
Abstract
The calculation of fruit growth rate (FGR) is the main part of fruit-bearing crop growth models. In this paper, greenhouse tomato was used as a research material. The purpose of this study is to better understand the regulation of single fruit growth response to environmental or genetic factors. A FGR model of tomato fruit was described based on Beta function law because it has flexible ability to generate different curve shape when adjusting its parameters. A field experiments with 4 planting densities in 2012 spring was carried out in China, using the cultivar Weichi. The parameters of Beta-law FGR model were estimated with data from the fruits in the first truss by using the optimization algorithm Nelder-Mead The results showed that the optimization procedure described in this paper found best fitted and robust parameters, which implied that those parameters are cultivar specific, and little environmental dependence. The validation against the data from the second truss and third truss with variations in planting density was found to be acceptable (R2 = 0.9117). This modeling methodology and software program have the potential to become a powerful tool for optimization of ideotype design.
You have full access to this open access chapter, Download conference paper PDF
Similar content being viewed by others

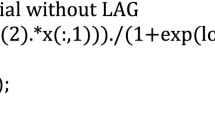
Keywords
1 Introduction
Tomato (solanum lycopersicum) is one of the most important horticultural crops and food preference in China. With the recent development of greenhouse technology, China has become the first tomato production country in the world as far as total amount of yields, but researches still indicated that the actual yield of the tomato in China is only 7.7 % of the average potential productivity, and there is still a larger space to improve the potential production [1].
Crop growth modeling provides a powerful research tool for studying the potential production. [2–5] Calculation of tomato fruit growth rate (FGR) is an important part of tomato simulation models, and the integration of FGR over time gives the accumulated fresh weight along different growth stages. It not only can be used to describe the potential performance in any given environment, but also to describe graded size distribution of final yields. Two kinds of models describing fruit dynamics can be distinguished (1) the empirical models and (2) physiological models. Empirical or regression models are built up based on direct field and experimental observation with some regression functions, for instance the typical S-shaped logistic curves [5].
Because they are descriptive and the parameters of regression models are possible to varies with different level of observation without botanical meaning. Physiological models, on the other hand, explain the observed growth rates from the underlying physiological process and in relation to the environmental factors. So the parameters in such models are usually divided into two categories: cultivar-specific parameters and environment-dependent parameters, where cultivar-specific parameters are assumed constant for some kinds of cultivar and will be estimated and evaluated through optimization technology [6].
Physiological models simulate physiological process by using mathematical equations with different hierarchical level. For instance, in famous tomato growth model of Tomsim and Tomgro, both simulate tomato plant process based on compartment level, and the realization of fruit dynamics depend upon the partitioning of the photosynthesis production into fruit growth phases according to sink strength ratio among compartments, so the final yields is the main simulation objective, while individual fruit growth rate is seldom considered [7, 8].
Tomato functional-structural model (TFSM) provides detailed hierarchical structure and it entails quantitative integration of mechanism at organ level to provide a better modeling frame explaining the behavior related to environment-dependent or genetic-control phenotypic plasticity. However, because of the complexity of the model and the data sets used to estimate model parameter values were limited, the prediction of fruit kinetics was not accurate enough, thus limiting the application into the real production [9, 10].
In practice, management requires well-parameterized predictive models. To accomplish this, a tailored fruit dynamic model from TFSM model for simulating FGR in tomato was presented, which objective is to determine and evaluate those parameters whether they are affected by environment conditions or not. If not, those parameters will be introduced into the physiological models as known invariable cultivar-specific parameters so that the number of parameters to be estimated and the complexity of optimization procedure will be reduced.
2 Materials and Methods
2.1 Field Experiments
An experiment was conducted during spring 2012 (sowing date, 18 January) on tomato (Solanum lycopersicum, WeiChi) at the Xiaotangshan Vegetables Production Zone, Beijing, China (39.55 N, 116.25E). Plants were planted in a multi-span greenhouse. Rock wool were used as soil-free cultivation medium and four density treatments were applied: high density (HD, 4.2 plants m−2), intermediate density (MD1, 3.5 plants m−2 and MD2, 2.8 plants m−2) and low density (LD; 2.1 plants m−2). Irrigation and fertilization were similar for all density treatments. Air temperature, relative humidity and CO2 were collected every ten minutes by AR5-type data logger. The instrument AV-19Q was used to measure photosynthetic photon flux density.
The first flowering and fruit-set date were recorded. For each treatment, there were three replications and in each replication block, 4 plants were sampled to calculate the average diameter of tomato fruit. The leaf appearance rate was recorded shortly after planting date. After fruit thinning, 8 measurement date were carried out to evaluate the influence of varying density on fruit growth rate (FGR).
2.2 Model Description
The FGR model is executed at time steps corresponding to organogenetic growth cycles (GC), equal to the thermal time (degree days, °Cd) needed to generate a new metamer (or a leaf). According to the measurements and analysis, we found that there is a strong linear relationship between number of leaf and accumulated thermal time (Fig. 1).

