
Abstract
Contrafreeloading is the willingness of animals to work for food when equivalent food is freely available. This behavior is observed in laboratory, domesticated, and captive animals. However, previous research found that six laboratory cats failed to contrafreeload. We hypothesized that cats would contrafreeload in the home environment when given a choice between a food puzzle and a tray of similar size and shape. We also hypothesized that more active cats would be more likely to contrafreeload. We assessed the behavior of 17 neutered, indoor domestic cats (Felis catus) when presented with both a food puzzle and a tray across ten 30-min trials. Each cat wore an activity tracker, and all sessions were video recorded. Cats ate more food from the free feed tray than the puzzle (t (16) = 6.77, p < 0.001). Cats made more first choices to approach and eat from the tray. There was no relationship between activity and contrafreeloading, and there was no effect of sex, age, or previous food puzzle experience on contrafreeloading. Our results suggest that cats do not show strong tendencies to contrafreeload in the home environment, although some cats (N = 4) ate most food offered in the puzzle or showed weak contrafreeloading tendencies (N = 5). Eight cats did not contrafreeload. Cats who consumed more food from the puzzle, consumed more food in general, suggesting a relationship between hunger and effort. Further research is required to understand why domestic cats, unlike other tested species, do not show a strong preference to work for food.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Foraging is a natural behavior: most animals in the wild must forage in some way to survive, whether by searching or ambush (Huey and Pianka 1981; Stephens and Krebs 1986). Domesticated and captive animals often are encouraged to engage in foraging behaviors as a form of enrichment. This may take the form of providing opportunities for animals to work for food, for example by hiding food in a substrate such as straw that can be foraged (e.g., domesticated pigs and hamadryas baboons, de Jonge et al. 2008; Jones and Pillay 2004); providing enrichment that allows animals to extract seeds from holes (parrots, Coulton et al. 1997); scattering food in enclosures (Malayan sun bears, Schneider et al. 2014); through presentation of live prey (African lions and Sumatran tigers, Bashaw et al. 2003); and with puzzle feeders that require manipulation to extract food (Gray parrots, van Zeeland et al. 2013). One goal of feeding enrichment is to adjust captive animals’ time budgets to more closely mimic those of freely living animals.
When tested, many animals will work for food when similar food is freely available, a phenomenon known as contrafreeloading (Inglis et al. 1997; Jensen 1963). A preference for contrafreeloading is indicated when an animal works for 50% or more of all obtained food (Osborne 1977). Contrafreeloading contradicts optimal foraging theory, which suggests that animals should maximize energy gained while minimizing costs (Stephens and Krebs 1986).
Some proposed reasons for why animals contrafreeload include boredom in captive environments (McGowan et al. 2010), reducing uncertainty while in captivity or uncertain environments (McGowan et al. 2010), stimulation-seeking (Inglis et al. 1997), and information gathering (Inglis et al. 2001). Specific environmental conditions can increase or decrease contrafreeloading behavior: for example, food deprivation and an increase in required effort to obtain food tend to decrease contrafreeloading whereas sensory deprivation increases it (Bean et al. 1999; Inglis et al. 1997).
Contrafreeloading is found in various captive and companion species, including wild or domesticated animals housed in sanctuaries, zoos, laboratories and homes. Species tested include humans, chimpanzees, macaques, chickens, jungle fowl, pigeons, grizzly bears, maned wolves, rats, giraffes, and pigs (de Jonge et al. 2008; Inglis et al. 1997; Jensen et al. 2002; McGowan et al. 2010; Sasson-Yenor and Powell 2019; Vasconcellos et al. 2012). Contrafreeloading is tested by providing animals with a simultaneous choice of freely available food and food that is acquired through some form of operant response (Inglis et al. 1997).
McGowan et al. found that captive-living, wild-born grizzly bears spent more time interacting with food boxes that required effort to obtain reinforcement than with freely available food (2010). A study of domestic pigs found that they tended to work for food even when the same type of food was available without effort (de Jonge et al. 2008). Contrafreeloading also has been observed in adult humans, although the effects were age-dependent, with younger participants preferring to press a lever for candy or cash rewards compared to older participants (Tarte 1981).
