
Abstract
IgE-mediated food allergy is a relevant health problem inducing symptoms ranging from mild local reactions up to severe life-threatening situations. Currently, no immunotherapy is available and avoidance of the incriminating food is the method of choice. Therefore, reliable diagnostic tools to formulate dietary recommendations and to avoid unnecessary exclusion diets for the individual patient are urgently needed. This review provides an update on the current knowledge on food allergens and their application in various diagnostic approaches such as skin prick test, basophil activation test, and serum IgE testing. Furthermore, these new approaches are discussed and compared to conventional extract-based assays and correlated to the gold standard of food allergy diagnosis, the double-blind placebo-controlled food challenge. Finally, the application of food allergens for preventive measurements such as allergen detection assays and the determination of threshold levels for allergen levels are discussed.
Similar content being viewed by others


Introduction
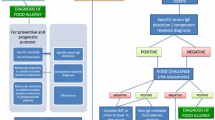
Allergic diseases are regarded as a relevant global burden to society and healthcare systems. The prevalence for asthma, allergic rhinitis, and atopic eczema has indeed increased in the recent past. However, for IgE-mediated food allergy, the picture is less clear. A recent systematic review and meta-analysis of food allergy in Europe identified a 17.3 % pooled lifetime prevalence of self-reported food allergy and a point prevalence of 5.9 % [1, 2]. In contrast, the point prevalence of food allergy (FA) diagnosis based on food challenges was around 0.9 %, yet differing on the food allergen source and depending on the age. While for diagnosis of food allergy, double-blind placebo-controlled food challenge (DBPCFC) is regarded as the gold standard; a careful case history is equally mandatory. The detection of allergen-specific IgE antibodies completes the state-of-the-art diagnosis of food allergy [3]. Besides symptomatic treatment, there is no causative immunotherapy available. Currently, avoiding the causative allergen sources is the method of choice.
In food allergy, two different routes of sensitization are known. While in primary food allergy, the atopic individual becomes sensitized against the culprit dietary protein through the gastrointestinal tract, and in secondary food allergy, sensitization occurs via pollen or latex allergens and the allergic reaction is induced by cross-reactive homologous food allergens [4].
The diagnostic relevance of specific IgE testing strongly depends on the quality of the analyte used. For more than 100 years, total extracts obtained from allergen sources were used. It is well known that these extracts are not very well standardized and may lack certain allergens, e.g., due to enzymatic activity [3].
During the past 25 years, a plethora of individual allergens has been identified and their physicochemical properties analyzed. Especially, up-to-date molecular biology methods facilitated the production of recombinant allergens for in vitro diagnosis. The determination of specific IgE to different allergens is called molecular diagnosis or component-resolved diagnosis (CRD).
Molecular diagnosis will help to increase the sensitivity of the testing and, on the other hand, provides a more-detailed patient-tailored risk profile. This in turn can help to formulate improved dietary recommendations and reduce unnecessary exclusion diets [3].
However, it has to be stated that most allergens described in CRD studies so far have been applied in experimental settings. Therefore, the recent EAACI Guidelines on the Diagnosis and Management of Food Allergy clearly emphasized the urgent need for well-designed randomized controlled studies to assess the diagnostic value of CRD-based tests [3].
Allergen Extract Versus Single Allergens
Production and Purification of Allergens for Molecular Diagnosis
Allergens used for CRD can be obtained from either natural sources or expressed in heterologous expression systems. The decision whether to purify natural or recombinant allergens has to be made on a case-by-case approach and depends on a number of aspects. For example, the presence and impact of any post translational modification on the IgE binding activity of the target protein need to be evaluated throughout the purification process. If a given allergen consists of a number of isoforms all relevant for allergy diagnosis, then the natural mix of proteins is to be preferred. In contrast, if one isoform is representative for IgE-based diagnosis, then the recombinant production can be chosen. Similarly, if the natural protein is sensitive to enzymatic degradation during the extraction process, then heterologous expression systems can be advantageous. If the correct three-dimensional structure of the protein is crucial for the known interaction with IgE antibodies, then either suitable expression systems or purification from natural sources can be chosen [5, 6]. Equally important is the development of a suitable purification protocol of the allergen of interest. Finally, the protein has to be tested for its physicochemical properties such as stability, purity, correct primary, secondary, and tertiary structure, enzymatic activity, and IgE binding capacity [7, 6]. As a proof of concept, an allergen library was developed within the EU-funded project EuroPrevall, collecting highly pure and well-characterized food allergens from the most important food allergen sources. Up to date, the integrity of the purified allergen batches was assessed by technologies such as HPLC, mass spectrometry, and infrared spectroscopy. CD spectroscopy and 1D-NMR can be applied to analyze the secondary structure of allergens [8].
As stated above, a considerable number of allergens have been identified from animal- and plant-derived foods. Consecutively, a number of allergen databases were established to collect and update the existing data on allergens, their physicochemical features, and allergenic relevance. The IUIS allergen database (http://www.allergen.org) is the official source, providing a systematic nomenclature for allergens, taking care of identification of allergens, and upon positive evaluation of the allergen nomenclature subcommittee, the official allergen designation is granted [9, 10]. The Allergome database (http://www.allergome.org) is a continuously updated non-peer-reviewed literature collection about all aspects of allergens, while the AllergenOnline (http://www.allergenonline.org) provides peer-reviewed information about allergens and sequence comparison tools for allergenic risk assessment. SDAP, the structural database of allergens, collects all the known structures of allergens (http://fermi.utmb.edu/SDAP/index.html), while others such as the InformAll link clinical data with allergens (http://www.inflammation-repair.manchester.ac.uk/informAll/).
