
Abstract
Purpose of Review
Osteoarthritis (OA) is a subset of joint disorders resulting in degeneration of synovial joints. This leads to pain, disability and loss of independence. Knee and hip OA are extremely prevalent, and their occurrence increases with ageing. Similarly, loss of muscle mass and function, sarcopenia, occurs during ageing.
Recent Findings
Little is known about the impact of muscle wasting on OA progression; nevertheless, it has been suggested that muscle wasting directly affects the stability of the joints and loss of mobility leads to gradual degeneration of articular cartilage. The molecular mechanisms underlying muscle wasting in OA are not well understood; however, these are probably related to changes in gene expression, as well as epigenetic modifications.
Summary
It is becoming clear that skeletal muscle wasting plays an important role in OA development and/or progression. Here, we discuss mechanisms, current interventions, such as exercise, and potentially novel approaches, such as modulation of microRNAs, aiming at ameliorating OA symptoms through maintaining muscle mass and function.
Similar content being viewed by others
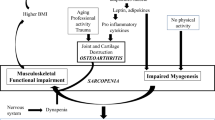


Introduction
Osteoarthritis (OA) is a multifactorial condition associated with degeneration of the joint, with pathological changes to multiple musculoskeletal tissues such as cartilage, meniscus, ligament and synovium. OA is characterised by degradation of articular cartilage, subchondral bone sclerosis, the formation of osteophytes at the joint margin and synovitis [1]. Synovial inflammation (synovitis) has been demonstrated during all stages of OA with synovium showing thickening and inflammation even before visible cartilage degeneration [2]. In addition, anterior cruciate ligament (ACL) injury, especially with concomitant ligament or meniscal pathology, increases the risk of knee OA suggesting an important role of healthy ligaments in maintaining joint tissue homeostasis [3]. OA patients and rodent models of OA are characterised by changes in cartilage, meniscus, synovium and ligament, suggesting common mechanisms of joint degeneration during OA development [1, 4]. Therefore, OA can be thought of as an organ failure of the whole synovial joint. Linked to the joint, muscles also play a major role in joint function during movement and in joint stability, but this link has been seldom explored. In this mini-review, we aim to provide current information linking muscle wasting and OA.
Muscle Wasting in OA
The progressive loss of periarticular muscle mass and function has consequences on joint stability and health. Muscle wasting is inevitably associated with ageing, and, more recently, it has been demonstrated in patients with OA [5,6,7]. The reduction in muscle mass and strength is attributed to myofibre atrophy, reduction of muscle quality and defective muscle regeneration (reviewed in [8]). Until recently, research investigating the association of muscle wasting with OA has been lacking. Currently, more and more data support the relationship between joint health and the surrounding skeletal muscle; however, functional and mechanistic studies are still lacking.
Muscle contractility is required for joint formation already during embryogenesis [9], and muscle weakness is an important determinant of pain and disability during OA [10]. Several studies have shown that a decrease in lower limb lean mass is frequent in OA patients [11], and this is associated with a greater risk of falls [12,13,14]. Progressive muscle weakness in OA is also associated with muscle fibre atrophy, with studies demonstrating 12–19% reduction in cross-sectional area of muscles in patients with hip and knee OA [6]. Moreover, muscle damage is associated with articular cartilage degeneration [15], whereas the increased cross-sectional area of vastus medialis is associated with improved structural changes at the knee and reduced pain in OA patients [16].
On the other hand, a large longitudinal cohort study found that, in 1653 subjects without radiographic knee OA (ROA) at baseline, an increased risk of ROA was not associated with sarcopenia alone, but rather with sarcopenic and body composition–based obesity [17]. Though, it should be noted that this study classified sarcopenia as the sum of absolute muscle mass of upper and lower limbs. However, findings from a previous study suggest that skeletal muscle mass of the lower limbs shows a higher correlation with knee OA than that of the lower and upper limbs combined [18]. Therefore, a statistical association between the risk of knee OA and sarcopenia may be observed, if the assessment of skeletal muscle mass focuses on the lower limbs.
A study investigating the influence of pain and ROA on muscle mass, strength, quality and risk of falls in older adults showed that hip and knee ROA are not significantly associated with changes in muscle strength and quality, despite the association of selfreported lower extremity joint pain, stiffness and dysfunction with declines in the aforementioned muscle parameters in older women. As such, it was concluded that pain may be the underlying mechanism via which OA leads to functional decline [19].
These contradictory results may be associated with a lack of a clear definition of sarcopenia. In recent years, many definitions of sarcopenia have been proposed, each one recommending diagnostic criteria based on muscle mass combined with measures of muscle strength, function or physical performance [20]. It is therefore crucial that criteria for the definition of sarcopenia are established and adopted in order for research to obtain results that are clinically relevant.
