
Abstract
Previous chapters have described a large number of transgenic animal models used to study specific cardiovascular syndromes. This chapter will fill in some gaps. Many of these transgenic animals were developed to study normal and/or abnormal physiological responses in other organ systems, or to study basic biochemical and molecular reactions or pathways. These models were then discovered to also have effects on the cardiovascular system, some of them unanticipated.
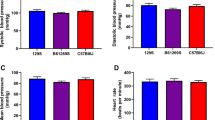
A word of caution, particularly when highly inbred mouse strains are used to develop transgenic models - not all strains of a particular species are created equal. When cardiovascular parameters of age- and sex-matched A/J and C57BL/6J inbred mice were compared the C57BL/6J mice demonstrated eccentric physiologic ventricular hypertrophy, increased ventricular function, lower heart rates, and increased exercise endurance.1
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- Transgenic Mouse
- Sarcoplasmic Reticulum
- Reverse Cholesterol Transport
- Regulatory Light Chain Phosphorylation
- BERKO Mouse
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
Previous chapters have described a large number of transgenic animal models used to study specific cardiovascular syndromes. This chapter will fill in some gaps. Many of these transgenic animals were developed to study normal and/or abnormal physiological responses in other organ systems, or to study basic biochemical and molecular reactions or pathways. These models were then discovered to also have effects on the cardiovascular system, some of them unanticipated.
A word of caution, particularly when highly inbred mouse strains are used to develop transgenic models - not all strains of a particular species are created equal. When cardiovascular parameters of age- and sex-matched A/J and C57BL/6J inbred mice were compared the C57BL/6J mice demonstrated eccentric physiologic ventricular hypertrophy, increased ventricular function, lower heart rates, and increased exercise endurance.1
It is also important to note that different species will react differently to the introduction of promoter genes. Human ICAM-2 promoter produces strong and uniform endothelial expression in all organs examined in mice. When the identical promoter was used to generate transgenic pigs only two of the pigs showed expression and that was significantly weaker than in the mice and restricted to vascular endothelium in the heart and kidney.2
Transgenic animals that harbor luciferase or other marker/reporter proteins have proven to be very useful when used to define in vivo responses to specific perturbations. The reporter protein in these transgenics is under the control of a specific promoter. A few examples of these transgenic animals include the development of CD2-enhanced green fluorescence protein transgenic mice used to characterize lymphocyte trafficking during inflammation.3 Placier et al.4 used transgenic mice harboring the luciferase reporter gene under the control of the collagen I-α-2 chain promoter to show that the combination of removing l-NAME and blocking endothelin in the l-NAME model of hypertension normalized collagen-I gene expression and reversed nephroangiosclerosis without marked changes in blood pressure. Pichler et al.5 describe a universal transgenic reporter mouse strain that expresses firefly luciferase under the regulatory control of a concatenated Gal-4 promoter. They use an adenovirus to deliver a fused binding-domain-activator chimera to induce bioluminescence in tissues that take up the Ga1-4 promoter. Beta-gal expression of the lacZ gene was used in two transgenic models (cGATA6/lacZ and cardiac conduction system/lacZ) and two targeted gene knock-in models (minK/lacZ and Hop/lacZ) in an attempt to mark portions of the proximal and distal atrioventricular conduction system (AVCS) in mice. Beta-gal expression was consistently observed in the proximal and distal AVCS but it was also observed in the working myocardium outside the AVCS. The reporter system was not expressed in the SA node, and thus it limits the technique.6
14.1 Sex-Related Responses
Studies using estrogen deletion,7 phytoestrogen feeding,8 genetic models, and estrogen receptor antagonists and agonists have demonstrated that both estrogen receptor subtypes (α and β) protect against various forms of cardiovascular disease including ischemia-reperfusion injury, hypertension, ventricular hypertrophy, and chronic heart failure.9 When low-density lipoprotein LDL−/− female mice were treated with estrogen (E2) there was a delay in atherosclerotic plaque formation. This delay was associated with an increase in DNA strand breaks in the arterial wall. When the same mouse model was fed a high-cholesterol diet and ovarioectomized there was an increase in atherogenesis.10
Aromatase knockout mice, both males and females, suffer from a variety of abnormalities including loss of fertility and libido, bone loss, cardiovascular and cerebrovascular problems, and development of metabolic syndrome.11 Treating arteries from male estrogen receptor-α knockout (ERKO) mice with specific aromatase short-interfering RNA for 72 h knocked down the aromatase mRNA and protein associated with elimination of epoxyeicosatrienoic acid (EET) mediation of flow-induced dilation. This indicates that flow-induced dilation in male ERKO mice arteries is maintained by an EDHF/EET-mediated mechanism compensating for reduced NO because of reduced estrogen aromatized from testosterone.12
We were able to demonstrate the important role of estrogen receptor-α in cardiac ischemia-reperfusion injury.13 Nuclear ER-α−/− (ERKO) mice were used to show that ER-α target genes involved in energy substrate oxidation, ATP synthesis, and phosphate transfer are downregulated at baseline and following pressure overload. The results indicate that ER-α is required for the adaptive bioenergetic response to pressure overload heart failure.14 During acute lipopolysaccharide (LPS)-induced inflammatory responses ER-α mediates NO synthesis and the vascular responses to NO involving soluble guanylate cyclase.15
ER-β−/− (BERKO) mice were used to show that ER-β-mediated mechanisms protect against left ventricular hypertrophy.16 E2 (estrogen) mitigates Angio-II signaling that produces hypertrophy and fibrosis in female BERKO mice.17 BERKO mice are protected against high-fat diet-induced insulin resistance and glucose intolerance. The protection involves augmented PPAR-γ signaling in adipose tissue and overexpression of nuclear coactivators such as SRC1 and TIF2.18 ER-β also plays a role in the cardioprotective effects of estrogen following global, warm ischemia-reperfusion injury.19 Myocardial infarction-induced heart failure in female BERKO mice resulted in increased mortality, increased levels of clinical and biochemical markers of heart failure, and impaired expression of Ca2+-handling proteins.20 Estrogenic effects on Ca2+ currents in isolated ventricular cardiomyocytes from WT, ERKO, and BERKO mice show that the inhibition of Ca2+ currents and decreased contractility are not dependent upon ER-α or β.21
ERKO and BERKO mice were used to reveal that ER-α- and ER-β-dependent pathways regulate distinct, and mostly nonoverlapping, sets of genes that function as an estrogen-regulatory transcriptional network in the vasculature.22 ER-β seems to modify adrenergic control of small artery tone in males but not in females. This is demonstrated by a gender-specific (male) enhanced response to phenylephrine23 and reduced endothelium-derived hyperpolarizing factor (EDHF)-mediated relaxation via gap junction communication.24 NO appears to contribute to the vasodilation response to E2 in femoral arteries from BERKO but not WT mice.25 ERs play a sex-specific role in small mesenteric arterial estrogen-mediated flow responses and distensibility. Deletion of ER-β affects arterial structure only in male BERKO.26
In vascular smooth muscle cells (VSMC) human-inducible nitric oxide synthase (iNOS) transcription is positively regulated by ER-β and negatively regulated by ER-α.27 Knocking out the gene encoding the catabolic enzyme CYP7B1 decreased estrogen-dependent expression of NOS and repressed carotid artery reendothelialization following endothelial injury.28
Female follitropin-receptor knockout (FORKO) mice have an impaired natriuretic peptide system. This could contribute to the susceptibility to develop age-related hypertension in female FORKO mice. This model shows a relationship between estrogen, adipose tissue, and ANP, and it is used as a model for menopause.29 Aged FORKO mice developed higher levels of Angio-II-induced hypertension, ventricular hypertrophy, and myocardial fibrosis than age-matched WT controls. The Angio-II-induced ventricular hypertrophy and myocardial fibrosis are associated with downregulation of the endogenous antioxidant thioredoxin (Trx) and upregulation of apoptosis-signal-regulated kinase-1 (ASK-1)/caspase signaling in the FORKO mice.30
14.2 Kinases