Analyzing all the existing allergen sequences, it became evident that only a minority of all known protein families contain intrinsic properties that render them allergenic [11–13].
Tests for Food Allergy Diagnosis
In the following, the application of allergen-specific testing for food allergy diagnosis will be discussed. Furthermore, an overview on the most important plant-derived food allergen sources and their respective allergen panels will be presented.
Skin Prick Testing
In vivo skin prick testing for food allergy diagnosis is routinely performed. The food extracts used for skin prick testing (SPT) are not all standardized and may vary from batch to batch missing some allergens [14–16]. Therefore, prick-to-prick testing with the food source is applied in daily practice, especially when testing plant food allergen sources (fresh fruits, vegetables, nuts).
In proof-of-concept studies, purified recombinant fruit allergens from apple and peach were tested in SPTs. Bolhaar and colleagues applied purified recombinant apple allergen Mal d 1, the Bet v 1 homologue, by SPT in 14 patients using serial dilutions ranging from 0.02, 0.2, 2, 20, to 100 μg/mL. Positive skin reactions were observed in a dose-dependent manner starting with the lowest concentration of 2 μg/mL [17]. In a similar study, purified recombinant Mal d 4, the apple profilin, was tested in 5 patients in the same serial dilutions 0.02, 0.2, 2, 20, and 100 μg/mL, giving positive reactions at a the dose of 0.2 μg/mL [18].
Garcia et al. performed SPTs with four peach extracts containing 0.4, 2, 10, and 50 μg/mL of Pru p 3, the non-specific lipid transfer protein (nsLTP) from peach [19], during a sublingual immunotherapy trial with natural Pru p 3. They showed that after 6 months of SLIT, the active group presented a significant decrease (5.3 times) in SPTs, which correlated with a significant increased tolerance of peach intake [19].
In a recent study, Kollmann et al. used purified recombinant Mal d 1 and Bet v 1 (birch pollen allergen) and birch pollen extract (BPE) in SPTs [20•]. For this study, the recombinant allergen was produced under good manufacturing practice (GMP) conditions suitable for clinical trials. Twenty out of 21 patients had a positive reaction to rMal d 1 (50 μg/mL) and to fresh apple. The overall sensitivity of the SPT was 95 % for rMal d 1 [20•].
There are not yet purified recombinant allergens approved for SPTs in clinical routine, while for some European countries, purified natural peach nsLTP (Pru p 3) and purified natural date profilin can be used [21].
Peeters and colleagues applied purified natural peanut allergens, Ara h 1, Ara h 2, Ara h 3, and Ara h 6, in SPTs in a collective of 30 peanut-allergic patients in serial dilutions ranging from 100 μg/mL down to 0.01 μg/mL [22]. The majority of the patients with severe reactions to peanuts had positive SPTs to Ara h 2 and Ara h 6, even at low concentrations. However, SPT reactions to Ara h 1 and Ara h 3, at higher concentrations, were also indicative for severe reactions [22]. In a pediatric study, the same set of natural peanut allergens was used for SPTs and provided a similar reactivity pattern as in the adult study group, with Ara h 2 and Ara h 6 inducing in most of the patients a positive reaction, while this was less pronounced for Ara h 1 and Ara h 3, respectively [23]. However, positive allergen-specific SPTs could not be related to the severity of peanut allergy.
Serum IgE Testing
Extract-based in vitro testing for specific IgE antibodies has been used for a long time and is still in use for routine diagnosis. In the past three decades, advanced molecular biology methods enabled a dramatic increase in our knowledge on individual allergens and their potential application in diagnosis and therapy. The application of individual purified allergens for in vitro diagnosis has been designated molecular diagnosis or CRD.
One application of purified, well-characterized allergens is their use as a certified reference material for existing and approved in vitro diagnostics. The European Union (EU)-funded project CREATE used well-defined inhalant allergens and provided a proof-of-concept study to improve standardization of existing extract-based diagnostics [24]. However, while this has been shown for inhalant allergies, no such tools are available for food allergy to date. One option is to spike a given food extract with one or several individual allergens. So far, only one example has entered the market, namely Cor a 1.04, the Bet v 1 homologous food allergen from hazelnut, which was added to hazelnut extract for improved in vitro diagnosis [25]. While the Cor a 1.04-specific IgE antibody detection improved upon spiking, no assay interference due to the spiking event was observed for the other allergens present in the hazelnut extract. In a clinical study, the results obtained with spiked hazelnut extract for specific IgE antibodies were compared with DBPCFCs in hazelnut-allergic children [26]. While the negative predictive value of in vitro diagnosis was improved with the spiked hazelnut extract, the positive predictive value was rather moderate to decreased [26].
