
Abstract
The temperature dependence of the rate constants for the formation of oligocytidylate (oligo(C)) from the 5′-monophosphorimidazolide of cytidine (ImpC) in the presence of Pb(II) ion catalyst has been investigated at 10–75°C. The rate constants for the formation of oligo(C) increased in the order of the formation of 2-mer < 3-mer ≤ 4-mer; this trend resembles the trend in the cases of the template-directed and the clay-catalyzed formations of oligonucleotides. While the rate constants of the formation of oligo(C) increased with increasing temperature, the yield of oligo(C) decreased with increasing temperature. This is due to the fact that the relative magnitude of the rate constants of the formation of 2-mer, 3-mer, and 4-mer to that of the hydrolysis of ImpC decreased with increasing temperature. This is probably due to the fact that association between ImpC with the elongating oligo(C) decreases with increasing temperature. The apparent activation energy was 61.9 ± 8.5 kJ mol−1 for the formation of 2-mer, 49.3 ± 2.9 kJ mol−1 for 3-mer, 51.8 kJ mol−1 for 4-mer, and 66.8 ± 4.5 kJ mol−1 for the hydrolysis of ImpC. The significance of the temperature dependence of the formation rate constants of the model prebiotic formation of RNA is discussed.
Similar content being viewed by others

Introduction
The RNA world hypothesis is generally accepted (Gilbert 1986; Cech 1986; Ellington and Szostak 1990; Gesteland and Atkins 1993). Experimental attempts have been made to study the non-enzymatic oligomerization of ribonucleotides to model the possible origins of an RNA world (Lohrmann and Orgel 1980; Inoue and Orgel 1982, 1983; Sawai 1976; Sawai et al. 1981, 1989, 1992; Sawai and Ohno 1985; Ertem and Ferris 1996; Ferris and Ertem 1992a,b; Ferris et al. 1996). However, the hypothesis that life originated under hydrothermal vent environments (the hydrothermal origin of life hypothesis) (Holm 1992; Holm and Andersson 1995; Imai et al. 1999) appears to be inconsistent with the RNA world hypothesis. The hydrothermal origin of life hypothesis was proposed based on the existence of thermophilic organisms (Corliss et al. 1981; Baross and Hoffman 1985; Nisbet 1986) and phylogenetic analysis of present organisms. The last common ancestor (LCA) is considered by many to have been a thermophilic organism (Pace 1991; Oshima 1994; Forterre 1995); however, this is widely disputed (Miller and Bada 1988; Galtier et al. 1999; Miyakawa et al. 2002a,b). In addition, it is difficult to determine the temperature at which life originated. The temperature of the primitive ocean in which life originated remains speculative (Walker 1985; Karhu and Epstein 1986; Kasting and Ackerman 1986).
A few investigations concerning the degradation of RNA and its precursors have been conducted from the standpoint of the hydrothermal origin of life (White 1984; Miller and Lazcano 1995; Larralde et al. 1995; Kawamura et al. 1997; Levy and Miller 1998; Kawamura 1998, 2000, 2003a,b). According to these empirical data, it is considered that RNA molecules are too labile under hydrothermal vent conditions for these two hypotheses to be compatible (Kawamura 2004, 2005). Furthermore, biologically important weak interactions such as hydrophobic interactions and hydrogen bonding are weaker at higher temperatures (Kawamura 2005). However, it should be noted that the accumulation behavior of RNA in an open system is kinetically controlled by both the formation and decomposition rates of RNA (Kawamura 2000, 2003a,b). If the formation rate of RNA is greater than the degradation rate, the accumulation of RNA molecules would be kinetically possible even at high temperatures.