Transgenic mice overexpressing cardiac-specific Lats2, a serine/threonine kinase, have significantly reduced left and right ventricular mass whereas transgenic mice overexpressing dominant-negative Lats2 demonstrate biventricular hypertrophy.31 G-protein-coupled receptor (GPCR) kinase 2 (GRK-2) is also a serine/threonine kinase. It phosphorylates and desensitizes agonist-bound GPCRs. GRK-2 levels in VSMC are increased in animal models of hypertension. In GRK-2 knockout mice β-adrenergic receptor-mediated dilation was increased while α-adrenergic-mediated vasoconstriction was also increased.32 Confocal studies of cardiac tissue from transgenic mice overexpressing GRK-2 show a clear increase in relative fluorescence intensity of GRK-2 in the arrhythmia-prone subepicardial border zone following coronary artery ligation.33 Myocardial thrombin signaling, as assessed by p42/p44 mitogen-activated protein kinase activation, was significantly attenuated in transgenic mice with cardiac specific overexpression of GRK-3.34 Transgenic mice overexpressing cardiac-specific dominant-negative glycogen synthase kinase-β (GSK-3-β DN) were produced. Persistent inhibition of GSK-3-β induced compensatory hypertrophy, inhibited apoptosis and fibrosis, and increased cardiac contractility. The antiapoptosis effects of GSK-3-β inhibition are mediated by myeloid cell leukemia-1.35 When GSK-3-β was used to create a double-transgenic mouse by breeding with a mouse model of hypertrophic cardiomyopathy (HCM) the male animals had reduced contractility, reduced sarcoplasmic (endo) reticulum Ca2+-ATPase expression, elevated ANF expression, and premature death. Female HCM/GSK-3-β double-transgenic mice demonstrated cardiac histology, function, and survival the same as their female HCM littermates.36
Hypertension was induced using uninephrectomy and DOCA-salt in Axl (a receptor tyrosine kinase) knockout (Axl−/−) mice. The results indicate a likely mechanism for Axl-dependent effects on hypertension.37 Studies conducted in serum- and glucocorticoid-inducible kinase-1 knockout mice (SGK-1−/−) indicate that this kinase may play a decisive role in fetal programming of hypertension induced by prenatal protein restriction.38
Reactive oxygen species (ROS) generated during ischemic preconditioning could be responsible for altering mitochondrial metabolism by oxidizing key mitochondrial enzymes in the hearts of protein kinase C-delta knockout (PKC-delta−/−) mice. Metabolic adaptation to preconditioning is also impaired in this animal model.39 In transgenic mice that overexpress specific cardiomyocyte Ca2+/calmodulin-dependent myosin light chain kinase-increased regulatory light chain phosphorylation was not responsible for cardiac hypertrophy. It apparently inhibits hypertrophy by contributing to enhanced contractility and myocardial efficiency.40
The hearts of transgenic mice overexpressing the regulatory γ-subunit of AMP-activated protein kinase (PPKAG2) resist low-flow ischemic injury better than controls. This resistance was associated with increased consumption of glycogen but this was unrelated to AMP-activated protein kinase activation.41 The intimal thickness-related receptor (ITR) is expressed in rabbit aortic smooth muscle cells following injury. The sequence contains a motif common to the Rhodopsin-like GPCR superfamily. ITR seems to be a novel receptor that could play a role in vascular remodeling.42
Cardiac-specific transgenic mice with increased or decreased phosphoinositide 3-kinase (PI3K) activity were crossed with a transgenic mouse model of dilated cardiomyopathy (DCM). Exercise training and increased PI3K activity prolonged survival in the DCM model by 15-20%. Reduced PI3K activity shortened survival by about 50%.43
14.3 Oxidases and Oxygenases
Transgenic mice deficient in gp91phox, an NADPH oxidase subunit protein, designated gp91−/− mice were used to define mechanisms involved in DOCA-salt-induced hypertension. The findings indicated that vascular superoxide ion overproduction via gp91phox-containing NADPH oxidase is involved in the development of hypertension in this model.44 Using mice lacking the p47phox subunit of the NADPH oxidase (p47phox−/−) Salguero et al.45 show that in WT mice 2K1C hypertension was associated with a fourfold increase in endothelial progenitor cells (EPCs). In p47phox−/− mice there was a significantly less increase in arterial pressures and no change in EPCs.
Coronary ligation was performed in gp91phox−/− and WT mice. Mortality was significantly higher in the gp91phox−/− mice but the progression of left ventricular remodeling was not affected by phenotype. Systemic oxidative stress was not reduced in gp91phox−/− mice since there was a significant increase in lipid peroxides probably mediated by the observed increase of the NADPH subunit nox-1 in gp91phox−/− mice.46
Heme oxygenase is a cytoprotective enzyme that degrades heme to generate carbon monoxide, bilirubin, and iron. Two rodent models, Gunn rats (hyperbilirubinemic) and heme oxygenase-1 knock out (HO-1−/−) mice, were used to evaluate DOCA-salt hypertension. Uninephrectomized WT rats treated with DOCA-salt had significantly increased arterial pressures but the response was attenuated in Gunn rats. The response in the Gunn rats was not due to a greater induction of HO-1 in the vasculature or due to an increase in renal Na+ excretion. Using HO-1−/− mice Wiesel et al.47 reported that chronic deficiency of HO-1 does not alter basal blood pressure, but when the HO-1−/− mice were exposed to 1K1C there was more severe renovascular hypertension and cardiac hypertrophy, an increase in ischemic damage, and higher mortality rates. DOCA-salt induced HO-1 protein in HO-1+/+ mice but there was no increase in arterial pressures. The same DOCA-salt regimen in HO-1−/− mice produced significant increases in arterial pressures.48
14.4 Adenosine and Adrenergic Receptors
Adenosine receptors belong to a large family of GPCRs containing four receptor subtypes. In keeping with the physiological model of yin and yang (accelerator and brake), receptor subtypes designated A1R and A3R mediate inhibition of adenylyl cyclase and subtypes A2aR and A2bR mediate stimulation of adenylyl cyclase. All of the identified receptor subtypes have now been used to create knockout mouse models and some have been used in overexpression models. Various activities related to normal cardiovascular function and to CNS regulation of CV function have been identified using these transgenic models. Some responses are reduced and others enhanced depending upon the specific receptor that has been deleted or overexpressed.49 Some examples are as follows: in the mouse heart, A1R inhibits β-adrenergic-induced myocardial contractility whereas A2aR opposes the action of A1R and enhances myocardial contractility. During low-flow myocardial ischemia A2aR supports myocardial contractility.50 Mice with deletion of adenylyl cyclase type V (AC-V−/−) demonstrate enhanced basal left ventricular function than WT controls, but there is a reduced response to β-adrenergic stimulation.51
In the subcommissural bed nucleus of the stria terminalis norepinephrine (NE)-induced decreases in synaptic transmission are markedly reduced in α2-adrenergic receptor knockout (α2-AR−/−) mice.52 α2-ARs directly regulate the release of NE, but not dopamine, at the terminal level in the nucleus accumbens in mice. α2-AR−/− mice have been used to show that these receptors regulate dopamine indirectly via their effects on dopamine neurons in the ventral tegmental area. The mechanism(s) for this indirect regulation were yet to be described in 2004.53 In WT mice rilmenidine, moxonidine, and clonidine all produce dose-dependent decreases in blood pressure and heart rate. In D79N α2-AR−/− mice there were no responses to these agents indicating that α2-ARs are responsible for the cardiovascular effects of these drugs.54 α2c-AR overexpression increased the development of behavioral despair and associated cardiovascular responses associated with increased corticosterone levels following stress. α-2c−/− mice demonstrated attenuated corticosterone levels and cardiovascular responses when exposed to equal levels of stress.55
The effects of alterations in the level of myocardial β-adrenergic receptor kinase (β-ARK-1) were studied in β-ARK-1+/− (heterozygous) mice and a mouse model heterozygous for β-ARK-1 and transgenic for cardiac-specific overexpression of β-ARK-1 COOH-terminal inhibitor peptide (β-ARK-1+/−/β-ARK-1ct). These studies suggest that the level of β-ARK-1 activity can modulate contractile function. Even partial inhibition of β-ARK-1 activity enhances β-adrenergic receptor signaling and results in improved catecholamine responsiveness.56
Dopamine (D2)-receptor knockout (D2R−/−) mice were used to demonstrate that pituitary vascular endothelial growth factor (VEGF) expression is under dopaminergic control and may be critical for pituitary angiogenesis via paracrine actions in female D2R−/− mice.57 “Beta-less” mice are triple adrenoreceptor knockouts (β-1/β-2/β-3 AR−/−). In these animals the lipolytic response to NE and β-AR agonists is blunted, but a residual low-affinity lipolytic effect is observed in the presence of catecholamines and β-3-AR agonists but not β-1- or β-2-AR agonists. The residual lipolytic effect is attributed to an unknown GPCR with low affinity for catecholamines.58
14.5 Nitric Oxide Synthase
In eNOS−/− mice, chronically treated with l-NAME, blood pressure decreased while eNOS+/− and eNOS+/+ mice both developed hypertension.59 eNOS−/− mice also develop fasting hyperinsulinemia, hyperlipidemia, and decreased insulin-stimulated glucose uptake.60 Pulmonary endothelial cells were isolated from eNOS−/− mice and grown without E2. In control WT cells E2 significantly increased telomerase activity but in the eNOS−/− cells exogenous eNOS or an NO donor were needed to normalize the level of telomerase activity.61 Billon et al.62 used eNOS−/− mice to document that the presence, but not the enzymatic activity, of eNOS is necessary for estrogen signaling in the endothelium.
