
Abstract
During the Holocene colonization of islands in the Pacific and Caribbean by agropastoral and horticulturalist communities, a variety of proxies (material, genetic, zoogeographic etc.) indicate substantial inter-island and inter-community contact. It has been suggested that this contact represents an adaptive response to mitigate intrinsic demographic fragility during the initial phases of island colonization, and that this exogamous imperative faded in the aftermath of initial dispersal as overall population density increased. Here, we evaluate this model by synthesizing and comparing increasingly available 87Sr/86Sr data from funerary populations in the Pacific and Caribbean. After performing basic hygienic discrimination, we conclude that the Caribbean data—both from across the region, and on an intra-site/island basis—lend support to this model, with high early in-migration succeeded by comparatively low in-migration in a relationship which, based on chi-square tests, is statistically significant (1, N = 290) = 4.046, p = 0.044). The Pacific situation is more complex, with data from the Bismarck Archipelago and northern Solomon Islands skewing the analysis. However, in considering these data in detail, we demonstrate that there are reasons to suppose that the Pacific data may also tentatively support a model of high rates of in-migration being replaced later in settlement histories by comparatively low rates. We conclude by highlighting future directions for this incipient research program.
Similar content being viewed by others


Introduction
Agropastoral and horticulturalist populations expanded from a number of mid-low latitude centers of initial domestication during the Holocene. These expanding populations were subject to similar trajectories of comparatively exaggerated demographic growth (e.g., Bocquet-Appel 2002) and clearly maintained preference for certain types of environmental organization (Diamond and Bellwood 2003; Isern et al. 2017). Because of these structural socioecological constraints, we might reasonably suppose the spatial and temporal dynamics of this expansion to be comparable. Certain types of environment in particular should be expected to parameterize Neolithic sociospatial dynamics to an unusual extent. Oceanic islands, organized into subduction arcs, hotspot-chains, or isolates, represent a very distinct type of environmental heterogeneity that imposes terrestrial biological constraints, especially in terms of niche size and intervening distance (Rybicki and Hanski 2013; Borregaard et al. 2016); in essence, islands represent the most extreme exemplification of “patchiness” on the surface of the geosphere. Accordingly, contexts such as the insular Pacific and Caribbean may represent likely cases in which large-scale and long-term post-initial settlement trajectories parallel—or even potentially converge upon—one another.
One type of constraint militating in favor of parallel socioecological development relates to how populations act cross-culturally to mitigate the effects of demographic stochasticity—the greater probability in smaller as opposed to larger populations that stochastic demographic perturbations will cause the population to fluctuate to zero (e.g., Demetrius et al. 2004; Lande 1993; Moore 2001). Considering the scale and organization of originating communities, colonizing agropastoral and horticulturalist populations in Pacific and Caribbean island contexts would have been comparatively small and highly fragmented, and consequently exposed to exaggerated threat of localized unviability and/or extinction (cf. McArthur et al. 1976; Richter-Dyn and Goel 1972). It has been proposed that a primary mitigating strategy would involve artificially increasing local population size by maintaining high levels of genetic flow between dispersed communities, essentially boosting overall metapopulation connectivity in the immediate aftermath of settlement (Leppard 2015). A corollary effect of this would be decreasing metapopulation connectivity over time-elapsed since initial colonization as discrete populations grew towards robustness. Relaxation of the imperative to drive up metapopulation resilience would result in a general decline of post-juvenile residence mobility. This would in turn have clear social consequences (not least relating to language evolution, dynamism in material-cultural traditions and/or “styles,” and sociopolitical organization) which—if shown to be emergent outcomes of initial ontogenetic environmental-demographic conditions—would have profound implications for our confidence in modeling general macro-scale processes in human social behavior over the long-term.
Because of the implications of this model, it is vital to establish the degree to which it is accurate. We can, then, use the predictive capacity of the model to generate a testable hypothesis:
Over time-elapsed since initial colonization/arrival of agropastoral and horticulturalist subsistence behaviors (assuming demographic growth is grossly a function of time), post-juvenile residence mobility will decline.
Such that the null hypothesis:
Over time-elapsed since initial colonization/arrival of agropastoral and horticulturalist subsistence behaviors (assuming demographic growth is grossly a function of time), there will be an increase/no discernible change in post-juvenile residence mobility.
We can test this hypothesis against the extant corpus of human 87Sr/86Sr data from funerary populations in the Pacific and Caribbean, and proceed to do this subsequently.
How do we facilitate and undertake this comparison? The model assumes the key variable to be time-elapsed since colonization, with growth understood to be a function of time-elapsed. This has two consequences. Firstly, we are interested less in calendar date of initial settlement and more in comparably deep (or shallow) post-colonization histories. Accordingly, we restrict ourselves here to 87Sr/86Sr data from funerary populations drawn from the initial, quasi-instantaneous (in radiocarbon terms). Agropastoral and horticulturalist settlement episodes of the Pacific and Caribbean, the Lapita and Saladoid phenomena respectively (see now Rieth and Athens 2017). Secondly, as published studies of human 87Sr/86Sr ratios cover a temporal continuum in both case studies, it is necessary to artificially group them, such that we can demarcate gross general temporal trends (i.e., decreasing or increasing post-juvenile residence mobility). Accordingly, we divide the settlement histories of both areas into artificial categories representing earlier in respective colonization histories (Lapita: 3200–2000 cal BP/Saladoid 2300–1200 cal BP) and later (post-Lapita: post-2000 cal BP/post-Saladoid: post-1200 cal BP), bearing in mind these chronological divisions are themselves estimates subject to margins of error.
