
Abstract
Given the accumulating evidence of climate change in sub-Saharan Africa, there is an urgent need to develop more climate resilient maize systems. Adaptation strategies to climate change in maize systems in sub-Saharan Africa are likely to include improved germplasm with tolerance to drought and heat stress and improved management practices. Adapting maize systems to future climates requires the ability to accurately predict future climate scenarios in order to determine agricultural responses to climate change and set priorities for adaptation strategies. Here we review the projected climate change scenarios for Africa’s maize growing regions using the outputs of 19 global climate models. By 2050, air temperatures are expected to increase throughout maize mega- environments within sub-Saharan Africa by an average of 2.1°C. Rainfall changes during the maize growing season varied with location. Given the time lag between the development of improved cultivars until the seed is in the hands of farmers and adoption of new management practices, there is an urgent need to prioritise research strategies on climate change resilient germplasm development to offset the predicted yield declines.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Over the next 50 years agriculture must provide for an additional 3.5 billion people (Borlaug 2007). Production of the three major cereal crops alone (maize, wheat and rice) will need to increase by 70 % by 2050 in order to feed the world’s growing rural and urban populations. However, climate change scenarios show that agricultural production will largely be negatively affected and will impede the ability of many regions to achieve the necessary gains for future food security (Lobell et al. 2008). The impact of climate change on agricultural production will be greatest in the tropics and subtropics, with sub-Saharan Africa (SSA) particularly vulnerable due to the range of projected impacts, multiple stresses and low adaptive capacity (IPCC 2007). Climate change scenarios for SSA include an increase in seasonal and extreme temperature events and intensity of droughts (IPCC 2007), and are likely to result in changes in production and the suitability of current crops. Africa is warming faster than the global average (Collier et al. 2008) and by the end of this century, growing season temperatures are predicted to exceed the most extreme seasonal temperatures recorded in the past century (Battisti and Naylor 2009). There remains greater uncertainty in projected changes in rainfall distribution patterns, with the outputs of climate models for future precipitation often not agreeing on the direction of change for SSA (IPCC 2007). There appears to be a general trend of increased precipitation in East Africa, with decreased precipitation in Southern Africa (IPCC 2007). For West Africa projected changes in rainfall vary greatly making it difficult to infer future climate scenarios (IPCC 2007; Cooper et al. 2008).
In large parts of SSA maize is the principal staple crop, covering a total of nearly 27 M ha (Table 1). Maize accounts for 30 % of the total area under cereal production in this region: 19 % in West Africa, 61 % in Central Africa, 29 % in Eastern Africa and 65 % in Southern Africa (FAO 2010). In Southern Africa maize is particularly important, accounting for over 30 % of the total calories and protein consumed (FAO 2010). Despite the importance of maize in SSA, yields remain low (Shiferaw et al. 2011). While maize yields in the top five maize producing countries in the world (USA, China, Brazil, Mexico and Indonesia) have increased three-fold since 1961 (from 1.84 t ha−1 to 6.10 t ha−1), maize yields in SSA have stagnated at less than 2 t ha−1, and less than 1.5 t ha−1 in Western and Southern Africa. In SSA maize is predominantly grown in small-holder farming systems under rainfed conditions with limited inputs. Low yields in this region are largely associated with drought stress, low soil fertility, weeds, pests, diseases, low input availability, low input use and inappropriate seeds. Reliance on rainfall increases the vulnerability of maize systems to climate variability and change. While farmers have a long record of adapting to the impacts of climate variability, current and future climate change represents a greater challenge because the probable impacts are out of the range of farmers’ previous experiences. Climate change will, therefore, severely test farmers’ resourcefulness and adaptation capacity (Adger et al. 2007).
While there is an urgent need to address policies and management strategies at both the country and international levels for agriculture adaptation to climate change, additional measures are also required to reduce the adverse effects of climate change on agricultural production. Past experience has demonstrated that the use of new varieties alongside improved management options can offset yield losses by up to 40 % (Thornton et al. 2009). Varieties with increased tolerance to abiotic stresses including heat and drought stress will play an important role in adaptation to climate change (Easterling et al. 2007; Fedoroff et al. 2010; Hellin et al. 2012). Given the time-lag between the development and the dissemination and adoption of new varieties, it is imperative that current research focuses on the development of improved climate-adapted germplasm and that greater attention is directed at facilitating farmers’ access to this germplasm. This paper reviews projected climate change scenarios for SSA and presents the possible future temperature, climates and monthly precipitation for maize mega-environments in SSA. Adaptation options for maize-based systems in SSA alongside present and future research requirements to develop maize germplasm with better adaptation to future African climates are discussed. The potential of improved maize germplasm to provide farmers with adaptation options to climate change is dependent on a functioning maize seed sector. One of the biggest challenges in the African maize seed sector is seed production. A case study is presented where the production of drought tolerant maize varieties in Eastern and Southern Africa (ESA) was scaled up to enable farmers’ access to improved varieties.
Predicted climate change scenarios in maize-growing regions of sub-Saharan Africa
During the last century, SSA has warmed at the average rate of 0.5°C per century (Hulme et al. 2001). Spatial pattern analysis revealed large variation in warming within SSA, with a maximum warming of 2°C per century in the Mediterranean basin of north-west Africa as well as over the interior of Southern Africa. Both day and night temperatures have continued to rise at the same rate (Solomon et al. 2007). Earlier climate change predictions showed land areas of the Sahara and semi-arid parts of Southern Africa warming by as much as 1.6°C by 2050 (Hernes et al. 1995; Ringius et al. 1996). Using greenhouse gas emission scenarios prepared for the Intergovernmental Panel on Climate Change (IPCC) Third Assessment Report, Hulme et al. (2001) forecast future warming across the Africa subcontinent at 0.2°C per decade (Special Report on Emissions Scenarios (SRES) B1 - low emissions scenario) to over 0.5°C per decade (SRES A2 – high emissions scenario). The greatest warming is predicted over Central and Southern Africa and the semi-arid tropical margins of the Sahara. Ruosteenoja et al. (2003) also showed higher levels of warming, at up to 7°C, in Southern Africa in September to November (SRES A1F1 emissions scenario).
Projected changes in precipitation during maize growing seasons vary greatly in SSA. Using the outputs of 21 global climate models (SRES A1B emissions scenario) the IPCC (2007) projected the median change in precipitation for West Africa to be −3 % during March–May and +2 % during June-August. In East Africa the median change in rainfall suggested precipitation would increase by 6 % between March and May and 4 % between June and August. In Southern Africa the median value of the 21 global climate models suggested no change in rainfall patterns between December and February and March and May. Collier et al. (2008) projected rainfall would increase by 2–4 % in Eastern Africa, but decrease by 5 % in Southern Africa during the main maize growing seasons (SRES A1B emissions scenario, 21 global climate models). Shongwe et al. (2009) estimated a general decrease in precipitation between September–November and March–May (12 global climate models, SRES A1B emissions scenario). The authors predict a notable delay in the onset of the rainy season in Southern Africa, with an early cessation of the rainy season in many areas within this region.
