
Abstract
We examined the justifications invoked by the German government in April 2009 to suspend the cultivation of the genetically modified maize varieties containing the Bt insect-resistance trait MON810. We have carried out a critical examination of the alleged new data on a potential environmental impact of these varieties, namely two scientific papers describing laboratory force-feeding trials on ladybirds and daphnia, and previous data on Lepidoptera, aquatic and soil organisms. We demonstrate that the suspension is based on an incomplete list of references, ignores the widely admitted case-by-case approach, and confuses potential hazard and proven risk in the scientific procedure of risk assessment. Furthermore, we did not find any justification for this suspension in our extensive survey of the scientific literature regarding possible effects under natural field conditions on non-target animals. The vast majority of the 41 articles published in 2008 and 2009 indicate no impact on these organisms and only two articles indicate a minor effect, which is either inconsistent during the planting season or represents an indirect effect. Publications from 1996 to 2008 (376 publications) and recent meta-analyses do not allow to conclude on consistent effects either. The lower abundance of some insects concerns mainly specialized enemies of the target pest (an expected consequence of its control by Bt maize). On the contrary, Bt maize have generally a lower impact than insecticide treatment. The present review demonstrates that the available meta-knowledge on Cry1Ab expressing maize was ignored by the German government which instead used selected individual studies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
MON810 is a transgenic trait introgressed into a number of maize varieties, which consists of a Bacillus thuringiensis-derived gene (Bt), namely a truncated cry1Ab gene encoding an insecticidal protein (δ-endotoxin) for control of some lepidopteran pest insects such as Ostrinia nubilalis, the European corn borer. This trait also offers protection against certain moths during maize grain storage (Hubert et al. 2008).
The Federal Office of Consumer Protection and Food Safety of the German government suspended, on 17 April 2009, the permit allowing the cultivation of the genetically modified (GM) maize varieties containing the MON810 trait. The official German suspension order (GSO) is based on Sect. 20 paragraph 2 of the Gentechnikgesetz (German Act on Genetic Engineering). The GSO states that the responsible German higher federal authority is authorized to suspend such a permit when there is justified reason to assume that the GM organism poses a hazard to human health or the environment. Such a justified reason needs to be based on new or additional scientific information becoming available. According to the GSO, this new and additional information gives rise to “the justified assumption that the cultivation of MON810 is a hazard (Gefahr) to the environment”, namely to non-target arthropods (www.bvl.bund.de/cln_027/DE/08__PresseInfothek/00__doks__downloads/mon__810__bescheid,templateId=raw,property=publicationFile.pdf/mon_810_bescheid.pdf).
A proper scientific risk assessment of the impact of Bt maize cultivation on non-target arthropods requires consideration of both the hazard (expression of toxicity) and the likelihood of exposure to the hazard (i.e. the Cry1Ab toxin). Laboratory studies are a useful initial step in the risk assessment of the impact of insect-resistant GM crops which allows the characterization of hazards to the environment. The exposure to potential hazards then needs to be assessed in realistic situations, namely under natural field conditions, before conclusions can be drawn on the risk presented by these crops (Romeis et al. 2006a, 2008). This is illustrated by the Monarch butterfly case (see Minorsky 2001). Laboratory assays (Losey et al. 1999; Jesse and Obrycki 2000) showed that Monarch larvae suffered higher mortality when reared on milkweed leaves dusted with pollen from Bt maize compared to non-transformed maize. However, a more complete risk assessment, including field studies, indicated that the effects of Bt maize cultivation on Monarch populations are negligible in the case of MON810, but not Bt176 (Hellmich et al. 2001; Pleasants et al. 2001; Sears et al. 2001; Stanley-Horn et al. 2001; Tschenn et al. 2001; Zangerl et al. 2001; Anderson et al. 2005; Dively et al. 2004). The Bt176 trait is also a Cry1Ab type but this GM maize (which is no longer cultivated) contains toxin levels 12–80 times higher in the pollen compared to MON810 (US EPA 2000). Thus, the Monarch case also illustrates the absolute requirement of a case-by-case risk assessment.
Since the German authorities had partially suspended and re-authorized cultivation of MON810 varieties in 2007, it has to be assumed that “new information” means that further scientific evidence has been made available since that date. Therefore, in the present article, we first provide a critical assessment of the new publications cited by the GSO document, namely Bohn et al. (2008) and Schmidt et al. (2009), which used daphnia and ladybirds, respectively, in laboratory experiments. Secondly, since the GSO surprisingly also cites earlier references, we also examine them in a scientific risk assessment perspective, along with the scientific literature (available when the GSO was issued) on the impact of the Cry1Ab-expressing maize varieties. We discuss the actual scientific evidence available in the literature and that which has been actually used for this policy-decision and examine the way the empirical validity of scientific knowledge has been taken into account.
Critical examination of a laboratory feeding study using Daphnia magna
Bohn et al. (2008) performed laboratory studies consisting of feeding daphnia (crustacean arthropods) with 100% ground maize suspension derived from either a GM (Bt) or a conventional cultivar. The methodology of Bohn et al. (2008) suffers from several weakness. First, the authors used a hybrid variety containing the MON810 trait (termed Dekalb 818 Yieldgard) and, as a control, a local (from the Philippines) variety termed Dekalb 818. No information is provided as to whether these lines are near isogenic. Secondly, the test material was not characterized for potential nutritional differences. Thirdly, in order to determine the effect of GM plants it is important in any research to not only compare the GM plant with its corresponding isogenic line but also to compare the effect between at least two non-isogenic lines. These comparisons would indicate whether the effect observed between the GM plant and its isogenic line is linked to the genetic modification or whether it is caused by other traits. Fourthly, no data were provided on the dose of toxins the daphnia were exposed to (and how much they ingested), which prevents any comparison with the exposure dose under field conditions.
The reported survival curves are characterized by a high mortality of daphnia, even when fed with conventional maize (about 55–90% mortality after ca. 40 days). It is therefore reasonable to consider only shorter experimental time points (without high mortality of control animals) to aim for a biological valid result. When these time points are considered (up to 14–21 days depending on the experiments), no statistically significant difference in mortality was observed between GM and non-GM feed. Another parameter considered by the authors (proportion of females reaching maturity) is dependent on survival, whereas fecundity parameters provided inconsistent results (some improved with the GM feed).