Another limitation of research in this area is the lack of a clear method to correlate the development of OA and incidence of sarcopenia. The current method used by research to investigate this relationship is to separately assess the OA severity grade and the lean body mass (LBM) of the area of interest [21]. Moreover, most of the research into OA and sarcopenia focuses on whether or not there is a correlation between the two disorders and has yet to fully investigate the molecular mechanisms behind the observed changes. It has been suggested that myokines, muscle-produced cytokines, peptides and growth factors communicate with the surrounding articular components such as the synovium, cartilage and bone through paracrine mediation, and thus may affect the signalling cascades implicated in OA [22].
Of the studies exploring molecular mechanisms underlying muscle wasting in OA, most have focused on inflammatory mediators as the molecular link between muscle function and OA. For example, Levinger et al. [23] observed increased inflammation in the vastus lateralis in patients with knee OA compared with a control group, based on increased protein abundance of p65 NF-κB, STAT-3 and JNK. However, a recent study attempted to investigate changes in muscle quality in OA by conducting the first cellular-level analysis of the vastus lateralis in adults with moderate knee OA [24]. Results of this study showed significant pathogenic fibrosis in the muscle of OA patients. Moreover, aberrant collagen deposition was noted in the extracellular matrix of OA muscle, which was significantly associated with decreased satellite cell density, as well as muscle strength. Increased expression levels of both CCN2 and TGFβ mRNAs were correlated positively to the amount of collagen deposition and inversely correlated with muscle strength. Finally, results indicated a muscle fibre type shift in the OA group, with significantly more type IIa/x hybrid fibres and fewer type I fibres in OA muscles relative to controls. Despite the limitations of this study, including low sample sizes and its cross-sectional nature, the data provide excellent molecular insight into the pathology of muscle changes that occur during OA. Nonetheless, there remains a large insufficiency of research in this area, making it essential that future studies focus on elucidating the mechanisms behind the influence of OA on skeletal muscle in order to develop more targeted therapeutic approaches. Interestingly, we have also shown that, in a spontaneous model of OA (STR/Ort mice), significant changes occur in the expression of muscle-related genes in the articular cartilage, suggesting at least some common molecular mechanisms underlying degeneration of musculoskeletal tissues during OA [25].
Another question remains to be answered: whether muscle changes precede OA, or vice versa. Studies have shown that low muscle quality and the depletion of lean body mass of lower extremities/body weight may be risk factors as important a contributor to OA pathogenesis as mechanical influences [26]. On the other hand, it has been suggested that the disuse of an OA affected joint, due to the pain of movement, may be the primary cause of the reduction in muscle strength associated with OA [27]. To add to the complexity, data from a study investigating muscle wasting in an anterior cruciate ligament transection (ACLT) model of OA suggest that disuse does not fully explain the muscle wasting observed in OA patients [6]. Results of this research show that rats in the OA group display the same pattern of movement as those of SHAM group (i.e., rats submitted to surgery without ALCT). Despite the similar locomotion (measured as spontaneous exploratory velocity and distance), the gastrocnemius crosssectional area was reduced by approximately 10% in the OA group. It was concluded, therefore, that muscle wasting may be a consequence of chronic, low-grade inflammation associated with OA, rather than solely joint disuse. However, the authors did acknowledge that the rats in the OA group displayed significantly increased nociception towards the end of the study, suggesting that the extended experimental period could have allowed for the detection of reduction in joint use.
The results of the above studies highlight the need to monitor the tendency of OA patients to avoid physical activity. It may prove beneficial for patients identified from this assessment to undergo therapeutic exercise intervention as a means to alleviate the symptoms of OA, as well as to improve their muscle strength.
Exercise Interventions to Maintain Muscle Mass and Function During OA
Current exercise interventions for OA focus highly on lifestyle modification, weight reduction and specific exercise interventions that focus on strengthening periarticular musculature [28, 29]. This is not without challenge, however, as many OA patients are unable to tolerate the load intensity required for progressive overloading of the relevant musculature [28]. Fortunately, the examination of modifications in low impact exercise, such as swimming and cycling, has shown a beneficial reduction in pain, stiffness and functional deficits in patients with knee OA [29, 30]. In a 12-week study examining swimming and cycling as exercise therapy for moderate knee OA, patients from both groups showed increased distance achieved in a 6minute walk test, as well as increased isokinetic knee extensor and flexor strength. Additionally, in the swimming cohort, there was an approximately 40% reduction in joint pain, as assessed by the Western Ontario and McMaster Universities Osteoarthritis Index (WOMAC) [29]. Limitations to this study, however, were the lack of participation of patients with advanced OA and the short intervention time. Other areas of research have evaluated similar low-load (2050% of one-repetition maximum) exercise training in knee OA patients utilising a partial vascular occlusion approach, wherein a pressure cuff or tourniquet is applied to the upper thigh, with similar results in pain reduction and functional improvements as compared with the conventional strength-training group (i.e., 70% of one-repetition maximum), yet with reduced knee pain whilst exercising [28]. The assumption is that this technique provides greater type II fibre activation via a generated anaerobic environment, whilst other possible mechanisms include increased intracellular metabolites (e.g., H+ protons, lactate and adenosine monophosphate), which in turn may stimulate growth hormone secretion [28, 31].