Contrary to other species tested, a study of six reproductively intact domestic cats found no evidence for contrafreeloading. Cats were first trained to offer paw touches to a switch plate for food on a continuous reinforcement schedule. When offered a choice between delivering an operant response or eating the same type of food that was freely available in a dish, cats consumed the freely available food first (Koffer and Coulson 1971). Based on this result, cats have been described as the only species to show no contrafreeloading (Inglis et al. 1997). However, the previous study had a relatively small sample size (N = 6), and the experiment was conducted in the laboratory environment with cats maintained at 85–90% of their free-feeding body weights. As previous studies have found that increased hunger reduces contrafreeloading (Inglis et al. 1997), food restriction may be just one factor that impacted cats’ willingness to work for food.
Based on studies of contrafreeloading and foraging behavior of wild-born species living in captivity, foraging enrichment is often recommended for companion animals, such as dogs, cats, and parrots (e.g., Dantas et al. 2016; Meehan and Mench 2007; Schipper et al. 2008). Although contrafreeloading has been studied in domesticated species such as swine and poultry (de Jonge et al. 2008; Lindqvist and Jensen 2008, 2009), there have been few studies of preferences to work for food in companion animals.
The goal of the present study was to further assess contrafreeloading in indoor-only domestic cats by testing normally fed cats in their home environments at multiple times. Contrafreeloading has previously been correlated with exploration (e.g., Bean et al. 1999) and may allow an individual to gather information about their environment (Inglis et al. 1997). For this reason, we also included a measure of each cat’s activity level to assess whether there is a relationship between activity and contrafreeloading. We predicted that most domestic cats would contrafreeload, preferring to eat food from a food puzzle designed for cats over freely available food from a tray of the same size and shape. Further, we predicted that cats with higher activity levels would show a stronger preference for contrafreeloading compared to cats with lower activity levels.
Methods
All animal procedures were approved by the Animal Care and Use Committee at the University of California, Davis, under Protocol #21433. Data were collected between December 2019 and March 2020.
Participants
Twenty domestic cats (11 male, 9 female) between 1 and 10 years old (average age: 5.1 years, SD: 3.1 years) were enrolled in the study. All cats were privately owned, neutered, living indoors, and free of any medical conditions. All cats were the only cat living in the home and regularly ate commercially available dry cat food. All cats were fed their regular diet for training and tests. Six of the 20 cats had previous experience with using a food puzzle.
Equipment acclimation
Cats were acclimated to the experimental equipment for 4–12 days, depending on how quickly the cat adjusted. Owners of the participating cats were provided with a 13-g activity tracker (Fitbark Inc., Kansas City, MO) which was secured to a breakaway collar with zip ties and placed on each cat. The Fitbark™ is designed to detect movement via tri-axial accelerometer technology, and provides amalgamated activity levels for 1-min epochs when the device is worn. The device uses a proprietary point system to create an activity measure. Activity levels can be downloaded from the company’s website. Cat owners did not have access to the raw data or website unless requested after their participation was complete.
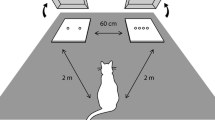
Cats were simultaneously introduced to the Trixie Pet Tunnel Feeder food puzzle (Trixie Pet Products, Fort Worth, TX) using a protocol similar to that used by Naik et al., 2018. The puzzle requires cats to scoop food out of tunnels/compartments using their paws. Cats were also given a round plastic tray of an identical diameter as the puzzle as the control (Fig. 1). For the first 1–3 days, cats were given their regular amount of food on the plastic tray with an additional 25% of their daily intake provided in the puzzle. Cats who successfully consumed some food from the puzzle by that time were next provided 25% of their daily food rations in the puzzle and 75% on the tray for 1–3 days. The food was then split evenly between the tray and puzzle for 1–3 days, and on the last 1–3 days of training, the cats were provided 75% of their food in the puzzle, and 25% on the tray. Three cats (2 female, 1 male) who refused to consume any food from the puzzle during training were withdrawn from the study for their safety. Cats that successfully completed the training were then exposed to the testing protocol.

An example of the experimental set up with the food puzzle (Trixie Tunnel Feeder) and the tray of identical size/shape
Procedures
Cat owners were provided with a WyzeCam camera (Wyze Labs, Seattle, WA) and a secure digital (SD) card to record all trials. For each trial, cats were provided with an equal amount of food in the free food tray and the puzzle. Cat owners recorded two to four trials per day over the course of 3–4 days and were asked to wait at least 2 h between trials. Depending on the number of trials per day, the cat was given an appropriate percentage of their daily rations at each trial (e.g., if the owner was conducting four trials in a day, they gave their cat 25% of their daily rations for one trial).
