
Abstract
There are many positive attributes to DNA vaccination that make it a conceptually desirable platform. In clinical studies, however, standard DNA injection alone generally induces low levels of transgene-specific immunity when compared to other vaccine approaches. In order to boost the immunogenicity of this platform, next-generation DNA vaccines require additional techniques such as the administration of electroporation. This new method involves the generation of a brief electric field in tissue around a local injection site that results in the transient poration, or permeabilization, of the cellular membranes. As a result, antigen-specific immune responses are greatly enhanced and are likely due to increased DNA uptake and antigen expression. Thus, electroporation-mediated DNA vaccination represents a promising new strategy for the elicitation of strong immune responses directed against the expressed antigen(s) and not the vector, and ongoing studies are currently underway to optimize the working parameters of this technique. Here, we review the uses of this technology in conjunction with vaccination and suggest future directions for its further exploration.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others

Keywords
Introduction
The discouraging results from the recent human HIV trial performed by Merck and collaborators, named “STEP,” raises serious questions about current vaccine approaches using replication-defective viral vectors [1]. This study was an international phase II “test of concept” trial in uninfected volunteers at high risk for acquiring HIV infection which showed that vaccination using a recombinant adenovirus serotype 5 vector was ineffective at preventing virus infection and even increased the rate of transmission in individuals exhibiting prior immunity to the viral vector. While this vaccine was safe and immunogenic in both humans and nonhuman primates, eliciting long-lasting and multifunctional CD8+ T cell responses that were partially protective in rhesus macaques, the discovery that the vaccine could possibly heighten HIV infection was both unexpected and alarming, resulting in the immediate discontinuation of vaccinations as recommended by the independent Data Safety Monitoring Board for STEP. Since preexisting immunity against a vaccine vector may compromise its efficacy in humans, future vaccine approaches should aim to utilize vectors that exhibit minimal or no reactivity in immunocompetent vaccines.
DNA vaccination consists of the administration of genetic material encoding a desired antigen that when expressed in the vaccine, is capable of eliciting an immune response. Compared with other approaches, the advantages of DNA vaccination are many [2]; no infectious agents are involved and vaccines are unable to revert into virulent form like live vaccines. They can prime both cytotoxic [3] and humoral responses [4], and DNA vectors are easily manipulated, can be tested rapidly, produced at high yield in bacteria, and are readily isolated. They are also more temperature-stable than conventional vaccines, easily stored and transported, and do not require a cold chain. Furthermore, DNA vaccines could potentially induce immunity in newborns in situations that usually neutralize conventional vaccines via the presence of high levels of maternal antibodies [5]. The introduction of exogenous DNA into cells or tissue can be achieved using DNA conjugates [6–10], virus-derived vectors [11], or naked plasmid DNA (pDNA) [12]. Naked pDNA shows variable and low transfection efficacy when administered by conventional means, such as needle injection or topical application. However, several strategies aimed to improve pDNA vaccine immunogenicity have been developed, including codon optimization [13], mRNA optimization [14], addition of leader sequences [15], and construction of consensus immunogens [16]. While these strategies help to boost the overall immunogenicity of a DNA vaccine, they may not be applicable to all antigens. Recently, electroporation, or electropermeabilization, has gained great interest in multiple research areas including gene therapy and vaccinology [17]. Although the precise mechanism of action has not yet been well defined, it is hypothesized that cell membranes in host tissue receiving electroporation, normally impermeable to charged molecules, form pores or functionally equivalent structural changes upon application of an external electric field which facilitate the influx of macromolecules. Thus, higher transfection efficacy of naked pDNA as a result of electroporation is thought to be the major contributor to the increased immunogenicity of electroporation-mediated DNA vaccination. In addition, it has been shown that electroporation increases vaccine potency by activating antigen-presenting cells (APCs) via danger signals and local inflammation [18] and by recruiting immune cells to the site of DNA administration [19, 20]. Furthermore, direct transfection of APCs could also be facilitated by electroporation. Currently, intensive investigation is focused upon utilizing electroporation of muscle and skin as an effective method for DNA vaccine delivery to small and large animals, and in humans. The safety and feasibility of electroporation in humans has recently been demonstrated, but not finally proven [21]. Thus, the paramount question for DNA vaccines at this time is whether a sufficient level of efficacy can be reached with the present methodology, or if further improvements or breakthroughs in vaccine design and/or electroporation delivery will be necessary.
In Vivo Transgene Expression After Gene Electrotransfer
In 1990, the first transgene expression detected in skeletal muscle after injection of naked mRNA or pDNA raised the possibility of using this method for certain gene therapies and DNA vaccinations [22]. Subsequently, transgene expression was also obtained in the same way in a wide variety of other tissues, but transgene expression was generally too low and variable to be useful for the envisioned purposes [12]. Attempts to sufficiently enhance pDNA uptake, and thus transgene expression, with cationic lipids or the gene gun have also proven unsuccessful to date. The first publications on a substantial increase in transgene expression (about 100-fold) when electroporation was applied in vivo after pDNA injection appeared as late as 1996 [23], although electroporation had been used for in vitro cell transfections since 1982 [24]. In addition, as pointed out by Bettan et al. [25], when using gene electrotransfer, a higher interindividual reproducibility in gene transduction can be observed.
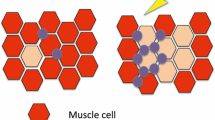
Skeletal muscle (Fig. 18.1) has been the most frequently targeted tissue in both gene therapy and DNA vaccine studies, either with or without electroporation. Some reasons why muscle cells (also known as myocytes) and muscle tissues continue to be attractive targets for transgene expression include: muscle tissue is easily accessible, plentiful, and well vascularized; the latter facilitates circulation of the antigens produced by the transfected muscle cells. More discussion of gene electrotransfer to muscle can be found in Chap. 16.