The majority of the literature suggests that sedentary lifestyle behaviours and immobilization are associated with cartilage disease and progression of OA, whereas controlled loading of the knee is associated with favourable changes to the extracellular matrix and molecular biomarkers associated with OA such as Creactive protein and IL-10 [32]. Therapeutic exercise is beneficial to cartilage health, as the literature suggests molecular biomarkers associated with inflammation (i.e., CRP and TNFα) and cartilage breakdown (C12C, CTX-II and COMP) are either unchanged or downregulated in studies examining these characteristics in exercise and OA [32]. Furthermore, exercise programmes focusing on neuromuscular control, such as closed chain kinetic and proprioceptive exercises, are generally well tolerated in OA patients and show results similar to more traditional strengthening exercises [33, 34]. This is consistent with literature suggesting that aberrations in the neuro-motor system are involved in changes to muscle force patterns, with altered joint reaction forces leading to focal alterations to cartilage loading over time [35]. This suggests that there is a complex relationship between both muscular strength and changes to motor unit physiology in OA, and strengthening alone may not be the only consideration in clinical practice [35]. Meta-analyses show that no specific protocol is deemed the best and current guidelines are ambiguous in recommending exercise programs [36]; thus, the best exercise programme may be the one that is met with high adherence by the individual and tolerated enough to provide consistent progression over time with graded adjustments to load and volume. Recent exercise interventions in OA patients are summarised in Table 1.
microRNAs as Novel Therapeutic Targets for Muscle Wasting During OA
MicroRNAs (also referred to as miRNAs or miRs) are a class of small non-coding RNA molecules, approximately 22 nucleotides long, which bind to their target messenger RNAs (mRNAs), induce mRNA degradation or inhibit protein translation, and thus exert post-transcriptional regulation of gene expression [38]. Studies report that specific miRNA-target interactions may regulate all major known contributors to the onset of the senescent phenotype, such as DNA damage, telomere shortening, protein misfolding and oxidative stress [39]. Therefore, recent research has focused on investigating whether restoring physiologic levels of specific miRNAs in these tissues can restore—at least partially—functional homeostasis [40]. Investigating the use of miRNA therapies in disorders such as OA is particularly important given that the only current treatment options for the condition are pain and symptom management and eventual joint replacement therapy [41]. It is anticipated that elucidating the underlying molecular mechanisms of the disorder, in particular the associated genetic and epigenetic dysregulation, will lead to the development of novel interventions to delay the need for invasive and transient total joint replacement procedures.
One of the most well-studied miRNAs in OA research, to date, is miR-140, a cartilage-specific miRNA [42], which was first implicated in OA pathogenesis in a study by Miyaki et al. This study showed that miR-140 expression in articular cartilage from OA donors is significantly lower than that in normal cartilage or following treatment with IL-1β, a cytokine known to be involved in the pathogenesis of OA. Interestingly, the transfection of chondrocytes with miR-140 downregulated IL-1β-induced ADAMTS5 gene expression. The correlation of abnormal miR-140 expression in OA cartilage with increased ADAMTS5, as first reported in this study [42], suggests that miR-140 may play an important role in maintaining cartilage homeostasis. Since then, multiple studies have shown that miRNAs play significant roles in cartilage homeostasis. Moreover, there is evidence that miRNAs are important players in maintaining the health of all joint tissues, including muscle [43,44,45,46,47], cartilage [48, 49] and ligament and tendon [50, 51]. Whilst there is very little known about the role of specific microRNAs in muscle wasting during OA, with only one microRNA, miR-141, suggested to have a function in the ageing of the multiple tissues of the musculoskeletal system [52], multiple studies have shown the involvement of miRNAs in muscle wasting [53,54,55]. The importance of microRNAs in the function of musculoskeletal tissues was also demonstrated in depletion studies of Dicer, a key enzyme in miRNA biogenesis. The deletion of Dicer in chondrocytes resulted in defective cell proliferation and differentiation [51], and the deletion of Dicer in bone progenitors was associated with defective bone formation [56]. A satellite cell–specific Dicer knockout resulted in mild muscle fibre atrophy over time [57], and microRNAs maintain muscle homeostasis through the control of muscle hypertrophy and atrophy and muscle adaptation to exercise [58,59,60]. Below, we focus on several examples of microRNAs with a potential role in regulating homeostasis of muscle and other joint tissues in the context of OA.
miR-143 is differentially expressed in the cartilage of older people suffering from OA [53]. Interestingly, miR-143 regulates the phenotype of synovial fibroblasts in rheumatoid arthritis through the regulation of IGFBP5 [61]. Our group has previously demonstrated the regulation of senescence of satellite cells, adult skeletal muscle stem cells, via the miR-143:Igfbp5 regulatory network [47]. Interestingly, miR-143:Igfbp5 interactions are regulated by inflammatory cytokines, including IL6.