Owners were provided with a standard food scale to weigh and record the amount of food offered in the puzzle and tray for each trial. Owners were also given a predetermined randomized sequence (random.org) for placing the puzzle on either the left or right side of the tray on each trial, to reduce any effects of side bias or preference. The cat was presented with the puzzle on the left or right an equal number of times. Owners were asked to place the puzzle and tray near each other, equidistant from the cat at the time of presentation. Owners were also asked to record the date and time each trial started, and the initial and final amount of food left on both the puzzle and tray. At the end of each trial, any remaining food was provided to the cat in their regular feeding dish. Cats were given 30 min to eat from the puzzle and tray for each trial, at which point both were removed and the recording was stopped.
Behavior coding
Videos of trials were coded using BORIS (Behavioral Observation Research Interactive Software; (Friard and Gamba 2016)). We coded anytime the cat was eating from either the puzzle or tray, as well as any sniffing or touching of the puzzle or tray. Since discriminating between sniffing and eating was not always possible depending on the camera angle, we consolidated these interactions (eating, sniffing, and touching) into one measure for both the puzzle and the free food tray.
Five coders assessed the videos. All coders trained on a subset of 20–30 min of video, then were provided with additional training if needed. Overall inter-rater reliability of coders, as calculated by BORIS, was substantial, average Cohen’s kappa = 0.81 (range 0.73—0.91).
Data analysis
To assess the contrafreeloading and activity of domestic cats, we used the following data: amount of food consumed from the puzzle and free food tray, time spent interacting with the puzzle and tray, proportion of first choice for the puzzle for interacting and eating during each trial, and the activity measure downloaded from the Fitbark™.
Consumption from puzzle (P total ) and tray (T total )
The total food consumed from the puzzle and free food tray for each cat was calculated by adding the amount of food eaten from each for all trials.
Strength of preference to feed and interact with puzzle (contrafreeloading)
The presence of contrafreeloading behavior was assessed using the following equations:
To explore the consistency of each cat’s behavior across trials, we also calculated CFfeeding for each trial for each cat, which is depicted in Fig. 2. If the cat spent more time at, or consumed more food from, the puzzle than the free food tray, CFfeeding and CFtime will be greater than 0.5. A complete preference for the puzzle would result in values of 1. Equal time and consumption from both puzzle and tray would lead to values of 0.5. If the cat did not interact with or eat from the puzzle, CFfeeding and CFtime would be 0 (complete preference for the tray).

Data for each cat for each trial of the study. The figure depicts the cats’ first choices for interacting or feeding, and presents the data for CFfeeding for each trial. A value of 1 indicates complete preference for feeding from the puzzle, a value of 0.50 indicates consumption of the same amount of food from both, and a value of 0 indicates complete preference for feeding from the tray
First choice for puzzle (C p )
Cp values were calculated by the following equation:
Cp = number of first choices made for the puzzle/total number of sessions.
We calculated Cp by observing both the first interaction a cat had (ICp; e.g., sniffing or touching either the puzzle or free food tray) as well as for which source they chose to eat from first (ECp). If the cat immediately started eating, then his/her choice (puzzle or tray) was marked as both the first choice for interacting and eating. Cp represents the proportion of first choices that were for the puzzle. Each cat’s choice behavior for each trial is depicted in Fig. 2, and the values for ICp and ECp are in Table 1.
Statistical analyses
All data were analyzed using SAS University Edition (SAS Institute Inc, Cary, NC). To assess the potential effects of individual differences, we used general linear models to determine if there were any effects of sex, age, and previous food puzzle experience on CFfeeding and CFtime. We used a paired t test to compare the amount of food eaten from the puzzle to that eaten from the free food tray by each cat. Finally, we used correlation tests to assess the relationships between activity and contrafreeloading (CFfeeding and CFtime).
Results
Due to equipment error, activity data were not collected for one cat (Pee Wee). In addition, for two cats (Orion and Benedict), video data were collected for only 9 out of 10 trials. All other participants had complete video data for all 10 trials. All data used for analyses are presented in Table 1 and the supplementary data file.