In conclusion, these laboratory results appear inconclusive: varietal effects (with no link to the transgenic toxin) that may affect feed composition cannot be excluded; reported differences in the survival rate of daphnia are not convincing due to the experimental design of the test. Furthermore, as the authors themselves admit “… this is an artificial situation. However, our study was not aimed at estimating the responses of D. magna under natural field conditions, where they would have a diverse diet.”
We found no other publication in 2008 and 2009 dealing with the impact of Bt maize on daphnia.
Critical examination of a laboratory toxicological test using Adalia bipunctata
Schmidt et al. (2009) performed tests consisting of measuring the mortality rate of preimaginal larval stages, development time of these stages and body weights of ladybirds fed in the laboratory in the presence of different concentrations of Cry1Ab or Cry3Bb.
Again, their methodology suffers from several weakness. First, the authors used protoxins produced in E. coli and trypsin-digested to yield an active toxin. They refer to Bosch et al. (1994) who refer to two further publications. However, none of these publications characterizes the bioactivity of the trypsinized protoxins in sensitive insect bioassays. No details are provided concerning the activity of the cleaved toxin and on the possible formation of unusual peptides as a consequence of the trypsic digestion, nor on the composition of the bacterial extracts, which do not seem to consist of pure protoxins. Secondly, ladybird larvae were fed Ephestia kuhniella eggs (a moth) sprayed with trypsinized Cry protoxins, but no data are provided on the dose of toxins ingested by the ladybirds. This is particularly important since first instars of ladybird larvae bite in their prey and suck out the liquid, which implies they are unlikely to ingest high amounts of Cry toxins sprayed on the prey’s surface. This unfortunately prevents any comparison with the exposure dose under field conditions where ladybirds feed on herbivores (which in turn have been feeding on maize) or directly on maize pollen.
Regarding the results, no statistically significant difference was reported for the development time of preimaginal larval stages, nor for body mass of newly emerged adults. Concerning the authors’ claim on higher larvae mortality when fed Cry1Ab, it should first be mentioned that these are not new data since they have been already published (Schmidt et al. 2004). It should be noted that mean mortality rates of control larvae (no toxin sprayed) vary from 7.5 to 20.8% in the three experiments presented (termed ‘a’, ‘b’ and ‘c’ in Table 1 published by Schmidt et al. 2009). This puts into question on the biological relevance of a mean mortality rate of 24.2% as reported in experiment ‘a’ for the lowest concentration of Cry1Ab (5 μg/ml). Furthermore, at higher doses the data appear inconsistent since mean mortality rate does not increase, but actually decreases, when the Cry1Ab concentration in the spray solution increases from 25 to 50 μg/ml.
In conclusion, the authors do not provide convincing data to support their claim of a toxicity of the Cry1Ab trypsinized protoxin for ladybirds (which are Coleopterans, whereas the bona fide toxin is considered specific for Lepidopterans): they used an insufficiently characterized laboratory test and their results on mortality are weakened by a high variability of the crucial parameter. Consequently, this article does not allow the drawing of ecotoxicological relevant conclusions on a negative impact of maize MON810 on ladybirds in a field setting where these insects actually feed mainly on aphids (which, in fact, contain little toxin).
Critical examination of the GSO arguments on the ecotoxicological effects of MON810
Coleoptera
The GSO refers solely to Schmidt et al. (2009), which is discussed above.
Lepidoptera
The GSO refers to Losey et al. (1999) and Jesse and Obrycki (2000) who concluded a potential threat to the Monarch butterfly in laboratory force-feeding experiments (see Introduction). The GSO cites a publication by Hellmich et al. (2001) which describes laboratory tests to establish the relative toxicity of various Cry toxins: their data suggest that pollen from the Cry1Ab varieties Bt11 and MON810 would have no acute effects on the Monarch butterfly in a field setting and that only the Bt176 varieties can affect these butterflies, a conclusion confirmed by Zangerl et al. (2001), also cited in the GSO, who found sublethal effects of Bt176 on black swallowtails (Papilio polyxenes) in the field. Surprisingly, the GSO does not mention Wraight et al. (2000) who, in contrast, failed to detect a harmful effect of MON810 pollen on black swallowtails. Dively et al. (2004) studied continuous exposure to pollen (during maize anthesis) of Monarch larvae in force-feeding trials. They found that 24% fewer larvae exposed to Bt pollen reached the adult stage. However, taking into account the high natural mortality of larvae and the proportion of larvae actually exposed to pollen, they calculated that the risk associated with long term exposure is only 0.6% additional mortality, a conclusion, again, not mentioned in the GSO.
The GSO also cites a number of publications by Felke and coworkers (see the German federal website www.gmo-safety.eu/en/safety_science/136.docu.html). A summary report by Felke and Langenbruch (2005) states: “We recommend only permitting cultivation of Bt-Maize with negligible toxin expression in pollen”, which is the case of MON810. The authors did not demonstrate harmful effects on butterflies with MON810 in contrast to Bt176.
The most recent references cited in the GSO are laboratory studies, namely Mattila et al. (2005) who used a MON810 hybrid producing two toxins (Cry1Ab and Cry2Ab2) and MON863 producing Cry3Bb1 (with force-feeding results similar to those of Dively et al. 2004), and Lang and Vojtech (2006) who used again Bt176 maize.
In conclusion, the GSO does not provide any evidence for deleterious effects of MON810 on Lepidoptera under field conditions. In addition, the GSO uses arguments concerning Bt176 to argue for a suspension of MON810 varieties, which is incorrect since scientific risk assessment follows a case-by-case procedure.
Aquatic organisms
The GSO mentions the publication by Rosi-Marshall et al. (2007) suggesting that toxins in transgenic crop byproducts may affect headwater stream ecosystems. Maize byproducts were measured in headwater streams adjacent to maize fields. This work also showed the presence of maize pollen grain in trichopteran insects collected in such streams. However, this work did not report deleterious effects on these insects under natural conditions. Only laboratory conditions (force-feeding experiments) with two trichopteran species (caddisflies) were reported. They showed no effect on mortality for either Lepidostoma liba fed maize litter or for Helicospyche borealis fed maize pollen at the highest concentration measured in water streams. In the latter case, a higher mortality of H. borealis was observed only at higher pollen concentrations, and in the former, L. liba had a 50% reduced growth rate.