Transgenic mice that overexpress eNOS were used to discover that ventilator-induced lung injury could be reduced significantly by excess eNOS. The mechanism for this protection seems to be inhibition of the production of inflammatory chemokines and cytokines associated with neutrophilic infiltration.63 eNOS overexpression is also able to prevent the development of 2K1C renovascular hypertension in mice.64 At the University of Missouri-Columbia transgenic pigs carrying an endogenous eNOS gene driven by a Tie-2 promoter and tagged with a V5 His tag have been developed.65
14.6 Metabolic Syndrome
Obesity and obesity-linked insulin resistance are significant causes of the metabolic syndrome characterized by adipocyte hypertrophy. Adipose tissue is an important regulator of energy homeostasis. It functions as an endocrine organ secreting adipokines including adiponectin (APN). Studies using adiponectin transgenic (overexpression) and knockout mice demonstrate that adiponectin is an insulin-sensitizing adipokine and that obesity can result in insulin resistance and diabetes. At least two different adiponectin receptors have been cloned (AdipoR1 and ApipoR2) comprising a novel cell-surface receptor family acting as receptors for globular and full-length adiponectin. These receptors mediate increased activity of AMP-activated protein kinase, and PPAR-α, as well as glucose uptake and fatty-acid oxidation by adiponectin.66,67 Intraglomerular macrophage infiltration and mRNA levels of VCAM-1, MCP-1, TNF-α, TGF-β, collagen type I/III, and NADPH oxidase are all increased in adiponectin knockout (APN−/−) mice than in WT, when subjected to subtotal nephrectomy and the resulting hypertension. Changes in the APN−/− mice resulted in glomerular and tubulointerstitial injury from increased inflammation and oxidative stress.68 Studies conducted on transgenic mice overexpressing native full-length APN, targeted to white adipose tissue, revealed that adipose mass and adipocyte size were reduced while caloric intake was maintained constant. APN−/− mice also show decreases in preadipocyte factor-1 mRNA and increased CCAAT/enhancer binding protein-α in white adipose tissue.69 Shinmura et al.70 compared adiponectin antisense transgenic mice with WT to demonstrate that the cardioprotective effects of short-term caloric restriction are mediated by increased production of adiponectin and the resulting activation of AMP-activated protein kinase.
The insulin receptor knockout (IR−/−) mouse is a genetic model of resistance to insulin’s indirect effects on hepatic glucose production. This model demonstrates high levels of adiponectin in the blood with normal adiponectin receptor levels and was used to provide evidence that adiponectin potentiates hepatic insulin sensitivity.71
Carvalho et al.72 crossed glucose transporter-4 knockout (GLUT-4−/−) mice with mice overexpressing GLUT-4 only in adipose tissue (AG-4-TG). Overexpression of GLUT-4 reduces fasting hyperglycemia and glucose intolerance in GLUT-4−/− mice to subnormal levels and reduces whole body insulin resistance without restoring glucose transport in muscle.
Adipose fatty acid binding protein knockout (A-FABP−/−) mice have increased fat mass, decreased lipolysis, increased muscle glucose oxidation, and attenuated insulin resistance. Mice overexpressing epithelial FABP in adipose tissue have the exact opposite phenotype. These observations suggest that the balance between adipocyte lipolysis and lipogenesis is remodeled in animal models that reprogram adipokine expression in fat cells and plasma adipokine homeostasis.73
Transgenic mice that overexpress protein phosphatase-1 (PP-1) glycogen-targeting subunit (PTG), driven by the adipocyte fatty acid binding protein promoter (PTG-TG), were found to have exogenous PTG in gonadal, perirenal, and brown fat depots, but not in any of the other tissues studied. PTG overexpression was associated with a modest redistribution of PP-1 to glycogen particles. The results of these experiments suggest that in vivo mechanisms may exist to maintain adipocyte glycogen storage at physiological set points.74
Regucalcin is a multifunctional regulatory protein in intracellular signaling pathways. Female regucalcin transgenic (RC-TG) rats were compared at 7 and 50 weeks of age. As these animals age disorders of lipid metabolism in adipose tissue and liver are observed and levels of gene expression of leptin and adiponectin are reduced.75
Hemochromatosis gene knockout (Hfe−/−) mice have increased plasma adiponectin levels and increased activation of AMP-dependent kinase. Following glucose challenge these animals have decreased glucose excursions compared with WT controls. The decreased glucose excursions are mediated by insulin-independent increased glucose disposal.76
Feeding a high-fat diet to low-density lipoprotein receptor knockout (LDL-R−/−) mice results in obesity, hyperglycemia, hyperlipidemia, and upregulation of serum TNF-α. Mice with augmented vascular TNF-α, produced by a transgene (SM22-TNF-α-TG) driven by the SM22 promoter, had upregulated aortic Msx2, Wnt3a, and Wnt7a transcription proteins that contribute to the aortic calcification seen in type-II diabetic mouse models.77
Infusion of a large number of CD4+ helper T (Th) cells overcomes ovalbumin (OVA)-specific immune tolerance in transgenic rat insulin promoter (RIP)-mOVA mice. This results in CD+ cytotoxic T lymphocyte-mediated destruction of pancreatic islet cells and diabetes. Major histocompatibility complex (MHC) gene knockout (MHC-II−/−) mice lack CD4+ T cells. When CD4+ Th cells are injected into MHC-II−/− mice the OVA-specific CD8+ T-cell response is stimulated. These findings could have significant implications in autoimmunity, antitumor immunity, and regulatory T-cell-dependent immune suppression, important in xenotransplantation studies.78
14.7 Xenotransplantation
Research in xenotransplantation is directed at solving the ever-increasing shortage of organs for human transplantation. It also offers new information related to the development of cell- and tissue-based therapies. Real progress in xenotransplantation has been delayed by the presence of xenoreactive natural antibodies that bind to the foreign cell surface and activate complement resulting in graft rejection. Genetic engineering of donor cells and animals to express human complement inhibitors such as hCD59 has significantly prolonged graft survival. Expression of human α, 2-fucosyltransferase (HT) in pigs modifies the cell-surface carbohydrate phenotype and results in reduced G-α-1, 3-Gal expression and decreased antibody binding. Hearts from transgenic pigs that coexpress hCD59 and HT were perfused with human blood. These hearts demonstrated increased resistance to human serum-mediated lysis.79
Targeting xenograft rejection using single genetic modifications has proven to be ineffective. This led to development of pigs that express human decay accelerating factor (hDAF). Hearts from hDAF transgenic pigs seem to be protected during perfusion with human blood or blood components. They are metabolically stable and maintain acceptable hemodynamic function longer than previous models.80–83 Expression of human complement regulating factor (hCRF) in pig organs seems to prevent hyperacute rejection of these organs after xenotransplantation to nonhuman primates. Arteries from pigs transgenic for human CD46 expression demonstrate augmented endothelium-dependent relaxation responses to bradykinin mediated by the release of endothelium-derived relaxing factors other than NO.84
Pigs transgenic for human membrane cofactor protein (hMCP) were crossed with pigs heterozygous for hDAF. Endothelial cells from this cross were exposed to complement-mediated damage. Cells expressing both hMCP and hDAF were not better protected than cells expressing only hDAF.85
Transgenic CMy-mOVA mice were subjected to adoptive transfer of OVA-specific CD8+ effector T cells expressing OVA in cardiac myocytes. The studies conducted on these animals indicate that granulocytic inflammation sustains CD8+ T-cell-mediated myocarditis. This is important knowledge related to allograft rejection.86
Hearts from transgenic mice expressing viral interleukin (vIL-10), encoded in the Epstein-Barr virus genome, were transplanted as vascularized allografts into unmodified MHC full-mismatched or MHC class II-disparate mice. Results from these studies indicate that a high level of expression of vIL-10 in grafts can exacerbate immunological rejection.87
Liver sinusoidal endothelial cells are capable of selectively suppressing the expansion of IFN-γ-producing cells but are also capable of promoting the outgrowth of IL-4-expression in T helper (Th) cells thus creating an immune suppressive milieu in the liver.88 Donor hearts from B6.C-H2 mice were transplanted into WT and IL-10 transgenic recipients. In IL-10 TG recipients murine IL-10 is produced under control of the human IL-2 promoter. IL-10 expression inhibited the development of allograft vasculopathy.89 Hearts from IL-4 transgenic mice, under the control of a cardiac α-myosin heavy chain promoter, were transferred into unmodified recipients. Although the investigators were able to establish that Th-2 bias may contribute to allograft acceptance in this model, probably by inducing the down-regulation of Th-1-cytokine mRNAs, they were not able to conclude that those reactions were able to confer long-term graft survival.90
Two lines of transgenic mice express β-galactosidase (β-gal) exclusively in endothelial cells (EC). TIE2-lacZ mice express β-gal in all EC and VWF-lacZ mice express β-gal only in the microvascular EC of the heart and brain. Using these animal models Rothermel et al.91 found that EC express intracellular “self” proteins to the immune system but this antigen expression does not delete or stimulate a large population of specific lymphocytes that respond to the same protein following conventional immunization with protein or expression vector DNA. The results of these experiments indicate that context sensitivity in the immune recognition of EC exists and that this phenomena needs to be considered in the context of xenotransplantation.
