
Abstract
To understand changes in biodiversity, we must take into consideration at least the effects of global warming like increased droughts, the shift in plant phenology with the lengthened vegetation period, and the possibility that alien species might benefit from them. For better predictions whether non-native species could benefit from global change and turn into invasive alien species we chose to study Aster squamatus, alien that is already present in Slovenia and is currently regarded as non-invasive. We studied the species’ reproductive success and micro-scale habitat properties. Results show high reproductive potential of A. squamatus and it is necessary to constrain accumulation of seeds in the soil seed bank for future invasions. According to habitat properties, especially C:N ratio, we can regard A. squamatus as an engineering species, fertilising its own habitat. Climate change, prolonging the vegetation period, would enable more biomass accumulation followed by rapid decomposition. Released nutrients can be better used by fast-growing species that start their development slightly later in the season than early spring species, which begin to grow by utilising nutrients from their storage organs. In such cases of engineering alien species, adapted management is needed.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Alien Plant Species Might Benefit from Global Warming
Since 2007, it has been widely accepted that the global climate is warming and that human-induced increases in greenhouse gas emissions are mostly the cause (Kerr 2007). Global warming has been recognised to have profound impact on physical and biological systems. Of particular concern is the effect of global warming on biological diversity. Predictions of extinction have estimated that 20–30 % of species might face increasingly high risk of extinction. However, mechanisms of species persistence could lower the estimated extinction rate, while taking into consideration other human impacts (e.g., habitat destruction, landscape fragmentation or alien species introduction) could account for additional biodiversity loss (Botkin et al. 2007). Invasive species have been recognised as one of the most salient threats to biodiversity. Non-native species occurring in habitats where they were not present before the introduction are termed invaders or alien species. Rapidly increasing problems caused by invasive alien species worldwide demand the effective implementation of various policies aimed at reducing the impact of potentially or currently problematic non-native species (Richardson and Pyšek 2004).
To understand changes in biodiversity, we must take into consideration at least the effects of global warming and the possibility that alien species might benefit from them. The future impact of global climate warming will vary from region to region. The most striking variation will involve changes in precipitation, and drought is expected in some areas like the Mediterranean (Kerr 2007). To these changes local species might respond with forced migrations. On the other hand, the alien invasive species already found in the Mediterranean might respond differently. In some cases, low soil water availability reduces their invasiveness (Alpert et al. 2000; Stohlgren et al. 2001), while some other alien species can tolerate drought better than local species (Williams and Black 1994; Milchunas and Lauenroth 1995; Nernberg and Dale 1997). In the latter case, such aliens might extend their range boundaries. Drought might have a particular effect on increased invasibility of fast-growing alien species if habitats are un-shaded (Schumacher et al. 2008).
Another recognised impact of global warming is the shift in plant phenology caused by an earlier biological spring and a delayed winter; while again, these effects will vary according to water availability and regional characteristics (Peñuelas et al. 2009). The lengthened vegetation period might represent additional benefits for invasive species in the Mediterranean, since these are mostly species which flower late and their flowering period is terminated by oncoming winter. The end of flowering in late season is one of the significant differences between native and non-native species in general (Knapp and Kühn 2012). Similarly, growth and biomass accumulation in non-natives are terminated by the first occurring low temperatures and are not ended gradually by plant senescence, as in most native species. This should be especially important for annual species.
Thorough studies are needed to better predict whether any alien species that are currently regarded as non-invasive non-native species and are already present could benefit from global change and turn into invasive alien species. This is why every alien species must be taken into consideration. In this chapter we discuss whether global warming might favour the non-invasive alien Aster squamatus, which is already present and whether it could potentially become invasive.
2 Aster squamatus, a Non-invasive Alien in Slovenia
Aster squamatus (Sprengel) Hieron. is a hemicryptophyte originating from central South America, occupying habitats like salt marsh landscapes from Central Argentina (Cantero et al. 1998). Even though A. squamatus does not usually form large dense stands in introduced habitats, the species is distributed widely in many European countries, among them Italy (Pace and Tammaro 2001), Spain (Molina et al. 2004), France (Bassett 1980), Malta (Deidun 2010), Cyprus (Hand 2000), and Greece (Theocharopoulos et al. 2006).