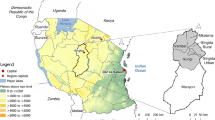
While large variation exists between global climate change models for regional application, the use of multiple models reduces the error in both the mean and variability (Pierce et al. 2009). At the country level, modelling using low spatial resolution masks large variation in key factors, such as climate and topography, and reduces the potential application of projections as decision making tools for identifying priority areas for research. To identify priority areas at the regional level for targeting drought and heat stress tolerant maize breeding programs, we used downscaled outputs from 19 SRES models and the A2 emissions scenario with data provided by the International Center for Tropical Agriculture (CIAT) (Ramirez and Jarvis 2008). The period 2040 to 2069 was chosen and the average of this period is referred to as 2050. Global climate models used were: BCCR-BCM 2.0, CCCMA-CGM2, CCCMA-CGCM3.1 T47, CCCMA-CGCM3.1 T63, CNRM-CM3, IAP-FGOALS-1.0G, GISS-AOM, GFDL-CM2.1, GFDL-CM2.0, CSIRO-MK3.0, IPSL-CM4, MIROC 3.2-HIRES, MIROC 3.2-MEDRES, MIUB-ECHO-G, MPI-ECHAM5, MIUB-ECHO-G, MPI-ECHAM5, MRI-CGCM2.3.2A, NCAR-PCM1, NIES99, UKMO-HADCM3. For temperature predictions countries were divided into maize production environments (also called maize mega-environments) (Bänziger et al. 2006; Hartkamp et al. 2000) based on rainfall and temperature data (Fig. 1). For rainfall predictions 16 areas across all mega-environments were selected. Areas chosen were important regions for maize production (Fig. 2). Climate data were downscaled to a 2.5 min (ca 5 km) resolution using an empirical statistical approach. For this, linear or other relationships were established between historically observed climate data at local scales, such as meteorological station measurements and climate model outputs (Ramirez and Jarvis 2010). ArcGIS software was used to calculate averages and standard deviations of model outputs for temperature (min, max, average) for all 19 models (Ormsby et al. 2009). Historical climate data (temperature and rainfall) which was obtained from the worldclim 1.4 dataset, at 2.5 min resolution, was used as a reference for long-term average values (1950–2000) (Hijmans et al. 2005). Values within mega-environments within the respective countries were averaged.

Maize environments within sub-Saharan Africa (adapted from Hodson et al. 2002)

Location of sites chosen for rainfall projections
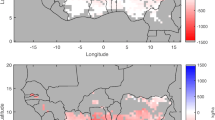
Average temperatures are predicted to increase by 2.4°C within the wet lowlands and wet low and upper mid-altitude environments, and by 2.6°C, 2.5°C and 2.4°C in the dry lowlands, dry mid-altitude and highland environments, respectively (Fig. 3). Both maximum and minimum temperatures are predicted to increase, with a greater increase in maximum temperatures. Maximum temperatures are predicted to increase by 2.5°C in wet-upper mid-altitudes, 2.6°C in wet lowlands and wet lowland mid-altitude, 2.7°C in dry lowlands and 2.8°C in dry mid-altitude environments (Fig. 4), while minimum temperatures are predicted to increase by 2.0°C in the wet lower and upper mid-altitude, wet lowland and dry mid-altitude, and dry lowland environments.

Increase in average temperatures in maize mega-environments between 2050 and 1960–200 using the outputs of 19 GCM’s and A2 emissions scenarios

Increase in maximum temperatured in maize mega-environments between 2050 and 1960–2000 using the outputs of GCM’s and A2 emissions scenarios. Regions with a 40 % probability of a failed season due to drought stress are shaded in grey (adapted from Jones 2008)
In maize, increasing maximum (day) temperatures have a greater, negative impact on yields than minimum (night) temperatures (Lobell et al. 2011). Average optimum temperatures for temperate, highland tropical and lowland tropical maize lie between 20 and 30°C, 17 and 20°C, and 30 and 34°C, respectively (Badu-Apraku et al. 1983; Brown 1977; Chang 1981; Chowdhury and Wardlaw 1978; Cross and Zuber 1972; Dale 1983; Jiang et al. 1999; Jones et al. 1984; Shaw 1983). Maximum temperatures currently exceed optimal temperature conditions for lowland tropical maize (34°C) in several countries (Burkina Faso, Chad, Eritrea, Gambia, Mali, Mauritania, Niger, Nigeria, Senegal and Sudan), however the area of maize grown within several of these regions is small. Lobell et al. (2011) showed that maize yields in Southern Africa declined linearly by 1 % under optimal conditions and 1.7 % under drought stress for every accumulated degree day above 30°C. In the dry lowlands of Burkina Faso, Cameroon, Chad, Gambia, Mali, Mauritania, Niger and Senegal mean temperatures are predicted to increase above 30°C and could reduce maize yields.
Precipitation projections suggest monthly rainfall patterns will change in all locations, however the direction and magnitude of change varies with location (Fig. 5). In the highlands of Ethiopia rainfall will decrease during the maize growing season (May–October), particularly during the critical reproductive stage. In West Africa rainfall will decrease during the maize growing season in the wet upper mid-altitude of Nigeria and the wet lowland of Benin; however in the wet lowland mid-altitude of Nigeria and the wet lowland of Ghana, total rainfall during the maize growing season will increase. In East Africa there is a consistent increase in rainfall between December and February across mega-environments. In the wet lowland mid-altitude of Kenya there is little change in total rainfall during the maize growing season, however in the dry lowlands rainfall is projected to decrease during the maize reproductive stage, with the onset of the short rainy season also delayed. In Southern Africa the maize growing season is November–April. Monthly rainfall projections suggest a general increase in rainfall during the maize growing season, ranging from 3 % in the dry lowlands of Zimbabwe to 18 % in the dry lowlands of Mozambique. However, in agreement with Shongwe et al. (2009), the onset of the rainy season is delayed. Low rainfall in May in all locations in Southern Africa is likely to result in a delay in planting. While rainfall during the maize growing season in the drought-prone lowlands Southern Africa may increase slightly, it is unlikely to translate into higher yields as evapotranspiration will increase under higher temperatures (Cook and Vizy 2012). Under climate change, increased extreme rainfall events are expected to increase (IPCC 2007), however, data available for modelling did not allow the prediction of rainfall distribution within each month thereby masking extreme events.

Monthly rainfall in 1960–2000 (black) and 2050 (white) using the outputs of 19 GCMs and A2 emissions scenarios in highlands (a, Ethiopia 1; b, Ethiopia 2), wet upper mid altitude (c Nigeria 1), wet lower mid altitude (d, Nigeria 2; e, Ethiopia 3; f, Kenya; g, Uganda; h, Tanzania; i, Malawi; j, Zambia), wet lowlands (k, Benin; l, Ghana) and the dry lowlands (m, Kenya 1; n, Kenya 2; o, Kenya3, p, Mozambique; q, Zimbabwe 1 and r, Zimbabwe 2). Details of each location are presented in Fig. 2
Reducing the effects of climate change on farmers in maize-based systems
Adapting maize germplasm to future climates
Substantial gains in cereal yields have been made through breeding (Fischer and Edmeades 2010). In 1997 CIMMYT initiated a product-orientated maize breeding programme in Southern Africa to increase yields in low input, drought-prone environments (Bänziger and Diallo 2001). Maize varieties were simultaneously selected for performance under optimal, low nitrogen and managed drought stress conditions. CIMMYT hybrids yielded more than commercial checks at all yield levels (Bänziger et al. 2006). Under severe drought stress CIMMYT hybrids had a 40 % yield advantage compared to commercially available hybrids. Since 2006, phenotyping capacity for managed drought screening in maize in ESA has increased from 6 ha to 35 ha under the Drought Tolerant Maize for Africa project (funded by the Bill and Melinda Gates Foundation). This increase has allowed greater efforts to be directed towards increasing drought tolerance in maize. Recent on-farm trials in ESA of new drought tolerant hybrids showed a 35 % yield advantage against farmers own varieties under low (<3 t ha−1) yield conditions (Setimela et al. 2012). The best hybrid (CZH0616) out-yielded the most popular commercial check by 26 %. These results highlight the potential of maize breeding to increase yields in unfavourable environments.