Cellular targets of electroporation-mediated DNA vaccination. Current electroporation approaches following i.d. and i.m. DNA vaccination target skin cells (keratinocytes), muscle cells, and APCs (including Langerhans cells) by direct transfection (DT). Application of electroporation at a local immunization site induces the formation of transient complexes between the DNA and the lipids in the pore edges of the hydrophilic pore zones [62] which facilitate the translocation of pDNA into the cell cytoplasm. Once in the cytosol, pDNA may gain access to the nucleus where transgene expression may occur
Electroporation and Plasmid DNA Vaccine Immunogenicity
In vivo electroporation has been used to deliver DNA vaccine encoding antigens from numerous infectious agents, summarized in Table 18.1. Enhanced immune responses to electroporation-mediated DNA vaccination have been observed both in small and large animals such as mice [26], pigs [27], and monkeys [28]. Widera et al. [26] demonstrated in mice that upon electroporative treatment, the delivery of a weakly immunogenic hepatitis B virus (HBV) surface antigen (Hbs Ag) DNA vaccine resulted in an increased humoral immune response, characterized by rapid onset and higher titers of anti-Hbs Ag antibodies. In addition, the authors observed in the same study that the potency of an HIV gag pDNA vaccine was increased as shown by the lower dosage of DNA required to induce higher antigen-specific antibody levels and increased CD8+ T cell responses. Similarly, in a study carried out with a bovine herpes virus-1 truncated glycoprotein D DNA vaccine, Tsang et al. [29] showed that the onset of the primary humoral response was earlier in the group treated with DNA followed by electroporation, and that this group produced higher antibody levels than those in the group receiving i.m. DNA immunization or a recombinant protein vaccine only; similar results were obtained earlier with an otherwise inert microparticulate adjuvant [30]. Interestingly, the efficiency of transfection by electroporation was not increased by doubling the dose of DNA administered; however, the duration of the antigen-specific antibody response was increased at a higher rate in comparison to the immunization with the same dose of plasmid without electroporation. Moreover, electroporation increased the degree of consistency among the individuals in the DNA-plus-electroporation group as seen in the 5 weeks of follow-up. Finally, a high correlation between the duration of the primary immune response and the magnitude of the secondary antibody response was observed, implying that electroporation could represent an effective approach to elicit a longer memory antibody response.
Capone et al. [31] have demonstrated that gene electrotransfer efficiently increased the cellular immune response both in mice and rhesus macaques vaccinated with a plasmid encoding a nonstructural region of hepatitis C virus (HCV). In particular, they showed by ex vivo interferon (IFN)-γ ELISPOT assay that electroporation in mice induced a fivefold more potent T cell response than DNA administration alone, and that the elicited response was directed against all six of the antigen pools spanning the HCV NS3-NS5B region. To assess whether electroporation treatment elicited similar responses in a nonhuman primate model, they immunized rhesus macaques three times with the vaccine and collected peripheral blood mononuclear cells at periodic intervals to test the T cell effector function. The immune responses observed in the electroporation-treated group showed a faster kinetic, with all the animals responding after the second challenge and reaching a peak after the third. Moreover, all animals treated with electroporation showed both CD4+ and CD8+ T cell responses, in comparison to the naked DNA group which showed a weaker CD8+ response and no CD4+ response. Finally, gene electrotransfer-immunized macaques maintained anti-HCV effector T cells for the entire observation period of 6 months, indicating that the gene electrotransfer efficiently elicited a strong memory T cell response.
DNA vaccination in association with electroporation represents an effective strategy to elicit strong, broad, and long-lasting B and T cell responses. Although muscle is the most common target for DNA vaccine immunizations [32], the presence of APCs in both the skin layers makes it an attractive target for nucleic acid vaccination, since direct transfection of APCs may be important for T cell priming upon skin DNA immunization [33] (Fig. 18.2). In a murine model using a viral challenge, Raz et al. [34] have demonstrated that a single intradermal (i.d.) injection (without electroporation) of naked DNA encoding the influenza nucleoprotein gene is sufficient to induce production of antigen-specific antibodies and cytotoxic T lymphocytes that persist for at least 68 weeks and are protective against a lethal challenge with a heterologous strain of influenza virus. Furthermore, immune responses to i.d. DNA vaccination have been recorded to be significantly enhanced by in vivo electroporation [35]; analysis of the antibody response to an Hbs Ag-encoding plasmid delivered i.d. upon electroporation in mice has revealed a strong enhancement of the Th1 response, which is mainly characterized by a strong cell-mediated response, compared to that elicited by protein immunization, which showed an exclusively Th2 pattern, characterized by a dominant humoral responses. Also, in a nonhuman primate model study carried out in rhesus macaques [36], the i.d.-plus-electroporation group developed 50% more IFN-γ-producing cells and twice more memory T cells than the group not treated with electroporation. Higher antibody responses were recorded in the i.d.-plus-electroporation group when compared to the i.m.-plus-electroporation group. Altogether, these results support the idea that electroporation following DNA injection, both in muscle and skin, represents an effective approach to large animal immunization.