The first record of A. squamatus in Slovenia was noted in 1973 (Wraber 1982), and the species has been constantly present since then, although not highly invasive (Kaligarič 1998; Glasnović 2006). This species has also been observed in urban areas (urbanophile species) like the old town centre of Izola. Even though it is not highly invasive, where present, its occurrence is disturbing, especially near the two most important coastal wetlands along the Slovenian coast: Sečovlje salina and Škocjan inlet, representing various Natura 2000 habitats. Sečovlje salina is a traditionally built salt-pan system where active salt-making is still partly practised. The salt-pan system includes natural salt pools (flooded and dried out), which are maintained exclusively by high tide, and salts flats which both offer habitats for halophile species, and the lower course of the Dragonja river with its river mouth and freshwater as well as brackish riparian habitats along the banks. Škocjan inlet was renatured some years ago and here, too, different Natura 2000 habitats developed. In some years A. squamatus was found to be very abundant, even exceeding the abundance of the native halophile vegetation (Glasnović and Fišer Pečnikar 2010). The occurrence of A. squamatus is especially dense within and around the port of Koper, possibly the source of the arrival of the species on the Slovenian coast.
Species-specific empirical data are important for individual-based modelling of future changes of biodiversity; in particular, data about dispersal and life history trade-offs might improve model realism (Botkin et al. 2007). Additionally, knowledge about preferred environmental conditions helps to improve niche-based models because it allows forecasting of the persistence and distribution of species (Botkin et al. 2007). Thus, knowledge of a species’ reproductive success is needed for better understanding of its invasive potential, while on the other hand, micro-scale habitat properties along with biotic interactions also influence trends in the richness and abundance observed in alien species (Aguiar et al. 2006). This is why we estimated the reproductive potential of A. squamatus by measuring seed production of individual plants in relation to plant height and soil properties, especially soil salinity and humidity. Additionally, we tested the seeds for their germination characteristics.
3 Determining the Reproductive Potential and Habitat Characteristics
In evaluating the reproductive potential of A. squamatus plants, we recognised five categories of plant height: 1 – below 50 cm; 2 – from 50 to 70 cm; 3 – from 70 to 110 cm; 4 – from 110 to 140 cm; and 5 – from 140 to 170 cm. The potential reproductive success of A. squamatus plants was estimated by counting the number of flower heads for plants belonging to each category, as well as seed number per single flower head. The data obtained were used in making a calculated estimation of the total seed number for an individual plant. Subsequently, we tested the seeds for their germination rate in laboratory conditions. We collected ripe seeds in the field in October 2011; these were dry-stored until the beginning of the experiment. Twenty seeds were allowed to germinate untreated on filter paper, watered with distilled water for 31 days. Observations were performed every 2 days, at which point germinated seeds were counted and removed.
Soil samples collected from plots where A. squamatus was absent (0) or present (1), were analysed for P, K, organic matter, C, total N, C:N content, Zn and pH. Additionally, we measured the salinity and humidity of the soil samples. Where present, the A. squamatus plants were categorised into different height classes (a – 50 cm or less, b – between 50 and 100 cm, and c – above 100 cm).
Mapping of A. squamatus occurrence was predominantly used to estimate its distribution, not its habitat preference. However, it was obvious that this species favours disturbed ruderalised habitats, including ruderalised grasslands with slight to moderate elevations in salinity level.
Results show high reproductive potential and low soil C:N ratio.
Aster squamatus plants show high reproductive potential, which increases with the plant’s height (Fig. 19.1). The relation between a plant’s height and its reproductive potential, measured through the number of flower heads, shows no significant differences between plants up to 110 cm tall; however, taller plants from the height categories 4 and 5 differ significantly from the first 3 categories and between each other (ANOVA, F(4,69) = 69.07, P < 0.001, post-hoc Unequal N HSD; Fig. 19.2). The average number of seeds (achenes) in a single flower head seems to be fixed (31 and 35 seeds in categories 1 and 2, respectively), since the number increases only slightly with plant height (38 seeds in categories 3–4).