We note two relevant caveats. Firstly, we are assuming that demographic growth under conditions of agropastoral and horticulturalist subsistence (which we define broadly, to encompass majority food-producing rather than acquiring groups) is essentially a function of time. At large spatial and temporal scales, this is demonstrably the case. However, at finer scales of analysis (i.e., sub-millennial), it may not be, especially considering cross-cultural evidence for “boom-and-bust” dynamics during initial and potentially pre-, agropastoral, and horticulturalist settlements (Timpson et al. 2014; Goldberg et al. 2016). This complicates our analysis, and we return to this in our discussion below, considering our results in the context of (a) general growth models and (b) initial r-type growth followed by collapse and gradual recovery. At the very large (supra-millennial) temporal scale in we are working, however, and having subdivided our data into temporal absolutes (rather than continua), these fine-grained temporal dynamics should neither be observable nor, from the perspective of total demographic growth over time and corresponding behavioral changes, vitally relevant. In any case, gradual settlement “in-filling,” evident in both study areas following initial colonization, may lend further support to a model of demographic change in which the general trend is towards growth.
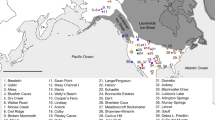
Secondly, it might be considered that the differences in spatial scale between our Pacific (Fig. 1) and Caribbean (Fig. 2) case studies complicate any attempt at comparing patterns in post-juvenile residence mobility, in that greater inter-site distance might be expected to impose some sort of structuring process or threshold on mobility generally. In response to this, we would highlight that the difference in scale between our Saladoid-area and Lapita-area case studies is probably within an order of magnitude, and that differences in range and speed of the relevant mobility technologies (i.e., Pacific sailed outrigger canoes vs. Caribbean paddled dugout canoes) either renders the Caribbean a more daunting space or the Pacific a more compact one, depending on perspective (e.g., Broodbank 2010). It is not clear to us that these differences are fatal for the process of comparison.

Map of the Pacific indicating the limit of Lapita expansion, and the location of islands and island groups discussed in the text

Map of the Caribbean indicating the limit of Saladoid expansion, and the location of islands and island groups discussed in the text
Data and methods
In order to test the proposed model, we gathered published strontium isotope data obtained from the analysis of archeological human dental enamel in the insular Pacific and Caribbean. Data mining aimed at inclusivity within the parameters of the model; accordingly, we have included all relevant agropastoral and horticulturalist datasets for both macro-regions. Datasets were excluded if the total number of samples per site was too small to conduct statistical analysis (n < 5), and if the chronological association of the analyzed population indicated that it did not correspond to the initial colonization period (i.e., post-settlement by agropastoral and horticulturalist or communities, but pre-Contact) for that location, or if the chronological association was unclear.
For the Pacific dataset, we chose to exclude the results from the site of Wairau Bar, South Island, New Zealand (Kinaston et al. 2013). This decision was based on the purported context as a likely “landfall site.” In comparing human 87Sr/86Sr data with the local baseline range obtained from dog (Canis lupus familiaris) from this site, the authors estimate that as many as 20 of the 24 (83.3%) humans were nonlocal. This proportion of nonlocals is, as far as we are aware, much higher than that reported for any other population from either the Pacific or the Caribbean, or, indeed, most other areas. Although the dating of Wairau Bar places it (late) within the parameters of our discussion, the disproportionate number of nonlocals relative to the other sample populations within the prehistoric Pacific may well indicate, as the authors suggest, that the site represents one of, or the, founder settlement(s) on South Island. As our interest is in inter-site migratory dynamics in the aftermath of colonization, not during initial colonization itself, including the Wairau Bar data would be erroneous. Additionally, Sr data from the site of Nebira on the southern coast of Papua New Guinea (Shaw et al. 2011) were also excluded. New Guinea not only has a much longer and more complicated demographic history—having been settled during the Late Pleistocene when it formed the northern spur of the paleo-continent Sahul, which now has settlement dates of ~ 65 kya (Clarkson et al. 2017)—it is also orders of magnitude larger than other islands in our Pacific sample, likely rendering it noncomparable in terms of spatial and demographic dynamics. Lastly and conversely, we should note that we have included sample populations from the Anir Islands, Tanga Islands, and Watom Island in the Bismarck Archipelago (Shaw et al. 2009, 2010), and from Buka in the Solomons. These island groups are located within the limits of human colonization of near Oceania during the Pleistocene (as with New Guinea, to which they may or may not have been attached at sea level lowstands during the Last Glacial Maximum), but the sample populations ostensibly date to Lapita (Shaw et al. 2009, 2010). Owing to the ambiguity of these three cases (i.e., early horticulturalist sites embedded in contexts with deep nonhorticulturalist settlement histories), we have conducted the calculations comparing the proportions of nonlocals from the Early and Late phase of the Neolithic Pacific colonization both including (complete) and excluding (“trimmed”) these datasets—we return to this topic below.
For the Caribbean dataset, we excluded populations that have been traditionally ascribed as “Archaic” or Archaic Age (including data from both Aruba and Cuba). These of course may be grossly contemporaneous with the other Ceramic Age populations in our study, but it is presently unclear if they actually represent cases of agropastoral and horticulturalist subsistence as defined above. Strontium isotope data derived from populations dating to the early contact (e.g., Valcárcel Rojas et al. 2011) and/or colonial period (e.g., Schroeder et al. 2009) were also excluded from our analysis. The final utilized datasets of archeological human 87Sr/86Sr results for both the Pacific and Caribbean macro-regions are listed in Table 1 and in the Appendix (supplementary table).