Modelling maize yields using climate projections for temperature and precipitation in ESA revealed that a 2°C increase in temperature will result in a greater reduction in maize yields than a 20 % decrease in precipitation (Lobell and Burke 2010; Rowhani et al. 2011). In the 1970s when CIMMYT initiated a maize drought breeding programme in Mexico, maize germplasm was screened across a range of environments including an environment with higher temperature (Edmeades et al. 1997). As a result of indirect selection for heat stress tolerance, drought tolerant maize developed through this program generally had a higher level of tolerance to higher temperatures (Cairns et al. 2013). In contrast, until recently maize breeding for drought tolerance in ESA did not include a high temperature environment. Screening a range of maize lines from breeding programs at CIMMYT and the International Institute for Tropical Agriculture (IITA) in high temperature environments revealed maize germplasm developed in ESA tended to be susceptible to both heat stress and drought stress at higher temperatures (Cairns et al. 2013). These results highlight the need to incorporate high temperature environments into the maize breeding pipeline within ESA. Direct selection for heat stress in breeding will ensure drought tolerant maize varieties and donors that are highly susceptible to heat stress are eliminated from the breeding pipeline and increase gains in tolerance to heat stress in maize. Breeding for heat stress tolerance requires knowledge of the potential effects of temperature on maize growth and development and genetic variability for traits of interest. The main effects of progressive heat stress on maize production are associated with reduced growth duration, reduced light interception and reproductive failure (Stone 2001). The reproductive phase is the most sensitive growth stage to heat stress. In contrast to drought stress, the male reproductive tissue is more sensitive to heat stress than the female reproductive tissue (Dupuis and Dumas 1990). Pollen is produced in the tassel, which is located above the leaf canopy, providing maximum exposure to elevated temperatures (Cairns et al. 2012). High temperatures reduce the quantity and viability of pollen produced resulting in reduced fertilisation of ovules and therefore reduced sink capacity (Schoper et al. 1986). High temperatures at the early reproductive stage also delay the emergence of silks (female reproductive tissues) resulting in decreased flowering synchrony and further reducing fertilisation (Cicchino et al. 2011). Temperature primarily affects growth by reducing the length of time the crop can intercept radiation and the duration of the grain filling period (Badu-Apraku et al. 1983). In the field, delayed planting can be used to screen for tolerance to heat stress by ensuring higher temperatures are experienced during critical stages of plant growth (Craufurd et al. 2013). Using this method Cairns et al. (2013) recently reported large genetic variability for tolerance to heat stress in sub-tropical and tropical maize suggesting gains in maize tolerance to heat stress could be achieved through conventional breeding.
In Southern Africa, yield losses of maize under drought stress doubled when temperatures were above 30°C (Lobell et al. 2011). In rainfed environments, elevated temperatures will increase the evapotranspirative demand from the atmosphere, negatively affecting crop water balance and thus inducing drought stress. In water-limited conditions plants partially close their stomata to reduce water loss through transpiration, resulting in the leaves becoming warmer (Jackson et al. 1981) and canopy temperature can rise by over 9°C (Garrity and O’Toole 1995; Zia et al. 2013). Atmospheric CO2 is expected to double by the end of this century (Meehl et al. 2007). Increased CO2 concentration will stimulate photosynthesis in C3 plants but in C4 plants, such as maize, photosynthesis is saturated at current CO2 concentrations (Zaman-Hussain et al. 2013) and increased CO2 concentration will have no effect on photosynthesis (Leakey et al. 2009). However, elevated CO2 concentration will decrease evapotranspiration (and increase water use efficiency) through the partial closure of stomata (Žalud and Dubrovsky 2002). While this may increase soil water availability and delay the onset of drought, partial stomatal closure will also decrease transpirational cooling and is likely to result in an increase in canopy temperature (Markelz, et al. 2011). It is highly likely that the combination of drought and heat stress will increase in many areas of SSA. For example, while maximum temperatures by 2050 will not increase above threshold temperatures for lowland tropical maize in the drought-prone dry lowlands of Southern Zimbabwe, northern Botswana and northern Namibia, maize crops are highly likely to experience heat stress during drought stress. In model species, the combination of drought and heat stress is found to be unique and could not be extrapolated from the additive effect of each stress individually (Mittler 2006). A recent study in maize confirmed that tolerance to combined drought and heat stress was genetically distinct from tolerance to each stress individually, and tolerance to either drought or heat stress did not confer tolerance to the combined effect of drought and heat stress (Cairns et al. 2013).
Mitigating the effects of climate change on maize-based systems
Agriculture is estimated to account for 20 % of anthropogenic greenhouse gases (IPCC 2007). The use of fertilisers, agricultural machinery for tillage practices and irrigation are associated with increased CO2 emissions through fuel consumption (Cairns et al. 2012). Improved agronomic practices such as conservation agriculture have the potential to mitigate global warming through reduced greenhouse gas emissions and increasing C sequestration. However maize production systems in SSA are predominately rainfed, relying on animal traction or manual land preparation, with very limited fertiliser use (smallholder farmers in SSA use less than 10 kg/ha (Morris et al. 2007)). Furthermore SSA accounts for just 3.6 % of the world emissions of CO2 (Collier et al. 2008). Thus adaptation to climate change may be more important than mitigation within this region.
New maize varieties with improved drought and heat tolerance will play an important role in adapting maize systems to climate change in SSA, however maize yields in this region are currently amongst the lowest in the world. Maize breeding alone will be unable to adequately contribute on the scale required to both increase current yield levels and offset potential yield losses associated with climate change and increased climate variability. As previously discussed, while growing season temperatures may not increase above the threshold temperatures for sub-tropical and tropical maize by 2050, the higher growing season temperatures will increase plant transpiration thereby increasing planting water use and reducing plant-available soil water. Improved soil water status has the potential to both buffer plants against the potential effects of drought stress under intermittent rainfall and reduce the severity of a drought event (Thierfelder and Wall 2010). Conservation agriculture is a system of agronomy based on minimum soil disturbance through ploughing, residue retention and crop rotations (Hobbs 2007). Conservation agriculture practices increase stored soil water by improved water infiltration, reduce evapotranspiration and reduce water runoff (Verhulst et al. 2010; Thierfelder and Wall 2012). Several studies have shown that conservation agriculture techniques are associated with increased soil moisture content (Ussiri and Lal 2009; Dendooven et al. 2012). Verhulst et al. (2011) showed that, under mild drought, yields of maize grain under conservation agriculture were 1.8 to 2.7 times higher than under conventional management practices. Higher soil water content under conservation agriculture may be an important mechanism by which maize production is buffered against short drought periods during the growing season (Fischer et al. 2002; Thierfelder and Wall 2009; Thierfelder and Wall 2010).