Relation between plant height and number of flower heads of Aster squamatus (equation of logarithmic fit is given)

Reproductive potential of A. squamatus plants from different height categories (1: <50 cm; 2: 50–70 cm; 3: 70–110 cm; 4: 110–140 cm; 5: 140–170 cm) measured through the number of flower heads per single plant (significant differences are marked with different letters; ANOVA, post-hoc Unequal N HSD)
Different height categories (a–c) did not differ among each other in any of the soil chemical properties tested (ANOVA, at P < 0.05). However, there is statistically significant difference (t-test, P < 0.05) in phosphorus content (t-value = 2.1; d.f. = 28; P = 0.045) and C:N ratio (t-value = −3.77; d.f. = 28; P = 0.0008) between the soil samples taken from sites where A. squamatus is present and these where it is absent, irrespective of plant height (Table 19.1). Additionally, differences in total N (t-value = 1.78; d.f. = 28; P = 0.086) and zinc content (t-value = 1.72; d.f. = 28; P = 0.098) were close to significance.
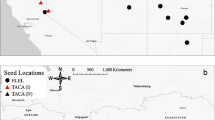
Until now, the occurrence of A. squamatus has been restricted by the low number of available habitats. In Slovenian coastal habitats A. squamatus was found restricted to semi-saline habitats, which are represented only by a narrow strip near the coast between the typical halophile vegetation and coastal grasslands or arable land farther from the sea. The habitat types where A. squamatus thrives belong to halophytic scrubs (1420), halophilous reeds and rush salt marshes dominated by Juncus maritimus (1410; Fig. 19.3).

Locations where A. squamatus is present on the Slovenian sea coast (a). Two protected areas are shown in detail: Sečovlje Salina (b) and Škocjan Inlet (c)
4 As an Engineering Species Aster squamatus Could Potentially Become Invasive
Since the first record of A. squamatus in Slovenia, the species has had a constant presence. Its abundance and distribution is limited by habitat availability, which is scarce along the Slovenian coast, since coastal grasslands are poorly represented on the Slovenian coast because of considerable human impact and because they belong to low-productive grasslands. Such specific habitat properties do not enable A. squamatus to become abundant or highly invasive, even though the species has an enormous seed production and fairly high germination rates. However, these low-productive habitats could be altered by species’ decomposing biomass, as discussed later. On the other hand, in several European countries, coastal grasslands are common habitats and could potentially be invaded by A. squamatus. It has been observed before that A. squamatus, sometimes together with Conyza canadensis – another alien species, invades vegetation with coarse perennial grasses and sedges or rushes (Bassett 1980). Along with such invasions, it was mainly the abundance of small annuals that declined: for example, Catapodium rigidum and Parapholis incurva. We made similar observations in Slovenian habitats for Spergularia marina and Parapholis strigosa, rare plants from semi-saline habitats. If in any way the preferred semi-saline habitats become more common, A. squamatus could rapidly invade favourable habitats. Our results show that A. squamatus has some of the characteristics that have been recognised by statistical models from Knapp and Kühn (2012) to be significant for non-native species. One of them is a high level of seed production, which may be explained by non-native species being more frequently able to self-pollinate (Knapp and Kühn 2012).
The study site is located in the Sub-Mediterranean part of Slovenia, and between 1951 and 2010 the temperature has risen and precipitation shifted to autumn. There are more sunny days between May and August; all these consequences could be attributed to global warming (Anonymous 2006). There is already a slight increase in autumn temperatures, and predictions for the years 2036–2065 by various models (C4I, ETHZ, KNMI, MPI, SMHI_BCM, SMHI_HadCM3Q3, DMI_ECHAM5, DMI) based on climate data from 1971 to 2000 do show an increase in mean temperature especially in the middle of the vegetation season (the ENSEMBLES dataset and the HABIT-CHANGE-database 2012). Even though models predict a slight increase in precipitation (about 20 mm/day), the number of dry days (<1 mm/day) per vegetation season is expected to increase by 4–5 days. When combining these models the predicted beginning of the vegetation period will shift from 22nd February (1971–2000) to 27th January (2036–2065). Additionally, the predicted water balance, calculated as the mean precipitation minus the potential evapotranspiration, is expected to decrease slightly in autumn.
Global-warming-related effects on biodiversity can be mitigated in the short term by designing an appropriate nature reserve (Botkin et al. 2007). However, the challenges of biodiversity loss are daunting, since biodiversity is decreasing even in protected areas. For this reason, alien invasive species found in the Mediterranean, even if not highly invasive, like A. squamatus, might nevertheless potentially become so. In Fig. 19.3 we can recognise the distribution of A. squamatus in areas adjacent to both protected areas of Sečovlje Salina and Škocjanski Inlet. As previously noted, in some years like in 2008 the abundance of A. squamatus increased (Glasnović and Fišer Pečnikar 2010). Our analysis of temperature showed a highly increased mean annual temperature the previous year 2007 (Fig. 19.4), which resulted in a prolonged vegetation period and must have resulted in high seed production as well.