There have been a number of successful studies of the condensation of activated nucleotides to form RNA oligonucleotides in the presence of RNA templates (TD reaction) (Inoue and Orgel 1982, 1983; Joyce et al. 1984), and in the presence of metal ion (Sawai 1976; Sawai et al. 1981, 1989, 1992; Sawai and Ohno 1985) or clay mineral catalyst (CL reaction) (Ferris and Ertem 1992a,b; Ertem and Ferris 1996). In addition, kinetic analysis of these model reactions has been important to evaluate the reaction mechanism (Kanavarioti et al. 1993; Kanavarioti 1994, 1998; Kawamura and Ferris 1994, 1999). However, these simulation reactions of the prebiotic formation of RNA were mainly studied at low temperatures (∼37°C) (Inoue and Orgel 1982, 1983; Fakharai et al. 1984; Kanavarioti et al. 1993; Kanavarioti 1994, 1998). Thus, in order to evaluate the possibility of the prebiotic formation of RNA under hydrothermal conditions, we have investigated the temperature dependence of the TD reaction of oligoguanylate (oligo(G)) formation on a polycytidylic acid (poly(C)) template and the cyclization reaction of hexanucleotides in the presence of a condensing agent at 40–80°C (Kawamura and Umehara 2001; Kawamura et al. 2003). These investigations have suggested that oligonucleotides are unstable in aqueous solution at high temperatures, but also the prebiotic formation rates of oligonucleotides in these simulation reactions can be greater than the degradation rate of RNA at up to near 300°C (Kawamura 2005). Naturally, the estimation does not straightforwardly elucidate that the accumulation of RNA molecules could have proceeded under the primitive hydrothermal conditions. However, to evaluate the meaning of the rate of prebiotic formation of RNA at high temperatures and to search plausible conditions for the accumulation of RNA molecules under hydrothermal environments, it is important to accumulate kinetic data regarding the temperature dependence of the prebiotic simulation reactions of RNA molecules. In the present study, the kinetic analysis of the formation of oligo(C) in the presence of Pb(II) ion catalysts has been carried out at 10–75°C (Sawai et al. 1981; Sawai and Ohno 1985).
Experimental
Materials and Equipment
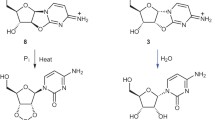
The activated nucleotide monomer was prepared by the techniques in previous studies (Joyce et al. 1984; Ferris and Ertem 1992b). All other reagents used were of analytical grade. High-performance liquid chromatography (HPLC) was performed by a LC10A HPLC system (Shimadzu, Japan) with a DNA-NPR anion-exchange column from TOSOH Co., Japan using a gradient of 0.3–1.5 M NaCl at pH 9 with 0.02 M 2-amino-2-hydroxymethyl-1,3-propanediol buffer and a ODS-2 column from GL Science Co., Japan using a gradient of 0.005 M NaH2PO4 in water at pH 3.5 mixed with 0.01 M NaH2PO4 in 40% CH3OH at pH 4.0.
Kinetics
Oligomerization of ImpC in the Presence of Pb(II) Ion at Elevated Temperatures
The formation of oligo(C) from ImpC in the presence of Pb(II) was performed in a solution containing 0.015 M ImpC, 0.001 M Pb(NO3)2, 0.001 M 2-[4-(2-hydroxyethyl)-1-piperazinyl]ethanesulfonic acid (HEPES), at pH 8.0. Six mg ImpC and 0.238 mg HEPES were placed in a 1.5 ml plastic vial (RNase free), which bears up to 120°C, and to it was added a 0.067 ml of 0.015 M Pb(NO3)2 solution and 0.933 ml distilled water. The pH of the mixture was adjusted to 8.0 using a small amount of NaOH. The vial was settled in a cartridge temperature controller at 10–75°C. The sample solution was withdrawn at regular intervals over 200 h at 10°C, 96 h at 25°C, 8 h at 50°C, and 1.5 h at 75°C, and then immediately quenched using liquid nitrogen. Control reactions without Pb(NO3)2 were also carried out. The samples were analyzed by both the anion-exchange HPLC (AE-HPLC) and the reversed-phase HPLC (RP-HPLC). The calculation of the rate constants was performed using a kinetic program SIMFIT (Terfort and von Kiedrowski 1992).
Degradation of Oligo(C)
A standard solution containing oligo(C) was prepared from the Pb(II)-catalyzed oligo(C) formation at 25°C for 7 days. Running solutions containing the standard oligo(C) were incubated at 60–90°C and aliquot samples were withdrawn at regular intervals. The samples were analyzed by AE-HPLC. The rate constants of the degradation of oligo(C) were determined using the previous method (Kawamura et al. 1999; Kawamura and Umehara 2001).