We used general linear models to assess the effects of sex, age, and prior food puzzle experience on the two measures of contrafreeloading, CFfeeding and CFtime. The overall model for the effects of sex, age and experience on CFfeeding was not statistically significant (F (3,13) = 2.93, p = 0.07). One factor, sex, had a p value < 0.05, but this should be interpreted with caution since the overall model was not statistically significant. The overall model for the effects of sex, age and experience on CFtime was not statistically significant (F (3,13) = 1.72, p = 0.21). None of the individual factors had a p value < 0.05.
We used a paired t test to compare the amount of food that cats consumed from the puzzle and the free food tray. Although most cats ate some food from both sources, the amount of food consumed from the tray was statistically higher than that eaten from the puzzle (t (16) = 6.77, p < 0.001). Almost half of the cats (N = 8, 5 females) consumed less than 10% of the food offered to them from the puzzle, and two of those cats (both female) ate no food offered from the puzzle during sessions.
To test the hypothesis that activity levels were related to contrafreeloading, we compared CFfeeding and CFtime with the average daily activity of each participating cat. Since larger values indicate more expression of contrafreeloading, CFfeeding and CFtime should be positively correlated with activity if cats contrafreeload. There was no correlation between CFfeeding and activity, Spearman correlation coefficient r (16) = −0.09, p = 0.75. There was no correlation between CFtime and activity, Spearman correlation coefficient r (16) = 0.21, p = 0.44. As an additional exploratory analysis, we assessed the correlation between the amount of food eaten from the tray and food eaten from the puzzle. The results suggested a positive relationship between the two, Spearman correlation coefficient r (16) = 0.56, p = 0.02; cats who ate more food from the tray also tended to eat more food from the puzzle.
Based on these results, we would classify four cats as willing to contrafreeload, five cats as weakly contrafreeloading, and eight cats as freeloaders (Table 1). Since no cats consumed more food from the puzzle than the free food tray, we would not classify any cats as strong contrafreeloaders.
Discussion
In this study, we tested for the first time whether domestic cats living in homes would contrafreeload, as has been demonstrated in several captive and domesticated species. We did not find strong evidence for contrafreeloading; instead, cats preferred to eat the food that was freely available with no required additional effort. This was true when looking at both the overall population of cats, and the behavior of individual cats across trials.
We were unable to identify specific individual traits (sex, age and previous experience with a food puzzle) that predicted whether cats would contrafreeload. The four cats who appeared to be contrafreeloaders had one trait in common; they ate most of the food available to them during trials. In fact, the strongest predictor of amount of food eaten from the puzzle was the amount of food eaten from the tray.
The unanswered question is why cats, among multiple species tested, appear to be the only one that does not reliably contrafreeload. This tendency appears to contradict the fact that cats naturally work for food by hunting and will stop eating to hunt additional prey (Adamec 1976; Leyhausen 1979). Some differences in contrafreeloading tendencies among species have been attributed to domestication, such as in one study where white leghorn layers contrafreeloaded less than their ancestral species, jungle fowl (Jensen et al. 2002). Possible explanations included selection for allocation of resources toward reproduction and individual growth, and less need for information gathering when food sources are stable.
The cats in our study were all spayed or neutered and housed indoors only, which might have impacted their behavior. Spaying and neutering decreases the metabolic rate (Fettman et al. 1997) and the physical activity of female cats (Belsito et al. 2009), while simultaneously increasing cats’ food intake (Alexander et al. 2011; Wei et al. 2014). However, the previous study that failed to find evidence of contrafreeloading in cats only included reproductively intact cats (Koffer and Coulson 1971). To date, no studies have directly compared the activity patterns of indoor cats with those who have outdoor access. Thus, it would be premature to predict different results from reproductively intact or outdoor cats.
We also found no relationship between activity and contrafreeloading behavior. This result supports a previous study that found no change in activity level after introducing food puzzles to cats (Naik et al. 2018). Another study found that when two laboratory cats were required to offer an increasing number of touches to a switchplate in exchange for free access to food, they decreased the number of meals per day, and consumed more food at each feeding period (Collier et al. 1997). Studies of free-roaming feral cats suggest they spend almost 90% of their time inactive, with < 1% of their time spent hunting (Hernandez et al. 2018). In general, cats appear to conserve energy to the greatest extent possible, minimizing the amount of time and effort required to meet their caloric requirements, whether by hunting or engaging with enrichment devices in homes.
