, Báldi A, Batáry P, Bommarco R, Gross N, et al. Density of insect-pollinated grassland plants decreases with increasing surrounding land-use intensity. Ecol Lett. 2014;17:1168–77.
Llusia D, Oñate JJ. Are the conservation requirements of pseudo-steppe birds adequately covered by Spanish agri-environmental schemes? An ex-ante assessment. Ardeola. 2005;52:31–42.
Siriwardena GM, Calbrade NA, Vickery JA. Farmland birds and late winter food: does seed supply fail to meet demand? Ibis. 2008;150:585–95.
Concepción ED, Díaz M. Relative effects of field- and landscape-scale intensification on farmland bird diversity in Mediterranean dry cereal croplands. Asp Appl Biol. 2010;100:245–52.
Swift TL, Hannon SJ. Critical thresholds associated with habitat loss: a review of the concepts, evidence, and applications. Biol Rev. 2010;85:35–53.
Benton TG, Vickery JA, Wilson JD. Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol. 2003;18:182–8.
Herzog F, Steiner B, Bailey D, Baudry J, Billeter R, Bukácek R, et al. Assessing the intensity of temperate European agriculture at the landscape scale. Eur J Agron. 2006;24:165–81.
Holling CS. Cross-scale morphology, geometry, and dynamics of ecosystems. Ecol Monogr. 1992;62:447–502.
Andrén H. Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos. 1994;71:355–66.
Covas R, Blondel J. Biogeography and history of the Mediterranean bird fauna. Ibis. 1998;140:395–407.
Tryjanowski P, Hartel T, Báldi A, Szymański P, Tobolka M, Herzon I, et al. Conservation of farmland birds faces different challenges in Western and Central-Eastern Europe. Acta Ornithol. 2011;46:1–12.
Batáry P, Matthiesen T, Tscharntke T. Landscape-moderated importance of hedges in conserving farmland bird diversity of organic vs. conventional croplands and grasslands. Biol Conserv. 2010;143:2020–7.
Díaz M, Carbonell R, Santos T, Tellería JL. Breeding bird communities in pine plantations of the Spanish plateaux: biogeography, landscape and vegetation effects. J Appl Ecol. 1998;35:562–74.
Caparrós A, Jacquemont F. Conflicts between biodiversity and carbon sequestration programs: economic and legal implications. Ecol Econ. 2003;46:143–57.
Reino L, Beja P, Osborne PE, Morgado R, Fabião A, Rotenberry JT. Distance to edges, edge contrast and landscape fragmentation: interactions affecting farmland birds around forest plantations. Biol Conserv. 2009;142:824–38.
Aviron S, Nitsch H, Jeanneret P, Buholzer S, Luka H, Pfiffner L, et al. Ecological cross compliance promotes farmland biodiversity in Switzerland. Front Ecol Environ. 2009;7:247–52.
Evans AD, Green RE. An example of a two-tiered agri-environment scheme designed to deliver effectively the ecological requirements of both localised and widespread bird species in England. J Ornithol. 2007;148:S279–86.
Ribeiro PF, Santos JL, Bugalho MN, Santana J, Reino L, Beja P, et al. Modelling farming system dynamics in high nature value farmland under policy change. Agric Ecosyst Environ. 2014;183:138–44.
Campos P, Huntsinger L, Oviedo JL, Díaz M, Starrs P, Standiford RB, et al., editors. Mediterranean oak woodland working landscapes: Dehesas of Spain and Ranchlands of California. New York: Springer; 2013.
Cardador L, De Cáceres M, Giralt D, Bota G, Aquilué N, Arroyo B, et al. Tools for exploring habitat suitability for steppe birds under land use change scenarios. Agric Ecosyst Environ. 2015;200:119–25.
Hancock D. Tame problems and wicked messes: choosing between management and leadership options. RMA J. 2004; 38–42.
Chapin III FS, Trainor SF, Huntington O, Lovecraft AL, Zavaleta E, Natcher DC, et al. Increasing wildfire in Alaska’s boreal forest: pathways to potential solutions of a wicked problem. Bioscience. 2008;58:531–40.
Pahl-Wostl C. Requirements for adaptive water management. In: Pahl-Wostl C, Kabat P, Möltgen J, editors. Adaptive and integrated water management. Berlin: Springer; 2008. p. 1–22.
Roling NG, Wagemakers MAE. Facilitating sustainable agriculture: participatory learning and adaptive management in times of environmental uncertainty. Cambridge: Cambridge Univ Press; 2000.
Acknowledgments
We would like to acknowledge the invitation by Gavin Siriwardena and Leonore Fahrig to start this review, as well as suggestions by Dr. Siriwardena and two anonymous referees on a first version. This work is still a direct consequence of the creative environment generated by the EU’s EASY project lead by David Kleijn in 2003–2006 (QLK5-CT-2002-01495). Interactions with Spanish and Portuguese colleagues along the last 10 years, especially Ana Carricondo, Irene Guerrero, Manuel Morales, Juan Oñate, Lluis Brotons, Pablo Campos, Gerardo Moreno and Francisco Moreira, have improved a great deal our views on the topic. This paper is a contribution to the Spanish projects REMEDINAL3-CM (S2013/MAE-2719) and GANGA, and to the European BiodivERsA project BIOGEA (testing BIOdiversity Gain of European Agriculture with CAP greening).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Human and Animal Rights and Informed Consent
This article contains no studies with human or animal subjects performed by the authors.
Additional information
This article is part of the topical collection on Landscape Design and Planning for Ecological Outcomes
Rights and permissions
About this article

Cite this article
Díaz, M., Concepción, E.D. Enhancing the Effectiveness of CAP Greening as a Conservation Tool: a Plea for Regional Targeting Considering Landscape Constraints. Curr Landscape Ecol Rep 1, 168–177 (2016). https://doi.org/10.1007/s40823-016-0017-6
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40823-016-0017-6