Results and Discussion
Formation of Oligo(C) in the Presence of Lead(II) Ion
During the reaction of the formation of oligo(C) in the presence of Pb(II), white precipitates of Pb(OH)2 were sometimes observed at different Pb(II) and salt concentration conditions. The presence of NaCl and/or MgCl2 and a high concentration of Pb(II) seem to enhance the formation of precipitates, where NaCl and MgCl2 probably act as a reagent to reduce the solubility of Pb(OH)2 with increasing the ionic strength or result in the precipitate formation of PbCl2. Since the precipitate would result in inaccurate kinetic analysis, the reaction conditions described in the experimental section were selected to avoid the formation of precipitates. Typical chromatograms are shown in Fig. 1. The formation of oligo(C) was monitored using AE-HPLC, and the disappearance of ImpC by the hydrolysis of ImpC to form 5′pC, the oligomerization to form oligo(C), and the formation of P 1,P 2-dicytidine 5′,5′-pyrophosphate (C5′ppC) were monitored using RP-HPLC (Kawamura and Umehara 2001). The extents of ImpC, 5′pC, oligo(C), and C5′ppC were summarized in Table 1 and the reaction curves are shown in Fig. 2. No oligo(C) was detected in the control reactions without the Pb(II) ion catalyst. The reaction times at different temperatures, where the amount of remaining ImpC is roughly the same (Table 1), were chosen to visualize the influence of temperature. Although the yield of oligo(C) decreases with increasing temperature, the extent of oligo(C) was a maximum at 25°C, where 5-mer was observed. The fact that the yield of oligo(C) at 10°C is smaller than that 25°C may be due to that a trace precipitate of Pb(OH)2 forms at 10°C. The trend that the yield of oligo(C) decreases with the length of oligo(C) is the same as shown in that in the cases of the TD reaction and the CL reaction (Kanavarioti et al. 1993; Kawamura and Ferris 1994, 1999).

HPLC charts for the oligo(C) formation from ImpC in the presence of Pb(II) catalyst. Reaction conditions: [ImpC] = 0.015 M, [Pb(NO3]2] = 0.001 M, [HEPES] = 0.001 M, pH = 8.0. A: 25°C, 96 h, B: 75°C, 1.5 h, C: 25°C, 96 h, no Pb(NO3)2 was added

Reaction curves for the oligo(C) formation from ImpC in the presence of Pb(II) catalyst. Reaction conditions are the same as shown in Fig. 1. a: 10°C, b: 25°C, c: 50°C, d: 75°C. The lines drawn through the experimental points were fit by SIMFIT. Closed circles: 5′pC, open squares: C5′ppC, closed squares: 2-mer, open triangles: 3-mer, closed triangles: 4-mer, small open circles: 5-mer
Kinetics
The reaction rate constants for the processes in the present reaction system were determined using the following model (Kawamura and Ferris 1994),
where (pC)2, (pC)3, (pC)n−1, (pC)n indicate 2-mer, 3-mer, (n-1)-mer, and n-mer oligo(C) and k 2, k 3, and k n are the rate constants for the formation of 2-mer, 3-mer, and n-mer, where k n was defined as a constant value for 4-mer and longer oligo(C) (k n = k 4). Here, the approximation that the rate constants of the formation of (pC)2 are equated in the pathways Eqs. 2 and 3 was applied in the previous studies since the values are not distinguishable by the present HPLC technique. It was evaluated that this approximation is reasonable. Thus, the same approximation has been adopted in the present study. The rate constant of the ImpC hydrolysis is k hy,ImpC and that for the formation of C5′ppC is k py. The reaction curves shown in Fig. 2 were fitted using a kinetic program SIMFIT (Terfort and von Kiedrowski 1992). The rate constants were summarized in Table 2.
The rate constant of the ImpC hydrolysis (k hy,ImpC) in the absence of Pb(II) was about two times greater than that in the presence of Pb(II), while no oligo(C) was detected in the absence of Pb(II) (Table 2). This fact suggests that Pb(II) ion possesses a weak inhibitory activity for the hydrolysis of ImpC.
Besides, the rate constants of oligo(C) formation up to k 4 were able to be determined at 10 and 25°C and k 2 and k 3 were determined at 50 and 75°C. The magnitude of oligo(C) formation increased in the order k 2 < k 3(≤k 4) (Fig. 3). This trend can be observed at 50–75°C although no data was obtained for k 4. This trend coincides with the cases of the TD reaction and the CL reaction (Kanavarioti et al. 1993; Kawamura and Ferris 1994), while the rate constants in the present reaction were somewhat smaller than those for the TD reaction and the CL reaction (Fig. 3). In addition, the difference between the magnitude of k 2 and that of k 4 in the present system is much smaller than that of the TD reaction and somewhat smaller than that of the CL reaction. This is probably due to that the importance of association between elongating oligonucleotide with the activated nucleotide monomer in the Pb(II)-catalyzed oligo(C) formation is weaker than that in the TD reaction and the CL reaction. The association would be likely enhanced in the presence of Pb(II) since the oligo(C) formation does not proceed at all in the absence of Pb(II) (Sawai 1976). On the other hand, the magnitude of the rate of the hydrolysis of ImpC to the rate of the formation of oligo(C) (Table 1) is relatively large as comparing to the TD and CL reactions. This is a reason that the yield of oligo(C) is smaller than that of oligonucleotides in the TD and CL reactions. Since the concentration of Pb(II) was decreased in the present study to avoid precipitation of Pb(OH)2, the efficiency of the catalytic formation of oligo(C) with Pb(II) was reduced. Thus, this fact would reflect that the relatively weak efficiency of the present reaction is due to the relatively large rate constant of the ImpC hydrolysis. Conclusively, it is reasonable that the associate formation between the activated monomer and elongating oligonucleotide would be as important as that in the TD reaction and in the CL reaction. A possible model is shown in Fig 4.