Our conclusion that cats prefer freely available food over that which requires effort is limited by some aspects of our study, such as a relatively small sample size. It is possible that the appearance of the food puzzle hindered consumption, as food was more visible in the tray. However, the cover of the puzzle was translucent, and several of the compartments face upward, such that the food would be visible from above. Although the novelty or shape of the food puzzle could have been a deterrent to the cats, we controlled for this by presenting all cats with a novel tray of the same shape and size simultaneously. All cats ate from both the puzzle and the tray during the training period, so we do not believe that appearance, novelty, or aversion to the device itself can explain the failure to contrafreeload.
We also did not control for food intake or hunger, as we did not want to increase the stress levels of cats, who can be sensitive to changes in feeding regimens (Stella et al. 2011). Food restriction can also reduce contrafreeloading (Inglis et al. 1997), so we did not want food withheld from cats longer than they were accustomed to. Since the cats ate a large percentage of the freely available food during trials, we cannot blame the failure to contrafreeload on a lack of interest in food. However, as we only gave cats access to the tray and puzzle for 30 min per trial, it is possible that cats would have engaged more with the puzzle if it continued to be available throughout the day.
Interpretation of our results may be dependent on how contrafreeloading is defined. Sometimes contrafreeloading is defined as when an animal will work for any food in the presence of freely available food, whereas some consider it a preference to work for food (Inglis et al., Osborne et al.). Most cats in our study did eat some food from the puzzle but none ate more food from the puzzle than the tray. Thus, although we have evidence for some willingness to work for food when freely available food is present (weak contrafreeloading), there is no evidence that cats preferred to work for food.
Of the species who have been tested for contrafreeloading, few predatory species (chimps, humans, and cats) are included, and most species tested are foragers who use extended search to acquire food (e.g., pigeons, rats, gerbils, Inglis et al. 1997). Sit-and-wait predation is considered low cost and is a common hunting style among felids (Williams et al. 2014). A full discussion of predatory energetics is beyond the scope of this manuscript, but contrafreeloading, which provides information about the quality of food patches, is expected to be weaker in species that do not engage in prolonged search (Inglis et al. 1997). Future studies should investigate whether foraging style is an important factor in contrafreeloading tendencies, and whether energy conservation is influential.
Like other studies, we did not find a statistically significant effect of sex on contrafreeloading (Lindqvist and Jensen 2008; Vasconcellos et al. 2012). However, in the current study, the four cats who did show a tendency to contrafreeload were all males, and the two cats who did not eat any food from the puzzle were both females. Male cats are more prone to obesity (Lund et al. 2005) and may be more food motivated in general. Our findings suggest the sex of the cat should be considered in future studies.
Future research can further explore contrafreeloading in cats by introducing different types of food puzzles or operant behaviors necessary for obtaining food. Changing the value of the food offered may also increase contrafreeloading, as novelty of food items increases the level of reinforcement (Inglis et al. 1997). The effects of foraging enrichment on cat welfare and health indicators should also be assessed. A presentation of case studies found positive effects of food puzzles for domestic cats, such as weight reduction and an improvement of behavioral health (Dantas et al. 2016), but puzzles do not appear to increase overall activity levels (Naik et al. 2018).
Understanding contrafreeloading is important for captive and domestic animal welfare as foraging enrichment is a frequently used tool to provide choice and mental stimulation. The effects of such enrichment on the behavior of captive animals are rarely tested. For domestic cats, the provision of foraging enrichment may depend on the needs and food motivation of the individual animal, and may be best introduced as a choice to enhance welfare, as foraging enrichment has been suggested to do in other species (e.g., Tarou and Bashaw 2007).
Availability of data and material
Data are available to download at https://doi.org/10.25338/B8GG92.
Code availability
Not applicable.