14.8 Na+/Ca2+ and Na+/H+ Exchangers
Studies on transgenic mice overexpressing canine cardiac Na+/Ca2+ exchanger (cNCX) indicate that this exchanger could be an important Ca2+ transport mechanism in myocardial dysfunction. During ischemia-reperfusion injury increased [Ca2+] in the sarcoplasmic reticulum, the result of increased Na+/Ca2+ exchange activity, can compromise contractile performance and result in cardiac hypertrophy and heart failure.92 The NCX gene contains at least three promoters, H1, K1, and Br1, resulting in multiple tissue-specific variants of the NCX. The H1 promoter directs cardiac-specific expression of NCX in both the embryo and adult and is probably responsible for upregulation of cNCX in response to pressure overload.93
Postrest potentiation was compared in transgenic mice overexpressing cNCX and mice with a deletion mutant of cNCX (Delta680-685). The Delta680-685 mice are devoid of intracellular Na+- and Ca2+-dependent regulatory properties. Postrest potentiation was greater in Delta680-685 mice than in the cNCX animals indicating that ionic regulation of sodium/calcium exchange plays a significant functional role in myocardial contractility.94 Other studies using isolated myocytes from the same models suggest that beat-to-beat changes in cNCX function can occur in vivo.95
Overexpression of cNCX in isolated myocytes from transgenic mice appears to accelerate the decline of [Ca2+]i during relaxation. Increased Ca2+ influx also appears to occur.96 Null mutation of the cNCX gene is lethal resulting in embryonic death (9.0-9.5 days). When transgenic mice expressing canine cNCX were bred to cNCX+/− mice this did not prevent the lethal results.97 Isolated thoracic aortic rings from transgenic mice that specifically overexpress NCX in smooth muscle were used to show that NCX was involved in the forskolin-induced reduction of tension resulting from decreased [Ca2+]i.98 The reverse NCX current was measured in isolated ventricular myocytes from WT and transgenic mice overexpressing NCX. Currents were significantly higher (>twofold) in the transgenic myocytes.99
Transgenic mice that overexpress the human sarcolemmal Na+/H+ exchanger (hNHX), with high cardiac activity, develop hypertrophy, contractile dysfunction, and heart failure. These changes are mainly the result of activation of calmodulin-dependent protein kinase-II (CaMK-II)-dependent phorphorylation of phospholamban (PLB).100
14.9 Inflammatory Cytokines
Several laboratories have reported the results of experiments using transgenic mice overexpressing cardiac-specific TNF-α or β using a variety of different strategies and/or promoters.101–107 These animals develop concentric ventricular hypertrophy,101,102 modify desmin so that it loses its intercalated disk localization and forms aggregates that colocalize with heat shock protein 25 and ubiquitin,106 and demonstrate atrial structural remodeling, downregulation of connexin-40, and an increased incidence of atrial arrhythmias.104
Sekiguchi et al.105 were able to demonstrate that overexpression of TNF-β signaling pathways suppresses PPAR-α activity and reduces cardiac fatty acid β-oxidation in cardiac myocytes. Increased matrix metalloproteinase (MMP)-3 in male TNF-α transgenic mice might regulate activation of MMP-9/gelatinase initiating the progression of cardiac remodeling and the development of heart failure.103 Sustained proinflammatory signaling in the hearts of adult transgenic mice overexpressing TNF-β is associated with a profibrotic phenotype that activates Smad 2/3 leading to increased myocardial fibrosis and loss of compliance in the left ventricle.107
Transgenic mice with murine IL-10 under the control of the human IL-2 promoter fed a high-fat diet demonstrate decreased atherosclerotic lesions compared with WT controls.108 Overexpression of IL-10 by T cells inhibits atherogenesis in LDLR−/− mice fed a high-fat diet.109
Willuweit et al.110 generated transgenic mice expressing a noncleavable transmembrane form of TNF under control of the endothelial-specific tie2 promoter. These mice develop chronic inflammatory disease in kidney and liver but were protected from immune-mediated liver injury following Con A-induced acute hepatitis. Interferon-γ transgenic mice constitutively express IFN-γ in their livers and therefore have high circulating serum levels and develop chronic active myocarditis.111
LPS-induced toxicosis is characterized by hypermetabolism and anorexia in mice. TNF-α−/− mice show a reduction in the amount of LPS-induced hypermetabolism but no change in anorexia. Lymphotoxin-α−/− mice show the same responses to LPS as the TNF-α−/− mice. Results from experiments using soluble tumor necrosis factor receptor-1 fusion protein (TNFR1-IgG−/−) transgenic mice indicate that the hypermetabolic and anorexic responses are independently regulated. Interferon-γ receptor knockout mice showed the strongest anorexic response following LPS exposure.112 TNF-α−/− mice show a reduction in reactive stenosis following endothelial injury.113
Intercellular adhesion molecule-1 knockout (ICAM-1−/−) mice were subjected to myocardial ischemia and reperfusion at 2 h, 1 week, and 3 weeks. The absence of ICAM-1 expression was associated with less myocardial damage following early reperfusion but the size of the infarction or scar formation was not changed.114 β-1-integrin knockout (β-1−/−) mice subjected to myocardial ischemia-reperfusion injury demonstrated reduced neutrophil infiltration in the ischemic regions but there was no effect on the severity of myocardial damage.115
14.10 Peroxisome Proliferator-Activated Receptor
Peroxisome proliferator-activated receptor (PPAR) alpha (PPAR-α) is a key driver of diabetes-related lipid metabolic dysregulation. Transgenic mice overexpressing PPAR-α (PPAR-α-TG) have decreased GLUT-4 mRNA levels and decreased glucose uptake.116 PPAR-α-TG mice also demonstrate reduced β-adrenergic responses in contractility and chronotropy compared with WT controls.117 Electrophysiological experiments conducted on myocytes from both the left and right ventricles of PPAR-α-TG and WT control mice reveal marked K+ current remodeling in the mice overexpressing PPAR-α.118
DOCA-salt hypertension was induced in WT controls and PPAR-α−/− mice. These studies reveal that the PPAR-α pathway induces renal tubular 20-hydroxyeicosatetranenoic acid (20-HETE) production regulating Na+ retention and blood pressure.119 The Na+ and blood pressure regulation in PPAR-α−/− mice is also affected via amiloride- and thiazide-sensitive mechanisms, so this model is not hypertensive but develops salt-sensitive hypertension despite defective fatty acid oxidation.120
Transgenic mice expressing PPAR-γ1 in the heart, via the cardiac α-myosin heavy chain promoter, have increased cardiac expression of fatty acid oxidation genes and increased lipoprotein triglyceride uptake. Heart glucose gransporter-4 (GLUT-4) mRNA expression and glucose uptake are not decreased. These animals develop DCM associated with increased lipid and glycogen stores, and abnormal ultrastructural changes in the mitochondria and cristae. Loss of PPAR-γ in myeloid cells impairs alternative macrophage activation and predisposes PPAR-γ−/− mice to the development of diet-induced obesity, insulin resistance, and glucose intolerance.121
14.11 Renin-Angiotensin System
Transgenic mice overexpressing angiotensinogen have decreased caspase recruitment domain (ARC), catalase expression, and protein kinase-2 (CK-2) levels. Catalase, CK-2, and ARC are an antihypertrophic pathway in the heart.122 Transgenic mice that overexpress rat angiotensinogen in their proximal tubule cells were used to demonstrate that the intrarenal renin-angiotensin system (RAS) stimulates proximal tubule cell apoptosis and tubulointerstitial fibrosis, at least in part, by enhanced NADPH oxidase activity and ROS generation independent of hypertension.123
Double-transgenic rats harboring both human renin and human angiotensinogen genes were compared with Sprague-Dawley normal controls in experiments with and without a p38 inhibitor. The p38 mitogen-activated protein kinase inhibitor improved survival, target organ damage, and arrhythmogenic potential in these double-transgenic rats.124