For the initial testing of the model, we have used the number and proportions of nonlocals for each population as presented in the original studies. It is important to note, however, that different studies have used (a) different approaches to estimate the local range of strontium isotope variation and (b) various methods for the identification of nonlocals. Among the datasets gathered, the most common approaches to outlier identification rely on some variant of the mean plus or minus two standard deviations from the mean. Some of the Pacific studies included herein (e.g., Jarić 2004; Shaw et al. 2009, 2010; Stantis et al. 2016) apply this method solely to the human 87Sr/86Sr data, while others (Phaff 2012; Fenner et al. 2015; Kinaston et al. 2013, 2014) apply it to comparative baseline or bioavailable 87Sr/86Sr data obtained from the analysis of associated (local) faunal and/or floral samples. For the Caribbean studies, most local range estimates rely on an iterative version of the mean ± 2σ method including both human and bioavailable data, or simply the absolute range of the associated bioavailable dataset. These differences in how nonlocals are identified and quantified represent a clear limitation of the current study, although an acceptable one from a perspective of inclusiveness and corresponding data richness. However, we have chosen to utilize the metrics adopted by each study, rather than recalculating based on our own criteria for two reasons: (a) the inherent variation in the analyzed sample types and materials (human data only, humans and archeological fauna, humans, and modern flora) vary widely between different studies making it difficult, if not impossible, to apply a single standard metric to all datasets; and (b) most of the studies for which the datasets are roughly comparable produce similar estimates of local ranges—and thereby identifications of which individuals are outliers (nonlocals)—regardless of which method is applied.
We have considered the option of applying a standard statistical method to all of the strontium isotope data sets as recently proposed by Lightfoot and O’Connell (2016) for oxygen isotope (δ18O) data. However, for archeological skeletal δ18O data, there is often little, if any, independent data for estimating local ranges of variation for a given site/location/area. This contrasts with strontium isotope studies, where generally several independent datasets (geological/bedrock, soils, fauna, flora) for estimating local ranges and thereby identifying nonlocals are often available. Additionally, there is an underlying assumption that intra-population variation in δ18O should be (approximately) normally distributed, which is an assumption that is likely generally valid. In contrast, bioavailable 87Sr/86Sr (including human Sr data), can be most simply thought of as the result of a two-end member mixing solution between different Sr sources: geological and nongeological (in island settings = seawater). Therefore, any ranges that fall near the end members (as opposed to near the middle of the mixing line) will not likely be normally distributed but rather should display a high degree of skewness even in the absence of nonlocals. For these reasons, we have chosen not to apply a standard statistical test for the identification of nonlocals to the various Sr isotope datasets.
Results
Results of the estimated number of analyzed individuals, and the number and proportion of nonlocals identified at each site, are presented in Table 2. A few patterns are evident from a simple perusal of this dataset. There is a high degree of variation both in the number of analyzed samples per population and in the absolute number and percentage of nonlocals. For several sites (n = 7), zero nonlocals were identified, while at only one site (Morel, Guadeloupe) were the majority (> 50%) of analyzed individuals identified as nonlocal. Comparison of the results between the Pacific and the Caribbean reveals several general trends. Most of the populations for which no nonlocals were identified (five out of seven) are in the Pacific dataset, there are as many studies in the Pacific reporting zero nonlocals as there are reporting any nonlocals, and only two populations in the Pacific are comprised of greater than 20% nonlocals. Conversely, for the Caribbean populations, there are not only fewer populations consisting of no nonlocals (n = 2) but also eight sample populations possess nonlocal proportions exceeding 20%. In terms of general comparisons between the Pacific and Caribbean datasets, the former possesses fewer nonlocals both in absolute and relative terms (n = 21, 12.4%) compared to the latter (n = 64, 22.1%).
We now turn to the chronological assessment of the nonlocal results vis-a-vis our proposed demographic model. The results of the overall number and percentage of nonlocals for the Pacific and Caribbean and for the Early and Late phase of each is presented in Table 3 and Fig. 3.

Percentage of individuals identified as having nonlocals origins for Early and Late contexts from the Pacific and Caribbean
For the total Pacific dataset, contrary to the proposed model there is not a decrease in the percentage of nonlocals between the early and late phase of the Neolithic when all data, including those from the Bismarcks and northern Solomons, are included in the analysis. In fact, there is a slightly higher percentage of nonlocals in the Late phase (13.7%) compared to the Early phase (10.3%). A chi-square test indicates that the difference in the proportions of nonlocals between the Early and Late Pacific data is not statistically significant; χ2 (1, N = 170) = 0.444, p = 0.505. A closer inspection of the site data reveals that a primary factor influencing this pattern is that four out of the six sample populations from the early Pacific have no nonlocals. Three of these are from the Bismarck Archipelago and one from the northern Solomon Islands, and all are located within or near the limits of Pleistocene colonization of Melanesia (Fig. 1). This potentially complicates our analysis (with regard to a more complex and deeper history of settlement than in Remote Oceania), which we consider in detail below. For the sake of comparison, we have recalculated the results with a trimmed dataset that excludes these four Early (Lapita) cases from offshore insular Papua New Guinea (PNG). When the circum-PNG data are excluded from the calculations for the Early Pacific, the proportion of nonlocals rises to 21.9%, a much higher percentage than for the Late Pacific sample set (13.7%). A chi-square test indicates that the difference in the proportions of nonlocals between the Early and Late Pacific for this trimmed dataset is also not statistically significant; χ2 (1, N = 134) = 1.224, p = 0.269.