Farmers coping strategies to reduce climate risk
The impacts of climate change and increased climate variability on farmers’ livelihoods are less clear because farmers are not passive players but rather active ‘agents’ whose strategies and interactions shape the outcome of development within the limits of the information and resources available to them (Long 1992). Over time, farmers have developed coping strategies to shield them against uncertainties induced by year to year variation in rainfall (Cooper et al. 2008). Climate change is likely to affect an array of farmer decision making and crop production/management choices. Dixon et al. (2001) identified the typology of livelihood improvement strategies farmers seek so as to alleviate or escape poverty. Using this model, livelihood improvement strategies that may be available to farmers under climate change in Africa are listed below. Livelihood improvement strategies include both ex-ante and ex-post risk management options (Cooper et al. 2008).
-
Intensification – households increase financial or physical productivity of existing production patterns e.g. farmers increase yields by applying external inputs e.g. irrigation and improved varieties of crops that farmers are already growing and adoption of water-conserving land management practices such as conservation agriculture (as previously discussed).
-
Diversification – farmers expand into new or existing market opportunities in order to increase income or decrease income variability. This may include the cultivation of new products and on-farm processing to add value to an existing product. In the case of Southern Africa, this may mean switching from maize to more heat- and drought-tolerant crops such as sorghum and millet (Burke et al. 2009). However, while both crops have shorter growing periods and require less water than maize, farmers prefer to grow maize.
-
Expansion – farmers increase income or resources by their lands, or their herd size. Expansion may come about through the distribution of new lands via land reform, the accumulation in fewer hands of land abandoned by migrating farmers, or through the clearing of previously unused land.
-
Increased off-farm income – farmers resort to seasonal or long-term off-farm employment to supplement farm incomes. The income generated may be re-invested in agriculture, or in other household needs.
-
Exit from agriculture - takes place when farmers work in another farming system or pursue a non-farming lifestyle. Migration is a means of coping with climate variability (Adger et al. 2003).
The aforementioned livelihood strategies are likely to mitigate the detrimental impacts of climate change but in terms of farmers’ adaptation strategies, plant breeding will continue to play a critical role.
Present and future research requirements for climate-adapted germplasm
Given the time lag between the development of improved germplasm and farmers’ access to the seed, there is an urgent need to strengthen breeding strategies to offset predicted yield declines, and also to realise predicted yield increases (Burke et al. 2009; Thornton et al. 2009). Geographic information systems (GIS) will play an integral role in targeting breeding programs by predicting regions of vulnerability, targeting germplasm movement and identifying future climates for agricultural production environments. Future vulnerable regions should become a focus of maize breeding efforts to increase adaptive capacity. The prediction of future climates will also allow the identification of current spatial analogues of these climates which can be used to facilitate germplasm exchange for breeding (Burke et al. 2009). The concept of mega-environments was developed to increase the efficiency of applying breeding resources by grouping maize production regions into agriculture production environments with broadly similar environmental characteristics and adaptation patterns (Hartkamp et al. 2000). Maize mega-environments are delineated by environmental characteristics (maximum temperature, rainfall, and soil pH) and performance of maize genotypes in multi-environment trials (Hodson et al. 2002; Setimela et al. 2005). Climate change projections in this study will be important for the identification of regions which will need to be reassigned to different mega-environments in the future. Similarly, the application of conservation agriculture technologies in maize-based systems relies on the concept of grouping sites with similar environmental characteristics (rainfall and soil type), known as recommendation domains. GIS will play an essential role in the identification of locations with similar soil types and rainfall patterns to facilitate promotion and uptake of conservation agriculture techniques within the most appropriate locations.
Breeding for climate-adapted germplasm can be achieved using a combination of conventional, molecular and transgenic approaches. Conventional breeding has played an important role in increasing yields in both temperate and tropical maize (Duvick and Cassman 1999; Bänziger et al. 2006). Upscaling conventional breeding methodologies, especially phenotyping and double haploid technology, and training in these technologies will play critical roles in accelerating the development of climate-adapted germplasm (Prasanna et al. 2012). Emerging molecular technologies, including discovery of marker-trait associations using genome-wide association analysis, coupled with advances being made in genome-wide selection, promise to provide significant thrust in the efforts to develop germplasm with tolerance to multiple abiotic and biotic stresses in much shorter time-frames. The vast unexploited genetic diversity in the maize gene banks, especially in the landraces and the wild relatives is waiting to be explored (Ortiz et al. 2009) through the rapid advances in genotyping and the progress being made in phenotyping. Enhanced understanding of the value of these genetic resources could pave way for their more effective utilisation in introgressing novel alleles for stress tolerance and broadening the genetic base of the elite cultivars. Transgenic approaches could also provide an opportunity to further increase genetic variation for key abiotic stresses in the breeding products, especially for those traits with limited genetic variation in the maize gene pool.
Climate-adapted maize germplasm will have to incorporate packages of traits. In Southern Africa farmers are likely to require maize with tolerance to drought stress at elevated temperatures. Tolerance to both drought and waterlogging is becoming increasingly important in Asia and may be required by farmers in small areas in ESA (Cairns et al. 2012). Separate breeding programs using initial selection under only one stress may result in the loss of valuable alleles or genetic variability for additional stresses. Given that the combination of heat and drought stress is distinctive, crop improvement efforts must focus on the identification of traits and donors associated with combined tolerance to both of these stresses. The development of managed stress screening sites and standardized protocols for specific combinations of stresses predicted in the target environment will be important to facilitate climate-adapted germplasm development. Key to the success of field phenotyping for combined drought and heat stress will be the ability to develop reproducible protocols in terms of both the intensity and duration of drought and heat stress (Bänziger et al. 2006). Unlike field screening for drought or heat where stress can be imposed by planting in the dry season or late planting to experience elevated temperatures, combined heat and drought stress screening requires a period of elevated temperatures and no rainfall. Free air temperature enrichment (FATE) facilities, which allow temperatures to be controlled in field plots without an enclosure, will be important in the development of phenotyping capacity for combined heat and drought stress (Rehmani et al. 2011). The frequency of extreme heat events is also expected to increase under climate change (IPCC 2007). Progressive heat stress and a single, rapid event of heat stress have different effects on plant growth and yield (Corbellini et al. 1997) and thus FATE facilities will be important for research and breeding for rapid heat stress events.
Breeding strategies for developing climate-adapted germplasm must take into account the predicted and significant regional variation in temperatures. While the overall crop yields in Africa may fall by 10–20 % by 2050 due to higher temperatures and reduced rainfall (Jones and Thornton 2003), this figure masks regional variation. In some areas crop reductions will be greater (northern Uganda, southern Sudan, and the semi-arid areas of Kenya and Tanzania) while in other areas crops yields may increase (southern Ethiopia highlands, central and western highlands of Kenya and the Great Lakes Region) (Thornton et al. 2009). However our results show maximum temperatures within the highlands of Kenya and Ethiopia will increase above optimum temperatures for highland germplasm (Fig. 2). Thus, to achieve predicted yield gains, highland germplasm with improved tolerance to higher temperatures is required. This can be achieved through two pathways – improving heat tolerance within highland germplasm and facilitating farmers’ access to lowland sub-tropical and tropical germplasm.