Recently, CRD for hazelnut and peanut allergy was compared to the outcome of DBPCFCs. For peanut allergy, Eller and Bindslev-Jensen identified anti-Ara h 2 IgE >1.63 kU/L as a clear decision point with both optimal specificity and high sensitivity to predict clinically relevant peanut allergy [27••]. In addition, applying this cutoff value, the number of necessary DBPCFCs could be significantly reduced. In contrast, Beyer and co-authors identified 14.4 kU/L anti-Ara h 2 IgE to be 90 % predictive for a positive peanut challenge [28••]. For hazelnut positive challenge outcome, these authors calculated Cor a 14-specific IgE at 47.8 kU/L to be 90 % predictive [28••].
Basophil Activation Test
Cellular tests are available to verify the presence of allergen-specific IgE in patients’ sera and the ability to activate basophils. These tests either use autologous cells from the donor or established cell lines. After passive sensitization with serum and the addition of allergen extracts or allergens, cell activation can be measured by the determination of histamine or sulfidoleukotriene release. Alternatively, expression of a surface marker such as CD63 or CD203c is detected by fluorescence-activated cell sorting (FACS) analysis. CD63, a glycoprotein, is present only in low levels in resting basophils. However, upon activation, it is upregulated. Similarly, CD203c, an ecto-enzyme, is expressed on the cell membrane of human basophils and mast cells and becomes upregulated upon cross-linking of the Fc-epsilon receptor alpha. Erdmann and co-authors tested sera from 32 patients with Bet v 1-related plant food allergies. The recombinant allergens Api g 1 from celery, Dau c 1 from carrot, and Mal d 1 from apple were applied in the basophil activation test (BAT) assay, and CD63 upregulation was determined by FACS analysis. In this study, sensitivity for BAT ranged from 65 to 75 % and specificity from 68 to 100 % [29].
Sato and co-workers evaluated the CD203c expression on basophils from 71 egg- and milk-allergic children in the BAT assay [30]. In parallel, egg extract and Gal d 1, ovomucoid, and cow’s milk extract and Bos d 8, casein, were applied. The authors determined the CD203c stimulation index and the threshold of CD203c expression. In their study population, they found no difference in sensitivity, specificity, and positive predictive values (PPV), when comparing milk extract to Bos d 8. In hen’s egg allergy, sensitivity, specificity, and PPV for Gal d 1 were superior as compared to those in hen’s egg extract [30].
Similar studies were also performed in peanut-allergic children. Glaumann and co-authors evaluated the basophil allergen threshold sensitivity, called CD-sens, and peanut allergen-specific IgE antibody levels in relation to DBPCFC outcome [31•]. Basophil tests were performed with both peanut extract and purified rAra h 2 and yielded positive results in 92 % of the tested samples and were in agreement with a positive DBPCFC. Negative outcome in CD-sens was paralleled by negative results in DBPCFCs. Finally, specific IgE raised against Ara h 1, Ara h 2, and Ara h 3 was clearly associated with peanut allergy [31•].
Santos and co-workers compared the threshold of allergic reactions to peanuts with peanut extract basophil reactivity and identified clear associations [32].
For milk allergy, CRD was performed in milk-allergic patients and compared to the outcome of the basophil degranulation tests [33]. Despite specific IgE directed against milk allergens, the basophil degranulation test could discriminate between milk-allergic patients and patients still sensitized but that had already outgrown. Ford and co-workers performed another study, including milk-allergic children and assessed, whether they had already acquired tolerance by specific IgE testing and BAT assays using baked milk antigens. Again, the results from the BAT test proved useful in the identification of different phenotypes of the patients [34].
Food Allergen Panels for CRD
For some foods, a range of allergens are available for CRD, while for others, only single allergens have been identified to date. In the recent past, efforts were undertaken to perform CRD studies in well-characterized patients with positive DBPCFCs. These studies contribute to compile a patient-tailored risk profile and allow to relate distinct sensitization patterns with either mild or severe and/or generalized symptoms. They also allow to identify cross-sensitizations such as pollen allergens and help to discriminate between primary and secondary food allergy. In addition, these studies provide useful information about the sensitivity and specificity of these allergen-specific IgE testing. In the following, the selected examples of plant food allergen panels which are summarized in Table 1 will be briefly discussed.
Apple and Peach
Recent prevalence data for fruit allergies identified peach and apple as the most frequent sensitizers in Europe [35]. Four allergens have been officially designated for apple. These proteins were applied for CRD studies in the SAFE study. The Bet v 1 homologue, Mal d 1, was the major allergen in Central Europe (Netherlands, Austria) and linked with Fagales pollen allergy. In contrast, sensitization to the nsLTP, Mal d 3, was frequent in Southern Europe (Spain, Italy) and linked with severe allergic symptoms [36]. Sensitization to profilin, Mal d 4, is evenly distributed across Europe, associated mostly with grass and tree pollen allergy. For the thaumatin-like protein, Mal d 2, the dataset is less clear.
Peach allergy is dominant in Southern Europe and correlates with high consumption rates of fresh fruits throughout the year. Pru p 3, the nsLTP, is the major allergen in clinically relevant peach allergy [37] and accumulates in the peach peel. Boyano-Martinez assessed a high Pru p 3 sensitization rate in children and confirmed that the majority of the patients tolerated peach pulp [38••]. Sensitization to Pru p 3 starts earlier in life as compared to Pru p 1 and Pru p 4, the Bet v 1 homologue and the profilin, respectively [39].