Angio-II subtype 1A receptor (ATR-1A)−/− and WT mice were subjected to 2K1C renovascular hypertension and were compared. Arterial blood pressures were significantly lower in the ATR-1A−/− mice. Clip placement, ATR-2 blockade, and NOS activity did not alter the hypertension in this knockout model.125,126
14.12 Bradykinin-2 Receptor
Myocardial eNOS and mitogen-activated protein kinases (MAPK), including ERK, p38, and JNK, protein expression were measured in bradykinin-2 receptor (BR-2) knockout (BR-2−/−) mice. Disruption of BR-2 resulted in maladaptive cardiac hypertrophy with downregulation of eNOS and upregulation of MAPK. These results were reversed by treatment with a NOS inhibitor.127 BR-2−/− and their normal 129/SvEvTac WT controls were compared to evaluate the hypertensive effects of DOCA-salt and aortic coarctation. In this model kinins, acting via B-2 receptors, do not participate in the maintenance of normal arterial pressures or the establishment and maintenance of hypertension resulting from either treatment. Both BR-2−/− and WT mice reacted similarly to DOCA-salt and aortic coarctation when treated with angiotensin-converting enzyme inhibition, i.e., hypertension and ventricular hypertrophy were prevented.128 However, Madeddu et al.129 used BR-2−/− and WT to conclude that kinins, acting on the BR-2, exert protection against excessive hypertension in the early phases of 2K1C.
14.13 Apolipoprotein-E and Low-Density Lipoprotein Knockout Models
Apolipoprotein-E (ApoE−/−), C57BL/6 WT, and sham-operated mice were compared following 2K1C renovascular hypertension. ApoE−/− mice developed less of an increase in aortic wall area than that seen early in the WT controls but the aortic lumen area was increased to a similar magnitude in both 2K1C models.130 The effects of 1K1C and 2K1C renovascular hypertension were compared in ApoE−/− mice. 2K1C animals developed significantly more atherosclerotic lesion area and enhanced accumulation of macrophages, accompanied by a parallel increase in scavenger receptor-A expression in the macrophages, in the aortic sinus than did the 1K1C mice. This suggests that increased generation of Angio-II in the 2K1C model may initiate and promote atherosclerosis via activation of VSMC.131 When ApoE−/− mice were fed a high-fat diet and subjected to 2K1C renovascular hypertension they were unable to increase renin secretion and blood pressure in response to diminished renal perfusion compared with ApoE−/− mice fed a regular diet.132
ABCG1 is a member of the G subfamily of ATP-binding cassette (ABC) transporters. Overexpression of ABCG1 alone can induce cholesterol efflux to HDL. When bone marrow from ABCG1−/− mice is transplanted into ApoE−/− or LDL−/− mice the amount of atherosclerotic lesion developed is reduced. ABCG1−/− mice were crossed with LDL−/− mice and the offspring (ABCG1/LDL−/−) were compared with LDL−/− mice after both were placed on a high-fat diet. No major differences were observed in total plasma lipids but IDL-LDL cholesterol was significantly increased in the double-knockout mice as was the amount of atherosclerotic lesions. Plasma levels of MCP-1 and TNF-α were also increased in the double-knockout mice.133
BACH-1 is a nuclear protein that directly interacts with the highly conserved C-terminal BRCT repeats of the tumor suppressor BRCA-1. BACH-1/ApoE−/− (double knockout) mice were generated by crossing BACH-1−/− and ApoE−/− mice. These mice were fed a high-fat diet for 8 weeks. The double-knockout mice had significantly less atherosclerotic lesion formation, a result of upregulation of heme oxygenase-1 in the endothelium and, to a lesser extent, in the vascular smooth muscle.134
Mice transgenic for lysozyme (LZ-TG) are resistant to both acute and chronic oxidative stress. They display decreased circulating levels of pro-oxidant advanced glycation end products (AGEs). The degree of atherosclerosis was evaluated in LZ-TG mice crossed with ApoE−/− mice (LZ/ApoE−/−). Serum levels of LZ were increased, levels of AGE and 8-isoprostanes were decreased, but hyperlipidemia was similar to ApoE−/− controls. The LZ/ApoE−/− transgenic mice had significantly less atherosclerotic plaque, neointimal lesions, and inflammatory infiltrates than the controls, and femoral arterial lesions following injury were also reduced.135
14.14 Toll-Like Receptors
Two strains of toll-like receptor TLR-4−/− mice (C57/BL10 ScCr and C3H/HeJ) were compared with control strains (C57/BL10 ScSn and C3H/OuJ). All four groups were subjected to 1 h of coronary ligation followed by 24 h of reperfusion. The TLR-4−/− animals had less infarcted area, less neutrophilic infiltration, less myeloperoxidase activity, and fewer lipid peroxides and complement deposition than controls.136 TLR-4−/− mice demonstrate improved left ventricular function and reduced LV remodeling following coronary ischemia-reperfusion injury than WT controls.137 When TLR-2−/− mice were compared with WT controls, following coronary ischemia-reperfusion injury, survival rates were significantly higher in the TLR-2−/− mice but infarct size and degree of inflammatory cell infiltration were similar. There were significantly less myocardial fibrosis, TGF-β, and collagen type-I mRNA expressions in the noninfarct area of the knockout mice.138 The same laboratory found that inflammatory responses and neointimal hyperplasia were reduced in TLR-2−/− mice following the placement of an external cuff around the femoral artery.139
14.15 Caveolin-1 (Cav-1)
Ectonucleoside triphosphate diphosphohydrolase-1 (CD39) is a plasma membrane ectoenzyme that regulates purinergic receptor signaling by controlling the levels of extracellular nucleotides. Using Cav-1−/− mice Papanikolaou et al.140 demonstrated that caveolae are not essential for the enzymatic activity of CD39 or for its targeting to plasma membrane. Cav-1 TG mice that overexpress Cav-1 show that endothelial-specific expression of Cav-1 impairs endothelial NOS activation, endothelial barrier function, and the angiogenic response to exogenous VEGF and ischemia. VEGF-mediated phosphorylation of Akt and eNOS were significantly reduced in Cav-1 TG mice compared WT littermates.141
14.16 Long QT Syndrome
Transgenic rabbits that lack slowly activating delayed rectifier K+ currents (IKs), designated LQT-1 (long QT-1), and another rabbit model that generates repolarizing K+ currents (IKr) (long QT-2)142 have been generated. These TG rabbits were produced by expression of pore mutants of the human genes designated KCNQ1 (KvLQT1-Y315S) and KCNH2 (HERG-G628S) in the heart. Both TG rabbits demonstrate prolongation of the QT interval and prolonged action potentials associated with the elimination of IKs and IKr currents. In these animals the elimination of one repolarizing current was associated with downregulation of the reciprocal repolarizing current. This is different from the compensatory upregulation that has been observed in long QT syndrome (LQTS) mouse models.143–145
14.17 Nuclear Factor Kappa-B
Mice expressing a luciferase reporter, whose transcription is dependent upon nuclear factor kappa-B (NF-κB) activation, were studied after myocardial infarction. A significant increase in NF-κB activity was observed with the maximum response 3 days following infarction.146 Mice with NF-κB subunit p50 knocked out (p50/NF-κB−/−) were compared with WT controls following coronary artery ligation and ischemia-reperfusion injury. Ventricular dilation, early mortality, and ischemia-reperfusion injury were significantly reduced in the p50/NF-κB−/− animals compared WT controls.46,147,148 Pharmacological inhibition of aldose reductase, or knockdown of the enzyme by small interfering RNA, prevents the activation of NF-κB and the release of TNF-α and increased survival in mice injected with lethal doses of LPS.149
Transgenic Cre/lox mice with endothelial cell-restricted NF-κB super-repressor IkappaBalphaDeltaN (Tie-1-DeltaN) overexpression were subjected to hypertension by feeding a high-salt diet and l-NAME plus infusion of Angio-II. Both Tie-1-DeltaN and control mice demonstrated equal levels of hypertension. Tie-1-DeltaN mice developed less renal injury, reduced inflammatory responses, and less albuminuria than controls. There was also a significantly reduced expression of ICAM-1 and VCAM-1, targets of NF-κB. The investigators concluded that in vivo NF-κB suppression in endothelial cells blocks a signaling cascade leading to reduced hypertension-induced renal damage in the face of hypertension.150