Under optimal conditions yield potential is positively associated with days to maturity. Earliness is therefore associated with a yield penalty. However, in rainfed areas small-holder farmers often favour early maturing maize varieties. In a survey of 500 households in drought-prone maize growing areas of Ethiopia, Kenya, Tanzania and Uganda, early maturity was the second most desired attribute of maize varieties after yield potential (Erenstein et al. 2011a). Farmers perceived this trait as more important than drought tolerance. Early maturing maize varieties provide an effective drought avoidance strategy by increasing the probability of plants completing flowering, the most sensitive stage to drought, prior to the onset of drought and escaping terminal drought (Morris 2001). While breeding programs have substantially improved yields of early maturing maize varieties under low rainfall conditions, earliness continues to carry a yield penalty when rainfall levels are above average (Bänziger et al. 2000). However, increasing temperatures have the potential to further increase the yield penalty. In the drought-prone lowlands of Zimbabwe, extra early varieties that mature within 60–65 days are preferred by resource-poor farmers with no access to irrigation. Theoretically, an increase of 2.9°C throughout the growing season could reduce the number of days to maturity of an extra early hybrid from 65 to 58, while an early maturity maize variety would mature in approximately 64 days (with a daily increase of 2.9°C) as compared to 72 days under current climates. Given this reduction in growth duration (and its associated reduction in grain yield), it may be important for breeding programs targeting some drought prone areas, such as the lowlands of Zimbabwe, to shift their emphasis towards early maturing maize varieties rather than extra early maturing varieties.
Understanding and overcoming the obstacles to farmers’ access to improved germplasm
Maize seed sector in SSA
The potential of improved maize varieties as an adaptation strategy for climate change in SSA relies on a functioning, efficient seed sector. The maize seed sector in SSA has developed more slowly compared to other regions in the world (Tripp and Rohrbach 2001). Current maize seed sales are estimated to account for only 33 % of the total seed requirement in West and Eastern Africa and 38 % of the total seed requirements in Southern Africa (Abdoulaye et al. 2009; Langyintuo et al. 2008). Since the liberalisation of the seed sector in SSA in the early 1990s the total number of private seed companies within SSA over the last 20 years has increased four-fold (Langyintuo et al. 2010). During the same period seed production has only doubled suggesting that new seed companies are not increasing production in similar proportion (Langyintuo et al. 2010). However, there are significant differences in the use of improved seed by farmers between countries in ESA. For example both Zimbabwe and Kenya have high seed purchase rates, while Angola, Mozambique and Tanzania have low seed purchase rates. The need to identify factors limiting the production, multiplication and marketing of improved seeds in SSA is essential for promoting maize production, improving farmers’ income, alleviating poverty and ensuring food security (Abdoulaye et al. 2009). Surveys of the seed sector in SSA identified major bottlenecks that affect the entire maize value chain (Langyintuo et al. 2008; Abdoulaye et al. 2009; Erenstein et al. 2011b). Seed production was identified as the biggest obstacle in the seed sector across all regions in SSA. Seed companies often lack adequate land and related resources to produce enough seed, hence, to meet set targets they need to contract farmers to produce the seed (Langyintuo et al. 2010). The majority of farmers in SSA operate on small farms, increasing problems of identifying suitable large farms which would allow problems associated with foreign pollen contamination to be minimised. To increase the capacity of local seed companies to disseminate improved maize varieties to a greater number of farmers, training of contract farmers in seed production is required (MacRobert 2009).
Facilitating farmers access to improved maize varieties
Recent experiences through the Drought Tolerant (DT) Maize for Africa project have shown that increased seed production and dissemination of improved maize varieties is possible within a short time period. This project has adopted a three-pronged approach to support National Agricultural Research Systems (NARS) and seed companies to increase seed production. First, registration of improved maize varieties was expedited through varietal development and generation of data for national release. Since 2007 a total of 105 improved maize varieties have been released in 13 countries in SSA (Tsedeke 2012). Of these, 57 were open pollinated varieties (OPVs) and 48 were hybrids. Not all of these varieties were unique, with the same varieties registered in a number of countries. Second, seed production has been promoted through provision of foundation seed and training of seed entrepreneurs and technicians. The amount of foundation seed provided to private seed companies for certified seed production increased from minimal quantities in 2007 and 2008 to 17 t in 2009, 42 t in 2010 and 64 t in 2011. This helped support seed companies in the production of certified seed, such that the total certified seed production estimated from company feedback increased from 840 t in 2007 to 17,055 t in 2012 (Table 2). Hybrids have largely dominated the production in these years, except in 2011. The most significant production of certified seed of improved DT varieties was in Malawi, Kenya, Zimbabwe and Zambia (Kassie et al. 2012). Training was provided for technical staff within NARS and seed companies on trial and nursery management, variety registration, seed production and seed entrepreneurship. On-farm trials and demonstrations of improved DT varieties were conducted by NARS and seed companies in the target countries. On-farm trials provided farmers with direct interaction with new varieties, and because there were close linkages with the NARS extension services and seed companies, farmers were able to access information on new varieties and seed markets.
These interventions concentrated on the seed value chain from product development to product marketing. Unfavourable seed policies remain a bottleneck to the maize seed sector in SSA (Langyintuo et al. 2008). Policies associated with taxation, import and export restrictions continue to restrict the development of the seed sector. Further work is also still required on credit provision to seed companies and farmers, fertiliser, chemical and machinery provision, and grain market development. The lack of access to credit for seed companies will remain a significant bottleneck and until governments and development partners make credit available to seed entrepreneurs, directly or through risk sharing arrangements with commercial banks, national seed companies will not be able to grow as rapidly as required. As a consequence the seed sector will continue to be monopolized by the regional and multinational seed companies with the danger that farmers living in marginal areas are ignored. Additionally lack of credit to small-holder farmers is also cited as a major limitation to increased productivity (Morris et al. 2007). Lastly, it is considered that improving the market for grain in Africa would create a greater demand for maize, improve price stability for farmers and enable them to have more cash to purchase crop inputs.
Conclusions
While maize yields have steadily increased in over 70 % of maize growing areas, in SSA maize yields remain the lowest in the world and have stagnated since the early 1990s (Ray et al. 2012). Accumulating evidence of climate change in SSA suggests maize yields will decrease in many regions without the development of more climate resilient maize systems. Adaptation to climate change will require cross-disciplinary solutions (Howden et al. 2007) that include the development of appropriate germplasm and mechanisms to facilitate farmers’ access to the germplasm. Given the time taken from the development of climate-adapted germplasm to the diffusion of this germplasm, there is an urgent need to identify the most vulnerable regions and ensure both breeding programs and seed delivery systems are able to deliver improved varieties to offset potential losses under climate change within these regions. Earlier climate projections for SSA have tended to focus on upscaled projections or downscaled projections for individual countries/regions. The use of downscaled climate projections for maize growing regions in SSA was used to identify high priority geographic areas for the development and deployment of improved germplasm suited to the future climates. While temperatures are likely to increase throughout SSA by 2050, the combination of increasing temperatures and low seasonal rainfall in Southern Africa suggest this region will be particularly vulnerable, in agreement with Lobell et al. (2008). Breeding for improved stress tolerance within SSA has an admirable track record. To ensure that new varieties are tolerant to drought under future environments, particularly in Southern Africa, is it essential that higher temperature environments are included within the drought breeding pipeline. The ability of the seed system to deliver new varieties to farmers is often overlooked. The seed system in SSA has developed more slowly compared to other regions. Seed production was previously identified as the major bottleneck within the seed system of SSA. With support it is possible to overcome many of the obstacles within seed production. However at present the seed policy environment is not conducive to facilitating small-holder farmers’ access to the improved germplasm.
References
Abdoulaye, T., Sanogo, D., Langyintuo, A., Bamire, S. A., & Olanrewaju, A. (2009). Assessing the constraints affecting production and deployment of maize seed in DTMA countries of West Africa. Ibadan: IITA.