Kiwifruit
The number of kiwifruit-allergic patients has remarkably increased in the last two decades, and the actual prevalence rate of sensitization in Europe is 5.2 % [35]. Since the extract-based IgE tests were of low sensitivity, efforts were undertaken to apply the existing kiwifruit allergen panel for CRD. Bublin and co-workers tested 237 sera from kiwifruit-allergic patients in the microarray format. The panel of allergens showed a diagnostic sensitivity of 66 %, a specificity of 56 %, and a positive predictive value of 73 % [40]. Actinidin, Act d 1, from kiwifruit is regarded as a marker for kiwifruit monosensitization and linked with generalized symptoms. In contrast, pollen-related kiwifruit allergy correlates with sensitization to the Bet v 1 homologue and profilin, Act d 8 and Act d 9, respectively [41]. Testing the large panel of different kiwifruit allergens showed that some allergens sensitize only in rare cases and may be specific for certain geographic area, such as Act d 6 [40]. In a European study, 311 sera from kiwifruit-allergic patients were tested. Comparing different geographic regions, sensitization to Act d 9, the kiwifruit nsLTP, was predominant for Southern Europe [42••]. Act d 1 was identified as a marker for severe symptoms upon kiwifruit consumption.
Vegetables
Celery is a frequent cause for food allergic reactions, especially in Europe with a sensitization prevalence of 6.3 % [35]. Food allergic symptoms can range from mild local reactions, usually restricted to the oral mucosa, up to severe anaphylactic reactions. Sensitization to the pollen-related allergens Api g 1, the birch pollen homologue, and Api g 4, the profilin, are usually found in patients with concomitant tree pollen allergy and account for rather mild symptoms. Api g 5 is a flavin adenine dinucleotide (FAD) containing oxidase and contains N-glycans, which account for the majority of its IgE binding capacity [43]. Api g 2 and Api g 6 are both nsLTPs, differently expressed throughout the plant tissue [44, 45]. So far, little is known about their relevance as major or minor allergens. However, it seems that the celeriac allergen panel is still incomplete; a marker allergen responsible for the mugwort–celery cross-reactivity, usually known to be linked with severe symptoms in allergic patients, is especially missing.
In parallel, carrot allergy is predominant in areas where Fagales pollen exposure is abundant. This cross-reactivity is due to the Bet v 1 homologue, Dau c 1, and the profilin, Dau c 4 [46••]. The diagnostic relevance of additional carrot allergens, such as Dau c 5, the isoflavone reductase, is not yet determined.
Tree Nuts
Among tree nuts, hazelnuts are a common source of food allergies, with and without concomitant pollen allergy with an overall sensitization prevalence of 9.3 % in Europe [35]. Hansen and co-authors investigated the sensitization profiles from 52 hazelnut-allergic patients from Denmark, Switzerland, and Spain [47]. While the Bet v 1 homologue, Cor a 1.04, was predominant in Northern and Central Europe, the sensitization to Cor a 8, the nsLTP from hazelnut, dominated in Spain. Profilin, Cor a 2, sensitization was evenly distributed in all countries (40–45 %). Only in 2 % sensitization to Cor a 11 was detected. Masthoff et al. investigated a total of 161 hazelnut-allergic patients, including children and adults, and identified sensitization to Cor a 14, the 2S albumin, and Cor a 9, the 11S globulin, as a marker for rather severe allergy symptoms [48••]. In a recent European study, Datema et al. used 7 hazelnut allergens to test sera from 423 hazelnut-allergic patients [49••]. IgE levels against Cor a 1 were the highest ones as compared to other hazelnut allergens. Cor a 8 sensitization was dominant in Greece and Spain. Cor a 14 sensitization was highly correlated with sensitization to Cor a 9. Cor a 11 sensitization was generally low, in all study centers. However, sensitization to Cor a 12, the oleosin from hazelnut, was detected in 10–25 % of the patients in most of the centers. Interestingly, sensitization to seed storage proteins and oleosins correlated with sensitization to other nuts, seeds, and pollens [49••].
Peanuts
Peanut is known as a highly allergenic food, tentatively inducing severe, even life-threatening symptoms. In some patients, even minute amounts of peanuts can induce allergic reactions. Therefore, improved in vitro diagnosis could help to reduce the number of DBPCFCs and help to determine individual threshold levels for the high-risk patients. Burney and co-authors identified a sensitization prevalence of 2.7 % [35], yet the prevalence of clinical relevant peanut allergy is expected to be around 1 % [50]. In order to get a deeper insight into actual population-based data on peanut allergy, Nicolaou and co-workers investigated the prevalence of peanut allergy versus tolerance in children at the age of 8 years (total 933 participants) [51••]. The sensitization profiles were assessed using peanut allergens, Ara h 1, Ara h 2, Ara h 3, and Ara h 8, and SPTs were performed with peanut extract. Within this collective, 11.8 % had specific IgE to at least one of the peanut allergens. However, only 22.4 % out of the sensitized individuals had a clinically relevant peanut allergy, as assessed by DBPCFCs. When correlating the sensitization profiles with the DBPCFC results, Ara h 2, the 2S albumin from peanut, was the best predictor for clinical peanut allergy. Since then, a number of studies confirmed the relevance of Ara h 2 as a predictor for peanut allergy in both pediatric and adult study groups [52, 53•, 27••, 28••]. When Ara h 6, another 2S albumin from peanut, became available for molecular diagnosis, it was found to be an equally important predictor for peanut allergy [54•, 55]. Specific IgE directed against Ara h 1, the 7S globulin, and Ara h 3, 11S globulin from peanut, are additional relevant markers for peanut allergy [56, 57•]. A recent European study analyzed the sensitization patterns of peanut allergy patients and confirmed the predictive value of spIgE against Ara h 2. Sensitization to Ara h 8 (Bet v 1 homologue) and Ara h 9 (nsLTP) was observed in different geographical regions and was more frequently found in peanut-tolerant subjects. Sensitization to Ara h 1 and Ara h 2 was exclusively observed in early onset of peanut allergy [57•].