Cellular targets of EP-mediated DNA vaccination. Current EP approaches following i.d. and i.m. DNA vaccination target skin cells (keratinocytes), muscle cells, and APC (including Langerhans cells) by direct transfection (DT). Application of EP at a local immunization site induces the formation of transient complexes between the DNA and the lipids in the pore edges of the hydrophilic pore zones [51] which facilitate the translocation of plasmid DNA (pDNA) into the cell cytoplasm. Once in the cytosol, pDNA may gain access to the nucleus where transgene expression may occur
Electroporation-Mediated Local Inflammation
Several authors have hypothesized that inflammation caused by electroporation is important to prime the immune response to DNA vaccination [19, 37, 38]. Local inflammation was previously proposed to augment immune responses in studies where pDNA was coinjected with bupivicaine-HCl [39–41]. The localized tissue damage induced by the electric field is thought to recruit CD3+ cells, increasing the number of infiltrating immune cells at the injection site [42]. Indeed, electroporation caused the activation of proinflammatory signals including the expression of chemokines such as MIP-1α, MIP-1β, MIP-1γ, IP-10, MCP-2, and XCL1 [18]. Liu et al. [20] characterized the extent and nature of the cellular infiltrates at the site of electroporative vaccine delivery in mice and found both polymorphonuclear and mononuclear cells localized in the perivascular spaces and throughout the muscle tissue. In particular, they observed a significant increase in B cells, CD4+ and CD8+ T cells, and a dramatic increase in macrophages and dendritic cells compared to vaccination alone. No difference, however, was recorded among cell populations of blood, spleen, and draining lymph nodes of the mice treated with or without electroporation, suggesting that only local factors are involved in the augmentation of immune responses following electroporation. Also, these authors observed that cell infiltrates were transient and resolved within 2 weeks. Thus, improved antigen presentation may represent one of the mechanisms by which electroporation may elicit a more potent immune response.
Typically, innate immune recognition of the adjuvant component of vaccine formulations has been shown to be critical for their immunogenicity [43]. Many adjuvants are ligands for Toll-like receptors (TLRs), like monophosphoryl lipid A and CpG DNA [44, 45], while some conventional adjuvants, such as aluminum hydroxide and incomplete Freud’s adjuvant are free of TLR ligands [46]. Therefore, these examples demonstrate that multiple innate immune recognition and signaling pathways are required for adjuvants to function [44]. In the case of DNA vaccines, it has been controversial as to the main vaccine component contributing most to the induction of both innate and adaptive immune responses; while CpG motifs expressed within the plasmid backbone can stimulate innate immunity through TLR 9, the induction of adaptive immune responses were unaffected in the absence of this innate receptor [47, 48]. However, it has recently been shown that the double-stranded structure of DNA, independently of CpG sequences, possesses immunomodulatory effects when administered intracellularly [44], which can trigger TLR-independent, TANK-binding kinase I (TBK1)- and INF regulatory factor 3-dependent innate activation of both immune and nonimmune cells to produce type I INFs and their inducible genes [49, 50]. Recently, Ishii et al. [51] have reported that the enhancement of DNA vaccine immunogenicity achieved by electroporation may be due to increased transfection rates resulting from this technique, which better contributes to local inflammation by activating cells to produce IFN through the TBK1-dependent signaling pathway. These data suggest that TBK1 is a key signaling molecule for DNA vaccination immunogenicity by regulating innate immune signaling, which is critical for the induction of adaptive immune responses, and that the enhanced immunogenicity of pDNA by electroporation may be a result of more pDNA interacting with intracellular TBK1.
In accordance with this hypothesis, Peng et al. [18] postulated that local inflammation is more important than the actual quantity of expressed transgene in determining the magnitude of the immune response, demonstrated by higher antibody titers and CD4+ T cell proliferation rates observed by applying electric pulses 3–7 days prior to i.m. DNA immunization. In this case, it can be postulated that both increased cross-presentation and direct transfection of infiltrating APCs resulting from increased local inflammation may contribute to the augmented immune response in electroporation-mediated DNA vaccination. It appears that the mechanisms by which electroporation enhance the responses to naked plasmid vaccination is by an increase in DNA transfection and possibly include local inflammation, which may be augmented by the magnitude or duration of transgene expression. Indeed, Babiuk et al. [19] observed that the highest level of lymphocytic infiltration was only in muscle tissue treated with electroporation, which elicited higher levels of transgene expression, as was expected considering that antigen production is critical for the retention of the cellular infiltrates at sites of local inflammation.
Electroporation-Mediated DNA Vaccination in Humans
Although two DNA vaccines have been recently approved in the USA and Canada for the vaccination of horses against West Nile virus [52] and salmon against infectious hematopoietic necrosis virus [53], no DNA vaccine has been approved for use in humans. However, encouraging results from preclinical trials using electroporation technology with DNA vaccination in large animal models has prompted much interest in the technique and its safety. Currently, tolerability in humans has been demonstrated in healthy volunteers [54], anti-DNA antibodies have not been detected in patients electroporated after muscle DNA injection, and the integration of pDNA into host chromosomes following electroporation-mediated delivery has not been observed [55]. Together, these results have been sufficient for the regulatory approval of several clinical trials [56]. As reported on clinicaltrials.gov, seven electrotransfer DNA vaccine trials for cancer and three clinical trials using DNA vaccine against infectious agents in association with electroporation are currently open. Ongoing clinical studies using electroporation-mediated DNA vaccination against infectious agents include three phase I studies involving muscle electroporation. The first will test safety and immunological effects of PENNVAX™-B, an HIV vaccine encoding Gag, Pol, and Env, in HIV-infected individuals (VGX Pharmaceuticals, Inc.); the second will assess safety, tolerability, and immunogenicity of human papillomavirus (HPV) DNA Plasmid (VGX-3100™) delivered by electroporation in adult females postsurgical or ablative treatment of Grade 2 or 3 Cervical Intraepithelial Neoplasia (VGX Pharmaceuticals, Inc.); the third one, a Phase I/II trial is testing tolerability and efficacy of i.m.-administered CHRONVAC-C™ in combination with electroporation in chronic HCV genotype 1 infected and naïve patients with low viral load (Tripep AB).