In two letters, five scientists criticized this article on several grounds, including the fact that the control maize was not isogenic to the Bt maize, raising the possibility that the observed differences were due to differences in the chemical composition of the leaves, with no link to the Cry1Ab toxin (Beachy et al. 2008; Parrott 2008). Rosi-Marshall et al. (2008) replied to these criticisms agreeing on the point that their article should not have suggested their observations have “ecosystem-scale consequences”. Indeed, the same laboratory failed to observe any effect on growth or mortality for two trichopteran species during in situ experiments (Pokelsek et al. 2007).
Griffiths et al. (2009) studied headwater streams draining agricultural landscapes and receiving maize leaves via wind and surface runoff and examined how substrate quality and in-stream nutrient concentrations influenced microbial respiration on maize. They found that Bt maize had a faster decomposition rate than non-Bt maize, while microbial respiration rates did not differ between Bt and non-Bt maize. Decomposition rates were not negatively affected by genetic engineering, most likely because the Bt toxin does not adversely affect the aquatic microbial assemblage involved in maize decomposition. Additionally, shredding caddisflies, which were studied by Rosi-Marshall et al. (2007), were depauperate in these agricultural streams, and most likely did not play a major role in maize decomposition.
The GSO also mentions a publication by Bohn et al. (2008), which was examined above, and by Douville et al. (2007). The latter study measured the persistence of DNA (not insecticidal proteins) in surface water. The half-life of the cry1Ab DNA was identical when produced by Bt maize or by B. thuringiensis (widely used as a biopesticide). Traces of transgenic DNA (0–0.0000005 ng/l) were found in a river up to 82 km from a field (however, the authors did not demonstrate that it originated from this actual field). The same surface water samples contained 0–30 ng/l total DNA. It is unclear why the mere presence of DNA, a normal component of terrestrial and aquatic ecosystems, should be viewed as a threat to these ecosystems.
Douville et al. (2005) published another similar study which measured levels of the insecticidal protein in surface water. They found that the Cry1Ab protein produced by Bt maize disappeared more rapidly than its counterpart produced by B. thuringiensis (which was expected from its crystal structure). The authors concluded that this protein is “fairly uncommon” in aquatic environments.
Soil organisms
The GSO refers to a poster by Büchs et al. (2004) on the effects observed in laboratory trials involving sciarid fly larvae fed on pollen or plant parts from different maize varieties. Two Bt varieties and their respective isogenic parent varieties were involved in the study, alongside a third conventional variety. The authors stressed only one of the results: sciarid fly larvae fed on straw from MON810 Bt maize took longer to pupate (ca. 21 days instead of ca. 18 on average). However, no correlation was found between the observed effect and the absolute toxin content of different maize varieties. This effect may, therefore, be linked to the nutritional quality of the feed. Similar effects were also observed between different conventional varieties in the same experiments. In an interview (www.gmo-safety.eu/en/maize/soil/308.docu.html), the authors reported inconsistent trends in field experiments over 3 years and suggested “closer observation in post-market monitoring”.
In summary, the references cited in the GSO do not validate its claim on ecotoxicological effects of MON810 maize on non-target arthropods. Since the GSO appears to be based on an incomplete list of references, we provide a more complete literature survey below.
Review of the current scientific literature on the impact of Cry1Ab expressing maize varieties
Recent publications (2008 and 2009)
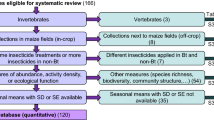
In our database of 15,000 references on transgenic plants collected since 1996 there are 683 references from 2009 (ended 31 April 2009) and 2293 from 2008. For these 2 years, using keywords ‘Maize or Corn’ and ‘Insect and Resistance’, 171 peer-reviewed references were selected, 137 from 2008 and 34 from 2009 (see Electronic supplementary material). These references are gathered as follows in the different fields of investigation (see Table 1).
Regarding the distribution of recent publications on the impact on non-target arthropods, we observe that the vast majority of the articles dealing with MON810 or Cry1 (n = 23) indicates that this ‘event’ has no impact on these organisms. Only two articles (noted “a” in the table) indicate a deleterious effect on non-target arthropods (Górecka et al. 2008; Zenner de Polania and Alvarez Alcaraz 2008). The other articles do not deal with Cry1.
Górecka et al. (2008) measured the braconid Aphidius colemani population when feeding on bird cherry-oat aphid (Rhopalosiphum padi), which were feeding on Bt or non-Bt maize under greenhouse conditions. In an experiment during the summer, they found a higher A. colemani population on Bt-fed aphids, while they found the opposite during the winter experiment. The authors conclude that “the observed effect of season on parasitation level by A. colemani on R. padi host feeding on Bt and non-Bt maize plants indicates that results obtained in a single greenhouse experiment may lead to questionable conclusions and should be confirmed by other experiments”.
Zenner de Polania and Alvarez Alcaraz (2008) carried out field observations and laboratory studies for 3 years to evaluate the effect, direct or indirect, of Bt maize (‘Yieldgard’ hybrid) on the natural enemies of the main insect pest, Spodoptera frugiperda. Their results indicated that the Cry1Ab toxin exhibits an indirect effect on these beneficial insect populations. They observed a tendency for a decrease in S. frugiperda populations in commercial fields of both transgenic and conventional varieties. The responsible factors are insecticidal applications and the control of the prey/host (S. frugiperda) by Bt maize varieties.
Survey of field evaluation of Cry1Ab-producing maize on ladybirds from 1996 to 2009
Our review leads to 75 articles. Among them, 27 concern Bt maize and ladybirds, of which 21 indicated that there is no effect of Bt maize on ladybirds. The remaining six articles mention some effects of Bt proteins on non-target organisms such as ladybirds and are discussed below. Among them, the publication of Schmidt et al. (2009) has already been discussed above.