Relationships between logarithmic values of k n and length n for the Pb(II) catalyzed oligo(C) formation, TD reaction, and CL reaction. Lines: Open circles (25°C), closed squares (10°C), open triangles (50°C), closed triangles (75°C): Pb(II) catalyzed oligo(C) formation, closed circles: TD reaction, 23°C (Kanavarioti et al. 1993), open squares: CL reaction, 25°C (Kawamura and Ferris 1994)

Possible associate formation for the Pb(II) catalyzed formation of oligo(C)
On the other hand, the degradation of oligo(C) formed in the presence of Pb(II) ion catalysts was also investigated at 60–90°C. Since oligo(C) molecules formed by the Pb(II) ion catalyzed reaction involve 2′,5′-linked oligo(C) isomers (Sawai et al. 1981; Sawai and Ohno 1985), the oligo(C) prepared by the Pb(II)-catalyzed reaction was used for the degradation study of oligo(C) instead of an authentic 3′,5′-linked oligo(C). The degradation rates were examined on the basis of the disappearance of phosphodiester bond. The mole concentration of the total number of phosphodiester bond (C bond) is calculated from the concentration of oligo(C) (C oligo) using Eq. 7 (Kawamura et al. 1999; Kawamura and Umehara 2001),
where Ai indicates the ratio of the oligo(G) in length i. Here, it was assumed that the rate constant of the cleavage of phosphodiester bond is independent on the values of i. The rate constants of the hydrolytic degradation of oligo(C) (k hy,oligo) are summarized in Table 3. The units of k hy,oligo (s −1) and k hy,ImpC (s −1) are the same, and these are different from the unit for k 2–k 4 (M −1 s −1). Thus, the direct comparison between k hy,oligo and k 2–k 4 is not meaningful. However, the magnitude of k hy,oligo is much smaller than that of k hy,ImpC, and the rate of the formation of oligo(C) is comparable to that of the hydrolysis of k hy,ImpC at 10–75°C, where oligo(C) indeed forms by the Pb(II)-catalyzed reaction. Thus, it is true that the rate of oligo(C) degradation is much smaller than that of the formation of oligo(C) from ImpC by the Pb(II) catalyzed reaction.
Temperature Dependence of the Rate Constants and the Chemical Evolution of RNA Under the Hyrothermal Conditions
The logarithmic values of the rate constants determined by the present study were plotted as a function of T−1 (Fig. 5). Based on the Arrhenius plots shown in Fig. 5 and Eyring plots, the apparent activated energy (Ea), enthalpy change (\( \Delta H^{\ddag } \)), and entropy changes (\( \Delta S^{\ddag } \)) were calculated. The activation parameters are summarized in Table 4. The magnitudes of Ea and \( \Delta H^{\ddag } \) in the present system are somewhat smaller than those in the TD reaction (Kawamura and Umehara 2001; Kawamura et al. 2003). The magnitude of Ea for k hy,ImpC, k py, and k 2 are comparable each other’s, but the magnitude of Ea for k 3 is smaller than that for k hy,ImpC, k py, and k 2. This is different from the trend that the magnitude of Ea for k 2 is much smaller than that for the hydrolysis of the activated nucleotide monomer, and k 3 and k 4 in the TD reaction. In addition, the magnitudes of Ea for k 2, k 3, and k 4 are comparable to the case of the TD reaction, where the magnitude of Ea for k 2 is clearly smaller than those for k 3 and k 4. Although the rate constants of oligo(C) formation increased with increasing temperature, the difference between k 2 and k 4 (or k 3) becomes small at high temperatures. This is a reason that the efficiency of oligo(C) formation decreases with temperature. This is consistent with the mathematical simulation that the yield of longer oligo(C) decreases when the ratio of the formation rate constant of longer oligo(C) to that of the formation of 2-mer decreases (Kawamura and Umehara 2001).