The amount of pain and distress associated with electroporation in humans has been of a tolerable level for the anticipated benefit [57]. To date, electroporation-mediated DNA vaccination in humans is performed administrating an injection volume of 0.2–0.5 mL followed by short (10–60 ms), low electric field strength (60–250 V/cm) pulses (2–10 pulses). Given that these conditions are efficient for the DNA vaccination of large animals, such as nonhuman primates, they should be sufficient in humans. Electroporation results in a sharp, but quick pain that is comparable to receiving a short electrical shock. While this sensation is transient, administration of short-acting sedative drugs or painkillers before treatment has been considered. Accordingly, as reported by Daud et al. [57], in a clinical trial using an interleukin-12-encoding plasmid delivered by electroporation in patients with metastatic melanoma, in order to limit patients’ discomfort, lidocaine was either administered topically or injected around each tumor site, and intravenous analgesic and/or anxiolytic drugs were offered to the patients before electroporation. Notably, previous studies have shown that pain is not a limiting factor as patient discomfort is limited to the period of electrical stimulation, and subjects have usually returned for repeated treatments without asking for sedation. Also, after muscle electroporation, like muscle injection, a mild ache may be experienced at the site of electroporation for some days, and similar to that following a strenuous workout. Several factors determine the strength of pain associated with electroporation, although there is a high interindividual variability in the perception of pain. Among these factors are the number, length, spacing, and thickness of the electrode needles, but primarily the electric pulse parameters dictate the pain threshold [58].
Exclusion criteria for electroporation treatment may include the presence of metal implants near the site of electrical delivery, the presence of a pacemaker, and in the case of muscle electroporation, obesity, since treatment efficacy may be decreased if muscle tissue is not reached for vaccine and/or electric pulse delivery.
Electrical Parameters and Electroporation Equipment
Electroporation-mediated DNA vaccine delivery requires a pulse generator that controls the parameters of individual pulses or pulse trains (amplitude, duration, number, polarity, wave form, frequency), and electrodes usually integrated into an applicator. Electrodes are in direct contact with the subject to be treated and it is their geometry (shape, size, and distance from each other) that ultimately determines the shape and strength of the electric field and the electrical currents in the target tissue (Fig. 18.3). Thus, both the properties of the pulse(s) and the electrodes are responsible for the desired enhancement of DNA delivery as well as undesirable side effects. Proper design of pulses and electrodes will maximize the effectiveness of a given DNA vaccine and minimize unwanted side effects, such as long-lasting histological changes, pain, and muscle contractions. Further discussion of electroporation parameters and equipment can be found in Chaps. 4 and 5.

Main types of electrodes used for EP. Examples of the two main types of electrodes used for delivering EP: Surface (a) and needle-type (b) electrodes. Surface electrodes come in the form of caliper electrodes or consist of an array of interweaving electrode fingers (meander). Needle-type electrodes consist of a variable number of (primarily) stainless steel needles arranged in different configurations. Picture depicts a 5-needle array firing rotating chord patterns, a 4-needle array fired in opposing pairs, crossing or triangular patterns, and a 2-needle array firing pin to pin
Skin Electroporation and Equipment
Skin is a potentially interesting target tissue for DNA vaccines because of its natural role in the immune defense of the body and its ready accessibility. Discussion of gene electrotransfer of DNA in general and to skin can be found in Chaps. 13 and 17, respectively.
Conclusion
The goal of any vaccination strategy is to prime a broad and long-lasting immune response that is capable of robust effector responses upon antigenic restimulation [59–61] and protect against infectious agents, while minimizing toxicity of the vaccine. Since complications may arise due to the use of viral-based vectors, such as preexisting immunity regulating vaccine effectiveness and the possibility of reversion into virulence that may decrease vaccine efficacy or safety, alternative approaches should be explored for future vaccine approaches. DNA vaccination appears an even more attractive candidate for future vaccines since it is safe, immunogenic, and does not stimulate vector-specific immunity. Extensive literature supports the hypothesis that electroporation represents a valid approach to vaccine administration in that it increases the consistency and potency of vaccination, inducing higher levels of both antibody-mediated and cytotoxic T cell responses. In this way, electroporation may help augment translocation of exogenous DNA into the nucleus. Furthermore, it has been proven to be effective in enhancing immune responses to antigens regardless of the degree of transgene expression achieved. Recent evidence suggests that innate immune recognition of the adjuvant component of vaccines and the danger signals provided by the method of vaccination may be important in determining the magnitude of the resultant immune response. Electroporation may enhance local inflammation at the site of immunization by facilitating the transfection of greater amounts of pDNA, which may be more readily available to interact with intracellular signaling proteins that trigger the secretion of inflammatory cytokines and their inducible genes. While electroporation-mediated DNA vaccination is currently the topic of intensive research, more theoretical studies and practical trials are required to optimize the delivery of the vaccine into the target tissue, and the electrical parameters facilitating DNA uptake while calibrating local tissue damage and reducing pain associated with vaccine delivery through electroporation.
Abbreviations
- Hbs Ag:
-
Hepatitis B surface antigen
- HBV:
-
Hepatitis B virus
- HCV:
-
Hepatitis C virus
- IFN:
-
Type I interferons
- pDNA:
-
Plasmid DNA
- TBK1:
-
TANK-binding kinase I
- TLR:
-
Toll-like receptors
References
Sekaly RP. The failed HIV Merck vaccine study: a step back or a launching point for future vaccine development? J Exp Med. 2008;205:7–12.