Wold et al. (2001) found no significant within-year differences in the overall density of beneficial insect populations including predatory coccinellids (Adalia bipunctata and Coleomegilla maculata), chrysopids and anthocorids between Bt and non-Bt sweet corn. Note that A. bipunctata was also used in the study of Schmidt et al. (2009). Although the test of mean density of insects per three plants in open plots detected a significant trend for a higher density of C. maculata in non-Bt maize (1.92) compared to Bt maize (1.17), the results were inconsistent from 1 year to another and it is unclear whether it is actually linked to the Bt trait.
Delrio et al. (2004) showed in field studies that the infestation of aphids, mainly Rhopalosiphum padi, was similar in both Bt and non-Bt plots. Similarly, no significant difference was found between Bt and non-Bt maize for predator counts (arachnids, mirids, anthocorids, syrphids, chrysopids and coccinellids) on sample plants. However, when predator abundance was estimated by their capture on yellow traps, a single difference was reported, namely in the total number of coccinellids, mainly Propylea quatuordecimpunctata, which was significantly lower in Bt plots.
Toth et al. (2004) compared the applicability of two arthropod sampling methods, namely “spider-web survey” and “whole plant visual sampling”, in a risk assessment study. Both methods were able to detect significant differences in the quantity of predatory insects in Bt (MON810) versus isogenic plots, but not in the same taxa. More Nabidae were found in Bt versus isogenic plots using the spider-web method, while more Coccinellidae were found in isogenic plots versus Bt using the plant sampling method. The authors conclude that “due to several biological uncertainties, interpretation and explanation of the results remain problematic”.
Hoheisel and Fleischer (2007) showed the influence of concurrent introduction of three transgenic vegetable varieties on the seasonal dynamics of coccinellids and their food, aphids and pollen, examined within diversified farm systems practicing insect pest management. The transgenic varieties used included sweet corn, potato and winter squash expressing Cry1Ab, Cry3A and plant viral coat proteins targeting Lepidoptera, Coleoptera and aphid-transmitted viruses, respectively. Transgenic systems reduced insecticides by 25%. Weekly differences in coccinellid density between transgenic and isogenic crops were rare and transitory, governed by the timing of planting or foliar insecticide use patterns.
Recently, under controlled conditions, Moser et al. (2008) confirmed that two species of coccinellids can feed directly on maize. Development time of one species increased after Bt maize treatments compared with non-Bt corn treatments.
Survey of field evaluation of Cry1Ab-producing maize on other non-target arthropods from 1996 to 2009
Our systematic review leads to 376 articles. Among these, 8 mentioned effects.
Orr and Landis (1997) recorded predation and parasitism of the European corn borer in Cry1Ab and isogenic maize fields. Levels of egg mass predation and parasitism, density of European corn borer predators and parasitism of larvae were not significantly different between the transgenic and isogenic plots. All observed differences in natural enemy population parameters were opposite to expected if transgenic plants had an adverse impact.
Bourguet et al. (2002) performed a field experiment at two sites comparing the temporal abundance of non-target arthropods in Bt maize (MON810 hybrid) and non-Bt maize. The non-target insects studied included the aphids Metopolophium dirhodum, Rhopalosiphum padi and Sitobion avenae, the bug Orius insidiosus, the syrphid Syrphus corollae, the ladybird Coccinella septempunctata, the lacewing Chrysoperla carnea (Stephens), thrips and hymenopteran parasitoids. For all but one species, the number of individuals varied greatly over the season but did not differ between the maize types. The only exception was thrips which, at one site, were significantly more abundant in Bt maize than in non-Bt maize. However, this difference did not remain significant when the multiple tests were taken into account.
Candolfi et al. (2004) found no effect of Bt maize on the communities of soil-dwelling and non-target plant-dwelling arthropods. A trend towards a community effect on flying arthropods was observed with a lesser abundance of adult Lepidoptera, flies in the families Lonchopteridae, Mycetophilidae and Syrphidae, and the hymenopteran parasitoids Ceraphronidae. However, the effects were slight and restricted to two sampling dates corresponding to anthesis. In contrast, a short but statistically significant effect of two insecticides was observed on the community of plant dwellers and a prolonged effect of one insecticide on the soil dwellers.
Pilcher et al. (2005), when measuring adult populations of five predator and one parasitoid species in maize plots in three locations in Iowa over several years, found few differences in abundance of the generalist predators between Bt and non-Bt maize. However, the specialist parasitoid of the European corn borer was significantly affected in Bt plots, which was not unexpected given its dependence on the presence of the European corn borer.
Bruck et al. (2006) found less Macrocentrotus cingulum and Nitidulidae in transgenic plots (MON810) probably due to the absence of the European corn borer which serves as a host for M. cingulum and provides a habitat for Nitidulidae, which are known to frequent European corn borer tunnels. Application of a conventional insecticide for European corn borer control had a broader impact on populations of various non-target arthropods.
Griffiths et al. (2007) found no effect of Bt maize (MON810) on microarthropods, enchytreids and earthworms, but they found a difference concerning the microbial community structure. However, this difference was not persistent and could not be distinguished from a varietal effect. They concluded that there were no soil ecological consequences for these communities associated with the use of Bt maize in place of conventional varieties. Other land management options, such as tillage, crop type and pest management regime, had significantly greater effects on the biology of the soil rather than the type of maize grown.
Toschki et al. (2007) recorded the activity abundance of spiders and carabid beetles in Bt maize (MON810), an isogenic variety and the isogenic variety treated with insecticide. Significantly different activity abundances in Bt plots compared with isogenic control plots were observed both for spiders and carabid beetles during 2001. However, in 2002 and 2003 they found no changes in community structure with any of the treatments. They hypothesized that the change in the first year may have been caused by the influence of a massive corn borer infestation and accompanying large changes in microclimatic factors.
Musser and Shelton (2003) found a positive effect on the Bt transgenic pest control in sweet corn. In field trials they found that transgenic Bt sweet corn and also the foliar insecticides indoxacarb and spinosad are all less toxic to the most abundant predators in sweet corn (Coleomegilla maculate, Harmonia axyridis, and Orius insidiosus) than the pyrethroid lambda cyhalothrin. Indoxacarb, however, was moderately toxic to coccinellids and spinosad, and indoxacarb was somewhat toxic to O. insidiosus nymphs at field rates. Their results “demonstrate that some of the new products available in sweet corn allow a truly integrated biological and chemical pest control program in sweet corn, making future advances in conservation, augmentation and classical biological control more feasible”.