Arrhenius plots for the Pb(II) catalyzed formation of oligo(C), the hydrolysis of ImpC and the hydrolysis of oligo(C). Lines: open circles: k hy,ImpC with Pb(II) catalyst, closed circles: k hy,ImpC without Pb(II) catalyst, open squares: k 2, closed squares: k 3, open triangles: k 4, open rhombus k py, closed rhombus: k hy,oligo
On the other hand, although the rate constant of the hydrolysis of ImpC in the presence of Pb(II) is smaller than that in the absence of Pb(II) at 10–25°C, the difference of the rate constants disappeared at 50–75°C. It was unexpected that Pb(II) ion inhibits the hydrolysis of ImpC at low temperatures. This is probably due to that Pb(II) is capable to form complex with ImpC, in which the hydrolytic stability of ImpC within Pb(II)-ImpC complex is greater than that of free ImpC. In addition, this fact suggests that the coordination with phosphate group and imidazole group to Pb(II) in the Pb(II)-ImpC is important as well as that with the pyrimidine base. As mentioned above, the relationship between logk n vs. length n indicates that the association between the activated monomer and elongating oligo(C) is important. However, for the case of ImpC the association would be not be so strong comparing to the cases of purine nucleotide monomers. Thus, the coordination of Pb(II) to the side of phosphate and imidazole group may be rather important to facilitate the formation of the associate between ImpC and elongating oligo(C) for the present system. It is reasonable that the stability of Pb(II)-ImpC complex becomes weak with increasing temperature so that the difference between the hydrolysis rates in the presence and absence of Pb(II) disappears with increasing temperature (Fig. 5).
The present kinetic analysis elucidated that the degradation of oligo(C) is much slower than that of the formation of oligo(C) by the Pb(II)-catalyzed formation from ImpC at high temperatures. This trend is the same as the case of the temperature dependence of the rate constants in the TD reaction (Kawamura and Umehara 2001). In the present system, the direct comparison of the rate constants for the formation and degradation of oligo(C) is not meaningful since the units of the rate constants are different. Thus, the half-lives (τ), which reflect the rate of the reactions, were compared. The values of log(τ) were plotted as a function of T −1 (Fig. 6). The values of τ for the hydrolysis of ImpC are much smaller than those for the formation of oligo(C) in the present system. This reflects the relatively low efficiency of the present reaction in contrast to the TD reaction. Here, it is noted that the half-lives of pseudo-second-order rate processes (k 2, k 3, k 4) are dependent on the concentration of the reactants, while the calculation was performed at 0.015 M ImpC. Thus, the magnitude of τ for the formation of oligo(C) decreases with increasing the ImpC concentration. The magnitude of τ of the formation of 2-mer is 23 times greater than that of the ImpC hydrolysis at 75°C. Thus, the magnitude of τ for the formation of 2-mer becomes comparable to that of the ImpC hydrolysis at 0.345 M ImpC at 75°C. This fact indicates that the oligonucleotide would have been formed at higher temperatures if a high concentration of the activated nucleotide monomers could have been supplied along with Pb(II) ion catalyst. While the concentration of Pb(II) ion in oceanic water is quite low, that in submarine hydrothermal fluid could have been higher than the bulk ocean water as shown in modern hydrothermal vent systems (Metz and Trefry 2000); although the concentration of Pb(II) found in these hydrothermal vent systems is lower than that used in the present simulation. It has been shown that the activated nucleotide monomer can be formed from inorganic phosphate, nucleotide monomer and imidazole (Lohrmann and Orgel 1973; Orgel and Lohrmann 1974). Further investigation has suggested that the activated monomer could have formed in fairly high concentrations at 37–100°C (Lohrmann 1977). Thus, it would be important to search for possible pathways where activated monomers could have formed under hydrothermal conditions.