Shedlock DJ, Weiner DB. DNA vaccination: antigen presentation and the induction of immunity. J Leukoc Biol. 2000;68:793–806.
Ulmer JB, Donnelly JJ, Parker SE, Rhodes GH, Felgner PL, Dwarki VJ, et al. Heterologous protection against influenza by injection of DNA encoding a viral protein. Science (New York, NY). 1993;259:1745–9.
Tang DC, DeVit M, Johnston SA. Genetic immunization is a simple method for eliciting an immune response. Nature. 1992;356:152–4.
Chen J, Zhang F, Fang F, Chang H, Chen Z. Vaccination with hemagglutinin or neuraminidase DNA protects BALB/c mice against influenza virus infection in presence of maternal antibody. BMC Infect Dis. 2007;7:118.
Nicolau C, Le Pape A, Soriano P, Fargette F, Juhel MF. In vivo expression of rat insulin after intravenous administration of the liposome-entrapped gene for rat insulin I. Proc Natl Acad Sci USA. 1983;80:1068–72.
Kaneda Y, Iwai K, Uchida T. Increased expression of DNA cointroduced with nuclear protein in adult rat liver. Science (New York, NY). 1989;243:375–8.
Mannino RJ, Gould-Fogerite S. Liposome mediated gene transfer. Biotechniques. 1988;6:682–90.
Benvenisty N, Reshef L. Direct introduction of genes into rats and expression of the genes. Proc Natl Acad Sci USA. 1986;83:9551–5.
Wu GY, Wu CH. Receptor-mediated gene delivery and expression in vivo. J Biol Chem. 1988;263:14621–4.
Schoenly KA, Weiner DB. Human immunodeficiency virus type 1 vaccine development: recent advances in the cytotoxic T-lymphocyte platform “spotty business”. J Virol. 2008;82:3166–80.
Wolff JA, Budker V. The mechanism of naked DNA uptake and expression. Adv Genet. 2005;54:3–20.
Deml L, Bojak A, Steck S, Graf M, Wild J, Schirmbeck R, et al. Multiple effects of codon usage optimization on expression and immunogenicity of DNA candidate vaccines encoding the human immunodeficiency virus type 1 Gag protein. J Virol. 2001;75:10991–1001.
Schneider R, Campbell M, Nasioulas G, Felber BK, Pavlakis GN. Inactivation of the human immunodeficiency virus type 1 inhibitory elements allows Rev-independent expression of Gag and Gag/protease and particle formation. J Virol. 1997;71:4892–903.
Yang JS, Kim JJ, Hwang D, Choo AY, Dang K, Maguire H, et al. Induction of potent Th1-type immune responses from a novel DNA vaccine for West Nile virus New York isolate (WNV-NY1999). J Infect Dis. 2001;184:809–16.
Gao F, Weaver EA, Lu Z, Li Y, Liao HX, Ma B, et al. Antigenicity and immunogenicity of a synthetic human immunodeficiency virus type 1 group m consensus envelope glycoprotein. J Virol. 2005;79:1154–63.
Wells DJ. Gene therapy progress and prospects: electroporation and other physical methods. Gene Ther. 2004;11:1363–9.
Peng B, Zhao Y, Xu L, Xu Y. Electric pulses applied prior to intramuscular DNA vaccination greatly improve the vaccine immunogenicity. Vaccine. 2007;25:2064–73.
Babiuk S, Baca-Estrada ME, Foldvari M, Middleton DM, Rabussay D, Widera G, et al. Increased gene expression and inflammatory cell infiltration caused by electroporation are both important for improving the efficacy of DNA vaccines. J Biotechnol. 2004;110:1–10.
Liu J, Kjeken R, Mathiesen I, Barouch DH. Recruitment of antigen-presenting cells to the site of inoculation and augmentation of HIV-1 DNA vaccine immunogenicity by in vivo electroporation. J Virol. 2008;82:5643–9.
Gehl J. Electroporation for drug and gene delivery in the clinic: doctors go electric. Methods Mol Biol. 2008;423:351–9.
Wolff JA, Malone RW, Williams P, Chong W, Acsadi G, Jani A, et al. Direct gene transfer into mouse muscle in vivo. Science (New York, NY). 1990;247:1465–8.
Heller R, Jaroszeski M, Atkin A, Moradpour D, Gilbert R, Wands J, et al. In vivo gene electroinjection and expression in rat liver. FEBS Lett. 1996;389:225–8.
Neumann E, Schaefer-Ridder M, Wang Y, Hofschneider PH. Gene transfer into mouse lyoma cells by electroporation in high electric fields. EMBO J. 1982;1:841–5.
Bettan M, Emmanuel F, Darteil R, Caillaud JM, Soubrier F, Delaere P, et al. High-level protein secretion into blood circulation after electric pulse-mediated gene transfer into skeletal muscle. Mol Ther. 2000;2:204–10.
Widera G, Austin M, Rabussay D, Goldbeck C, Barnett SW, Chen M, et al. Increased DNA vaccine delivery and immunogenicity by electroporation in vivo. J Immunol. 2000;164:4635–40.
Babiuk S, Baca-Estrada ME, Foldvari M, Storms M, Rabussay D, Widera G, et al. Electroporation improves the efficacy of DNA vaccines in large animals. Vaccine. 2002;20:3399–408.
Zhao YG, Peng B, Deng H, Chen G, Yang F, Shao M, et al. Anti-HBV immune responses in rhesus macaques elicited by electroporation mediated DNA vaccination. Vaccine. 2006;24:897–903.