Our group has also shown that miR-181 is downregulated in muscle during ageing and regulates the expression of Sirt1 [46]. Interestingly, another group has shown that the expression of miR-181 is also altered in the joints of OA patients and that inhibition of this microRNA in animal models of OA attenuates OA in facet and knee joints [62].
Another microRNA downregulated in muscle of humans during ageing is miR-378 [63], which controls myogenesis, metabolism and autophagy [55, 64, 65]. Indeed, molecules regulating miR-378 have been patented for the use in regulating metabolism in different tissues, including skeletal muscle (patent number WO2011153542A3). The changes in the regulation of cellular metabolism, for example through autophagy, have also been noted during OA, and miR-378 has been detected in synovium in late-stage OA patients [66, 67]. Moreover, miR-378 promotes osteogenesis in human bone marrow–derived mesenchymal stem cells, suggesting that it is a pro-bone regeneration molecule [68]. Together, these data indicate a potential common mechanism, via miR-378 and other microRNAs, underlying musculoskeletal tissues homeostasis.
Whilst multiple studies have focused on the role of microRNAs in muscle development and regeneration, very few functional studies exist that have characterised the role of microRNAs in muscle wasting during ageing or OA [69, 70].
It also has to be noted that Soares et al. [45] demonstrated that microRNA function is context dependent in different models of muscle atrophy; therefore, a question arises whether the miR-associated mechanisms of muscle wasting during ageing are similar or different to those observed during muscle wasting in OA. Functional studies of microRNAs in muscle during OA are needed.
Conclusions
OA is a progressive and debilitating disease and the most common cause of chronic disability in adults. Despite this, pain management and total joint replacement procedures are the only current treatment options for the disease. As such, a lot of research into OA has focused on the development of novel interventions to delay the need for invasive total joint replacement procedures. The association between skeletal muscle mass and function and OA development and progression remains to be elucidated; however, it is becoming clearer that changes in skeletal muscle are important players in OA. As such, exercise interventions could benefit OA patients not only due to benefits on muscle tissues, but also through all joint tissues affected by OA. An emerging approach to interventions against OA includes the regulation of expression and function of microRNAs. Although the role of microRNAs in muscle wasting in OA is not yet understood, given that a single microRNA can regulate multiple genes and dysregulation of microRNA expression occurs in a variety of pathological conditions, microRNAs are emerging powerful regulatory molecules and potential novel therapeutic agents; indeed, microRNAbased therapies are currently undergoing clinical trials [71].
Though currently in its infancy, the use of miRNA-based therapeutics in OA, including but not limited to regulating muscle mass and function, is a promising avenue of research, with many recent studies suggesting that their use may overcome the limitations of many traditional therapies. However, functional approaches are lacking. Moreover, experiments using more relevant animal OA models and large-scale or cohort-based clinical trials should be conducted in order to comprehensively evaluate the efficacy of miRNA therapeutics in OA.
References
Loeser RF, Goldring SR, Scanzello CR, Goldring MB. Osteoarthritis: a disease of the joint as an organ. Arthritis Rheum. 2012;64(6):1697–707. https://doi.org/10.1002/art.34453.
Mathiessen A, Conaghan PG. Synovitis in osteoarthritis: current understanding with therapeutic implications. Arthritis Res Ther. 2017;19(1):18. https://doi.org/10.1186/s13075-017-1229-9.
Simon D, Mascarenhas R, Saltzman BM, Rollins M, Bach BR Jr, MacDonald P. The relationship between anterior cruciate ligament injury and osteoarthritis of the knee. Adv Orthop. 2015;2015:928301. https://doi.org/10.1155/2015/928301.
Poulet B, Staines KA. New developments in osteoarthritis and cartilage biology. Curr Opin Pharmacol. 2016;28:8–13. https://doi.org/10.1016/j.coph.2016.02.009.
Narici MV, Maffulli N. Sarcopenia: characteristics, mechanisms and functional significance. Br Med Bull. 2010;95:139–59. https://doi.org/10.1093/bmb/ldq008.
Silva JMS, Alabarse PVG, Teixeira VON, Freitas EC, de Oliveira FH, Chakr R, et al. Muscle wasting in osteoarthritis model induced by anterior cruciate ligament transection. PLoS One. 2018;13(4):e0196682. https://doi.org/10.1371/journal.pone.0196682.