One should keep in mind that, compared to these 7 publications mentioning some minor negative effects, there are 37 other papers indicating no effect of Bt maize on non-target organisms. Nevertheless, in order to clarify whether the effects mentioned occasionally in research studies represent a real trend or not, we have examined the conclusions of recently published meta-analyses.
Recent comprehensive reviews and meta-analyses of the impact of Cry1Ab-producing maize on non-target arthropods
We will focus here on field experiments. However, it should be mentioned that Lövei et al. (2009) reviewed 55 laboratory studies on the impact of GM plants on arthropod natural enemies. They conclude that these proteins “often have non-neutral effects on natural enemies” and “although there are data on 48 natural enemy species, the database is still far from adequate to predict the effect of a Bt toxin or proteinase inhibitor on natural enemies”. Shelton et al. (2009) published a rebuttal of these interpretations stating that Lövei et al. (2009) conducted a data-mining exercise without prior elaboration of a risk hypothesis framework (Romeis et al. 2008), used inappropriate and unsound methods for risk assessment that have led them to reach conclusions that are in conflict with those of several recent comprehensive reviews and meta-analyses. The latter are summarized below.
In a systematic review, Romeis et al. (2006b) found that field studies generally confirmed that the abundance and activity of parasitoids and predators are similar in Bt and non-Bt crops. As far as MON810 is concerned, they compiled two field studies (Bourguet et al. 2002; Manachini 2003) which observed a lesser abundance of specialist enemies of the pest targeted by Bt maize. This is a consequence of the efficient control of target pest insects such as the European corn borer by Cry1Ab maize. Indeed, without their favorite prey or host, these specialists will not be attracted to the maize field.
Marvier et al. (2007) published a meta-analysis of 42 field experiments examining the effects of insect-protected cotton and maize. They noted that certain non-target taxa were less abundant in Bt fields compared to insecticide-free fields. For Cry1Ab plants, they emphasized the less abundant parasitic wasps of the braconidae and ichneumonidae.
In another meta-analysis, Wolfenbarger et al. (2008) also observed that insecticide effects were much larger than those of Bt crops. In maize, these analyses also revealed a reduction of parasitoids in Bt fields. Examination of the 116 observations showed that most of these reductions (n = 93) concern Macrocentrus grandii, a specialist parasitoid of the Bt target, the European corn borer. Higher numbers of the generalist predator, Coleomegilla maculata, were associated with Bt maize but numbers of other common predatory genera were similar in Bt and non-Bt maize. No significant effects of Bt crops on detritivores were found.
Naranjo (2009) agrees with the above analysis in concluding that “the minor negative effects of Bt crops demonstrated in the field pale in comparison with alternative pest suppression measures based on insecticides”. These systematic reviews and meta-analyses agree on the fact that the lower abundance of some insects mainly concerns specialized enemies of the target pest (an expected consequence of its control).
Concluding remarks
We demonstrate that the suspension of cultivation of maize MON810 in Germany in April 2009 is based on an incomplete list of references and ignores the widely accepted case-by-case approach. We did not find any justification for this suspension in the scientific literature regarding possible effects under natural field conditions on non-target animals, including ladybirds and daphnia. Regarding the latter organisms, the GSO invoked hazard effects on the basis of two inconclusive laboratory studies, which were also by nature insufficient to evaluate ecosystem-scale consequences. Thus, the German authorities’ risk management option is based on confusion between a potential hazard and a proven risk in the scientific procedure of risk assessment.
Laboratory studies are necessary to set up diagnostic tests (Brun-Barale et al. 2005) and to detect toxicological impacts. In the tiered approach (step-wise), if early tests in the laboratory yield uncertain results, further well-designed laboratory studies could ensure that results are relevant to in natura observations. Subsequently, if effects are seen in laboratory assays, in natura studies should be implemented. If no effect is seen under laboratory worst-case exposure conditions, then effects are unlikely to be detected in the field. The German government suspension does not fit logically in such a scientific approach, especially since its justifications are contradicted by its own publication, e.g. the recently published BEETLE report co-authored by the Federal Office of Consumer Protection and Food Safety of the German government. This report states that “an extensive body of research data has been assembled on non-target organism impacts of insect resistant maize. The majority of laboratory studies and all the field studies reviewed did not reveal any unexpected adverse or long-lasting effect. One important lesson is that even if negative effects were observed in the laboratory (e.g. under worst-case conditions) no similar quantitative or qualitative adverse were necessarily detected in the field” (http://ec.europa.eu/environment/biotechnology/pdf/beetle_report.pdf).
In addition, the GSO fails to take into account that many publications have shown that the differences are more significant between two non-Bt varieties than between isogenic Bt and non-Bt varieties at the farm scale. As Rauschen et al. (2009) showed recently, there are major differences between the two conventional maize varieties used in their field experiment, with regard to the densities of the mirid Trigonotylus caelestialium. One can also note that gene expression profiles of MON810 and comparable non-GM maize varieties cultivated in the field are more similar than are those of conventional lines (Coll et al. 2009). The GSO also fails to recognize an important conclusion of all recent meta-analyses (cited above), namely that (authorized) insecticide effects are much larger than those of Bt crops.
To encourage evidence-based risk analyses, we have constructed a systematic compilation of publications dealing with Cry proteins from B. thuringiensis or maize (see Supplementary electronic material). Our updated systematic review on impacts of MON810 and other Cry1Ab maize on non-target organisms complements recent reviews/meta-analyses (see above; Sanvido et al. 2007, which cites references published up to 2005; Icoz and Stotzky 2008, which analyzes impacts of soil organisms up to 2007).