Change history
18 August 2021
A Correction to this paper has been published: https://doi.org/10.1007/s10071-021-01546-9
References
Adamec RE (1976) The interaction of hunger and preying in the domestic cat (Felis catus): an adaptive hierarchy? Behav Biol 18:263–272. https://doi.org/10.1016/S0091-6773(76)92166-0
Alexander LG, Salt C, Thomas G, Butterwick R (2011) Effects of neutering on food intake, body weight and body composition in growing female kittens. Br J Nutr 106:S19–S23. https://doi.org/10.1017/S0007114511001851
Bashaw MJ, Bloomsmith MA, Marr MJ, Maple TL (2003) To hunt or not to hunt? A feeding enrichment experiment with captive large felids. Zoo Biol 22:189–198. https://doi.org/10.1002/zoo.10065
Bean D, Mason GJ, Bateson M (1999) Contrafreeloading in starlings: testing the information hypothesis. Behaviour 136:1267–1282. https://doi.org/10.1163/156853999500712
Belsito KR, Vester BM, Keel T, Graves TK, Swanson KS (2009) Impact of ovariohysterectomy and food intake on body composition, physical activity, and adipose gene expression in cats. J Anim Sci 87:594–602. https://doi.org/10.2527/jas.2008-0887
Collier G, Johnson DF, Morgan C (1997) Meal patterns of cats encountering variable food procurement cost. J Exp Anal Behav 67:303–310. https://doi.org/10.1901/jeab.1997.67-303
Coulton LE, Waran NK, Young RJ (1997) Effects of foraging enrichment on the behaviour of parrots. Anim Welf 6:357–363
Dantas LM, Delgado MM, Johnson I, Buffington CT (2016) Food puzzles for cats: feeding for physical and emotional wellbeing. J Feline Med Surg 18:723–732. https://doi.org/10.1177/1098612X16643753
de Jonge FH, Tilly SL, Baars AM, Spruijt BM (2008) On the rewarding nature of appetitive feeding behaviour in pigs (Sus scrofa): do domesticated pigs contrafreeload? Appl Anim Behav Sci 114:359–372. https://doi.org/10.1016/j.applanim.2008.03.006
Fettman M, Stanton C, Banks L, Hamar D, Johnson D, Hegstad R, Johnston S (1997) Effects of neutering on bodyweight, metabolic rate and glucose tolerance of domestic cats. Res Vet Sci 62:131–136
Friard O, Gamba M (2016) BORIS: A free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol Evol 7:1325–1330. https://doi.org/10.1111/2041-210X.12584
Hernandez SM, Loyd KAT, Newton AN, Gallagher MC, Carswell BL, Abernathy KJ (2018) Activity patterns and interspecific interactions of free-roaming, domestic cats in managed Trap-Neuter-Return colonies. Appl Anim Behav Sci 202:63–68. https://doi.org/10.1016/j.applanim.2018.01.014
Huey RB, Pianka ER (1981) Ecological Consequences of Foraging Mode. Ecology 62:991–999
Inglis IR, Forkman B, Lazarus J (1997) Free food or earned food? a review and fuzzy model of contrafreeloading. Anim Behav 53:1171–1191. https://doi.org/10.1006/anbe.1996.0320
Inglis IR, Langton S, Forkman B, Lazarus J (2001) An information primacy model of exploratory and foraging behaviour. Anim Behav 62:543–557. https://doi.org/10.1006/anbe.2001.1780
Jensen GD (1963) Preference for bar pressing over “freeloading” as a function of number of rewarded presses. J Exp Psychol 65:451–454. https://doi.org/10.1037/h0049174
Jensen P, Schütz K, Lindqvist C (2002) Red jungle fowl have more contrafreeloading than white leghorn layers: Effect of food deprivation and consequences for information gain. Behaviour 139:1195–1209. https://doi.org/10.1163/15685390260437335
Jones M, Pillay N (2004) Foraging in captive hamadryas baboons: Implications for enrichment. Appl Anim Behav Sci 88:101–110. https://doi.org/10.1016/j.applanim.2004.03.002
Koffer K, Coulson G (1971) Feline indolence: cats prefer free to response-produced food. Psychonom Sci 24:41–42
Leyhausen P (1979) Cat behaviour. Garland, New York
Lindqvist C, Jensen P (2008) Effects of age, sex and social isolation on contrafreeloading in red junglefowl (Gallus gallus) and White Leghorn fowl. Appl Anim Behav Sci 114:419–428. https://doi.org/10.1016/j.applanim.2008.03.002
Lindqvist C, Jensen P (2009) Domestication and stress effects on contrafreeloading and spatial learning performance in red jungle fowl (Gallus gallus) and White Leghorn layers. Behav Processes 81:80–84. https://doi.org/10.1016/j.beproc.2009.02.005