Larsson L, Degens H, Li M, Salviati L, Lee YI, Thompson W, et al. Sarcopenia: aging-related loss of muscle mass and function. Physiol Rev. 2019;99(1):427–511. https://doi.org/10.1152/physrev.00061.2017.
Brown DM, Goljanek-Whysall K. microRNAs: Modulators of the underlying pathophysiology of sarcopenia? Ageing Res Rev. 2015;24(Pt B):263–73. https://doi.org/10.1016/j.arr.2015.08.007.
Kahn J, Shwartz Y, Blitz E, Krief S, Sharir A, Breitel DA, et al. Muscle contraction is necessary to maintain joint progenitor cell fate. Dev Cell. 2009;16(5):734–43. https://doi.org/10.1016/j.devcel.2009.04.013.
Fisher NM, White SC, Yack HJ, Smolinski RJ, Pendergast DR. Muscle function and gait in patients with knee osteoarthritis before and after muscle rehabilitation. Disabil Rehabil. 1997;19(2):47–55.
Toda Y, Kobayashi T. The usefulness of walking for preventing sarcopenia in dieting postmenopausal women complaining of knee pain. Ann N Y Acad Sci. 2000;904:610–3.
O’Reilly SC, Jones A, Muir KR, Doherty M. Quadriceps weakness in knee osteoarthritis: the effect on pain and disability. Ann Rheum Dis. 1998;57(10):588–94.
Segal NA, Glass NA, Torner J, Yang M, Felson DT, Sharma L, et al. Quadriceps weakness predicts risk for knee joint space narrowing in women in the MOST cohort. Osteoarthritis Cartilage. 2010;18(6):769–75. https://doi.org/10.1016/j.joca.2010.02.002.
Conroy MB, Kwoh CK, Krishnan E, Nevitt MC, Boudreau R, Carbone LD, et al. Muscle strength, mass, and quality in older men and women with knee osteoarthritis. Arthritis Care Res. 2012;64(1):15–21. https://doi.org/10.1002/acr.20588.
Isaac C, Wright A, Usas A, Li H, Tang Y, Mu X, et al. Dystrophin and utrophin "double knockout" dystrophic mice exhibit a spectrum of degenerative musculoskeletal abnormalities. J Orthop Res. 2013;31(3):343–9. https://doi.org/10.1002/jor.22236.
Wang Y, Wluka AE, Berry PA, Siew T, Teichtahl AJ, Urquhart DM, et al. Increase in vastus medialis cross-sectional area is associated with reduced pain, cartilage loss, and joint replacement risk in knee osteoarthritis. Arthritis Rheum. 2012;64(12):3917–25. https://doi.org/10.1002/art.34681.
Misra D, Fielding RA, Felson DT, Niu J, Brown C, Nevitt M, et al. Risk of knee osteoarthritis with obesity, sarcopenic obesity, and sarcopenia. Arthritis Rheum. 2019;71(2):232–7. https://doi.org/10.1002/art.40692.
Lee SY, Ro HJ, Chung SG, Kang SH, Seo KM, Kim DK. Low skeletal muscle mass in the lower limbs is independently associated to knee osteoarthritis. PLoS One. 2016;11(11):e0166385. https://doi.org/10.1371/journal.pone.0166385.
Scott D, Blizzard L, Fell J, Jones G. Prospective study of self-reported pain, radiographic osteoarthritis, sarcopenia progression, and falls risk in community-dwelling older adults. Arthritis Care Res. 2012;64(1):30–7. https://doi.org/10.1002/acr.20545.
Cruz-Jentoft AJ, Bahat G, Bauer J, Boirie Y, Bruyere O, Cederholm T, et al. Sarcopenia: revised European consensus on definition and diagnosis. Age Ageing. 2019;48(1):16–31. https://doi.org/10.1093/ageing/afy169.
Papalia R, Zampogna B, Torre G, Lanotte A, Vasta S, Albo E, et al. Sarcopenia and its relationship with osteoarthritis: risk factor or direct consequence? Musculoskelet Surg. 2014;98(1):9–14. https://doi.org/10.1007/s12306-014-0311-6.
Krishnasamy P, Hall M, Robbins SR. The role of skeletal muscle in the pathophysiology and management of knee osteoarthritis. Rheumatology. 2018;57(suppl_4):iv124. https://doi.org/10.1093/rheumatology/key039.
Levinger P, Caldow MK, Feller JA, Bartlett JR, Bergman NR, McKenna MJ, et al. Association between skeletal muscle inflammatory markers and walking pattern in people with knee osteoarthritis. Arthritis Care Res. 2011;63(12):1715–21. https://doi.org/10.1002/acr.20625.
Noehren B, Kosmac K, Walton RG, Murach KA, Lyles MF, Loeser RF, et al. Alterations in quadriceps muscle cellular and molecular properties in adults with moderate knee osteoarthritis. Osteoarthritis and cartilage / OARS. Osteoarthritis Res Soc. 2018;26(10):1359–68. https://doi.org/10.1016/j.joca.2018.05.011.