Tsang C, Babiuk S, van Drunen Littel-van den Hurk S, Babiuk LA, Griebel P. A single DNA immunization in combination with electroporation prolongs the primary immune response and maintains immune memory for six months. Vaccine. 2007;25:5485–94.
Zhang L, Widera G, Rabussay D. Enhancement of the effectiveness of electroporation-augmented cutaneous DNA vaccination by a particulate adjuvant. Bioelectrochemistry (Amsterdam, Netherlands). 2004;63:369–73.
Capone S, Zampaglione I, Vitelli A, Pezzanera M, Kierstead L, Burns J, et al. Modulation of the immune response induced by gene electrotransfer of a hepatitis C virus DNA vaccine in nonhuman primates. J Immunol. 2006;177:7462–71.
Danko I, Williams P, Herweijer H, Zhang G, Latendresse JS, Bock I, et al. High expression of naked plasmid DNA in muscles of young rodents. Hum Mol Genet. 1997;6:1435–43.
Porgador A, Irvine KR, Iwasaki A, Barber BH, Restifo NP, Germain RN. Predominant role for directly transfected dendritic cells in antigen presentation to CD8+ T cells after gene gun immunization. J Exp Med. 1998;188:1075–82.
Raz E, Carson DA, Parker SE, Parr TB, Abai AM, Aichinger G, et al. Intradermal gene immunization: the possible role of DNA uptake in the induction of cellular immunity to viruses. Proc Natl Acad Sci USA. 1994;91:9519–23.
Drabick JJ, Glasspool-Malone J, King A, Malone RW. Cutaneous transfection and immune responses to intradermal nucleic acid vaccination are significantly enhanced by in vivo electropermeabilization. Mol Ther. 2001;3:249–55.
Hirao LA, Wu L, Khan AS, Hokey DA, Yan J, Dai A, et al. Combined effects of IL-12 and electroporation enhances the potency of DNA vaccination in macaques. Vaccine. 2008;26:3112–20.
Gronevik E, Mathiesen I, Lomo T. Early events of electroporation-mediated intramuscular DNA vaccination potentiate Th1-directed immune responses. J Gene Med. 2005;7:1246–54.
McMahon JM, Wells KE, Bamfo JE, Cartwright MA, Wells DJ. Inflammatory responses following direct injection of plasmid DNA into skeletal muscle. Gene Ther. 1998;5:1283–90.
Caselli E, Betti M, Grossi MP, Balboni PG, Rossi C, Boarini C, et al. DNA immunization with HIV-1 tat mutated in the trans activation domain induces humoral and cellular immune responses against wild-type Tat. J Immunol. 1999;162:5631–8.
Chattergoon MA, Robinson TM, Boyer JD, Weiner DB. Specific immune induction following DNA-based immunization through in vivo transfection and activation of macrophages/antigen-presenting cells. J Immunol. 1998;160:5707–18.
Coney L, Wang B, Ugen KE, Boyer J, McCallus D, Srikantan V, et al. Facilitated DNA inoculation induces anti-HIV-1 immunity in vivo. Vaccine. 1994;12:1545–50.
Ahlen G, Soderholm J, Tjelle T, Kjeken R, Frelin L, Hoglund U, et al. In vivo electroporation enhances the immunogenicity of hepatitis C virus nonstructural 3/4A DNA by increased local DNA uptake, protein expression, inflammation, and infiltration of CD3+ T cells. J Immunol. 2007;179:4741–53.
Pulendran B, Ahmed R. Translating innate immunity into immunological memory: implications for vaccine development. Cell. 2006;124:849–63.
Kawai T, Akira S. Signaling to NF-kappaB by Toll-like receptors. Trends Mol Med. 2007;13:460–9.
Mata-Haro V, Cekic C, Martin M, Chilton PM, Casella CR, Mitchell TC. The vaccine adjuvant monophosphoryl lipid A as a TRIF-biased agonist of TLR4. Science (New York, NY). 2007;316:1628–32.
Gavin AL, Hoebe K, Duong B, Ota T, Martin C, Beutler B, et al. Adjuvant-enhanced antibody responses in the absence of toll-like receptor signaling. Science (New York, NY). 2006;314:1936–8.
Babiuk S, Mookherjee N, Pontarollo R, Griebel P, van Drunen Littel-van den Hurk S, Hecker R, et al. TLR9−/− and TLR9+/+ mice display similar immune responses to a DNA vaccine. Immunology. 2004;113:114–20.
Spies B, Hochrein H, Vabulas M, Huster K, Busch DH, Schmitz F, et al. Vaccination with plasmid DNA activates dendritic cells via Toll-like receptor 9 (TLR9) but functions in TLR9-deficient mice. J Immunol. 2003;171:5908–12.
Ishii KJ, Coban C, Kato H, Takahashi K, Torii Y, Takeshita F, et al. A Toll-like receptor-independent antiviral response induced by double-stranded B-form DNA. Nat Immunol. 2006;7:40–8.
Stetson DB, Medzhitov R. Recognition of cytosolic DNA activates an IRF3-dependent innate immune response. Immunity. 2006;24:93–103.
Ishii KJ, Kawagoe T, Koyama S, Matsui K, Kumar H, Kawai T, et al. TANK-binding kinase-1 delineates innate and adaptive immune responses to DNA vaccines. Nature. 2008;451:725–9.
Minke JM, Siger L, Karaca K, Austgen L, Gordy P, Bowen R, et al. Recombinant canarypoxvirus vaccine carrying the prM/E genes of West Nile virus protects horses against a West Nile virus-mosquito challenge. Arch Virol Suppl. 2004;18:221–30.