A20 was first described as a TNF-inducible gene in human umbilical vein endothelial cells that inhibits NF-κB signaling and protects against apoptosis, inflammation, and cardiac hypertrophy. Transgenic mice containing the human A20 gene, under the control of the α-myosin heavy chain promoter, exhibit cardiac-specific overexpression of A20. Following acute coronary artery ligation these mice have improved cardiac function and less cardiac remodeling, apoptosis, inflammation, and fibrosis than controls.151
14.18 Orphan Nuclear Receptors
Orphan nuclear receptors, ERR-α and ERR-γ, act as nonobligatory heterodimers and target a common set of promoters involved in energy substrate uptake, the production and transport of ATP across mitochondrial membranes, intracellular fuel sensing, Ca2+ handling, and myocardial contractility.152 Disruption of the ERR-γ gene, highly expressed in fetal and postnatal hearts of mice, results in lactatemia, cardiac arrhythmias, and mortality during the first week of life.153
Steroid receptor coactivator-1 (SRC-1) is a transcriptional coactivator for nuclear receptors including estrogen receptors (ER). SRC-1−/− mice were used in studies to demonstrate that SRC-1 is expressed in endothelial cells, VSMC, and neointimal cells. Neointimal growth, induced by injury to the common carotid artery, was almost completely inhibited by estrogen in WT mice but only partially inhibited in the SRC-1−/− mice.
14.19 Troponin
Troponin T (TnT) is essential to the Ca2+ regulatory system of striated muscle. Three specific fiber-type TnT genetic isoforms encoding cardiac, slow- and fast-twitch skeletal muscle have been identified. Intact cardiac muscle strips from transgenic mice overexpressing fast-twitch skeletal muscle TnT show decreased contractility at acidic pH than the same preparations from WT control mice.154
Transgenic mice that overexpress cardiac troponin-I (TnI) have serine residues normally targeted by protein kinase A (PKA) that are mutated to aspartic acid and mimic constitutive phorphorylation. These animals have slightly enhanced basal systolic and diastolic left ventricular function but display a marked increase in frequency-dependent inotropy and relaxation compared with control mice.155
Hemodynamic studies were conducted in mice that express normal levels of PLB and express either cardiac troponin-I (PBL/cTnI) or the slow skeletal isoform of TnI (PBL/ssTnI). The latter cannot be phosphorylated by PKA. Another construct of mice that do not express PBL and express either cTnI (PBL−/−/cTnI) or ssTnI (PBL−/−/ssTnI) was also studied. PBL/ssTnI hearts demonstrated a significant reduction in -dP/dt compared with PLB/cTnI hearts. β-adrenergic stimulation increased HR similarly in both groups but at the highest levels of stimulation -dP/dt was significantly less in PLB/ssTnI than in PLB/cTnI hearts. The authors concluded that phosphorylation of cTnI significantly contributes to the enhanced rate of left ventricular relaxation during β-adrenergic stimulation.156
14.20 Chromogranin A
Genetic ablation of the chromogranin A (Chga −/−) gene in mice results in hypertension, reduced myocardial function, and reduced responses to β-adrenergic and ET-1 stimulation. Pretreatment of Chga−/− mice with catestatin (human chromogranin A352-372) prevents the adverse effects of removing this gene.157,158
14.21 Lectin-Like Oxidized Low-Density Lipoprotein Receptor
ATR-1 activation upregulates the expression of lectin-like oxidized low-density lipoprotein receptor LOX-1. LOX-1 activation upregulates ATR-1 expression. Angio-II-induced hypertension, but not NE-induced hypertension, was attenuated in LOX-1−/− mice. This indicates that LOX-1 is a key modulator of Angio-II-induced hypertension and cardiac remodeling.159
LOX-5−/− mice were used to demonstrate that the loss of this receptor has no effect on cardiac ischemia-reperfusion injury, but neutrophil infiltration and TNF-α expression were greater in the LOX-5−/− animals.160
14.22 Junctin
Junctin (JCN) is a transmembrane protein located in the cardiac junctional sarcoplasmic reticulum (SR). It binds to the ryanodine receptor [calsequestrin (CSQ)] and to triadin-1. Atria from 3-week-old transgenic mice that overexpress junctin (JCN-TG) demonstrate reduced expression of triadin-1. This results in a higher SR Ca2+ load but no changes in contractility or HR. Atria from 6-week-old JCN-TG show a compensatory downregulation of the ryanodine receptor that seems to offset the effects of junctin overexpression. In 18-week-old JCN-TG the progressive decrease in ryanodine receptor density seems to contribute to decreases in atrial contractility and HR during stress.161 JCN-TG mice demonstrate impaired myocardial relaxation possibly as a result of reduced NCX expression and/or increased leak of SR Ca2+. Altered shortening-frequency excitation-contraction coupling in these animals may be a consequence of impaired excitation-contraction coupling with depressed SR Ca2+ release at higher stimulation rates.162 Transgenic mice overexpressing canine junctin also show changes in the packing of CSQ in the junctional SR and a facilitation of the association of SR and T tubules.163
Crossbreeding of mice with heart-specific overexpression of both triadin and junctin resulted in offspring (JxT-TG) with a stable threefold expression of total triadin but normal levels of junctin. These animals display cardiac hypertrophy, prolonged basal relaxation, depressed responses to β-adrenergic stimulation, and altered Ca2+ transients.164
14.23 Connexin
Connexins (Cx) are labeled with a number that represents the molecular mass of the amino acid sequences, in kDa. When identical connexin isotypes form both Connexins of a gap junction a homomeric gap junction channel is formed. When different connexin isotypes combine a heteromeric channel is produced. In the mammalian heart seven connexins have been identified: Cx37, Cx40, Cx43, Cx45, Cx46, Cx50, and Cx57. The Cx40−/− mouse has been used to study mechanisms of arrhythmogenesis.165,166
Aside from being components of gap junctions, connexins can also comprise large, nonselective pores known as hemi channels. The Cx30.3/lacZ transgenic approach was used to identify Cx30.3 in the renal medulla in mice, rats, and rabbits.167 Cx45 expression has been found in the juxtaglomerular apparatus (JGA), as well as in the heart. Transgenic mice that express lacZ coding DNA under the control of the Cx45 promoter were used to discover that Cx45 is involved in the propagation of JGA vascular signals and thus in the regulation of renin release and the control of blood pressure.168
Cx40−/− mice have a misdirection of renin-expressing cells from the medial layer of afferent arterioles to the perivascular tissue, extraglomerular mesangium, and periglomerulular and peritubular interstitium. The aberrant renin-producing cells in these animals also express significant amounts of COX-2 mRNA and this, apparently, contributes to the renin secretion.169 Cx40−/− mice were also used to show that renin-secreting cells are coupled to each other and to endothelial cells by Cx40-containing channels.170 Transgenic mice were produced in which the coding region for Cx43 was replaced by Cx26. Both neonatal and adult Cx43 knock-in Cx26 mice demonstrated slowed ventricular conduction patterns, and both males and females were infertile from impaired gametogenesis.171
Calrectin is a Ca2+-binding protein of the endoplasmic reticulum and plays an essential role in cardiac development. Cre-loxP transgenic mice with spatiotemporal overexpression of calrectin were created. Protein levels of Cx40, Cx43, and myocyte-enhancer factor-2C were lower than the level of these proteins in control hearts.172 The overexpression of calrectin and decreased levels of Cx were associated with arrhythmias, ventricular dilatation, and sudden death in 6-10-week-old offspring, and marked edema at 7 weeks of age. The expression of hyperpolarization-activated cyclic nucleotide-gated channel-1, an essential component for cardiac pacemaker activity, was also decreased in the hearts of these mice.