Method of calculating the growth cycles (GCs) according to the relationship between accumulated thermal time and number of leaves
According to Fig. 1, the definition of one GC was derived as follows:
where \( GC \) is the definition of one growth cycle (about 24.78 degree days); \( GC \) is accumulated thermal time ((degree days, °Cd) which base temperature is 10°C. \( N \) is the total number of leaves along main stem.
With the definition of time step \( GC \), we formulate a dynamic model for tomato fruit growth rate as the following form:
where \( GC_{i} \) means \( i^{th} \) growth cycle; \( \Delta D \) is the real incremental size(mm) at the \( GC_{i} \);
\( P \) is unit cycle expansion strength(mm/GC) determining the potential maximum incremental size for each cycle; \( f_{c} \left( \ldots \right) \) will determine the variation shape of growth rate and is assumed as inner law only determined by cultivar type or gene control but not affected by environmental conditions; Here we apply Beta function to model the normalized growth rate for tomato fruit along \( GCx \) because it can generate flexible curve shape when adjusting its parameters.
where \( x = (1,2, \ldots T) \) and T is maximum expansion cycles and can be observed from field experiments; \( (p,q) \) are the parameters of Beta function to be estimated.
2.3 Parameter Calibration and Evaluation
To obtain the parameters of Beta law, a reliable calibration procedure was required. We firstly defined a criteria to evaluate the performance of the FGR model where non-linear least square sum was used as the objective function [14].
Where \( y_{i} \) is the observation (\( i = 1,2, \ldots N \) = 1); \( f(x_{i} ;\theta ) \) is the output of model; \( \theta \) is the parameter vector to be estimated which final calibrated value will minimize this objective function. To implement this, the optimization algorithm Nelder-Mead was introduced into model calibration procedure to find stable parameters. Finally, The test of calibration results against new data set was the root mean square error (RMSE), calculated as [11].
where \( p \) is predicted data, \( o \) is observation data and \( n \) is the number of observations. The smaller the \( RMSE \), the better the prediction.
2.4 Statistical Analysis
The data were analyzed using R package ‘stats’ (version 3.0.2). R Core Team (2013). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL http://www.R-project.org/.
3 Results and Discussion
In this section, the FGR model’s ability based on beta-law to simulate the expansion rate of tomato fruit was evaluated in two steps. First, the measurements on the fruits’ dynamics of the first truss were used for calibration. Second, a validation with parameterization was made using the observations from the fruits in the second and third truss along the main stem.
Those data for calibration were collected every five days from the date of fruit-set (20 Feb. 2012) to the maturity date (11 Apr. 2012). To convert the calendar date to growth cycles which are required in the model, we first compute the accumulated thermal time from fruit-set for each observation date, then use the relationship described in Fig. 1 to calculated the passing growth cycles. So the incremental diameter on each measurement will be divided by number of growth cycles and the calculated value was considered as the average growth rate on the observation date. To capture the essence of the tomato fruit’s characteristics and to investigate if the model parameters are environment-dependent or not, the FGR model’s parameters were estimated in each condition against target files composed of 4 planting density (LD,MD1,MD2,HD).
The estimated curves for four densities were shown in Fig. 2. Those curves have similar shape and a good agreement between observation and model results was obtained for MD1,MD2,HD (a RMSE of 25 % and below was defined as acceptable agreement), while the RMSE for LD is a little bigger, the possible reason for which is that the date of the first fruit set should be earlier than the others, but in real measurement and calibration procedure, it is recorded the same as the other density. But anyway, we can concluded that the Beta law could be possible to model the growth rate of tomato fruits’ dynamics, because it can basically track the variation of tomato fruit’s kinetics.

Fruit growth rate of the first truss as a function growth cycles after fruit-set: measurement verse fitted results with Beta-law FGR model.
Model parameters were determined for each growing condition by statistical optimization. Table 1 presents the optimized parameter values and their respective standard deviation for each situation. Coefficient of variation (CV%) shows a higher stability of model parameters, and it indicates that increasing densities had little influence on the parameters defining the kinetics of organ fruit (beta law parameters.
Figure 3 shows the iterations process of the optimism algorithm, i.e. Nelder-Mead applied in the non-linear least square calibration procedure. The curve indicated the good performance because the optimization algorithm converged very quickly and found the best fitted values of parameters to minimize the objective function, and it successfully found global minimum while avoiding local one. So this optimization algorithm is suitable for crop growth models.

Convergence paths of the Nelder-Mead optimization algorithm applied to the FGR model.
We made an average on parameters’ values for four density and the determined model was validated by the fruits dynamics in the second truss and third truss. The simulation results against the observation were shown in Fig. 4. The simulation of tomato fruit’s growth rate generally described the observed field data and the regression of observed data on predicted ones indicated a one to one correspondence (R2 = 0.91). But we still can observe from Fig. 4 that some points are over-predicted or largely under-predicted. One of the possible reasons for inaccurate prediction is that observation is the average diameter of total truss fruits, while in the calibration set, the observation is made from regular date and a single fruit, another reason is that a different nutrient solution was applied in the medium-later growth stages for the second truss and third truss from the one applied in the first truss. However, despite some inaccuracies, the model still provided a reasonable simulation of fruit expansion patterns.