The question of which scientific knowledge is mobilized for the implementation of public policies in agriculture is crucial. Ecology concentrates on the relationships between organisms and between organisms and their environments (see Burel et al. 2009). However, Raybould (2007) noted that, although transgenic crops have been the subject of much publicly funded research, this seems to have increased controversy rather than assisted decision-makers. Environmental risk assessment research has often attempted to describe the multitude of potential interactions between transgenic plants and the environment, rather than to test simple hypotheses selected by policy relevance. Raybould (2007) argued that a more effective method of research for decision-making follows principles exemplified by ecotoxicology. In either case, it matters how designers of measures mobilize sources of scientific knowledge, assess their quality and take their empirical validity into account when drawing up public policies (Laurent et al. 2008). It should be kept in mind that scientific knowledge and its use by policy makers change with time. This is attested by the emerging concepts of ‘evidence-based policy’ and ‘evidence-based decisions’—evidence being scientifically validated knowledge (Davies and Nutley 2001). Quantitative reviews of existing studies are necessary for better gauging risks and improving future risk assessments.
However, it is not enough: the present review demonstrates that the available meta-knowledge on Cry1Ab expressing maize was ignored by the German government who instead used selected individual studies which fit what seems to be a political decision. Similarly, the French government organized a stakeholder dialogue in 2007, which did not allow the emergence of the best scientific data available on GMOs (Kuntz 2008), but led the French government to suspend the cultivation of MON810 in 2008 on the basis of selected studies fitting the political agreement of this stakeholder dialogue. This strategy of political authorities does not take into account the findings of a recent unbiased stakeholder consultation in France (Ricroch and Jésus 2009) which established that, for all stakeholders, raising the objectivity of the debate on GMOs is the most important request.
References
Anderson PL, Hellmich RLII, Prasifka JR, Lewis LC (2005) Effects on fitness and behavior of monarch butterfly larvae exposed to a combination of Cry1ab-expressing corn anthers and pollen. Environ Entomol 34:944–952
Beachy RN, Fedoroff NV, Goldberg RB, McHughen A (2008) The burden of proof: a response to Rosi-Marshall et al. Proc Natl Acad Sci USA 105, 7:9
Bōhn T, Primicerio R, Hessen DO, Traavik T (2008) Reduced fitness of Daphnia magna fed a Bt-transgenic maize variety. Arch Environ Contam Toxicol 55:584–592
Bosch D, Schipper B, van der Kleij H, de Maagd RA, Stiekema WJ (1994) Recombinant Bacillus thuringiensis crystal proteins with new properties: possibilities for resistance management. Bio/Technology 12:915–918
Bourguet D, Chaufaux J, Micoud A, Delos M, Naïbo B, Bombarde F, Marque G, Eychenne N, Pagliari C (2002) Ostrinia nubilalis parasitism and the field abundance of non-target insects in transgenic Bacillus thuringiensis corn (Zea mays). Environ Biosaf Res 1:49–60
Bruck DJ, Lopez MD, Lewis LC, Prasifka JR, Gunnarson RD (2006) Effects of transgenic Bacillus thuringiensis corn and permethrin on nontarget arthropods. J Agr Urban Entomol 23:111–124
Brun-Barale A, Bouvier JC, Pauron D, Bergé JB, Sauphanor B (2005) Involvement of a sodium channel mutation in pyrethroid resistance in Cydia pomonella L., and development of a diagnostic test. Pest Manag Sci 61:549–554
Büchs W, Prescher S, Müller A (2004) Entwicklungsverzögerungen bei Zersetzern und ihren Räubern nach Aufnahme von MON 810 Bt-Maisstreu. Folgen für das Ökosystem. SiFo Project Status Seminar. Sicherheitsforschung und Monitoring. Berlin, June 16th 2004. http://www.gmo-safety.eu/en/maize/soil/308.docu.html
Burel F, Allsopp N, Ricroch A, Baudry A (2009) Research programmes in ecology. In: Burel F (ed) Plurality of science for interdisciplinarity and policy making. Theoretical findings from analyses of sustainable development in agriculture. Bentham E-book, Paris (in press)
Candolfi MP, Brown K, Grimm C, Reber B, Schmidli H (2004) A faunistic approach to assess potential side-effects of genetically modified Bt-corn on non-target arthropods under field conditions. Biocontrol Sci Tech 14:129–170
Coll A, Nadal A, Collado R, Capellades G, Messeguer J, Melé E, Palaudelmàs M, Pla M (2009) Gene expression profiles of MON810 and comparable non-GM maize varieties cultured in the field are more similar than are those of conventional lines. http://www.springerlink.com/content/f5l108080521538u/?p=b57038dca946410d90b10c84d2c790a9&pi=3
Davies HTO, Nutley SM (2001) Evidence-based policy and practice: moving from rhetoric to reality. In: Proceedings of the third international Interdisciplinary Evidence-based Policies and Indicator Systems Conference, pp 86–95
Delrio G, Verdinelli M, Serra G (2004) Monitoring of pest and beneficial insect populations in summer sown Bt maize. IOBC/WPRS Bull 27:43–48
Dively GP, Rose R, Sears MK, Hellmich RL, Stanley-Horn DE, Calvin DD, Russo JM, Anderson PL (2004) Effects on monarch butterfly larvae (Lepidoptera: Danaidae) after continuous exposure to Cry1Ab—expressing corn during anthesis. Environ Entomol 33:1116–1125
Douville M, Gagné F, Masson L, McKay J, Blais C (2005) Tracking the source of Bacillus thuringiensis Cry1Ab endotoxin in the environment. Biochem Syst Ecol 33:219–232