Mean annual temperature from 1998 until 2012 at the meteorological station in Portorož (Graph constructed from data provided by Slovenian Environmental Agency freely available at http://meteo.arso.gov.si/met/sl/app/webmet/)
To prevent a potential invasion by A. squamatus plants should be removed before reaching the height of 110 cm to prevent or at least minimise seed production, since smaller plants have more than five times fewer flower heads (Fig. 19.2), even though the number of seeds in a single flower head is more or less the same. Calculation show that plants smaller than 110 cm could bear from about 700 to 8,700 seeds, while plants taller than 110 cm produce at least five times more seeds from about 47,000 to 70,000 seeds. Obviously, it is necessary to constrain eventual accumulation of seeds in the soil seed bank and to prevent plants from forming a reservoir for future invasions if appropriate conditions might recur.
On the other hand, where A. squamatus plants are present, the soil C:N ratio is significantly lower (Table 19.1), indicating high biomass production and high quality litter, which enables faster decomposition and nutrient cycle rates. This is consistent with other invasive species, especially if they are capable of N-fixing (Williams and Baruch 2000). It is generally accepted that many invasive species benefit from high levels of nutrients (Schumacher et al. 2008 and references therein). We can regard A. squamatus as an engineering species, fertilising its own habitat. We can expect that the longer the species would be present in the habitat, the more the nitrogen content in the soil would increase and further promote A. squamatus growth. Climate change, prolonging the vegetation period, would enable more biomass accumulation followed by rapid decomposition. These nutrients from decomposed biomass can be better used by fast-growing species that start their development slightly later in the season than early spring species, which often begin to grow by utilising nutrients accumulated in their storage organs. In such cases of engineering alien species, adapted management is needed. Plants should be removed from occupied habitats irrespective of height, so that biomass accumulation cannot promote their own growth. This also includes dead plants from the previous season. However, this is not always feasible, so again plants should not be allowed to attain 1 m in height. Additionally, fertilising urban and ruderal sites could also promote invasions by alien species.
References
Aguiar, F. C., Ferreira, M. T., & Albuquerque, A. (2006). Patterns of exotic and native plant species richness and cover along a semi-arid Iberian river and across its floodplain. Plant Ecology, 184, 189–202.
Alpert, P., Bone, E., & Holzapfel, C. (2000). Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants. Perspectives in Plant Ecology, Evolution and Systematics, 3, 52–66.
Anonymous. (2006). Podnebne razmere v Sloveniji (Obdobje 1971–2000). Ljubljana: Slovenian Environmental Agency. Retrieved July 6, 2012, from http://meteo.arso.gov.si/uploads/probase/www/climate/text/sl/publications/podnebne_razmere_v_sloveniji_71_00.pdf
Bassett, P. A. (1980). Effects of grazing on vegetation dynamics in the Camargue, France. Vegetatio, 43, 173–184.
Botkin, D., Saxe, H., Araújo, M. B., Betts, R., Bradshaw, R., Cedhagen, T., Chesson, P., Davis, M. B., Dawson, T., Etterson, J., Faith, D. P., Guisan, A., Ferrier, S., Hansen, A. S., Hilbert, D., Kareiva, P., Margules, C., New, M., Skov, F., Sobel, M. J., & Stockwell, D. (2007). Forecasting effects of global warming on biodiversity. Bioscience, 57, 227–236.
Cantero, J. J., Cisneros, J. M., Zobel, M., & Cantero, A. (1998). Environmental relationships of vegetation patterns in saltmarshes of central Argentina. Folia Geobotanica, 33, 133–145.
Deidun, A. (2010). Challenges to the conservation of biodiversity on small islands: The case of the Maltese islands. International Journal of Arts and Sciences, 3, 175–187.
Glasnović, P. (2006). Flora slovenskega dela Miljskega polotoka (kvadranta 0448/1 in 0448/2). Dissertation, University of Ljubljana, Ljubljana.