In terms of chronological assessment of the Caribbean data, the results are clearer. For the Earlier component of the Caribbean, 28.6% of individuals have been identified as nonlocal, whereas in the Later component the proportion drops considerably to 18.4%. A chi-square test indicates that the difference in the proportions of nonlocals between the Early and Late Caribbean is statistically significant; χ2 (1, N = 290) = 4.046, p = 0.044. The results from the Caribbean conform to the proposed model in that a significant decrease in residential mobility, based on the relative proportions of individuals with nonlocal origins, is observed from the Early to the Late Ceramic Age. The implications of these results are further explored in the Discussion section.
In order to explore the patterns in the results at finer spatial scales, we also present the results of five case studies (two from the Pacific and five from the Caribbean) representing the individual islands or discrete island groups in the total datasets for which there are results from both an earlier and later phase of agropastoral/horticulturalist occupation. It is important to note that for the sake of this comparison the terms early and late do not necessarily correspond to the sites’ chronological placement in the broader chronological framework of the two macro-regions, but instead to relative earlier and later phase of that particular island’s (or island group’s) colonization history. For the Vanuatu example, the comparison is based on two sites on two different islands (Efate and Malakula) dating to the early Lapita and late Lapita periods respectively. For Fiji, the two sites are from the same island (Viti Levu) with one dating to an earlier and the other to a later post-Lapita occupation. For the Caribbean case studies, the Guadeloupe example is represented by one site dating primarily to the Early Ceramic Age and the other to a nearby Late Ceramic Age site. For both the St. Thomas and Puerto Rico samples, the chronological comparison is based on the splitting of the sampled skeletal populations into Early and Late categories based on associated remains and radiocarbon dates.
The results of these five smaller scale case studies are presented in Table 3 and displayed in Fig. 4. For all five cases, there is a decrease in the proportion of nonlocals from the earlier to the later phase of occupation. For some of the case studies, such as Fiji, the difference in the proportions of nonlocals from early (23.5%) to late (20.0%) is minor. For others, e.g. Guadeloupe (early = 57.1%; late = 26.4%) and Puerto Rico (early = 46.2%; late = 18.8%) the differences in the proportions of nonlocals are quite pronounced, with more than twice as many nonlocals in the early versus the late category. However, based on Fisher’s exact tests, none of the observed differences are statistically significant. This may partly reflect the much smaller sample sizes obtained by splitting the data into ever smaller groups.

Percentage of individuals identified as having nonlocals origins for Early and Late contexts from five island-scale case studies in the Pacific and Caribbean
Discussion
Overall, the results of our comparative analyses initially vary from the somewhat equivocal to the tentatively promising with regard to the proposed hypothesis of decreasing rates of post-juvenile residential mobility relative to time-elapsed since initial (agropastoral and horticulturalist) colonization. For the full Pacific dataset, the inverse pattern was observed with a slight increase in the proportions of nonlocals in the Late period. In contrast, for the trimmed Pacific dataset, the results are consistent with the proposed hypothesis, and a clear decrease in mobility was observed, although the difference in the relative proportions of nonlocals between Early and Late is not statistically significant. The results from the Caribbean most closely conformed to the proposed hypothesis, with a clear decrease in mobility rates and a statistically significant difference in nonlocal percentages between the Early and Late Ceramic Age. Shifting the scale of analysis from the macro-regional to individual islands or island groups, comparative analyses of mobility rates for five case studies consistently showed decreased rates of mobility over time-elapsed since initial colonization for each of them. None of the observed differences at these smaller spatial scales were statistically significant, although the greatly reduced sample sizes of the individual case studies may have influenced the lack of significance. In this section, we discuss several factors that may have influenced the results and explore the implications of the observed patterns in reference to the proposed model.
Methodological considerations
First, we consider the problem of false negatives. For the category nonlocal, in most cases, we can have a relatively high degree of confidence that an individual possessing an enamel strontium isotope value that is a statistical outlier is likely not locally born and thus must have migrated to the place of burial sometime after the age of dental crown formation and before final burial. However, the opposite cannot be said for the category of local individuals, which may contain a mix of actually locally born individuals and nonlocal migrants who originated from areas which are isotopically indistinguishable from the local range. In the absence of other relevant evidence, it is neither possible to determine if any actual migrants are present in the local category (as defined by their enamel Sr isotope signature) nor to quantify how many of these false negatives might be present. This is a problem inherent to strontium isotope studies of paleomobility generally, and it has important implications for both intra- and inter-population assessments of the absolute or relative representation of nonlocals (and thus inferred rates of paleomobility/migration).
A second, but potentially equally complicating variable is the intrinsic spatial variability of (bioavailable) strontium isotopes relative to the scales at which people migrate. More specifically, there is an enormous degree of variation in the spatial extent of local ranges. For example, for some small islands, the spatial extent of the local range may be coeval with the size of the island itself as, in essence, a single geological phenomenon. In fact, for some geologically homogenous areas, especially those underlain by relatively young marine sediments, the spatial extent of the local 87Sr/86Sr range may include not only an entire island but also an entire island group or even an archipelago and may extend up to hundreds of square kilometers. On the other hand, for geologically complex regions the spatial extent of the local 87Sr/86Sr range may be highly circumscribed to a relatively small area around the vicinity of the site. This problem is distinct from—but very much related to—that of false negatives, and it presents a potentially confounding factor for our model because it influences the distances that must be traveled for an individual to be identifiable as a nonlocal (immigrant) based on Sr isotope signals. In other words, in some contexts, individuals who have only migrated a few kilometers could potentially be identified as nonlocals, while in other contexts individuals would have to have migrated much further distances (tens or perhaps even hundreds of kilometers) to be identifiable as nonlocals based on the strontium isotope approach. As with false negatives, this is not a problem specific to our study but one that impinges upon strontium-based approached to paleomobility more generally.