Purified Food Allergens for Food Safety Aspects
In order to protect the allergic consumer from unintended exposure to allergenic food, legislation on allergen labeling has been set in place in many countries across the world. Consecutively, food industry has started initiatives to disseminate best practices for allergenic risk assessment and management among food producers. However, to implement risk assessment strategies during food production, processing, and packaging, reliable allergen detection assays and certified reference materials are still lacking. In addition, an agreed definition of a tolerable allergenic risk including threshold levels for each allergenic food is not yet in place [58]. For all these preventive measurements, purified well-defined food allergens can be used, and by replacing total extracts or spiking, total extracts with individual allergen test systems could be improved and provide more accurate determinations.
Conclusions
In the last two decades, our knowledge on individual allergens has dramatically increased. As a consequence, the application of well-defined molecules for in vitro diagnosis was started for the most important food allergen sources. While for some foods, the allergen panels are considerable complex, and for others, only a few allergens are identified yet. Nevertheless, it became evident that molecular diagnosis provides additional information as compared to conventional extract-based testing. It enables to identify cross-sensitization to inhalant allergen sources such as pollens. Furthermore, for some allergens, it emerged that they predominantly evoke mild allergic symptoms, while for others, they are inducers for severe symptoms. Nevertheless, these molecular diagnosis approach needs to be assessed on a food-by-food approach and the sensitivity and specificity of this test has to be compared to the conventional diagnostic tests, including the DBPCFCs. Therefore, more clinical studies are needed for assessing the relevance of molecular diagnosis for each food allergen source.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance •• Of major importance
Nwaru BI, Hickstein L, Panesar SS, Roberts G, Muraro A, Sheikh A. Prevalence of common food allergies in Europe: a systematic review and meta-analysis. Allergy. 2014;69(8):992–1007.
Nwaru BI, Hickstein L, Panesar SS, Muraro A, Werfel T, Cardona V, et al. The epidemiology of food allergy in Europe: a systematic review and meta-analysis. Allergy. 2014;69(1):62–75.
Muraro A, Werfel T, Hoffmann-Sommergruber K, Roberts G, Beyer K, Bindslev-Jensen C, et al. EAACI food allergy and anaphylaxis guidelines: diagnosis and management of food allergy. Allergy. 2014;69(8):1008–25.
Steckelbroeck S, Ballmer-Weber BK, Vieths S. Potential, pitfalls, and prospects of food allergy diagnostics with recombinant allergens or synthetic sequential epitopes. J Allergy Clin Immunol. 2008;121(6):1323–30.
Hoffmann-Sommergruber K, Mills EN, Vieths S. Coordinated and standardized production, purification and characterization of natural and recombinant food allergens to establish a food allergen library. Mol Nutr Food Res. 2008;52 Suppl 2:S159–65.
Sancho AI, Hoffmann-Sommergruber K, Alessandri S, Conti A, Giuffrida MG, Shewry P, et al. Authentication of food allergen quality by physicochemical and immunological methods. Clin Exp Allergy. 2010;40(7):973–86.
Hoffmann-Sommergruber K, Mills EN. Food allergen protein families and their structural characteristics and application in component-resolved diagnosis: new data from the EuroPrevall project. Anal Bioanal Chem. 2009;395(1):25–35.
Alessandri S, Sancho A, Vieths S, Mills CE, Wal JM, Shewry PR, et al. High-throughput NMR assessment of the tertiary structure of food allergens. PLoS One. 2012;7(7):e39785.
Chapman MD, Pomes A, Breiteneder H, Ferreira F. Nomenclature and structural biology of allergens. J Allergy Clin Immunol. 2007;119(2):414–20. doi:10.1016/j.jaci.2006.11.001.
Radauer C, Nandy A, Ferreira F, Goodman RE, Larsen JN, Lidholm J, et al. Update of the WHO/IUIS allergen nomenclature database based on analysis of allergen sequences. Allergy. 2014;69(4):413–9.
Breiteneder H, Radauer C. A classification of plant food allergens. J Allergy Clin Immunol. 2004;113(5):821–30.
Radauer C, Breiteneder H. Evolutionary biology of plant food allergens. J Allergy Clin Immunol. 2007;120(3):518–25.
Jenkins JA, Breiteneder H, Mills EN. Evolutionary distance from human homologs reflects allergenicity of animal food proteins. J Allergy Clin Immunol. 2007;120(6):1399–405.