Douville M, Gagné F, Blaise C, André C (2007) Occurrence and persistence of Bacillus thuringiensis (Bt) and transgenic Bt corn cry1Ab gene from an aquatic environment. Ecotoxicol Environ Saf 66:195–203
Felke M, Langenbruch GA (2005) Auswirkungen des Pollens von transgenem Mais auf ausgewählte Schmetterlingslarven. BfN Skripten 157. http://www.bfn.de/fileadmin/MDB/documents/skript157.pdf
Górecka J, Godzina M, Dabrowski ZT (2008) Effect of Bt maize MON 810 expressing Cry 1 Ab toxin on Aphidius colemani in tritrophic plant–herbivore–parasitoid system. J Plant Prot Res 48:130–136
Griffiths BS, Caul S, Thompson J, Birch ANE, Cortet J, Andersen MN, Krogh PH (2007) Microbial and microfaunal community structure in cropping systems with genetically modified plants. Pedobiologia 51(3):195–206
Griffiths NA, Tank JL, Royer TV, Rosi-Marshall EJ, Whiles MR, Chambers CP, Frauendorf TC, Evans-White MA (2009) Rapid decomposition of maize detritus in agricultural headwater streams. Ecol Appl 19:133–142
Hellmich RL, Siegfried BD, Sears MK, Stanley-Horn DE, Daniels MJ, Mattila HR, Spencer T, Bidne KG, Lewis LC (2001) Monarch larvae sensitivity to Bacillus thuringiensis—purified proteins and pollen. Proc Natl Acad Sci USA 98:11925–11930
Hoheisel GA, Fleischer SJ (2007) Coccinellids, aphids, and pollen in diversified vegetable fields with transgenic and isoline varieties. J Insect Sci 7: Article 61. http://www.benthos.org/database/allnabstracts.cfm/db/Columbia2007abstracts/id/370
Hubert J, Kudlíková-Křížková I, Stejskal V (2008) Effect of MON 810 Bt transgenic maize diet on stored-product moths (Lepidoptera: Pyralidae). Crop Prot 27:489–496
Icoz I, Stotzky G (2008) Fate and effects of insect-resistant Bt crops in soil ecosystems. Soil Biol Biochem 40:559–586
Jesse LCH, Obrycki JJ (2000) Field deposition of Bt transgenic corn pollen: lethal effects on the monarch butterfly. Oecologia 125:241–248
Kuntz M (2008) Grenelle de l’environnement et plantes génétiquement modifiées. In: Les Cahiers Rationalistes n°593, March–April. http://www.union-rationaliste.org/index.php/Cahiers-Rationalistes/Les-Cahiers-Rationalistes-n-593/Fiche-detaillee.html
Lang A, Vojtech E (2006) The effects of pollen consumption of transgenic Bt maize on the common swallowtail, Papilio machaon L. (Lepidoptera, Papilionidae). Basic Appl Ecol 7:296–306
Laurent C, Baudry J, Berriet-Solliec M, Kirsch M, Perraud D, Tinel B, Trouvé A, Allsopp N, Bonnafous P, Burel F, Carneiro MJ, Giraud C, Labarthe P, Matose F, Ricroch A (2008) Around the notion of “Evidence based policy”. Communication to the colloquium. ‘The words of development: origins, usage and trajectories’. Paris, 13–14 Dec 2008
Losey JE, Rayor LS, Carter ME (1999) Transgenic pollen harms monarch larvae. Nature 399:214
Lövei GL, Andow DA, Arpaia S (2009) Transgenic insecticidal crops and natural enemies: a detailed review of laboratory studies. Environ Entomol 38(2):293–306
Manachini B (2003) Effects of transgenic corn on Lydella thompsoni Hertig (Diptera: Tachinidae) parasitoid of Ostrinia nubilalis Hb. (Lepidoptera: Crambidae). Boll Zool Agrar Bachic 35:111–125
Marvier M, McCreedy C, Regetz J, Kareiva P (2007) A meta-analysis of effects of Bt cotton and maize on non-target invertebrates. Science 316:1475–1477
Mattila HR, Sears MK, Duan JJ (2005) Response of Danaus plexippus to pollen of two new Bt corn events via laboratory bioassay. Entomol Exp Appl 116:31–41
Minorsky PV (2001) The hot and the classic. The monarch butterfly controversy. Plant Physiol 127:709–710
Moser SE, Harwood JD, Obrycki JJ (2008) Larval feeding on Bt hybrid and non-Bt corn seedlings by Harmonia axyridis (Coleoptera: Coccinellidae) and Coleomegilla maculata (Coleoptera: Coccinellidae). Environ Entomol 37:525–533
Musser FR, Shelton AM (2003) Bt-sweet corn and selective insecticides: impacts on pests and predators. J Econ Entomol 96:71–80
Naranjo SE (2009) Impacts of Bt crops on non-target invertebrates and insecticide use patterns. CAB Rev Perspect Agric Vet Sci Nutr Nat Resour 4:23
Orr DB, Landis DA (1997) Oviposition of European corn borer (Lepidoptera: Pyralidae) and impact of natural enemy populations in transgenic versus isogenic corn. J Econ Entomol 90:905–909
Parrott W (2008) Study of Bt impact on caddisflies overstates its conclusions: response to Rosi-Marshall et al. Proc Natl Acad Sci USA 105 7: E10
Pilcher CD, Rice ME, Obrycki JJ (2005) Impact of transgenic Bacillus thuringiensis corn and crop phenology on five nontarget arthropods. Environ Entomol 34:1302–1316
Pleasants JM, Hellmich RL, Dively GP, Sears MK, Stanley-Horn DE, Mattila HR, Foster JE, Clark TL, Jones GD (2001) Corn pollen deposition on milkweeds in and near cornfields. Proc Natl Acad Sci USA 98:11913–11918
Pokelsek JD, Rosi-Marshall EJ, Chambers CP, Griffiths NA, Evans-White MA, Tank JL, Whiles MR, Royer TV (2007) Effects of Bt corn pollen on caddisfly growth rates in Midwestern agricultural streams. Poster presented at the NABS Annual meetings, Columbia, South Carolina
Rauschen S, Schultheis E, Pagel-Wieder S, Schuphan I, Eber S (2009) Impact of Bt-corn MON88017 in comparison to three conventional lines on Trigonotylus caelestialium (Kirkaldy) (Heteroptera: Miridae) field densities. Transgenic Res 18:203–214
Raybould A (2007) Ecological versus ecotoxicological methods for assessing the environmental risks of transgenic crops. Plant Sci 173:589–602
Ricroch A, Jésus F (2009) Consultations of stakeholders on the roles of research in relation to genetically modified plants in France. Publ Underst Sci 18(1):91–102