Lastly, we should emphasize that other inherent geographical, geological, geochemical, and socio-technological factors may also add further complications to the testing of the model as proposed herein. The much larger distances and distinct seafaring technologies of the Pacific and Caribbean have already been mentioned. Perhaps, more relevant for the patterns revealed by our analyses is the variable distances between geologically (and by proxy isotopically) distinct regions and how these may or may not map onto the source and destination locales for prehistoric (Neolithic) migrants differently within and between the Pacific and the Caribbean. As with any such model, the inherent ambiguities of the archeological record can make it difficult to control for socio-economic, spatial, and temporal parameters. In terms of subsistence, for example, there is still considerable debate concerning the antiquity, origins, and spatio-temporal distribution of animal and plant domestication and cultivation, as well as human colonization histories, among Caribbean scholars (see now e.g., Giovas et al. 2016; Pagán-Jiménez et al. 2015). One important distinction between the Pacific and Caribbean Neolithic (or more specifically between the Lapita and Saladoid migrations and colonizations) is that while the former primarily colonized previously uninhabited islands, the latter settled on many islands that were already inhabited by so-called “Archaic” populations, the presence of which likely affected the organization of Saladoid settlement in the insular Caribbean, although which effects we cannot necessarily trace in the current analysis.
Given the considerable number and variation in types of complicating factors and caveats, it is perhaps somewhat surprising and potentially promising that two of the main macro-regional chronological comparisons (the trimmed Pacific and complete Caribbean datasets) and all five smaller spatial scale (island/island group) comparisons can be tentatively (but not unequivocally) taken to correspond to the demographic model of a correlation between time-elapsed since colonization and decreased mobility. Before concluding, and in so doing highlighting future avenues of research, we turn to two larger issues which impinge upon the significance (or lack thereof) of this observation: (a) the rationales for exclusion or inclusion of the Bismarck and northern Solomon data and (b) broader evidence for more complicated trajectories of demographic growth during incipient agropastoral and horticulturalist colonization than we assume here.
Broader complicating factors
As we have stressed throughout our discussion, four strontium datasets from sites in the circum-PNG archipelago substantially affect the Pacific analyses: Kamgot/Balbalankin, in the Anir Islands; Lifafaesing, in the Tanga Islands; Reber-Rakival, on Watom Island; and Sohano, on Buka in the northern Solomons (with all the former in the Bismarck Archipelago). With these sites—all of which purport to have a Lapita date, and consequently qualify as Early—included in the analysis, overall frequencies of nonlocals are reduced, the sites in question containing no nonlocals. With these sites excluded from the analysis (thus the “trimmed” Pacific dataset), local/nonlocal frequencies in Early vs. Late periods conform more substantially to the model, although this is not statistically significant in χ2 terms, being (1, N = 134) = 1.224, p = 0.269. How, if at all, can we justify this trimming?
The general settlement dynamics of Remote Oceania are moderately well-understood, being absent of human populations until approximately 3100–3000 bp, with Lapita representing the initial colonization horizon and subsequent forays into the uninhabited Pacific (i.e., into East Polynesia) undertaken by populations to whom Lapita colonists were, in part, ancestral. The situation in the Bismarcks and northern Solomons is, however, substantially more complex. Radiometric dates suggest that these island groups (which were either attached to, or only narrowly separated from, Sahul at the Last Glacial Maximum) were in part settled during the Pleistocene, with the colonization horizon for Sahul itself now pushed back to ~ 65 kya (Clarkson et al. 2017). We emphasize that we have no intention here of becoming involved in what has, at times, becomes a contentious debate surrounding the nature of the Lapita phenomenon, whose origins probably lie in the Taiwan-Philippines complex but whose movement from the Southeast Asian littoral to the borders of Near Oceania (in terms of its rapidity and demic organization) has been the subject of extensive discussion (now Cochrane 2017; Spriggs 2011). Suffice it to say here that, if the Bismarcks and Solomons possessed established pre-Lapita populations, then whatever the Lapita complex represents in demic terms (and here we recognize the important data from Teouma, which suggest little PNG genetic input; Skoglund et al. 2016) the overall robustness and organization of the metapopulations supporting individual sites, such as Sohano would be substantively and qualitatively different to the structure of metapopulations supporting individual sites in previously uninhabited Remote Oceania, such as Sigatoka. Put simply, we wonder whether nonlocals (in Sr terms) are absent from the Bismarck and Solomon datasets because a deep history of settlement allowed communities to draw exogamously on nearby populations within an isotopically indistinguishable area, in contrast to more dispersed communities in Remote Oceania having to rely on much more distant (and therefore more likely to be isotopically distinguishable) populations. In the current format, there is no space to evaluate whether or not this may be the case, but the striking divergence in terms of locals/nonlocals between Lapita sites on islands with Pleistocene settlement and Lapita sites on islands with no previous settlement leads us to suspect that differing metapopulation dynamics and mean post-juvenile migration distances may be at work. Accordingly, we prefer here the “trimmed” Pacific data as likely more accurately reflecting expansion of colonizing populations into uninhabited areas. Interestingly, the distribution of Archaic sites in the Caribbean has, as far as we can tell, little to no discernible effect on resulting frequencies of locals/nonlocals at Saladoid sites.