Larenas-Linnemann D, Cox LS. European allergen extract units and potency: review of available information. Ann Allergy Asthma Immunol. 2008;100(2):137–45.
Sander I, Fleischer C, Meurer U, Bruning T, Raulf-Heimsoth M. Allergen content of grass pollen preparations for skin prick testing and sublingual immunotherapy. Allergy. 2009;64(10):1486–92.
Heinzerling L, Mari A, Bergmann KC, Bresciani M, Burbach G, Darsow U, et al. The skin prick test—European standards. Clin Transl Allergy. 2013;3(1):3.
Bolhaar ST, van de Weg WE, van Ree R, Gonzalez-Mancebo E, Zuidmeer L, Bruijnzeel-Koomen CA, et al. In vivo assessment with prick-to-prick testing and double-blind, placebo-controlled food challenge of allergenicity of apple cultivars. J Allergy Clin Immunol. 2005;116(5):1080–6.
Ma Y, Zuidmeer L, Bohle B, Bolhaar ST, Gadermaier G, Gonzalez-Mancebo E, et al. Characterization of recombinant Mal d 4 and its application for component-resolved diagnosis of apple allergy. Clin Exp Allergy. 2006;36(8):1087–96.
Garcia BE, Gonzalez-Mancebo E, Barber D, Martin S, Tabar AI, de Durana AM D, et al. Sublingual immunotherapy in peach allergy: monitoring molecular sensitizations and reactivity to apple fruit and Platanus pollen. J Investig Allergol Clin Immunol. 2010;20(6):514–20.
Kollmann D, Geroldinger-Simic M, Kinaciyan T, Huber H, Ebner C, Lidholm J, et al. Recombinant Mal d 1 is a reliable diagnostic tool for birch pollen allergen-associated apple allergy. J Allergy Clin Immunol. 2013;132(4):1008–10. This is an interesting proof of concept study—using recombinant Mal d 1, obtained by GMP conditions successfully for SPT—showing the potential of recombinant allergen testing in vivo.
Asero R. Peach-induced contact urticaria is associated with lipid transfer protein sensitization. Int Arch Allergy Immunol. 2011;154(4):345–8.
Peeters KA, Koppelman SJ, Penninks AH, Lebens A, Bruijnzeel-Koomen CA, Hefle SL, et al. Clinical relevance of sensitization to lupine in peanut-sensitized adults. Allergy. 2009;64(4):549–55.
Flinterman AE, van Hoffen E, den Hartog Jager CF, Koppelman S, Pasmans SG, Hoekstra MO, et al. Children with peanut allergy recognize predominantly Ara h 2 and Ara h 6, which remains stable over time. Clin Exp Allergy. 2007;37(8):1221–8.
Chapman MD, Ferreira F, Villalba M, Cromwell O, Bryan D, Becker WM, et al. The European Union CREATE project: a model for international standardization of allergy diagnostics and vaccines. J Allergy Clin Immunol. 2008;122(5):882–9 e2.
Andersson K, Ballmer-Weber BK, Cistero-Bahima A, Ostling J, Lauer I, Vieths S, et al. Enhancement of hazelnut extract for IgE testing by recombinant allergen spiking. Allergy. 2007;62(8):897–904.
Masthoff LJ, Pasmans SG, van Hoffen E, Knol MJ, Bruijnzeel-Koomen CA, Flinterman AE, et al. Diagnostic value of hazelnut allergy tests including rCor a 1 spiking in double-blind challenged children. Allergy. 2012;67(4):521–7. doi:10.1111/j.1398-9995.2011.02766.x.
Eller E, Bindslev-Jensen C. Clinical value of component-resolved diagnostics in peanut-allergic patients. Allergy. 2013;68(2):190–4. This study compares the outcome of molecular diagnosis and DBPCFCs and highlights that application of molecular diagnosis can reduce the number of necessary DBPCFCs considerably. In this study, a cutoff for Ara h 2 >1.63 kU/L would have reduced the number of challenges performed from 205 to 92.
Beyer K, Grabenhenrich L, Hartl M, Beder A, Kalb B, Ziegert M, et al. Predictive values of component-specific IgE for the outcome of peanut and hazelnut food challenges in children. Allergy. 2015;70(1):90–8. Prospective and multicenter German study, highlighting the predictive value of CRD in hazelnut and peanut allergy and reducing the number of necessary DBPCFCs accordingly.
Erdmann SM, Sachs B, Schmidt A, Merk HF, Scheiner O, Moll-Slodowy S, et al. In vitro analysis of birch-pollen-associated food allergy by use of recombinant allergens in the basophil activation test. Int Arch Allergy Immunol. 2005;136(3):230–8.
Sato S, Tachimoto H, Shukuya A, Ogata M, Komata T, Imai T, et al. Utility of the peripheral blood basophil histamine release test in the diagnosis of hen’s egg, cow’s milk, and wheat allergy in children. Int Arch Allergy Immunol. 2011;155 Suppl 1:96–103.
Glaumann S, Nopp A, Johansson SG, Rudengren M, Borres MP, Nilsson C. Basophil allergen threshold sensitivity, CD-sens, IgE-sensitization and DBPCFC in peanut-sensitized children. Allergy. 2012;67(2):242–7. In a peanut allergic collective, results were obtained from basophil test, molecular diagnosis, and DBPCFCs. When comparing the results, negative BAT outcome and spIgE testing for peanut allergens were identified as safe, time-saving, and cost-effective complements to DBPCFCs.