14.24 Phospholamban
Mouse studies have shown that PLB is a key regulator of SR Ca2+ cycling and cardiac function but mice differ significantly from humans in how they regulate calcium. Rabbits have proven to be a better model for the study of calcium handling. Transgenic rabbits that overexpress PLB die early from severe skeletal muscle wasting and myocardial dysfunction. A viable transgenic line exhibiting only a 30% increase in cardiac PLB shows isolated foci of cardiac pathology but cardiac function and response to β-adrenergic stimulation are normal.173 PLB phosphorylation is a CaMKII-dependent process. Mice with transgenic overexpression of CaMKII-delta-C (chronic overexpression) develop heart failure with impaired SR Ca2+ loading during late acidosis compared with WT controls. This response may be associated with decreased SR Ca-ATPase and decreased PLB expression. However, adenovirus-mediated gene transfer of CaMKII-delta-C in isolated mouse and rabbit cardiomyocytes (acute overexpression) actually slightly improves recovery following increased SR Ca2+ loading during late acidosis.174
14.25 Fas Ligand
Fas ligand (FasL) is a potent proapoptotic type-II transmembrane protein. Its forced expression can induce a dramatic inflammatory response.175 Transgenic mice with cardiac-specific expression of soluble Fas (sFas), a competitive inhibitor of FasL, were injected with either LPS or control vehicle. Neutralization of FasL by expression of sFas significantly preserved cardiac function and reduced inflammatory responses to LPS in the heart.176
Mice with cardiac-specific expression of sFas and double homozygous TG mice that express both monocyte chemoattractant protein-1 (MCP-1) and sFas were used to show that FasL released from infiltrating monocytes plays a critical role in the adverse effects of MCP-1 expression. The results suggest that Fas/FasL signaling could be a novel therapeutic target for certain types of heart failure.177,178
14.26 Proteases, Metalloproteinases, and ATPases
A novel class of protease-activated receptors (PARs) characterized by seven transmembrane G-protein-coupled domains are activated by serine proteases such as thrombin, trypsin, and tryptase. PAR-1, PAR-2, and PAR-4 have been implicated in vascular development via studies using transgenic mice and synthetically tethered ligand domains.179 PAR-4−/− mice have prolonged bleeding times because of the lack of PAR-4 function in platelets and are thereby protected against thrombosis.180 Male PAR-2−/− mice have a moderate elevation of systolic arterial and pulse pressures, but no change in diastolic pressures along with a lack of hypertensive response to high-salt diet. This model displays only a subtle difference in the time course of hypertension in response to Angio-II infusion.181
Extravascular inflammation was induced by injection of recombinant soluble murine tissue factor (TF) in the hind paw of PAR-1−/−, PAR-2−/−, PAR-3−/−, and PAR-4−/− mice. There was no effect on the inflammatory response in PAR-1−/−, PAR-2−/−, or PAR-3−/− mice but the PAR-4−/− mice demonstrated significantly less soluble TF (sTF)-induced swelling of the injected paw. These results indicate that sTF induces inflammation via a thrombin-dependent pathway, and both fibrin deposition and platelet activation are essential steps with the activation of PAR-4 essential to the process.182
Corin is a transmembrane serine protease that has been isolated from the heart. It is responsible for converting proatrial natriuretic peptide (pro-ANP) to active ANP. Corin−/− mice do not convert pro-ANP to ANP resulting in salt-sensitive hypertension. Interestingly the hypertension is exacerbated in pregnant corin−/− mice.183
Matrix metalloproteases (MMPs) are enzymes that regulate matrix remodeling in response to hypertension. Transgenic mice that overexpress cardiac MMP-1 and matched WT were subjected to hypertension induced by suprarenal banding of the aorta. Overexpression of MMP-1 attenuated the development of myocardial fibrosis, prevented left ventricular dilatation, and preserved myocardial function.184 Transgenic rabbits that overexpress human matrix metalloproteinase-12 (hMMP-12-TG) were fed a 1% cholesterol diet for 6 weeks. They developed larger atherosclerotic plaques, more significant degradation of the aortic internal elastic layer, and increased numbers of infiltrating macrophages and smooth muscle cells into the atherosclerotic lesions than WT control rabbits.185 Tissue inhibitor of metalloprotease-3 (TIMP-3) inhibits MMP and regulates angiogenesis. TIMP-3−/− mice demonstrate enhanced MMP activity in the retinal choroids and develop abnormal vessels with dilated capillaries.186
Strain differences are demonstrated between FVB and B6 mice where disintegrin and metalloproteinase-17 (ADAM-17) mRNA expression is higher in the liver, macrophages, and aorta in FVB.LDLR−/− mice than in B6.LDLR−/− mice. Macrophages from FVB.LDLR−/− mice demonstrate fivefold increases in PMA-induced shedding of TNF-α and 32% increases in release of TNF-receptor-I than B6.LDLR−/− animals.187
The copper transporter Menkes ATPase (MNK) is overexpressed in MNK-mutant mice. This model was used to demonstrate that MNK plays an important role in modulating Angio-II-induced hypertension and endothelial function by regulating extracellular superoxide dismutase (SOD-3) activity and vascular superoxide production.188
14.27 Binary Calsequestrin/P2Xr-Purinergic Receptor (CSQ/P2X4R) Transgenics
P2X4R is a ligand-gated ion channel. When activated by extracellular ATP it allows Ca2+ influx. Transgenic cardiac overexpression of human P2X4R results in increased ventricular contractility. Transgenic mice with cardiac overexpression of canine CSQ have ventricular hypertrophy, heart failure, and early mortality. Crossing P2X4R-TG with CSQ-TG produced CSQ/P2X4R-TG offspring with prolonged survival, less ventricular hypertrophy, and restored β-adrenergic responsiveness.189
14.28 pro-ANP Gene Disrupted Mouse
ANP−/− mice received either vehicle or an Angio-II type-2 receptor (ATR-2) antagonist. Both groups were placed on a high-salt diet. The studies indicate that ATR-2 blockade in salt-sensitive hypertensive/ventricular hypertrophic ANP−/− mice results in changes in myocardial extracellular matrix components that translate into decreased ventricular hypertrophy.190
14.29 Macrophage Colony-Stimulating Factor
Homozygous osteopetrotic mice (Op/Op) are deficient in macrophage colony-stimulating factor (M-CSF). Adult Op/Op, heterozygous (Op/+), and WT mice were subjected to 14 days of Angio-II (1,000 ng/kg/min) or vehicle infusion. Op/Op mice demonstrated less endothelial dysfunction, less vascular remodeling, and less oxidative stress than WT.191 M-CSF has also been shown to stimulate differentiation and proliferation of monocyte/macrophage lineage and is involved in neointimal formation following vascular injury. Recombinant human M-CSF (500 μg/kg/day) or vehicle was administered for 10 consecutive days, starting 4 days prior to mechanical injury to the femoral artery, in C57BL/6 mice. M-CSF accelerated early neointimal formation after vascular injury suggesting that inhibition of this system might have therapeutic potential in the treatment of a number of cardiovascular diseases.192
14.30 Endothelin-1
Endothelin-1 (ET-1) is a potent endogenous vasoconstrictor that contributes to vascular remodeling in hypertension and other cardiovascular diseases. Amiri et al.193 targeted expression of the human pre-pro-ET-1 gene to the endothelium using the Tie-2 promoter in C57BL/6 mice. Compared with WT littermates the TG mice exhibited threefold higher vascular tissue ET-1 mRNA and sevenfold higher ET-1 plasma levels, but there was no significant difference in blood pressures between the two groups. Higher ET-1 levels resulted in structural remodeling and endothelial dysfunction in resistance vessels. The experiments conducted suggest a direct, nonhemodynamic, effect of ET-1 on the vasculature involving activation of vascular NADPH-oxidase.