Glasnović, P., & Fišer Pečnikar, Ž. (2010). Akebia quinata (Houtt.) Dcne., new species for Slovenian flora, and contribution to the knowledge of the neophytic flora of Primorska region. Hladnikia, 25, 31–44.
Hand, R. (2000). Contributions to the flora of Cyprus I. Willdenowia, 30, 53–65.
Kaligarič, M. (1998). Botanični pogled na možne ureditve naravnega rezervata Škocjanski zatok. Annales, Series Historia Naturalis, 13, 131–142.
Kerr, R. A. (2007). Global warming is changing the world. Science, 316, 187–190.
Knapp, S., & Kühn, I. (2012). Origin matters: Widely distributed native and non-native species benefit from different functional traits. Ecology Letters, 15, 696–703.
Milchunas, D. G., & Lauenroth, W. K. (1995). Inertia in plant community structure: State changes after cessation of nutrient enrichment stress. Ecological Applications, 5, 452–458.
Molina, J. A., Pertíñez, C., Díez, A., & Casermeiro, M. A. (2004). Vegetation composition and zonation of a Mediterranean braided river floodplain. Belgian Journal of Botany, 137, 140–154.
Nernberg, D., & Dale, M. R. T. (1997). Competition of five native prairie grasses with Bromus inermis under three moisture regimes. Canadian Journal of Botany, 75, 2140–2145.
Pace, L., & Tammaro, F. (2001). The main invasive alien plants in the protected areas in central Italy (Abruzzo). In G. Visconti, M. Benistom, E. D. Iannorelli, & D. Barba (Eds.), Advances in global change research series global change and protected areas (pp. 495–504). Dordrecht: Kluwer.
Peñuelas, J., Rutishauser, T., & Filella, I. (2009). Phenology feedbacks on climate change. Science, 324, 887–888.
Richardson, D. M., & Pyšek, P. (2004). What is an invasive species? CABI invasive species compendium. Retrieved July 6, 2012, from http://www.cabicompendium.org/cpc/aspects.asp?
Schumacher, E., Kueffer, C., Tobler, M., Gmür, V., Edwards, P. J., & Dietz, H. (2008). Influence of drought and shade on seedling growth of native and invasive trees in the Seychelles. Biotropica, 40, 543–549.
Stohlgren, T. J., Otsuki, Y., Villa, C., Lee, M., & Belnap, J. (2001). Patterns of plant invasions: A case example in native species hotspots and rare habitats. Biological Invasions, 3, 37–50.
Theocharopoulos, M., Georgiadis, T., Dimitrellos, G., Chochliouros, S., & Tiniakou, A. (2006). Vegetation types with Cladium mariscus (Cyperaceae) in Greece. Wilidenowia, 36, 247–256.
Williams, D. G., & Baruch, Z. (2000). African grass invasion in the Americas: Ecosystem consequences and the role of ecophysiology. Biological Invasions, 2, 123–140.
Williams, D. G., & Black, R. A. (1994). Drought response of a native and introduced Hawaiian grass. Oecologia, 97, 512–519.
Wraber, T. (1982). Aster squamatus (Sprengel) Hieron. tudi v Sloveniji (Prispevek k poznavanju adventivne flore Koprskega). Biološki vestnik, 30, 125–136.
Acknowledgements
The authors would like to thank Maja Dragšič for her help in germination studies and Daniel Veselič for helping with plant sampling. The ENSEMBLES data used in this work was funded by the EU FP6 Integrated Project ENSEMBLES (Contract number 505539) whose support is gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is distributed under the terms of the Creative Commons Attribution Noncommercial License, which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Copyright information
© 2014 The Author(s)
About this chapter
Cite this chapter
Šajna, N., Kaligarič, M., Ivajnšič, D. (2014). Reproduction Biology of an Alien Invasive Plant: A Case of Drought-Tolerant Aster squamatus on the Northern Adriatic Seacoast, Slovenia. In: Rannow, S., Neubert, M. (eds) Managing Protected Areas in Central and Eastern Europe Under Climate Change. Advances in Global Change Research, vol 58. Springer, Dordrecht. https://doi.org/10.1007/978-94-007-7960-0_19
Download citation
DOI: https://doi.org/10.1007/978-94-007-7960-0_19
Published:
Publisher Name: Springer, Dordrecht
Print ISBN: 978-94-007-7959-4
Online ISBN: 978-94-007-7960-0
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)