Santos AF, Du Toit G, Douiri A, Radulovic S, Stephens A, Turcanu V, et al. Distinct parameters of the basophil activation test reflect the severity and threshold of allergic reactions to peanut. J Allergy Clin Immunol. 2015;135(1):179–86.
Hochwallner H, Schulmeister U, Swoboda I, Spitzauer S, Valenta R. Cow’s milk allergy: from allergens to new forms of diagnosis, therapy and prevention. Methods. 2014;66(1):22–33.
Ford LS, Bloom KA, Nowak-Wegrzyn AH, Shreffler WG, Masilamani M, Sampson HA. Basophil reactivity, wheal size, and immunoglobulin levels distinguish degrees of cow’s milk tolerance. J Allergy Clin Immunol. 2013;131(1):180–6 e1-3. doi:10.1016/j.jaci.2012.06.003.
Burney PG, Potts J, Kummeling I, Mills EN, Clausen M, Dubakiene R, et al. The prevalence and distribution of food sensitization in European adults. Allergy. 2014;69(3):365–71.
Fernandez-Rivas M, Bolhaar S, Gonzalez-Mancebo E, Asero R, van Leeuwen A, Bohle B, et al. Apple allergy across Europe: how allergen sensitization profiles determine the clinical expression of allergies to plant foods. J Allergy Clin Immunol. 2006;118(2):481–8.
Fernandez-Rivas M, Gonzalez-Mancebo E, Rodriguez-Perez R, Benito C, Sanchez-Monge R, Salcedo G, et al. Clinically relevant peach allergy is related to peach lipid transfer protein, Pru p 3, in the Spanish population. J Allergy Clin Immunol. 2003;112(4):789–95.
Boyano-Martinez T, Pedrosa M, Belver T, Quirce S, Garcia-Ara C. Peach allergy in Spanish children: tolerance to the pulp and molecular sensitization profile. Pediatr Allergy Immunol. 2013;24(2):168–72. Oral food challenges were performed and results compared with CDR and SPTs using purified nPru p 3. Even with Pru p 3 sensitization, only a subset of patients reacted with peach during challenges. In addition, most of the patients tolerated peach pulp—showing that Pru p 3 is enriched in peach peel.
Pastorello EA, Farioli L, Stafylaraki C, Mascheri A, Scibilia J, Pravettoni V, et al. Anti-rPru p 3 IgE levels are inversely related to the age at onset of peach-induced severe symptoms reported by peach-allergic adults. Int Arch Allergy Immunol. 2013;162(1):45–9.
Bublin M, Dennstedt S, Buchegger M, Antonietta Ciardiello M, Bernardi ML, Tuppo L, et al. The performance of a component-based allergen microarray for the diagnosis of kiwifruit allergy. Clin Exp Allergy. 2011;41(1):129–36.
Bublin M, Pfister M, Radauer C, Oberhuber C, Bulley S, Dewitt AM, et al. Component-resolved diagnosis of kiwifruit allergy with purified natural and recombinant kiwifruit allergens. J Allergy Clin Immunol. 2010;125(3):687–94. 94 e1.
Le TM, Bublin M, Breiteneder H, Fernandez-Rivas M, Asero R, Ballmer-Weber B, et al. Kiwifruit allergy across Europe: clinical manifestation and IgE recognition patterns to kiwifruit allergens. J Allergy Clin Immunol. 2013;131(1):164–71. This European study included 311 individuals from outpatient clinics. CRD highlighted different sensitization patterns in different geographic regions and correlated sensitization to Act d 1 with severe symptoms.
Bauermeister K, Ballmer-Weber BK, Bublin M, Fritsche P, Hanschmann KM, Hoffmann-Sommergruber K, et al. Assessment of component-resolved in vitro diagnosis of celeriac allergy. J Allergy Clin Immunol. 2009;124(6):1273–81. e2.
Gadermaier G, Egger M, Girbl T, Erler A, Harrer A, Vejvar E, et al. Molecular characterization of Api g 2, a novel allergenic member of the lipid-transfer protein 1 family from celery stalks. Mol Nutr Food Res. 2011;55(4):568–77.
Vejvar E, Himly M, Briza P, Eichhorn S, Ebner C, Hemmer W, et al. Allergenic relevance of nonspecific lipid transfer proteins 2: identification and characterization of Api g 6 from celery tuber as representative of a novel IgE-binding protein family. Mol Nutr Food Res. 2013;57(11):2061–70.
Ballmer-Weber BK, Skamstrup Hansen K, Sastre J, Andersson K, Batscher I, Ostling J, et al. Component-resolved in vitro diagnosis of carrot allergy in three different regions of Europe. Allergy. 2012;67(6):758–66. Diagnosis of carrot allergy was assessed by DBPCFCs and CRD test sensitivity and specificity determined and compared to extract-based approach. This study highlighted different sensitization patterns in different geographic regions and between carrot-tolerant and carrot-allergic individuals.
Hansen KS, Ballmer-Weber BK, Sastre J, Lidholm J, Andersson K, Oberhofer H, et al. Component-resolved in vitro diagnosis of hazelnut allergy in Europe. J Allergy Clin Immunol. 2009;123(5):1134–41. 41 e1-3.