Poulet B, Westerhof TA, Hamilton RW, Shefelbine SJ, Pitsillides AA. Spontaneous osteoarthritis in Str/ort mice is unlikely due to greater vulnerability to mechanical trauma. Osteoarthritis Cartilage. 2013;21(5):756–63. https://doi.org/10.1016/j.joca.2013.02.652.
Toda Y, Segal N, Toda T, Kato A, Toda F. A decline in lower extremity lean body mass per body weight is characteristic of women with early phase osteoarthritis of the knee. J Rheumatol. 2000;27(10):2449–54.
Pisters MF, Veenhof C, van Dijk GM, Dekker J, Group CS. Avoidance of activity and limitations in activities in patients with osteoarthritis of the hip or knee: a 5 year follow-up study on the mediating role of reduced muscle strength. Osteoarthritis Cartilage. 2014;22(2):171–7. https://doi.org/10.1016/j.joca.2013.12.007.
Bryk FF, Dos Reis AC, Fingerhut D, Araujo T, Schutzer M, Cury Rde P, et al. Exercises with partial vascular occlusion in patients with knee osteoarthritis: a randomized clinical trial. Knee Surg Sports Traumatol Arthrosc. 2016;24(5):1580–6. https://doi.org/10.1007/s00167-016-4064-7.
Alkatan M, Baker JR, Machin DR, Park W, Akkari AS, Pasha EP, et al. Improved function and reduced pain after swimming and cycling training in patients with osteoarthritis. J Rheumatol. 2016;43(3):666–72. https://doi.org/10.3899/jrheum.151110.
Al-Khlaifat L, Herrington LC, Hammond A, Tyson SF, Jones RK. The effectiveness of an exercise programme on knee loading, muscle co-contraction, and pain in patients with medial knee osteoarthritis: a pilot study. Knee. 2016;23(1):63–9. https://doi.org/10.1016/j.knee.2015.03.014.
Laurentino G, Ugrinowitsch C, Aihara AY, Fernandes AR, Parcell AC, Ricard M, et al. Effects of strength training and vascular occlusion. Int J Sports Med. 2008;29(8):664–7. https://doi.org/10.1055/s-2007-989405.
Bricca A, Struglics A, Larsson S, Steultjens M, Juhl CB, Roos EM. Impact of exercise therapy on molecular biomarkers related to cartilage and inflammation in people at risk of, or with established, knee osteoarthritis: a systematic review and meta-analysis of randomized controlled trials. Arthritis Care Res. 2018. https://doi.org/10.1002/acr.23786.
Bennell KL, Kyriakides M, Metcalf B, Egerton T, Wrigley TV, Hodges PW, et al. Neuromuscular versus quadriceps strengthening exercise in patients with medial knee osteoarthritis and varus malalignment: a randomized controlled trial. Arthritis Rheum. 2014;66(4):950–9. https://doi.org/10.1002/art.38317.
Clausen B, Holsgaard-Larsen A, Roos EM. An 8-week neuromuscular exercise program for patients with mild to moderate knee osteoarthritis: a case series drawn from a registered clinical trial. J Athl Train. 2017;52(6):592–605. https://doi.org/10.4085/1062-6050-52.5.06.
Miller MS, Callahan DM, Tourville TW, Slauterbeck JR, Kaplan A, Fiske BR, et al. Moderate-intensity resistance exercise alters skeletal muscle molecular and cellular structure and function in inactive older adults with knee osteoarthritis. J Appl Physiol. 2017;122(4):775–87. https://doi.org/10.1152/japplphysiol.00830.2016.
Bartholdy C, Juhl C, Christensen R, Lund H, Zhang W, Henriksen M. The role of muscle strengthening in exercise therapy for knee osteoarthritis: a systematic review and meta-regression analysis of randomized trials. Semin Arthritis Rheum. 2017;47(1):9–21. https://doi.org/10.1016/j.semarthrit.2017.03.007.
Koli J, Multanen J, Kujala UM, Hakkinen A, Nieminen MT, Kautiainen H, et al. Effects of exercise on patellar cartilage in women with mild knee osteoarthritis. Med Sci Sports Exerc. 2015;47(9):1767–74. https://doi.org/10.1249/MSS.0000000000000629.
Friedman RC, Farh KK, Burge CB, Bartel DP. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009;19(1):92–105. https://doi.org/10.1101/gr.082701.108.
Williams J, Smith F, Kumar S, Vijayan M, Reddy PH. Are microRNAs true sensors of ageing and cellular senescence? Ageing Res Rev. 2017;35:350–63. https://doi.org/10.1016/j.arr.2016.11.008.