A second, and we think more problematic, from the perspective of our model, issue concerns the precise nature of demographic trajectories in the immediate aftermath of the establishment of agropastoral and horticulturalist settlement. As we have stressed, in working at large temporal and spatial scales, we are assuming demographic growth to be a function of time-elapsed, and patterns in the data appear in general to corroborate this (as regards decreasing mobility as an index of dispersed populations growing towards robustness). However, there is evidence to suggest that at smaller scales (decadal and centennial), post-colonization dynamics are more complicated. Timpson et al. (2014) chart immediate and rapid growth in the aftermath of the LBK colonization of central Europe, followed by a collapse and then a subsequent up-tick in growth. Goldberg et al. (2016), albeit working with pre-horticulturalist data, track similar boom-bust dynamics in South America. Translating these dynamics into the terms implied by Leppard (2015) involves supposing initially high rates of post-juvenile migration collapsing rapidly in the face of demographic booms, only to re-assert themselves and then erode slowly during bust and gradual recovery. We stress that we do not have enough data, refined to the appropriate degree, to evaluate this—with current data (and these deriving from areas where boom-bust dynamics have not necessarily been demonstrated)—we can only chart fairly gross overall trends. However, if there does exist a substantive and causal link between demographic robustness and degree of in-migration, then we would expect the strontium data to reflect this nuance. We look forward to testing a revised model, based on more complicated demographic trends, as more Sr data become available.
Conclusions
Based on arguments from first principles, we have suggested that, in highly fragmented island environments newly colonized by agropastoral and horticulturalist populations, the fragile demographic situation of these populations would have encouraged exaggerated rates of post-juvenile residence mobility within a context of exogamy; as populations grew towards robustness over time, this imperative would have relaxed. This should be testable against burgeoning strontium datasets from island contexts. We argue that, based on the strontium data, we have gathered for the initial settlement of the Pacific and Caribbean islands by food-producing communities, trends in the Sr data in general support this model, with some caveats that we have discussed above. As such, we propose several avenues for future research that would permit us to more robustly reject or confirm our main hypothesis.
First, a more standard and consistent method for the estimation of local ranges, and the subsequent identification and quantification of nonlocals should be developed. It is difficult to define a priori what an ideal method would look like, given the inherent variation in the nature of the various datasets investigated for this study. This variation includes some studies which totally lack any baseline Sr data to those that base their local range estimates on very different types of bioavailable datasets (e.g., fauna versus flora, modern versus archeological, etc.); nonetheless, a call for greater methodological coherence is worthwhile, as more Sr datasets become available and questions in which comparison is implicit become more pressing. Second, and related, larger and more geographically representative sample populations and datasets would permit for more statistically robust model testing and more fine-grained and nuanced exploration of variations in mobility patterns at multiple scales. Lastly, further development and refinement of spatial maps of strontium isotope variation (i.e., isoscapes; e.g., Nafplioti 2011) would allow for more explicit consideration of the extent to which the underlying isotopic variation explains the observed patterns relative to the variable that is of interest, namely past human migrations.
These recommendations notwithstanding, what are the implications of our initial attempt to evaluate a model of paleomobility as a function of demographic robustness? In general, the apparent trend from relatively high in-migration rates earlier during insular settlement histories to relatively low rates later in settlement histories suggests that we might indeed be justified in understanding the mechanics of inter-settlement residence mobility as an outcome of varying socio-demographic calculus. As we have emphasized, based on the size of the dataset, this is preliminary, and a first task must be the accumulation and compilation of more relevant data, but nonetheless this observation leads to further, substantive questions regarding sociopolitical organization. First, assuming we are witnessing institutionalized long-distance exogamy, should we understand this to be patrilocal or matrilocal (cf. Jordan et al. 2009)? If we consider division of labor and craft-production to be likely distinguished in terms of gender in the prehistoric Caribbean and Pacific, then long-distance exogamy on matri- or patrilocal principles has significant implications for how we understand large-scale cultural complexes such as Lapita; here we underscore, but make no attempt to resolve, the interpretive problem of Cedrosan versus Huecoid Saladoid traditions. Second, if we are able to trace dynamics in which a significant minority of a population is nonlocal and transitions over time to a much less significant minority, what are the likely outcomes in terms of property and land-tenure rights, and indeed the extent to which local/nonlocal status is ascribed and utilized in social interactions? Here this research nudges towards a consideration of emergent complexity, and the development of vertically exaggerated hierarchies in both the Pacific and Caribbean (Kirch 2010; Siegel 2010); what role does population robustness play in this emergence? Finally, we have addressed here island environments, as types of physiographic organization which are intrinsically heterogeneous and consequently likely to encourage or constrain dispersed modes of settlement during Neolithic colonization. However, other examples of agropastoral and horticulturalist colonization also exhibit clear preference for certain types of environment (for example, the preference of LBK settlement for loessial pedologies), driving comparably patchy dynamics. We propose that exploring mobility strategies via summed strontium datasets is likely to be a powerful tool in elucidating the nature and dynamics of agropastoral and horticulturalist expansion, not only in island contexts but more generally.