Masthoff LJ, Mattsson L, Zuidmeer-Jongejan L, Lidholm J, Andersson K, Akkerdaas JH, et al. Sensitization to Cor a 9 and Cor a 14 is highly specific for a hazelnut allergy with objective symptoms in Dutch children and adults. J Allergy Clin Immunol. 2013;132(2):393–9. An important study showing the diagnostic value of spIgE against Cor a 9 and Cor a 14 for hazelnut allergic patients, both in adults and in children.
Datema MR, Zuidmeer-Jongejan L, Asero R, Barreales L, Belohlavkova S, de Blay F, et al. Hazelnut allergy across Europe dissected molecularly: a EuroPrevall outpatient clinic survey. J Allergy Clin Immunol. 2015. Prospective study recruiting participants form outpatient clinics. A subset of those underwent DBPCFCs. In parallel, sensitization pattern to 17 hazelnut allergens was assessed, and in addition, spIgE levels of Cor a 1, Cor a 2, Cor a 8, Cor a 9, Cor a 11, and Cor a 14 also Cor a 12 were assessed.
Rona RJ, Keil T, Summers C, Gislason D, Zuidmeer L, Sodergren E, et al. The prevalence of food allergy: a meta-analysis. J Allergy Clin Immunol. 2007;120(3):638–46.
Nicolaou N, Poorafshar M, Murray C, Simpson A, Winell H, Kerry G, et al. Allergy or tolerance in children sensitized to peanut: prevalence and differentiation using component-resolved diagnostics. J Allergy Clin Immunol. 2010;125(1):191–7. e1-13. Birth cohort study, investigating peanut allergy versus peanut sensitization in 8-year-old children.
Hong X, Caruso D, Kumar R, Liu R, Liu X, Wang G, et al. IgE, but not IgG4, antibodies to Ara h 2 distinguish peanut allergy from asymptomatic peanut sensitization. Allergy. 2012;67(12):1538–46.
Klemans RJ, Liu X, Knulst AC, Knol MJ, Gmelig-Meyling F, Borst E, et al. IgE binding to peanut components by four different techniques: Ara h 2 is the most relevant in peanut allergic children and adults. Clin Exp Allergy. 2013;43(8):967–74. This is the first part of two studies dealing with the diagnostic value of spIgE against Ara h 2 as a marker of peanut allergy.
Klemans RJ, Knol EF, Bruijnzeel-Koomen CA, Knulst AC. The diagnostic accuracy of specific IgE to Ara h 6 in adults is as good as Ara h 2. Allergy. 2014;69(8):1112–4. This is the second part of a peanut study, identifying the role of Ara h 6, another 2S albumin from peanut, in in vitro diagnosis.
Ackerbauer D, Bublin M, Radauer C, Varga EM, Hafner C, Ebner C, et al. Component-resolved IgE profiles in Austrian patients with a convincing history of peanut allergy. Int Arch Allergy Immunol. 2015;166(1):13–24.
Ebisawa M, Moverare R, Sato S, Maruyama N, Borres MP, Komata T. Measurement of Ara h 1-, 2-, and 3-specific IgE antibodies is useful in diagnosis of peanut allergy in Japanese children. Pediatr Allergy Immunol. 2012;23(6):573–81.
Ballmer-Weber BK, Lidholm J, Fernandez-Rivas M, Seneviratne S, Hanschmann KM, Vogel L, et al. IgE recognition patterns in peanut allergy are age dependent: perspectives of the EuroPrevall study. Allergy. 2015;70(4):391–407. European study on peanut allergy, including patients with positive DBPCFCs. Geographic and age-related differences were observed. Role of Ara h 2 as a marker allergen confirmed; sensitization to Ara h 1, 2, and 3 is acquired in childhood.
Muraro A, Hoffmann-Sommergruber K, Holzhauser T, Poulsen LK, Gowland MH, Akdis CA, et al. EAACI food allergy and anaphylaxis guidelines. Protecting consumers with food allergies: understanding food consumption, meeting regulations and identifying unmet needs. Allergy. 2014;69(11):1464–72.
Palacin A, Rodriguez J, Blanco C, Lopez-Torrejon G, Sanchez-Monge R, Varela J, et al. Immunoglobulin E recognition patterns to purified kiwifruit (Actinidinia deliciosa) allergens in patients sensitized to kiwi with different clinical symptoms. Clin Exp Allergy. 2008;38(7):1220–8.
Acknowledgments
Karin Hoffmann-Sommergruber and Sabine Pfeifer were supported by the Austrian Science Fund (grant SFB F4603-B19).
Compliance with Ethics Guidelines
ᅟ
Conflict of Interest
Drs. Hoffmann-Sommergruber, Pfeifer, and Bublin declare no conflicts of interest.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article is part of the Topical Collection on Immunologic/Diagnostic Tests in Allergy
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Hoffmann-Sommergruber, K., Pfeifer, S. & Bublin, M. Applications of Molecular Diagnostic Testing in Food Allergy. Curr Allergy Asthma Rep 15, 56 (2015). https://doi.org/10.1007/s11882-015-0557-6
Published:
DOI: https://doi.org/10.1007/s11882-015-0557-6