Relationships between logarithmic values of τ and T−1 for the Pb(II) catalyzed formation of oligo(C), the hydrolysis of ImpC and the hydrolysis of oligo(C). Lines: open circles: k hy,ImpC with Pb(II) catalyst, closed circles: k hy,ImpC without Pb(II) catalyst, open squares: k 2, closed squares: k 3, open triangles: k 4, closed rhombus: k hy,oligo
In addition, the value of τ for the formation of oligo(C) is much smaller than that for the degradation of oligo(C) at higher concentrations of the activated monomers. This fact is consistent with the cases of the TD reaction and the CL reaction; the formation rates of oligonucleotide in these simulation reactions are much greater than the degradation rates of oligonucleotides formed by these reactions. Thus, it is generally true that the reaction rates of oligonucleotide formation in the prebiotic model reactions examined in our group are greater than the degradation rate of oligonucleotide. In the TD reaction, the relatively slow rate of the formation of 2-mer was a main reason of the low yield of oligonucleotide at high temperatures (Kawamura and Umehara 2001). Besides, the increase of the ImpC hydrolysis with increasing temperature is a main reason of the low yield of oligo(C) at high temperatures in the present reaction system. These facts suggest that the formation of oligonucleotides would be possible if the relative reaction rate of 2-mer formation is enhanced in the TD reaction and the hydrolysis of the activated nucleotide monomer is inhibited in the present system by some additives, such as protein-like molecules and minerals (Kawamura 2005; Kawamura et al. 2005). We are currently searching such materials as well as continuing kinetic analyses of temperature dependence of other prebiotic models of the oligonucleotide formation.
The yield and length of oligonucleotides from the activated nucleotide monomer in the presence of Pb(II) catalyst are dependent on temperature, since the relative magnitude of the rate constants of the processes in the present system are dependent on temperature. Naturally, temperature varies widely in natural environments, such as in hydrothermal vent systems and surrounding deep oceans. Thus, it would be important to take this into account in the comparison of the rate constants or that of the values of τ. However, the trend that the values of τ increased in the order of the hydrolysis of ImpC < the formation of oligo(C) < the degradation of oligo(C) does not change at 0–300°C. In addition, the trend that the rate of phosphodiester bond formation is greater than that of phosphodiester bond degradation remains roughly unchanged in the TD reaction at up to 300°C. Therefore, the aspect derived from the present study on the prebiotic formation of oligonucleotide under hydrothermal environments is important for further evaluations of the hypothesis of the hydrothermal origin of life as well as the RNA world hypothesis.
Conclusions
Oligo(C) formation from ImpC was investigated at 10–75°C and a small amount of oligo(C) was detected even at 75°C. The rate constants for the hydrolysis of ImpC with and without Pb(II) and the formation of oligo(C) were determined at 10–75°C. In addition, the rate constants of the hydrolytic degradation of oligo(C) were determined at 60–90°C. Based on the temperature dependence of these rate constants, the activation parameters were determined. The main reason that the yield of oligo(C) decreases with temperature is due to the fact that the ratio of the rate of the hydrolysis of ImpC to that of the formation of oligo(C) increases with temperature, but not because of the degradation of oligo(C). This fact implies that oligonucleotide formation would have been possible at high temperatures if some accelerators such as protein-like molecules and/or minerals along with Pb(II) catalyst could have facilitated the formation of oligo(C) rather than the hydrolysis of ImpC.
References
Baross JA, Hoffman SE (1985) Submarine hydrothermal vents and associated gradient environments as sites for the origin and evolution of life. Orig Life 15:327–345
Cech TR (1986) A model for the RNA-catalyzed replication of RNA. Proc Natl Acad Sci 83:4360–4363
Corliss JB, Baross JA, Hoffman SE (1981) An hypothesis concerning the relationship between submarine hot springs and the origin of life on Earth. Ocean. Acta 4(suppl.):59–69
Ellington AD, Szostak JW (1990) In vitro selection of RNA molecules that bind specific ligands. Nature 346:818–822