Adger, W. N., Agrawala, S., Mirza, M. M. Q., et al. (2007). Assessment of adaptation practices, management options, constraints and capacity. Climate Change 2007: impacts, adaptation and vulnerability. Contribution of Working Group II to the Forth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK and New York, USA.
Adger, W. N., Huq, S., Brown, K., Conway, D., & Hulme, M. (2003). Adaptation to climate change in the developing world. Progress in Development Studies, 3, 179–195.
Badu-Apraku, B., Hunter, R. B., & Tollenaar, M. (1983). Effect of temperature during grain filling on whole plant and grain yield in maize (Zea mays L.). Canadian Journal of Plant Science, 63, 357–363.
Bänziger, M., & Diallo, A. O. (2001). Progress in developing drought and stress tolerant maize cultivars in eastern and southern Africa. Seventh Eastern and Southern Africa Regional Maize Conference, 11th–15th February. pp. 189–194.
Bänziger, M., Mugo, S., & Edmeades, G. O. (2000). Breeding for drought tolerance in tropical maize: conventional approaches and challenges to molecular approaches. In J. M. Ribaut & D. Poland (Eds.), Molecular approaches for the genetic improvement of cereals for stable production in water-limited environments (pp. 69–72). Mexico D.F. Mexico: CIMMYT.
Bänziger, M., Setimela, P. S., Hodson, D., & Vivek, B. (2006). Breeding for improved abiotic stress tolerance in Africa in maize adapted to southern Africa. Agriculture and Water Management, 80, 212–214.
Battisti, D. S., & Naylor, R. L. (2009). Historical warnings of future food insecurity with unprecedented seasonal heat. Science, 323, 240–244.
Borlaug, N. (2007). Feeding a hungry world. Science, 318, 359.
Brown, D. M. (1977). Response of maize to environmental temperatures: a review. Agrometeorology of the Maize (Corn) Crop. World Meteorological Organization, 481, 15–26.
Burke, M. B., Lobell, D. B., & Guarino, L. (2009). Shifts in African crop climates by 2050, and the implications for crop improvements and genetic resources conservation. Global Environmental Change, 19, 317–325.
Cairns, J. E., Crossa, C., Zaidi, P. H., Grudloyma, P., Sanchez, C., Araus, J. L., et al. (2013). Identification of drought, heat and combined drought and heat tolerance donors in maize (Zea mays L.). Crop Science. doi:10.2135/cropsci2012.09.0545.
Cairns, J. E., Sonder, K., Zaidi, P. H., Verhulst, N., Mahuku, G., Babu, R., et al. (2012). Maize production in a changing climate. Advances in Agronomy, 144, 1–58.
Chang, J. H. (1981). Corn yield in relation to photoperiod, night temperature and solar radiation. Agricultural Meteorology, 24, 253–262.
Chowdhury, S. I., & Wardlaw, I. F. (1978). The effect of temperature on kernel development in cereals. Australian Journal of Agricultural Research, 29, 205–223.
Cicchino, M., Rattalino Edreria, J. I., Uribelarrea, M., & Otegui, M. E. (2011). Heat stress in field-grown maize: response of physiological determinants of grain yield. Crop Science, 50, 1438–1448.
Collier, P., Conway, G., & Venables, T. (2008). Climate change and Africa. Oxford Review of Economic Policy, 24, 337–353.
Cook, K. H., & Vizy, E. K. (2012). Impact of climate change on mid-twenty first century growing seasons in Africa. Climate Dynamics, 39, 2937–2955.
Cooper, P. J. M., Dimes, J., Rao, K. P. C., Shapiro, B., Shiferaw, B., & Twomlow, S. (2008). Coping better with current climatic variability in the rain-fed farming systems of sub-Saharan Africa: an essential first step in adapting to future climate change? Agriculture, Ecosystems and Environment, 126, 24–35.
Corbellini, M., Canevar, M. G., Mazza, L., Ciaffi, M., Lafiandra, D., & Borghi, B. (1997). Effect of the duration and intensity of heat shock during grain filling on dry matter, protein accumulation, technological quality and protein composition in bread and durum wheat. Australian Journal of Plant Physiology, 23, 245–260.
Craufurd, P. Q., Vadez, V., Jagadish, S. V. K., Prasad, P. V. V., & Zaman-Allah, M. (2013). Crop science experiments designed to inform crop modeling. Agriculture and Forestry Meteorology, 170, 8–18.
Cross, H. Z., & Zuber, M. S. (1972). Prediction of flowering dates in maize based on different methods of estimating thermal units. Agronomy Journal, 61, 351–355.
Dale, R. F. (1983). Temperature perturbations in the midwestern and southeastern United States important for corn production. In C. D. Raper & P. J. Kramer (Eds.), Crop reactions to water and temperature stresses in humid and temperate climates (pp. 21–32). Colorado: Westview Press.
Dendooven, L., Patiño-Zúñiga, L., Verhulst, N., Luna-Guido, M., Marsch, R., & Govaerts, B. (2012). Global warming potential of agricultural systems with contrasting tillage and residue management in the central highlands of Mexico. Agriculture, Ecosystems and Environment, 152, 50–58.
Dixon, J., Gulliver, A., & Gibbon, D. (2001). Farming systems and poverty: improving farmers livelihoods in a changing world. Rome, Italy and Washington DC, USA: FAO and World Bank.
Dupuis, I., & Dumas, C. (1990). Influence of temperature stress on in vitro fertilization and heat shock protein synthesis in maize (Zea mays L.) reproductive tissues. Plant Physiology, 94, 665–670.
Duvick, D. N., & Cassman, K. G. (1999). Post-green revolution trends in yield potential of temperate maize in North-Central United States. Crop Science, 39, 1622–1630.
Easterling, W., Aggarwal, P., Batima, P., et al. (2007). Food fibre and forest products. In M. L. Parry, O. F. Canziani, J. P. Palutikof, P. J. van Linden, & C. E. Hansen (Eds.), Impacts, adaptation and vulnerability’ contribution of working group II to the fourth assessment report of the intergovernmental panel on climate change (pp. 273–313). Cambridge, UK: Cambridge University Press, Cambridge, UK.
Edmeades, G. O., Bänziger, M., Cortes, M., & Ortega, A. (1997). From stress-tolerant populations to hybrids: The role of source germplasm. In G. O. Edmeades, M. Bänziger, H. R. Mickelson, and C. B. Peña-Valdivia (Eds.). Developing drought and low-N tolerant maize (pp. 263–273). Mexico D.F., Mexico: CIMMYT.
Erenstein, O., Kassie, G. T., Langyintuo, A., & Mwangi, M. (2011a). Characterization of maize producing households in drought prone regions of Eastern Africa. CIMMYT Socio-Economics Working Paper 1. Mexico, D.F., Mexico: CIMMYT.
Erenstein, O., Kassie, G. T., & Mwangi, W. (2011b). Challenges and opportunities for maize seed sector development in eastern Africa. Paper presented at the conference: Increasing Agricultural Productivity & Enhancing Food Security in Africa: New Challenges and Opportunities, 1–3 November 2011, Africa Hall, UNECA, Addis Ababa, Ethiopia.
FAO. (2010). FAO statistical database. Rome: Food and Agricultural Organization of the United Nations (FAO).