Lund EM, Armstrong P, Kirk CA, Klausner J (2005) Prevalence and risk factors for obesity in adult cats from private US veterinary practices. Intern J Appl Res Vet Med 3:88–96
McGowan RT, Robbins CT, Alldredge JR, Newberry RC (2010) Contrafreeloading in grizzly bears: implications for captive foraging enrichment. Zoo Biol 29:484–502. https://doi.org/10.1002/zoo.20282
Meehan CL, Mench JA (2007) The challenge of challenge: can problem solving opportunities enhance animal welfare? Appl Anim Behav Sci 102:246–261. https://doi.org/10.1016/j.applanim.2006.05.031
Naik R et al (2018) Pilot study evaluating the effect of feeding method on overall activity of neutered indoor pet cats. J Vet Behav 25:9–13. https://doi.org/10.1016/j.jveb.2018.02.001
Osborne SR (1977) The free food (contrafreeloading) phenomenon: a review and analysis. Anim Learn Behav 5:221–235. https://doi.org/10.3758/BF03209232
Sasson-Yenor J, Powell DM (2019) Assessment of contrafreeloading preferences in giraffe (Giraffa camelopardalis). Zoo Biol 38:414–423. https://doi.org/10.1002/zoo.21513
Schipper LL, Vinke CM, Schilder MBH, Spruijt BM (2008) The effect of feeding enrichment toys on the behaviour of kennelled dogs (Canis familiaris). Appl Anim Behav Sci 114:182–195. https://doi.org/10.1016/j.applanim.2008.01.001
Schneider M, Nogge G, Kolter L (2014) Implementing unpredictability in feeding enrichment for Malayan sun bears (Helarctos malayanus). Zoo Biol 33:54–62. https://doi.org/10.1002/zoo.21112
Stella JL, Lord LK, Buffington CAT (2011) Sickness behaviors in response to unusual external events in healthy cats and cats with feline interstitial cystitis. J Am Vet Med Assoc 238:67–73. https://doi.org/10.2460/javma.238.1.67
Stephens DW, Krebs JR (1986) Foraging theory. Princeton University Press, Princeton, NJ
Tarou LR, Bashaw MJ (2007) Maximizing the effectiveness of environmental enrichment: suggestions from the experimental analysis of behavior. Appl Anim Behav Sci 102:189–204. https://doi.org/10.1016/j.applanim.2006.05.026
Tarte RD (1981) Contrafreeloading in Humans. Psychol Rep 49:859–866. https://doi.org/10.2466/pr0.1981.49.3.859
van Zeeland YRA, Schoemaker NJ, Ravesteijn MM, Mol M, Lumeij JT (2013) Efficacy of foraging enrichments to increase foraging time in Grey parrots (Psittacus erithacus erithacus). Appl Anim Behav Sci 149:87–102. https://doi.org/10.1016/j.applanim.2013.09.005
Vasconcellos AS, Harumi Adania C, Ades C (2012) Contrafreeloading in maned wolves: implications for their management and welfare. Appl Anim Behav Sci 140:85–91. https://doi.org/10.1016/j.applanim.2012.04.012
Wei A, Fascetti AJ, Kim K, Lee A, Graham JL, Havel PJ, Ramsey JJ (2014) Early effects of neutering on energy expenditure in adult male cats. PLoS ONE 9:e89557. https://doi.org/10.1371/journal.pone.0089557
Williams TM et al (2014) Mammalian energetics. Instantaneous energetics of puma kills reveal advantage of felid sneak attacks. Science 346:81–85. https://doi.org/10.1126/science.1254885
Acknowledgements
The authors thank Alice Mathews, Jin Seo, and Yucheng Celia Fan for video coding, and all the cat owners and cats who participated in this study. We thank the reviewers who provided helpful feedback on this manuscript.
Funding
MMD was supported by funding from Maddie’s Fund. The project described was supported by the National Center for Advancing Translational Sciences, National Institutes of Health, through Grant number UL1 TR001860 and linked award TL1 TR001861 to MMD. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH. The funding sources had no role in study design or in the collection, analysis, or interpretation of data.
Author information
Authors and Affiliations
Contributions
MMD, BH, and MJB designed the study. MMD and BH collected the data. MMD conducted the data analysis. All the authors discussed the results and contributed to the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
We declare no conflicts of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Delgado, M.M., Han, B.S.G. & Bain, M.J. Domestic cats (Felis catus) prefer freely available food over food that requires effort. Anim Cogn 25, 95–102 (2022). https://doi.org/10.1007/s10071-021-01530-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-021-01530-3