Rupaimoole R, Slack FJ. MicroRNA therapeutics: towards a new era for the management of cancer and other diseases. Nat Rev Drug Discov. 2017;16(3):203–22. https://doi.org/10.1038/nrd.2016.246.
Hermann W, Lambova S, Muller-Ladner U. Current treatment options for osteoarthritis. Curr Rheumatol Rev. 2018;14(2):108–16. https://doi.org/10.2174/1573397113666170829155149.
Tuddenham L, Wheeler G, Ntounia-Fousara S, Waters J, Hajihosseini MK, Clark I, et al. The cartilage specific microRNA-140 targets histone deacetylase 4 in mouse cells. FEBS Lett. 2006;580(17):4214–7. https://doi.org/10.1016/j.febslet.2006.06.080.
Drummond MJ, McCarthy JJ, Sinha M, Spratt HM, Volpi E, Esser KA, et al. Aging and microRNA expression in human skeletal muscle: a microarray and bioinformatics analysis. Physiol Genomics. 2011;43(10):595–603. https://doi.org/10.1152/physiolgenomics.00148.2010.
McCarthy JJ, Esser KA. MicroRNA-1 and microRNA-133a expression are decreased during skeletal muscle hypertrophy. J Appl Physiol. 2007;102(1):306–13. https://doi.org/10.1152/japplphysiol.00932.2006.
Soares RJ, Cagnin S, Chemello F, Silvestrin M, Musaro A, De Pitta C, et al. Involvement of microRNAs in the regulation of muscle wasting during catabolic conditions. J Biol Chem. 2014;289(32):21909–25. https://doi.org/10.1074/jbc.M114.561845.
Soriano-Arroquia A, House L, Tregilgas L, Canty-Laird E, Goljanek-Whysall K. The functional consequences of age-related changes in microRNA expression in skeletal muscle. Biogerontology. 2016;17(3):641–54. https://doi.org/10.1007/s10522-016-9638-8.
Soriano-Arroquia A, McCormick R, Molloy AP, McArdle A, Goljanek-Whysall K. Age-related changes in miR-143-3p:Igfbp5 interactions affect muscle regeneration. Aging Cell. 2016;15(2):361–9. https://doi.org/10.1111/acel.12442.
Philipot D, Guerit D, Platano D, Chuchana P, Olivotto E, Espinoza F, et al. p16INK4a and its regulator miR-24 link senescence and chondrocyte terminal differentiation-associated matrix remodeling in osteoarthritis. Arthritis Res Ther. 2014;16(1):R58. https://doi.org/10.1186/ar4494.
Barter MJ, Bui C, Young DA. Epigenetic mechanisms in cartilage and osteoarthritis: DNA methylation, histone modifications and microRNAs. Osteoarthritis and cartilage / OARS. Osteoarthritis Res Soc. 2012;20(5):339–49. https://doi.org/10.1016/j.joca.2011.12.012.
Dakin SG. MicroRNA replacement: a new era of molecular therapy for tendon disorders? Mol Ther. 2017;25(10):2243–4. https://doi.org/10.1016/j.ymthe.2017.09.004.
Ng TK, Yang Q, Fortino VR, Lai NY, Carballosa CM, Greenberg JM, et al. MicroRNA-132 directs human periodontal ligament-derived neural crest stem cell neural differentiation. J Tissue Eng Regen Med. 2019;13(1):12–24. https://doi.org/10.1002/term.2759.
Fariyike B, Singleton Q, Hunter M, Hill WD, Isales CM, Hamrick MW, et al. Role of MicroRNA-141 in the aging musculoskeletal system: a current overview. Mech Ageing Dev. 2019;178:9–15. https://doi.org/10.1016/j.mad.2018.12.001.
Balaskas P, Goljanek-Whysall K, Clegg P, Fang Y, Cremers A, Emans P, et al. MicroRNA profiling in cartilage ageing. Int J Genomics. 2017;2017:2713725. https://doi.org/10.1155/2017/2713725.
McCormick R, Goljanek-Whysall K. MicroRNA dysregulation in aging and pathologies of the skeletal muscle. Int Rev Cell Mol Biol. 2017;334:265–308. https://doi.org/10.1016/bs.ircmb.2017.03.005.
Proctor CJ, Goljanek-Whysall K. Using computer simulation models to investigate the most promising microRNAs to improve muscle regeneration during ageing. Sci Rep. 2017;7(1):12314. https://doi.org/10.1038/s41598-017-12538-6.
Gaur T, Hussain S, Mudhasani R, Parulkar I, Colby JL, Frederick D, et al. Dicer inactivation in osteoprogenitor cells compromises fetal survival and bone formation, while excision in differentiated osteoblasts increases bone mass in the adult mouse. Dev Biol. 2010;340(1):10–21. https://doi.org/10.1016/j.ydbio.2010.01.008.