References
Bentley RA, Buckley HR, Spriggs M, Bedford S, Ottley CJ, Nowell GM, Macpherson CG, Pearson DG (2007) Lapita migrants in the Pacific’s oldest cemetery: isotopic analysis at Teouma, Vanuatu. Am Antiq 72(4):645–656
Bocquet-Appel JP (2002) Paleoanthropological traces of a Neolithic demographic transition. Curr Anthropol 43:637–650
Booden MA, Panhuysen RGAM, Hoogland ML, de Jong HN, Davies GR, Hofman CL (2008) Tracing human mobility with 87Sr/86Sr at Anse á la Gourde, Guadeloupe. In: Hofman CL, van Duijvenbode A (eds) Crossing the Borders: new methods and techniques in the study of archaeological materials from the Caribbean. University of Alabama Press, Tuscaloosa, pp 214–225
Borregaard MK, Amorim IR, Borges PAV, Cabral JS, Fernández-Palacios JM, Field R, Heaney LR, Kreft H, Matthews TJ, Olesen JM, Price J, Rigal F, Steinbauer MJ, Triantis KA, Valente L, Weigelt P, Whittaker RJ (2016) Oceanic island biogeography through the lens of the general dynamic model: assessment and prospect. Biol Rev 92(2):830–853
Broodbank C (2010) ‘Ships a-sail from over the rim of the sea’: voyaging, sailing and the making of Mediterranean societies c. 3500–800 bc. In: Anderson A, Barrett JH, Boyle KV (eds) The global origins and development of seafaring. McDonald Institute for Archaeological Research, Cambridge, pp 249–264
Clarkson C, Jacobs Z, Marwick B, Fullagar R, Wallis L, Smith M, Roberts RG, Hayes E, Lowe K, Carah X, Florin SA, McNeil J, Cox D, Arnold LJ, Hua Q, Huntley J, Brand HEA, Manne T, Fairbairn A, Shulmeister J, Lyle L, Salinas M, Page M, Connell K, Park G, Norman K, Murphy T, Pardoe C (2017) Human occupation of northern Australia by 65,000 years ago. Nature 547:306–310
Cochrane E (2017) The evolution of migration: the case of Lapita in the Southwest Pacific. J Archaeol Method Theory. https://doi.org/10.1007/s10816-017-9345-z
Demetrius L, Gundlach VM, Ochs G (2004) Complexity and demographic stability in population models. Theor Popul Biol 65:211–225
Diamond J, Bellwood P (2003) Farmers and their languages: the first expansions. Science 300:597–603
Fenner JN, Clark G, Cressey A, Valentin F, Olesen SH, Armstrong R (2015) Isotopic uniformity and segregation in Tongan mounds. J Archaeol Sci Rep 2:644–653
Giovas CM, Kamenov GD, Fitzpatrick SM, Krigbaum J (2016) Sr and Pb isotopic investigation of mammal introductions: pre-Columbian zoogeographic records from the Lesser Antilles, West Indies. J Archaeol Sci 69:39–53
Goldberg A, Mychajliw AM, Hadly EA (2016) Post-invasion demography of prehistoric humans in South America. Nature 532:232–235
Hofman CL, Hoogland MLP, Mickleburgh HL, Laffoon JE, Field MH, Weston DA (2012) Life and death at Lavoutte, Saint Lucia, Lesser Antilles, as revealed from rescue excavations. J Field Archaeol 37(3):209–225
Isern N, Zilhão J, Fort J, Ammerman AJ (2017) Modeling the role of voyaging in the coastal spread of the Early Neolithic in the West Mediterranean. Proc Natl Acad Sci 114:897–902
Jarić J (2004) Use of Pb and Sr isotopes in human teeth as an indicator of Pacific Islander population dynamics. Unpublished PhD dissertation, University of Western Sydney
Jordan FM, Gray RD, Greenhill SJ, Mace R (2009) Matrilocal residence is ancestral in Austronesian societies. Proc R Soc B 276:1957–1964
Kinaston RL, Walter RK, Jacomb C, Brooks E, Tayles N, Halcrow SE, Stirling C, Reid M, Gray AR, Spinks J, Shaw B, Fyfe R, Buckley HR (2013) The first New Zealanders: patterns of diet and mobility revealed through isotope analysis. PLoS One E8(5):e64580. https://doi.org/10.1371/journal.pone.0064580
Kinaston RL, Bedford S, Richards M, Hawkins S, Gray A, Jaouen K, Valentin F, Buckley H (2014) Diet and human mobility from the Lapita to the early historic period on Uripiv Island, Northeast Malakula, Vanuatu. PLoS One 9(8):e104071. https://doi.org/10.1371/journal.pone.0104071
Kirch PV (2010) How chiefs became kings: divine kingship and the rise of archaic states in ancient Hawai’i. University of California Press, Berkeley
Laffoon JE (2012) Patterns of paleomobility in the ancient Antilles: an isotopic approach. Unpublished PhD dissertation. University of Leiden
Laffoon JE (2016) Human mobility and dietary patterns in precolonial Puerto Rico: integrating multiple isotope data. In: Roksandic I (ed) Cuban archaeology in the Caribbean. University of Florida Press, Gainesville, pp 147–167
Laffoon JE, Hoogland MLP (2012) Migration and mobility in the circum-Caribbean: integrating archaeology and isotopic analysis. In: Kaiser E, Burger J, Schier W (eds) Population dynamics in prehistory and early history: new approaches using stable isotopes and genetics. de Gruyter, Berlin, pp 337–353
Laffoon JE, Hoogland ML, Davies GR, Hofman CL (2017) A Multi-Isotope Investigation of Human and Dog mobility and diet in the pre-colonial Antilles. Environ Archaeol:1–17
Lande R (1993) Risks of population extinction from demographic and environmental stochasticity and random catastrophes. Am Nat 142(6):911–927
Leppard TP (2015) Adaptive responses to demographic fragility: mitigating stochastic effects in early island colonization. Hum Ecol 43(5):721–734