Garver KA, LaPatra SE, Kurath G. Efficacy of an infectious hematopoietic necrosis (IHN) virus DNA vaccine in Chinook Oncorhynchus tshawytscha and sockeye O. nerka salmon. Dis Aquat Organ. 2005;64:13–22.
Rune K, Torunn ET, Dag K, and Iacob M. Clinical Evaluation of Pain and Muscle Damage Induced by Electroporation of Skeletal Muscle in Humans Abstracts from the American Society of Gene Therapy 7th Annual Meeting. June 2–6, 2004, Minneapolis, Minnesota, USA. Mol Ther. 2004;9 Suppl 1:S1–435.
Strain AJ. The uptake and fate of exogenous cellular DNA in mammalian cells. Dev Biol (Basel). 2006;123:23–8. discussion 55–73.
ClinicalTrials.gov. U.S. National Institutes of Health; 2008.
Daud AI, DeConti RC, Andrews S, Urbas P, Riker AI, Sondak VK, et al. Phase I trial of interleukin-12 plasmid electroporation in patients with metastatic melanoma. J Clin Oncol. 2008;26:5896–903.
Rabussay D. Applicator and electrode design for in vivo DNA delivery by electroporation. Methods Mol Biol. 2008;423:35–59.
Janssen EM, Lemmens EE, Wolfe T, Christen U, von Herrath MG, Schoenberger SP. CD4+ T cells are required for secondary expansion and memory in CD8+ T lymphocytes. Nature. 2003;421:852–6.
Shedlock DJ, Shen H. Requirement for CD4 T cell help in generating functional CD8 T cell memory. Science (New York, NY). 2003;300:337–9.
Sun JC, Bevan MJ. Defective CD8 T cell memory following acute infection without CD4 T cell help. Science (New York, NY). 2003;300:339–42.
Hristova NI, Tsoneva I, Neumann E. Sphingosine-mediated electroporative DNA transfer through lipid bilayers. FEBS Lett. 1997;415:81–6.
Rosati M, Valentin A, Jalah R, Patel V, von Gegerfelt A, Bergamaschi C, et al. Increased immune responses in rhesus macaques by DNA vaccination combined with electroporation. Vaccine. 2008;26:5223–9.
Cristillo AD, Weiss D, Hudacik L, Restrepo S, Galmin L, Suschak J, et al. Persistent antibody and T cell responses induced by HIV-1 DNA vaccine delivered by electroporation. Biochem Biophys Res Commun. 2008;366:29–35.
Otten G, Schaefer M, Doe B, Liu H, Srivastava I, zur Megede J, et al. Enhancement of DNA vaccine potency in rhesus macaques by electroporation. Vaccine. 2004;22:2489–93.
zur Megede J, Sanders-Beer B, Silvera P, Golightly D, Bowlsbey A, Hebblewaite D, et al. A therapeutic SIV DNA vaccine elicits T-cell immune responses, but no sustained control of viremia in SIVmac239-infected rhesus macaques. AIDS Res Hum Retroviruses. 2008;24:1103–16.
Wang S, Zhang C, Zhang L, Li J, Huang Z, Lu S. The relative immunogenicity of DNA vaccines delivered by the intramuscular needle injection, electroporation and gene gun methods. Vaccine. 2008;26:2100–10.
Chen J, Fang F, Li X, Chang H, Chen Z. Protection against influenza virus infection in BALB/c mice immunized with a single dose of neuraminidase-expressing DNAs by electroporation. Vaccine. 2005;23:4322–8.
Kadowaki S, Chen Z, Asanuma H, Aizawa C, Kurata T, Tamura S. Protection against influenza virus infection in mice immunized by administration of hemagglutinin-expressing DNAs with electroporation. Vaccine. 2000;18:2779–88.
Qiu M, Fang F, Chen Y, Wang H, Chen Q, Chang H, et al. Protection against avian influenza H9N2 virus challenge by immunization with hemagglutinin- or neuraminidase-expressing DNA in BALB/c mice. Biochem Biophys Res Commun. 2006;343:1124–31.
Zhao YG, Xu Y. Electroporation-mediated HBV DNA vaccination in primate models. Methods Mol Biol. 2008;423:487–95.
Peng JL, Zhao YG, Mai JH, Pang WK, Guo W, Chen GM, et al. Non-cytolytic antigen clearance in DNA-vaccinated mice with electroporation. Acta Pharmacol Sin. 2007;28:1024–30.
Babiuk S, Tsang C, van Drunen Littel-van den Hurk S, Babiuk LA, Griebel PJ. A single HBsAg DNA vaccination in combination with electroporation elicits long-term antibody responses in sheep. Bioelectrochemistry (Amsterdam, Netherlands). 2007;70:269–74.
Luxembourg A, Hannaman D, Ellefsen B, Nakamura G, Bernard R. Enhancement of immune responses to an HBV DNA vaccine by electroporation. Vaccine. 2006;24:4490–3.
Medi BM, Hoselton S, Marepalli RB, Singh J. Skin targeted DNA vaccine delivery using electroporation in rabbits. I: efficacy. Int J Pharm. 2005;294:53–63.
Zucchelli S, Capone S, Fattori E, Folgori A, Di Marco A, Casimiro D, et al. Enhancing B- and T-cell immune response to a hepatitis C virus E2 DNA vaccine by intramuscular electrical gene transfer. J Virol. 2000;74:11598–607.
Dutta NK, Mazumdar K, Lee BH, Baek MW, Kim DJ, Na YR, et al. Search for potential target site of nucleocapsid gene for the design of an epitope-based SARS DNA vaccine. Immunol Lett. 2008;118:65–71.
He H, Tang Y, Qin X, Xu W, Wang Y, Liu X, et al. Construction of a eukaryotic expression plasmid encoding partial S gene fragments of the SARS-CoV and its potential utility as a DNA vaccine. DNA Cell Biol. 2005;24:516–20.