Ertem G, Ferris JP (1996) Synthesis of RNA oligomers on heterogeneous templates. Nature 379:238–240
Fakharai H, Inoue T, Orgel LE (1984) Temperature-dependence of the template-directed synthesis of oligoguanylates. Tetrahedron 40:39–45
Ferris JP, Ertem G (1992a) Oligomerization of ribonucleotides on montmorillonite: reaction of the 5′-phosphorimidazolide of adenosine. Science 257:1387–1389
Ferris JP, Ertem G (1992b) Oligomerization reactions of ribonucleotides: the reaction of the 5′-phophirimidazolide of nucleotides on montmorillonite and other minerals. Orig Life Evol Biosph 22:369–381
Ferris JP, Hill JAR, Liu R, Orgel LE (1996) Synthesis of long prebiotic oligomers on mineral surfaces. Nature 381:59–61
Forterre P (1995) Thermoreduction, hypothesis for the origin of prokaryotes, CR sciences de la vie/life sciences. Acad Sci Paris 318:415–422
Galtier N, Tourasse N, Gouy M (1999) A nonhyperthermophilic common ancestor to extant life forms. Science 283:220–221
Gesteland RF, Atkins JF (eds) (1993) The RNA world. Cold Spring Harbor Laboratory, New York
Gilbert W (1986) The RNA world. Nature 319:618
Holm NG (ed) (1992) Special issue-marine hydrothermal systems and the origin of life. Orig Life Evol Biosph 22:5–242(and therein)
Holm NG, Andersson EM (1995) Abiotic synthesis of organic compounds under the conditions of submarine hydrothermal systems: a perspective. Planet Space Sci 43:153–159
Imai E, Honda H, Hatori K, Brack A, Matsuno K (1999) Elongation of oligopeptides in a simulated submarine hydrothermal system. Science 283:831–833
Inoue T, Orgel LE (1982) Oligomerization of (guanosine 5′-phosphor)-2-methylimidazolide on Poly(C), An RNA polymerase model. J Mol Biol 162:201–217
Inoue T, Orgel LE (1983) A nonenzymatic RNA polymerase model. Science 219:859–862
Joyce GF, Inoue T, Orgel LE (1984) Non-enzymatic template-directed synthesis on RNA random copolymers, Poly(C,U) templates. J Mol Evol 176:278–306
Kanavarioti A (1994) Template-directed chemistry and the origins of the RNA world. Orig Life Evol Biosph 24:479–494
Kanavarioti A (1998) Kinetic preference for the 3′–5′-linked dimer in the reaction of guanosine 5′-phosphorylmorpholinamide with deoxyguanosine 5′-phosphoryl-2-methylmidazolide as a function of Poly(C). J Org Chem 63:6830–6838
Kanavarioti A, Bernasconi CF, Alberas DJ, Baird EE (1993) Kinetic dissection of individual steps in Poly(C)-directed oligoguanylate synthesis from guanosine 5′-Monophosphate 2-Methylimidazolide. J Am Chem Soc 115:8537–8546
Karhu J, Epstein S (1986) The implication of the oxygen isotope records in coexisting cherts and phosphates. Geochim Cosmochim Acta 50:1745–1756
Kasting JF, Ackerman TP (1986) Climatic consequences of very high-carbon dioxide levels in the Earth’s early atmosphere. Science 234:1383–1385
Kawamura K (1998) Kinetic analysis of hydrothermal reactions by flow tube reactor -hydrolysis of adenosine 5′-triphosphate at 398-573 K–. Nippon Kagaku Kaishi 1998:255–262
Kawamura K (2000) Monitoring hydrothermal reactions on the millisecond time scale using a micro-tube flow reactor and kinetics of ATP hydrolysis for the RNA world hypothesis. Bull Chem Soc Jpn 73:1805–1811
Kawamura K (2003a) Kinetics and activation parameter analyses of hydrolysis and interconversion of 2′,5′- and 3′,5′-linked dinucleoside monophosphate at extremely high temperatures. Biochim Biophys Acta 1620:199–210
Kawamura K (2003b) Kinetic analysis of cleavage of ribose phosphodiester bond within guanine and cytosine rich oligonucleotides and dinucleotides at 65–200°C and its implications on the chemical evolution of RNA. Bull Chem Soc Jpn 76:153–162
Kawamura K (2004) Behavior of RNA under hydrothermal conditions and the origins of life. Int J Astrobiol 3:301–309
Kawamura K (2005) A new probe for the indirect measurement of the conformation and interaction of biopolymers at extremely high temperatures using a capillary flow hydrothermal reactor system for UV-visible spectrophotometry. Anal Chim Acta 543:236–241
Kawamura K, Ferris JP (1994) Kinetics and mechanistic analysis of dinucleotide and oligonucleotide formation from the 5′-Phosphorimidazolide of adenosine on Na+-montmorillonite. J Am Chem Soc 116:7564–7572
Kawamura K, Ferris JP (1999) Clay catalysis of oligonucletide formation: Kinetics of the reaction of the 5′-phosphorimidazolide of nucleotides with the non-basic heterocycles uracil and hypoxanthine. Orig Life Evol Biosph 29:563–591