Lightfoot E, O’Connell TC (2016) On the use of biomineral oxygen isotope data to identify human migrants in the archaeological record: sample variation, statistical methods and geographical considerations. PLoS One 11(4):e0153850
McArthur N, Saunders IW, Tweedie RL (1976) Small population isolates: a micro-simulation study. J Polynesian Soc 85:307–326
Moore JH (2001) Evaluating five models of human colonization. Am Anthropol 103(2):395–408
Nafplioti A (2011) Tracing population mobility in the Aegean using isotope geochemistry: a first map of local biologically available 87Sr/86Sr signatures. J Archaeol Sci 38:1560–1570
Pagán-Jiménez JR, Rodríguez-Ramos R, Reid BA, van den Bel M, Hofman CL (2015) Early dispersals of maize and other food plants into the Southern Caribbean and Northeastern South America. Quat Sci Rev 123(1):231–246
Phaff BN (2012) Human dietary and mobility patterns of a prehistoric population from Sigatoka, Fiji: a reconstruction using stable isotope analysis. Unpublished MA thesis. University of British Columbia
Richter-Dyn N, Goel NS (1972) On the extinction of a colonizing species. Theor Popul Biol 3:406–433
Rieth T, Athens JS (2017) Late Holocene human expansion into Near and Remote Oceania: a Bayesian model of the chronologies of the Mariana Islands and Bismarck Archipelago. J Isl Coast Archaeol:1–12. https://doi.org/10.1080/15564894.2017.1331939
Rybicki J, Hanski I (2013) Species–area relationships and extinctions caused by habitat loss and fragmentation. Ecol Lett 16:27–38
Schroeder H, O’Connell T, Evans JA, Shuler KA, Hedges REM (2009) Trans-Atlantic slavery: isotopic evidence for forced migration to Barbados. Am J Phys Anthropol 139(4):547–557
Schroeder H, Sikora M, Gopalakrishnan S, Cassidy LM, Delser PM, Velasco MS et al (2018) Origins and genetic legacies of the Caribbean Taino. Proc Natl Acad Sci 115(10):2341–2346
Shaw B, Summerhayes G, Buckley HR, Baker JA (2009) The use of strontium isotopes as an indicator of migration in human and pig Lapita populations in the Bismarck Archipelago, Papua New Guinea. J Archaeol Sci 36(4):1079–1091
Shaw B, Buckley H, Summerhayes G, Anson D, Garling S, Valentin F, Mandui H, Stirling C, Reid M (2010) Migration and mobility at the Late Lapita site of Reber-Rakival (SAC), Watom Island using isotope and trace element analysis: a new insight into Lapita interaction in the Bismarck Archipelago. J Archaeol Sci 37(3):605–613
Shaw B, Buckley H, Summerhayes G, Stirling C, Reid M (2011) Prehistoric migration at Nebira, South Coast of Papua New Guinea: new insights into interaction using isotope and trace element concentration analyses. J Anthropol Archaeol 30(3):344–358
Siegel PE (2010) Continuity and change in the evolution of religion and political organization on pre-Columbian Puerto Rico. J Anthropol Archaeol 29:302–326
Skoglund P, Posth C, Sirak K, Spriggs M, Valentin F, Bedford S, Clark GR, Reepmeyer C, Petchey F, Fernandes D, Fu Q, Harney E, Lipson M, Mallick S, Novak M, Rohland N, Stewardson K, Abdullah S, Cox MP, Friedlaender FR, Friedlaender JS, Kivisild T, Koki G, Kusuma P, Merriwether DA, Ricaut FX, Wee JTS, Patterson N, Krause J, Pinhasi R, Reich D (2016) Genomic insights into the peopling of the Southwest Pacific. Nature 538:510–513
Spriggs M (2011) Archaeology and the Austronesian expansion: where are we now? Antiquity 85(328):510–528
Stantis C, Buckley HR, Kinaston RL, Nunn PD, Jaouen K, Richards MP (2016) Isotopic evidence of human mobility and diet in a prehistoric/protohistoric Fijian coastal environment (c. 750–150 BP). Am J Phys Anthropol 159:478–495
Timpson A, Colledge S, Crema E, Edinborough K, Kerig T, Manning K, Thomas MG, Shennan S (2014) Reconstructing regional population fluctuations in the European Neolithic using radiocarbon dates: a new case-study using an improved method. J Archaeol Sci 52:549–557
Valcárcel Rojas R, Weston DA, Mickleburgh HL, Laffoon JE, van Duijvenbode A (2011) El Chorro de Maíta: a diverse approach to a context of diversity. In: Hofman CL, van Duijvenbode A (eds) Communities in Contact. Essays in archaeology, ethnohistory and ethnography of the Amerindian circum-Caribbean. Sidestone Press, Leiden, pp 225–252
Funding
JEL is funded in part by the European Research Council under the European Union’s Seventh Framework Programme (FP7/2007–2013)/ERC Synergy grant agreement no. 319209 under the auspices of the “NEXUS1492: New World Encounters in a Globalizing World” research project. TPL is funded by a Renfrew Fellowship in the McDonald Institute for Archaeological Research and Homerton College, University of Cambridge, for which support he is grateful and acknowledges.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Supplementary table 1
(XLSX 29 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Laffoon, J.E., Leppard, T.P. 87Sr/86Sr data indicate human post-juvenile residence mobility decreases over time-elapsed since initial Holocene island colonization in the Pacific and Caribbean. Archaeol Anthropol Sci 11, 1757–1768 (2019). https://doi.org/10.1007/s12520-018-0623-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12520-018-0623-9