The validation of model against the data from the second and third truss
4 Conclusion
Form the parameterization of the tomato growth rate model and results’ analysis, it is concluded that the FGR beta-law model by using the concepts of growth cycles is able to accurately simulate the expansion rate of greenhouse tomato’s fruit. The optimized parameters of model are similar for different planting densities which further strengthened the assumption that those parameters are cultivar- specific and not affected by external environment conditions. The modeling methodology described in this paper will find applications on yield and quality prediction and gene-based ideotype optimization design.
References
Chu, J.X., Sun, Z.F., Du, K.M., Jia, Q.: Spatial distribution of potential productivity for greenhouse tomato variety Zhongza No. 9 Based on TOMSIM. China J. Agrometeorol. 30(1), 49–53 (2009). (in Chinese)
Pogson, M., Hastings, A., Smith, P.: Sensitivity of crop model predictions to entire meteorological and soil input datasets highlights vulnerability to drought. Environ. Model Softw. 29(1), 37–43 (2012)
Luquet, D., Rebolledo, M.C., Soulie, J.C.: Functional-structural plant modeling to support complex trait phenotyping: case of rice early vigour and drought tolerance using ecomeristem model. In: Proceedings - 2012 IEEE 4th International Symposium on Plant Growth Modeling, Simulation, Visualization and Applications, PMA 2012, pp. 270–277 (2012)
Soltani, A., Hoogenboom, G.: Assessing crop management options with crop simulation models based on generated weather data. Field Crops Res. 103, 198–207 (2007)
van Ittersum, M.K., Leffelaar, P.A., van Keulen, H., Kropff, M.J., Bastiaansb, L., Goudriaana, J.: On approaches and applications of the Wageningen crop models. Eur. J. Agron. 18, 201–234 (2003)
He, C., Zhang, Z.: Modeling the relationship between tomato fruit growth and the effective accumulated temperature in solar greenhouse. Acta Hortic. 718, 581–588 (2006)
Acutis, M., Confalonieri, R.: Optimization algorithm for calibrating cropping systems simulation models. A case study with simplex-derived methods integrated in the warm simulation environment Acutis M.e Confalonieri R. Ital. J. Agrometeorol. 3, 26–34 (2006)
Jones, J.W., Dayan, E., Allen, L.H., et al.: A dynamic tomato growth and yield model (TOMGRO). Trans. ASAE 34, 663–672 (1991)
Heuvelink, E.: Evaluation of a dynamic simulation model for tomato crop growth and development. Ann. Bot. 83, 413–422 (1999)
Dingkuhn, M., Luquet, D., Quilot, B., de Reffye, P.: Environmental and genetic control of morphogenesis in crops: towards models simulating phenotypic plasticity. Aust. J. Agric. Res. 56, 1289–1302 (2005)
Dong, Q.X., Louarn, G., Wang, Y.M., Barczi, J.F., de Reffye, P.: Does the structure-function model GREENLAB deal with crop phenotypic plasticity induced by plant spacing? A case study on tomato. Ann. Bot. 101, 1195–1206 (2008)
Janssen, P.H.M., Heuberge, P.S.C.: Calibration of process- oriented models. Ecol. Model. 83, 55–56 (1995)
Ma, L.L., Ji, J., He, C.: Studies on tomato leaf area index measurement method based on BP neural network. China Veg. 16, 45–50 (2009)
Wallach, D., Makowski, D., Jones, J.W.: Working with Dynamic Crop Models: Evaluations, Analysis, Parameterization, and Applications. Elsevier, Amsterdam (2006)
ACKNOWLEDGMENTS
This study was supported by the National Natural Science Foundation of China (#61174088 and #31200543), and Special Found for Beijing Common Construction Project. The authors would also like to thank students from College of Agronomy, China Agricultural University involved in the plant measurements.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2016 IFIP International Federation for Information Processing
About this paper
Cite this paper
Dong, Q., Yang, L., Qu, M., Shi, Q., Du, S. (2016). Beta Function Law to Model the Dynamics of Fruit’s Growth Rate in Tomato: Parameter Estimation and Evaluation. In: Li, D., Li, Z. (eds) Computer and Computing Technologies in Agriculture IX. CCTA 2015. IFIP Advances in Information and Communication Technology, vol 478. Springer, Cham. https://doi.org/10.1007/978-3-319-48357-3_3
Download citation
DOI: https://doi.org/10.1007/978-3-319-48357-3_3
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-48356-6
Online ISBN: 978-3-319-48357-3
eBook Packages: Computer ScienceComputer Science (R0)