Fedoroff, N. V., Battisti, D. S., Beachy, R. N., et al. (2010). Radically rethinking agriculture for the 21st century. Science, 327, 833–834.
Fischer, R. A., & Edmeades, G. O. (2010). Breeding and cereal yield progress. Crop Science, 50, S85–S98.
Fischer, R. A., Santiveri, F., & Vidal, I. R. (2002). Crop rotation, tillage and crop residue management for wheat and maize in the sub-humid tropical highlands: I. Wheat and legume performance. Field Crops Research, 79, 107–122.
Garrity, D. P., & O’Toole, J. C. (1995). Selection for reproductive-stage drought avoidance in rice, using infrared thermometry. Agronomy Journal, 87, 773–779.
Hartkamp, A. D., White, J. W., Rodríguez Aguilar, A., Bänziger, M., Srinivasan, G., Granados, G., et al. (2000). Maize production environments revisited: a GIS-based approach. Mexico: CIMMYT.
Hellin, J., Shiferaw, B., Cairns, J. E., Reynolds, M., Ortiz-Monasterio, I., Bänziger, M., et al. (2012). Climate change and food security in the developing world: potential of maize and wheat research to expand options for adaptation and mitigation. Journal of Development and Agricultural Economics, 4, 311–321.
Hernes, H., Dalfelt, A., Berntsen, T., Holtsmark, B., Otto Naess, L., Selrod, R., et al. (1995). Climate strategy for Africa. CICERO Report 1995:3. Norway: University of Oslo.
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G., & Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25, 1965–1978.
Hobbs, P. R. (2007). Conservation agriculture: what is it and why is it important for future sustainable food production? Journal of Agricultural Science, 145, 127–137.
Hodson, D. P., Martinez-Romero, E., White, J. W., Jones, P. G., Bänziger, M. (2002). Asia Maize Research Atlas. Version 1.0. http://www.cimmyt.org/ru/services/geographic-information-systems/resources/maize-research-atlas. Accessed 15 January 2012.
Howden, S. M., Soussana, J. F., Tubiello, F., Chhetri, N., Dunlop, M., & Meinke, H. (2007). Adapting agriculture to climate change. Proceedings of the National Academy of Sciences, 104, 19691–19696.
Hulme, M., Doherty, R. M., Ngara, T., New, M. G., & Lister, D. (2001). African climate change: 1900–2100. Climate Research, 17, 145–168.
IPCC. (2007). Fourth assessment report: synthesis. http://www.ipcc.ch/pdf/assessment-report/ar4/syr/ar4_syr.pdf. Accessed 17 November 2007.
Jackson, R. D., Idso, S. B., Reginato, R. J., & Pinto, P. J. (1981). Canopy temperature as a crop water stress indicator. Water Resources Research, 17, 1133–1138.
Jiang, C., Edmeades, G. O., Armstead, I., Lafitte, H. R., Hayward, M. D., & Hoisington, D. (1999). Genetic analysis of adaptation differences between highland and lowland tropical maize using molecular markers. Theoretical and Applied Genetics, 99, 1106–1119.
Jones, P. (2008). Failed seasons drought model. GCP, Generation Atlas Beta. Texcoco, Mexico. http://gismap.ciat.cgiar.org/egiron/GenerationAtlas/.
Jones, R. J., Quatter, S., & Crookston, R. K. (1984). Thermal environment during endosperm division and grain filling in maize: effects of kernel growth and development in Vitro. Crop Science, 24, 133–137.
Jones, P. G., & Thornton, P. K. (2003). The potential impacts of climate change on maize production in Africa and Latin America in 2055. Global Environmental Change, 13, 51–59.
Kassie, G. T., Erenstein, O., Mwangi, W., La Rovere, R., Setimela, P., & Langyintuo, A. (2012). Characterization of maize production in Southern Africa: synthesis of CIMMYT/DTMA household level farming system surveys in Angola, Malawi, Mozambique, Zambia and Zimbabwe. Socio-Economics Program Working Paper 4, Mexico, D.F.: CIMMYT.
Langyintuo, A. S., Mwangi, W., Diallo, A., MacRobert, J., Dixon, J., & Bänziger, M. (2008). An analysis of the bottlenecks affecting the production and deployment of maize seed in eastern and southern Africa. Harare: CIMMYT.
Langyintuo, A. S., Mwangi, W., Diallo, A., MacRobert, J., Dixon, J., & Bänziger, M. (2010). Challenges of the maize seed industry in eastern and southern Africa: a compelling case for private–public intervention to promote growth. Food Policy, 35, 323–331.
Leakey, A. D. B., Ainsworth, E. A., Bernacchi, C. J., Rogers, A., Long, S. P., & Ort, S. R. (2009). Elevated [CO2] effects on plant carbon, nitrogen, and water relations: six important lessons from FACE. Journal of Experimental Botany, 60, 2859–2876.
Lobell, D. B., Bänziger, M., Magorokosho, C., & Vivek, B. (2011). Nonlinear heat effects on African maize as evidenced by historical yield trials. Nature Climate Change, 1, 42–45.
Lobell, B., & Burke, M. B. (2010). On the use of statistical models to predict crop yield responses to climate change. Agricultural and Forestry Meteorology, 150, 1443–1452.
Lobell, D. B., Burke, M. B., Tebaldi, C., Mastrandrea, M. D., Falcon, W. P., & Naylor, R. L. (2008). Prioritizing climate change adaptation and needs for food security in 2030. Science, 319, 607–610.
Long, N. (1992). From paradigm lost to paradigm regained: The case for an actor-oriented sociology of development. In N. Long & A. Long (Eds.), Battlefields of knowledge: the interlocking of theory and practice in social research and development’ (pp. 16–45). London: Routledge.
MacRobert, J. F. (2009). Seed business management in Africa. Harare: CIMMYT.
Markelz, R. J. C., Strellner, R. S., & Leakey, A. D. B. (2011). Impairment of C4 photosynthesis by drought is exacerbated by limiting nitrogen and ameliorated by elevated [CO2] in maize. Journal of Experimental Botany, 62, 3235–3246.
Meehl, G. A., Stocker, T. F., Collins, W. D., et al. (2007). Global climate projections. In S. Solomon, M. Qin, R. B. Manning, et al. (Eds.), Climate change 2007: the physical science basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press.
Mittler, R. (2006). Abiotic stress, the field environment and stress combination. Trends in Plant Science, 11, 15–19.
Morris, M. L. (2001). Assessing the benefits of international maize breeding research: an overview of the global maize impacts study. Part II of the CIMMYT 1999–2000 world maize facts and trends. Mexico City: CIMMYT.
Morris, M. L., Kelly, V. A., Kopicki, R. J., & Byerlee, D. (2007). Fertilizer use in African agriculture: lessons learned and good practice guidelines. Washington DC: World Bank.
Ormsby, T., Napoleon, E., Burke, R., & Groessl, C. (2009). Getting to know ArcGIS desktop. Updated for ArcGIS 9.3. Basics of ArcView, ArcEditor, ArcInfo. Redmond: ESRI Press.
Ortiz, R., Taba, S., Tovar, V. H. C., Mezzalama, M., Xu, Y., Yan, J., et al. (2009). Conserving and enhancing maize genetic resources as global public goods – a perspective from CIMMYT. Crop Science, 50, 13–28.
Pierce, D. W., Barnett, T. P., Santer, B. D., & Gleckler, P. J. (2009). Selecting global climate models for regional climate change studies. Proceedings of the National Academy of Sciences, 106, 8441–8446.