Kawamura K, Umehara M (2001) Kinetic analysis of the temperature dependence of the template-directed formation of oligoguanylate from the 5′-phosphorimidazolide of guanosine on a Poly(C) template with Zn2+. Bull Chem Soc Jpn 74:927–935
Kawamura K, Yosida A, Matumoto O (1997) Kinetic investigations for the hydrolysis of adenosine 5′-triphosphate at elevated temperatures: prospects for the chemical evolution of RNA. Viva Orig 25:177–190
Kawamura K, Kameyama N, Matumoto O (1999) Kinetics of hydrolysis of ribonucleotide polymers in aqueous solution at elevated temperatures: implications of chemical evolution of RNA and primitive ribonuclease. Viva Orig 27:107–118
Kawamura K, Nakahara N, Okamoto F, Okuda N (2003) Temperature dependence of the cyclization of guanine and cytosine mix hexanucleotides with water-soluble carbodiimide at 0–75°C. Viva Orig 31:221–232
Kawamura K, Nagahama M, Kuranoue K (2005) Chemical evolution of RNA under hydrothermal conditions and the role of thermal copolymers of amino acids for the prebiotic degradation and formation of RNA. Adv Space Res 35:1626–1633
Larralde R, Robertson MP, Miller SL (1995) Rates of decomposition of ribose and other sugars: implications for chemical evolution. Proc Natl Acad Sci USA 92:8158–8160
Levy M, Miller SL (1998) The stability of the RNA bases: Implications for the origin of life. Proc Natl Acad Sci USA 95:7933–7938
Lohrmann R (1977) Formation of nucleoside 5′-Phosphorimidates under potentially prebiological conditions. J Mol Evol 10:137–154
Lohrmann R, Orgel LE (1973) Prebiotic activation process. Nature 244:418–420
Lohrmann R, Orgel LE (1980) Efficient catalysis of polycytidylic acid-directed oligoguanylate formation by Pb2+. J Mol Biol 142:555–567
Metz S, Trefry JH (2000) Chemical and mineralogical influences on concentrations of trace metals in hydrothermal fluids. Geochim Cosmochim Acta 64:2267–2279
Miller SL, Bada JL (1988) Submarine hot springs and the origin of life. Nature 334:609–611
Miller SL, Lazcano A (1995) The origin of life-did it occur at high temperatures? J Mol Evol 41:689–692
Miyakawa S, Cleaves HJ, Miller SL (2002a) The cold origin of life: A. Implications based on the hydrolytic stabilities of hydrogen cyanide and formamide. Orig Life Evol Bios 32:195–208
Miyakawa S, Cleaves HJ, Miller SL (2002b) The cold origin of life: B. Implications based on pyrimidines and purines produced from ammonium cyanide solutions. Orig Life Evol Bios 32:209–218
Nisbet EG (1986) RNA and hot-water springs. Nature 322:206
Orgel LE, Lohrmann R (1974) Prebiotic chemistry and nucleic acid replication. Acc Chem Res 7:368–377
Oshima T (1994) Origins and early evolution of life. Trans Mater Res Soc Jpn 19B:1069–1077
Pace NR (1991) Origin of life-facing up to the physical setting. Cell 65:531–533
Sawai H (1976) Catalysis of internucleotide bond formation by divalent metal ions. J Am Chem Soc 98:7037–7039
Sawai H, Ohno M (1985) Preparation and properties of oligocytidylates with 2′–5′ internucleotide linkage. Bull Chem Soc Jpn 58:361–366
Sawai H, Shibata T, Ohno M (1981) Preparation of oligoadenylares with 2′–5′ linkage using Pb2+ ion catalyst. Tetrahedron 37:481–485
Sawai H, Kuroda K, Hojo H (1989) Uranyl ion as a highly effective catalyst for internucleotide bond formation. Bull Chem Soc Jpn 62:2018–2023
Sawai H, Higa K, Kuroda K (1992) Synthesis of cyclic and acyclic oligocytidylates by uranyl ion catalyst in aqueous solution. J Chem Soc Perkin Trans 1992:505–508
Terfort A, von Kiedrowski G (1992) Self-replication by condensation of 3-Amino-Benzamidines and 2-Formylphenoxyacetic acids. Angew Chem Int Ed Engl 31:654–656
Walker JCG (1985) Carbon-dioxide on the early Earth. Orig Life Evol Biosph 16:117–127
White RH (1984) Hydrolytic stability of biomolecules at high temperatures and its implication for life at 250°C. Nature 310:430–432
Acknowledgement
Professor G. von Kiedrowski at Ruhr-Universitaet Bochum generously provided SIMFIT. This research was partially supported by the Grant-in-Aid for Scientific Research (C) (1550150) from Japan Society for the Promotion of Science (JSPS).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kawamura, K., Maeda, J. Kinetic Analysis of Oligo(C) Formation from the 5′-Monophosphorimidazolide of Cytidine with Pb(II) Ion Catalyst at 10–75°C. Orig Life Evol Biosph 37, 153–165 (2007). https://doi.org/10.1007/s11084-006-9063-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11084-006-9063-0