Hu H, Lu X, Tao L, Bai B, Zhang Z, Chen Y, et al. Induction of specific immune responses by severe acute respiratory syndrome coronavirus spike DNA vaccine with or without interleukin-2 immunization using different vaccination routes in mice. Clin Vaccine Immunol. 2007;14:894–901.
Shiau YT, Huang YH, Wu JC, Tao MH, Syu Jr W, Chang FY, et al. Analysis of humoral immunity of hepatitis D virus DNA vaccine generated in mice by using different dosage, gene gun immunization, and in vivo electroporation. J Chin Med Assoc. 2006;69:7–13.
Kim JY, Jeon ES, Lim BK, Kim SM, Chung SK, Kim JM, et al. Immunogenicity of a DNA vaccine for coxsackievirus B3 in mice: protective effects of capsid proteins against viral challenge. Vaccine. 2005;23:1672–9.
Wu CJ, Lee SC, Huang HW, Tao MH. In vivo electroporation of skeletal muscles increases the efficacy of Japanese encephalitis virus DNA vaccine. Vaccine. 2004;22:1457–64.
Kim SA, Liang CM, Cheng IC, Cheng YC, Chiao MT, Tseng CJ, et al. DNA vaccination against foot-and-mouth disease via electroporation: study of molecular approaches for enhancing VP1 antigenicity. J Gene Med. 2006;8:1182–91.
Yang NS, Wang JH, Lin KF, Wang CY, Kim SA, Yang YL, et al. Comparative studies of the capsid precursor polypeptide P1 and the capsid protein VP1 cDNA vectors for DNA vaccination against foot-and-mouth disease virus. J Gene Med. 2005;7:708–17.
Muthumani K, Lankaraman KM, Laddy DJ, Sundaram SG, Chung CW, Sako E, et al. Immunogenicity of novel consensus-based DNA vaccines against Chikungunya virus. Vaccine. 2008;26:5128–34.
Luxembourg A, Hannaman D, Nolan E, Ellefsen B, Nakamura G, Chau L, et al. Potentiation of an anthrax DNA vaccine with electroporation. Vaccine. 2008;26:5216–22.
Hooper JW, Golden JW, Ferro AM, King AD. Smallpox DNA vaccine delivered by novel skin electroporation device protects mice against intranasal poxvirus challenge. Vaccine. 2007;25:1814–23.
Zhang X, Divangahi M, Ngai P, Santosuosso M, Millar J, Zganiacz A, et al. Intramuscular immunization with a monogenic plasmid DNA tuberculosis vaccine: enhanced immunogenicity by electroporation and co-expression of GM-CSF transgene. Vaccine. 2007;25:1342–52.
Li Z, Song D, Zhang H, He W, Fan X, Zhang Y, et al. Improved humoral immunity against tuberculosis ESAT-6 antigen by chimeric DNA prime and protein boost strategy. DNA Cell Biol. 2006;25:25–30.
Tollefsen S, Tjelle T, Schneider J, Harboe M, Wiker H, Hewinson G, et al. Improved cellular and humoral immune responses against Mycobacterium tuberculosis antigens after intramuscular DNA immunisation combined with muscle electroporation. Vaccine. 2002;20:3370–8.
Saha S, Takeshita F, Sasaki S, Matsuda T, Tanaka T, Tozuka M, et al. Multivalent DNA vaccine protects mice against pulmonary infection caused by Pseudomonas aeruginosa. Vaccine. 2006;24:6240–9.
LeBlanc R, Vasquez Y, Hannaman D, Kumar N. Markedly enhanced immunogenicity of a Pfs25 DNA-based malaria transmission-blocking vaccine by in vivo electroporation. Vaccine. 2008;26:185–92.
Dobano C, Doolan DL. Identification of minimal CD8+ and CD4+ T cell epitopes in the Plasmodium yoelii hepatocyte erythrocyte protein 17kDa. Mol Immunol. 2007;44:3037–48.
Yen HH, Scheerlinck JP. Co-delivery of plasmid-encoded cytokines modulates the immune response to a DNA vaccine delivered by in vivo electroporation. Vaccine. 2007;25:2575–82.
Scheerlinck JP, Karlis J, Tjelle TE, Presidente PJ, Mathiesen I, Newton SE. In vivo electroporation improves immune responses to DNA vaccination in sheep. Vaccine. 2004;22:1820–5.
Acknowledgments
We would like to thank Karuppiah Muthumani and Shaheed Abdulhaqq for critical review of the manuscript and members of the DBW laboratory for helpful discussion.
Competing financial interests: The laboratory of D.B.W. has grant funding and collaborations, advising, or consulting including serving on scientific review committees for commercial entities and therefore notes possible conflicts associated with this work with Pfizer, Inovio, BMS, Virxsys, Ichor, Merck, Althea, VGXI, J&J, Aldevron, and possibly others. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed. No writing assistance was utilized in the production of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer Science+Business Media, LLC
About this chapter
Cite this chapter
Fagone, P., Shedlock, D.J., Kemmerer, S., Rabussay, D., Weiner, D.B. (2011). Electroporation-Mediated DNA Vaccination. In: Kee, S., Gehl, J., Lee, E. (eds) Clinical Aspects of Electroporation. Springer, New York, NY. https://doi.org/10.1007/978-1-4419-8363-3_18
Download citation
DOI: https://doi.org/10.1007/978-1-4419-8363-3_18
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4419-8362-6
Online ISBN: 978-1-4419-8363-3
eBook Packages: MedicineMedicine (R0)