Prasanna, B. M., Chaikam, V., & Mahuku, G. (2012). Doubled haploid technology in maize breeding: theory and practice. Mexico, D.F.: CIMMYT.
Ramirez, J., & Jarvis, A. (2008). High resolution statistically downscaled future climate surfaces, International Centre for Tropical Agriculture CIAT, http://gisweb.ciat.cgiar.org/GCMPage, accessed 30 November 2010.
Ramirez, J., & Jarvis, A. (2010). Downscaling global circulation model outputs: the delta method. Decision and policy analysis Working Paper No.1. DAPA, CIAT, Cali, Colombia. http://gisweb.ciat.cgiar.org/GCMPage/docs/Downscaling-WP-01.pdf. Accessed 30 January 2012.
Ray, D. K., Ramankutty, N., Mueller, N. D., West, P. C., & Foley, J. A. (2012). Recent patterns of crop yield growth and stagnation. Nature Communications, 3, 1293.
Rehmani, M. I. A., Zhang, J., Li, G., Ata-Ul-Karim, S. T., Wang, S., Kimball, B. A., et al. (2011). Simulation of future global warming scenarios in rice paddies with an open-field warming facility. Plant Methods, 7, 1–16.
Ringius, L., Downing, T. E., Hulme, M., Waughray, D., & Selrod, R. (1996). Climate change in Africa – issues and regional strategy (pp. 154). CICERO Report No. 1996:8, CICERO, Oslo, Norway.
Rowhani, P., Lobell, D. B., Linderman, M., & Ramankutty, N. (2011). Climate variability and crop production in Tanzania. Agriculture and Forest Meteorology, 151, 449–460.
Ruosteenoja, K., Carter, T. R., Jylhä, K., & Tuomenvirt, H. (2003). Future climate in world regions: an intercomparison of model-based projections for the new IPCC emissions scenarios. Helsinki: Finnish Environment Institute.
Schoper, J. B., Lambert, R. J., & Vasilas, B. L. (1986). Maize pollen viability and ear receptivity under water and high temperature stress. Crop Science, 26, 1029–1033.
Setimela, P., Chitalu, Z., Jonazi, J., Mambo, A., Hodson, D., & Bänziger, M. (2005). Environmental classification of maize-testing sites in the SADC region and its implications for collaborative maize breeding strategies in the subcontinent. Euphytica, 145, 123–132.
Setimela, P., MacRobert, J., Atlin, G. N., Magorokosho, C., Tarekegne, A., Makumbi, D., & Taye, G. (2012). Performance of elite maize varieties tested on-farm trials in eastern and southern Africa. Presented at the ASA, CSSA, and SSSA International Annual Meetings, 21–24 October, 2012. Cincinnati, Ohio, USA.
Shaw, R. H. (1983). Estimates of yield reductions in corn by water and temperature stress. In C. D. Raper & P. J. Kramer (Eds.), Crop reactions to water and temperature stresses in humid and temperate climates (pp. 49–65). Colorado: Westview Press.
Shiferaw, B., Prasanna, B., Hellin, J., & Bänziger, M. (2011). Crops that feed the world 6. Past successes and future challenges to the role played by maize in global food security. Food Security, 3, 307–327.
Shongwe, M. E., van Oldenborgh, G. J., van der Hurk, B. J. J. M., de Boer, B., Coelho, C. A. S., & van Aalst, M. K. (2009). Projected changes in mean and extreme precipitation in Africa under global warming. Part I. Southern Africa. Journal of Climate, 22, 3819–3837.
Solomon, S. D., Qin, M., Manning, R. B., et al. (2007). Technical summary. In S. Solomon, M. Qin, R. B. Manning, et al. (Eds.), Climate change 2007: the physical science basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press.
Stone, P. (2001). The effects of heat stress on cereal yield and quality. In A. S. Basara (Ed.), Crop responses and adaptations to temperature stress (pp. 243–291). Binghamton: Food Products Press.
Thierfelder, C., & Wall, P. C. (2009). Effects of conservation agriculture techniques on infiltration and soil water content in Zambia and Zimbabwe. Soil and Tillage Research, 105, 217–227.
Thierfelder, C., & Wall, P. C. (2010). Investigating Conservation Agriculture (CA) Systems in Zambia and Zimbabwe to mitigate future effects of climate change. Journal of Crop Improvement, 24, 113–121.
Thierfelder, C., & Wall, P. C. (2012). Effects of conservation agriculture on soil quality and productivity in contrasting agro-ecological environments of Zimbabwe. Soil Use and Management, 28, 209–220.
Thornton, P. K., Jones, P. G., Alagarswamy, G., & Andersen, J. (2009). Spatial variation of crop yield response to climate change in East Africa. Global Environmental Change, 19, 54–65.
Tripp, R., & Rohrbach, D. (2001). Policies for African seed enterprise development. Food Policy, 26, 147–161.
Tsedeke, A. (2012). Five years of addressing the needs of smallholder farmers in Africa. DTMA Brief 1, Nairobi, CIMMYT-Kenya.
Ussiri, D. A. N., & Lal, R. (2009). Long-term tillage effects on soil carbon storage and carbon dioxide emissions in continuous corn cropping system from an alfisol in Ohio. Soil and Tillage Research, 104, 39–47.
Verhulst, N., Govaerts, B., Verachtert, E., Castellanos-Navarrete, A., Mezzalama, M., Wall, P., et al. (2010). Conservation agriculture, improving soil quality for sustainable production systems? In R. Lal & B. A. Stewart (Eds.), Advances in soil science: food security and soil quality (pp. 137–208). Boca Raton: CRC Press.
Verhulst, N., Nelissen, V., Jespers, N., Haven, H., Sayre, K. D., Raes, D., et al. (2011). Soil water content, maize yield and its stability as affected by tillage and crop residue management in rainfed semi-arid highlands. Plant and Soil, 344, 73–85.
Žalud, Z., & Dubrovsky, M. (2002). Modelling climate change impacts on maize growth and development in the Czech Republic. Theoretical and Applied Climatology, 72, 85–102.
Zaman-Hussain, M., Van Loocke, A., Siebers, M. H., Ruiz-Vera, U., Markelz, R. J. C., Leakey, A. D. B., et al. (2013). Future carbon dioxide concentration decreases canopy evapotranspiration and soil water depletion by field-grown maize. Global Change Biology. doi:10.1111/gcb.12155.
Zia, S., Romano, G., Spreer, W., Sanchez, C., Cairns, J., Araus, J. L., et al. (2013). Infrared thermal imagery as a rapid tool for identifying water-stress tolerant maize genotypes of different phenology. Journal of Agronomy and Crop Science, 199, 75–84.
Acknowledgments
This work was supported in part by the Climate Change, Agricultural and Food Security (CCAFS) programme of the Consultative Group on International Agricultural Research (CGIAR), the MAIZE CGIAR research program, the Regional Fund for Agricultural Technology (FONTAGRO), and the Drought Tolerance Maize for Africa (DTMA) project funded by the Bill and Melinda Gates Foundation and USAID. We thank Drs. Kevin Pixley, Gary Atlin and Milos Coric for their helpful advice and comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Cairns, J.E., Hellin, J., Sonder, K. et al. Adapting maize production to climate change in sub-Saharan Africa. Food Sec. 5, 345–360 (2013). https://doi.org/10.1007/s12571-013-0256-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12571-013-0256-x