Romeis J, Bartsch D, Bigler F, Candolfi MP, Gielkens MMC, Hartley SE, Hellmich RL, Huesing JE, Jepson PC, Layton R, Quemada H, Raybould A, Rose RI, Schiemann J, Sears MK, Shelton AM, Sweet J, Vaituzis Z, Wolt JD (2006a) Moving through the tiered and methodological framework for non-target arthropod risk assessment of transgenic insecticidal crops. In: Ninth international symposium on biosafety of genetically modified organisms biosafety research and environmental risk assessment. Jeju Island, Korea, pp 64–69. http://www.isbr.info/isbgmo/docs/9th_isbgmo_program.pdf
Romeis J, Meissle M, Bigler F (2006b) Transgenic crops expressing Bacillus thuringiensis toxins and biological control. Nat Biotechnol 24:63–67
Romeis J, Bartsch D, Bigler F, Candolfi MP, Gielkens MMC, Hartley SE, Hellmich RL, Huesing JE, Jepson PC, Layton R, Quemada H, Raybould A, Rose RI, Schiemann J, Sears MK, Shelton AM, Sweet J, Vaituzis Z, Wolt JD (2008) Assessment of risk of insect-resistant transgenic crops to non-target arthropods. Nat Biotechnol 26:203–208
Rosi-Marshall EJ, Tank JL, Royer TV, Whiles MR, Evans-White M, Chambers C, Griffiths NA, Pokelsek J, Stephen ML (2007) Toxins in transgenic crop byproducts may affect headwater stream ecosystems. Proc Natl Acad Sci USA 104:16204–16208
Rosi-Marshall EJ, Tank JL, Royer TV, Whiles MR (2008) Reply to Beachy et al. and Parrott: Study indicates Bt corn may affect caddisflies. Proc Natl Acad Sci USA 105:11
Sanvido O, Romeis J, Bigler F (2007) Ecological impacts of genetically modified crops: ten years of field research and commercial cultivation. Adv Biochem Eng 107:235–278
Schmidt JEU, Braun CU, L’Abate C, Whitehouse LP, Hilbeck A (2004) Studies on effects of Bacillus thuringiensis-toxins from transgenic insect-resistant plants on predaceous lady beetles (Coleoptera: Coccinellidae). Mitt Dtsch Ges Allg Angew Entomol 14(1–6):419–422
Schmidt JEU, Braun CU, Whitehouse LP, Hilbeck A (2009) Effects of Activated Bt Transgene Products (Cry1Ab, Cry3Bb) on immature stages of the ladybird Adalia bipunctata in laboratory ecotoxicity testing. Arch Environ Contam Toxicol 56:221–228
Sears MK, Hellmich RL, Stanley-Horn DE, Oberhauser KS, Pleasants JM, Mattila HR, Siegfried BD, Dively GP (2001) Impact of Bt corn pollen on monarch butterfly populations: a risk assessment. Proc Natl Acad Sci USA 98:11937–11942
Shelton AM, Naranjo SE, Romeis J, Hellmich RL, Wolt JD, Federici BA, Albajes R, Bigler F, Burgess EP, Dively GP, Gatehouse AM, Malone LA, Roush R, Sears M, Sehnal F (2009) Setting the record straight: a rebuttal to an erroneous analysis on transgenic insecticidal crops and natural enemies. Transgenic Res 18:317–322
Stanley-Horn DE, Dively GP, Hellmich RL, Mattila H, Sears MK, Rose R, Jesse LC, Losey JE, Obrycki JJ, Lewis L (2001) Assessing the impact of Cry1Ab-expressing corn pollen on monarch butterfly larvae in field studies. Proc Natl Acad Sci USA 98:11931–11936
Toschki A, Hothorn LA, Roß-Nickoll M (2007) Effects of cultivation of genetically modified Bt maize on epigeic arthropods (Araneae; Carabidae). Environ Entomol 36(4):967–981
Toth F, Arpas K, Szekeres D, Kadar F, Szentkiralyi F, Szenasi A, Kiss J (2004) Spider web survey or whole plant visual sampling. Impact assessment of Bt corn on non-target predatory insects with two concurrent methods. J Environ Biosaf Res 3:225–231
Tschenn J, Losey JE, Jesse LH, Obrycki JJ, Hufbauer R (2001) Effects of corn plants and corn pollen on monarch butterfly (Lepidoptera : Danaidae) oviposition behavior. Environ Entomol 30:495–500
US EPA (2000) Biopesticides Registration Document: preliminary risks and benefits sections; Bacillus thuringiensis plant pesticides.US Environmental Protection Agency and Office of Pesticide Progrems, Biopesticide and Pollution Prevention Division, Washington, DC
Wold SJ, Burkness EC, Hutchison WD, Venette RC (2001) In-field monitoring of beneficial insect populations in transgenic corn expressing a Bacillus thuringiensis toxin. J Ent Sci 36:2
Wolfenbarger LL, Naranjo SE, Lundgren JG, Bitzer RJ, Watrud LS (2008) Bt crop effects on functional guilds of non-target arthropods: a meta-analysis. PLoS ONE 3:218
Wraight CL, Zangerl AR, Carroll M, Berenbaum MR (2000) Absence of toxicity of Bacillus thuringiensis pollen to black swallowtails under field conditions. Proc Natl Acad Sci USA 97:7700–7703
Zangerl AR, McKenna D, Wraight CL, Carroll M, Ficarello P, Warner R, Berenbaum MR (2001) Effects of exposure to event 176 Bacillus thuringiensis corn pollen on monarch and black swallowtail caterpillars under field conditions. Proc Natl Acad Sci USA 98:11908–11912
Zenner de Polania I, Alvarez Alcaraz G (2008) Analysis of the influence on the main beneficial fauna by two transgenic varieties, cotton and corn, at El Espinal (Tolima). Rev UDCA Actual Divulg Cient 11:133–142
Acknowledgments
We thank the three anonymous reviewers for their valuable comments. Part of this work was carried out with the financial support of the “ANR—Agence Nationale de la Recherche—The French National Research Agency” under the “Programme Agriculture et Développement Durable”, project “ANR-06-PADD-EBP-BIOSOC”.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License ( https://creativecommons.org/licenses/by-nc/2.0 ), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Ricroch, A., Bergé, J.B. & Kuntz, M. Is the German suspension of MON810 maize cultivation scientifically justified?. Transgenic Res 19, 1–12 (2010). https://doi.org/10.1007/s11248-009-9297-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11248-009-9297-5