Cheung TH, Quach NL, Charville GW, Liu L, Park L, Edalati A, et al. Maintenance of muscle stem-cell quiescence by microRNA-489. Nature. 2012;482(7386):524–8. https://doi.org/10.1038/nature10834.
Russell AP, Lamon S, Boon H, Wada S, Guller I, Brown EL, et al. Regulation of miRNAs in human skeletal muscle following acute endurance exercise and short-term endurance training. J Physiol. 2013;591(18):4637–53. https://doi.org/10.1113/jphysiol.2013.255695.
Ultimo S, Zauli G, Martelli AM, Vitale M, McCubrey JA, Capitani S, et al. Influence of physical exercise on microRNAs in skeletal muscle regeneration, aging and diseases. Oncotarget. 2018;9(24):17220–37. https://doi.org/10.18632/oncotarget.24991.
Widmann M, Niess AM, Munz B. Physical exercise and epigenetic modifications in skeletal muscle. Sports Med. 2019. https://doi.org/10.1007/s40279-019-01070-4.
Hong BK, You S, Yoo SA, Park D, Hwang D, Cho CS, et al. MicroRNA-143 and -145 modulate the phenotype of synovial fibroblasts in rheumatoid arthritis. Exp Mol Med. 2017;49(8):e363. https://doi.org/10.1038/emm.2017.108.
Nakamura A, Rampersaud YR, Nakamura S, Sharma A, Zeng F, Rossomacha E, et al. microRNA-181a- 5p antisense oligonucleotides attenuate osteoarthritis in facet and knee joints. Ann Rheum Dis. 2019;78(1):111–21. https://doi.org/10.1136/annrheumdis-2018-213629.
Drummond MJ, McCarthy JJ, Fry CS, Esser KA, Rasmussen BB. Aging differentially affects human skeletal muscle microRNA expression at rest and after an anabolic stimulus of resistance exercise and essential amino acids. Am J Physiol Endocrinol Metab. 2008;295(6):E1333–40. https://doi.org/10.1152/ajpendo.90562.2008.
Li Y, Jiang J, Liu W, Wang H, Zhao L, Liu S, et al. microRNA-378 promotes autophagy and inhibits apoptosis in skeletal muscle. Proc Natl Acad Sci U S A. 2018;115(46):E10849–E58. https://doi.org/10.1073/pnas.1803377115.
Carrer M, Liu N, Grueter CE, Williams AH, Frisard MI, Hulver MW, et al. Control of mitochondrial metabolism and systemic energy homeostasis by microRNAs 378 and 378*. Proc Natl Acad Sci U S A. 2012;109(38):15330–5. https://doi.org/10.1073/pnas.1207605109.
Li YH, Tavallaee G, Tokar T, Nakamura A, Sundararajan K, Weston A, et al. Identification of synovial fluid microRNA signature in knee osteoarthritis: differentiating early- and late-stage knee osteoarthritis. Osteoarthritis Cartilage. 2016;24(9):1577–86. https://doi.org/10.1016/j.joca.2016.04.019.
Cheng NT, Meng H, Ma LF, Zhang L, Yu HM, Wang ZZ, et al. Role of autophagy in the progression of osteoarthritis: the autophagy inhibitor, 3-methyladenine, aggravates the severity of experimental osteoarthritis. Int J Mol Med. 2017;39(5):1224–32. https://doi.org/10.3892/ijmm.2017.2934.
Zhang B, Li Y, Yu Y, Zhao J, Ou Y, Chao Y, et al. MicroRNA-378 promotes osteogenesis-angiogenesis coupling in BMMSCs for potential bone regeneration. Anal Cell Pathol (Amst). 2018;2018:8402390. https://doi.org/10.1155/2018/8402390.
Jung HJ, Lee KP, Kwon KS, Suh Y. MicroRNAs in skeletal muscle aging: current issues and perspectives. J Gerontol A Biol Sci Med Sci. 2018. https://doi.org/10.1093/gerona/gly207.
Goldring MB, Marcu KB. Epigenomic and microRNA-mediated regulation in cartilage development, homeostasis, and osteoarthritis. Trends Mol Med. 2012;18(2):109–18. https://doi.org/10.1016/j.molmed.2011.11.005.
Bouchie A. First microRNA mimic enters clinic. Nat Biotechnol. 2013;31(7):577. https://doi.org/10.1038/nbt0713-577.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
This article is part of the Topical Collection on Osteoarthritis
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Shorter, E., Sannicandro, A.J., Poulet, B. et al. Skeletal Muscle Wasting and Its Relationship With Osteoarthritis: a Mini-Review of Mechanisms and Current Interventions. Curr Rheumatol Rep 21, 40 (2019). https://doi.org/10.1007/s11926-019-0839-4
Published:
DOI: https://doi.org/10.1007/s11926-019-0839-4