Aryl hydrocarbon receptor (AHR) knockout (AHR−/−) mice develop cardiac hypertrophy of unknown origin. When AHR−/− mice were exposed to a modest decrease in inspired pO2 they became hypoxic and hypertensive and plasma ET-1 levels were significantly increased. The hypertension was not associated with increased pulmonary pre-pro-ET-1 mRNA expression.194
14.31 Elastin
Mice harboring targeted deletion of the elastin (ELN) gene (ELN+/−) show many of the same features of supravalvular aortic stenosis (SVAS) seen in humans, including changes in cardiovascular hemodynamics and arterial wall structure. ELN+/− mice are hypertensive at birth compared with WT counterparts. The inner diameters of arteries from ELN+/− mice are smaller at any given intravascular pressure than that of WT mice.195,196
Exact modeling of SVAS and other elastin gene abnormality diseases in mice is difficult because of structural differences between the human and mouse genes. A humanized elastin mouse with elastin production controlled by the human elastin gene has been developed. The human elastin protein appears to interact with the mouse elastin to form functional elastic fibers. When the h-ELN is expressed in the elastin haploinsufficient background (ELN+/−) the hypertension and cardiovascular changes are reversed.197
14.32 α-2-Antiplasmin
α-2-Antiplasmin (α-2-AP) is the major circulating inhibitor of plasmin. Plasmin plays a significant role in the regulation of intravascular fibrinolysis. Angio-II-induced arterial wall thickening, vascular cell proliferation, apoptosis, c-Myc, and collagen-I expression are all significantly decreased, along with perivascular fibrosis, in α-2-AP−/− mice compared to WT mice.198
14.33 cAMP Response Element Binding Protein
cAMP response element binding protein (CREB), at serine-133, regulates gene expression in the heart. The functional significance of CREB-S133 phosphorylation was studied by comparing TG mice in which a phosphorylation-resistant (CREB-S133A) mutant, containing either an intact or a mutated leucine zipper domain (CREB-S133A-LZ), was expressed in the heart. The 1-year survival of mice from two CREB-S133A-LZ (TG) lines was equivalent to non-TG littermate controls. TG CREB-S133A mice died from heart failure at about 30 weeks of age. CREB-S133A mice demonstrate altered gene expression characteristic of heart failure whereas the CREB-S133A-LZ mice did not.199
14.34 Fatty Acid Transport Protein: CD36
Hearts from otherwise normal aged mice show intramyocardial lipid accumulation, reduced fatty acid and glucose oxidation, and deterioration in supplies of cardiac ATP. These changes accompany myocardial remodeling and impaired myocardial function. Hearts from CD36−/− mice have lower levels of intramyocardial lipids, improved mitochondrial-derived ATP production, improved myocardial function, and a blunted hypertrophic response compared with age-matched WT. This indicates CD36 is a potential mediator of these aging-associated functional, structural, and metabolic changes.200
14.35 Clotting Factor XIII
Clotting factor XIII (FXIII) is a transglutaminase involved in wound healing. FXIII−/− and FXIII+/− mice suffer left ventricular rupture and die within 5 days following ligation of the left coronary artery. FXIII−/− mice receiving intravenous FXIII replacement therapy for 5 days have normal survival rates but still have adverse ventricular remodeling compared with WT controls.201
14.36 Apelin
Apelin describes a novel endogenous peptide system thought to be involved in a number of physiological processes, including cardiovascular development and function, control of fluid homeostasis, and obesity. It is also a catalytic substrate for ACE (angiotensin-converting enzyme)-2. Endogenous Apelin is essential for the maintenance of cardiac contractility during pressure overload and aging. Apelin−/− mice develop a progressive loss of cardiac contractility associated with systolic dysfunction, but without histological abnormalities, or heart failure. Pressure overload normally induces upregulation of Apelin expression in the heart, but when Apelin−/− mice were exposed to pressure-overload-induced heart failure there was no significant difference in the LV hypertrophy response compared with WT.202
14.37 T-Box Transcription Factor
Mutations in human TBX-5, a member of the T-box transcription factor gene family, are associated with congenital cardiac septal defects and isomerism in the autosomal dominant Holt-Oram syndrome. Overexpression of TBX-5 in embryonic chick hearts inhibits myocardial growth and trabeculation and suppresses embryonic cardiomyocyte proliferation.203
14.38 Thrombospondin-1 and Its Receptor CD47
Thrombospondin-1 is a key regulator of NO signaling. It limits the angiogenic activity of NO in EC, its vasodilator activity in VSMC, and its antithrombotic activity in platelets. Thrombospondin-1−/− or CD47−/− mice have hyperdynamic responses to NO and improved ability to respond to ischemic stress.204
14.39 Polyomavirus Middle T Antigen
The pronuclear microinjection method was used to introduce purified polyomavirus middle T antigen (PyMT) transgene into the chromosomes of fertilized mice ova. The transgenic positive animals were used to develop future generations by hybridization. PyMT expression in the F1 generation produced mice with venous malformations.205
14.40 Thrombopoietin Receptor
Mice that express a full-length thrombopoietin receptor (Mpl) transgene, in the absence of other Mpl isoforms, were generated under the control of a 2-kb Mpl promoter in Mpl−/− background animals. Transgene expression was decreased during late megakaryocyte maturation. This resulted in diminished Mpl expression in platelets. Platelets exert a negative feedback on thrombopoiesis by binding and consuming thrombopoietin in the circulation via the action of Mpl. The significant reduction of Mpl protein in platelets from Mpl-TG mice results in a marked elevation of the numbers of megakaryocytes and platelets during the steady state.206
14.41 Vascular Endothelial Growth Factor
A VEGF transgenic mouse, rhodopsin promoter, model (tro29VEGF) that mimics nonproliferative diabetic retinopathy and mild proliferative diabetic retinopathy was used to characterize early stages of concomitant vascular and neural retinal damage. Rhodopsin was detected at postnatal day (P)-5 and reached mature levels by P-15. VEGF protein expression was transient but peaked at P-10 to P-15. In tro29VEGF mice the formation of capillary beds was accelerated compared with that in WT controls. Vascular lesions were more numerous by P-28 in the TG mice. Retinal layer thinning tracked abnormal vessel growth.207
14.42 Osteopontin
Osteopontin (OPN) is a noncollagenous adhesion molecule protein identified in atherosclerotic lesions. OPN-TG and OPN-WT mice fed a 1.25% cholesterol diet for 16 weeks develop atherosclerotic lesion but the lesions in the OPN-TG mice were significantly larger. This study also showed that OPN induces fatty-streak lesion formation and inhibits IL-10 production by macrophages.208
14.43 ATP-Binding Membrane Cassette Transporter-A1
Atherosclerotic disease involves a host of processes including reverse cholesterol transport (RCT). RCT is a pathway for transport of cholesterol that has accumulated in the vessel wall to the liver where it can be excreted. Major constituents of RCT include HDL and apolipoprotein A-I (ApoA-I), considered acceptors, and enzymes including cholesterol acyltransferase (LCAT), phospholipid transfer protein (PLTP), hepatic lipase (HL), and cholesterol ester transfer protein (CETP). A critical portion of this process is cholesterol efflux. Accumulated cholesterol is removed from macrophages in the subintima of the vessel wall by ATP-binding membrane cassette transporter-A1 (ABC-A1), or by other mechanisms including passive diffusion, scavenger receptor-B1, caveolins, and sterol 27-hydroxylase. The cholesterol is then available to be collected by HDL and/or ApoA-I. Studies in ABC-A1−/− mice demonstrate that disruption of ABC-A1 genes induces atherosclerosis and that proinflammatory cytokines exert atherosclerotic effects, at least partially, by impairing RCT and cholesterol efflux.209
14.44 The K+/Cl− Cotransporter KCC3
Mice with a targeted disruption of the KCC3 gene (KCC3−/−) develop neurodegeneration of the peripheral and CNS and develop hypertension similar to Andermann syndrome in humans. Data from these animals indicate that local control of vascular myogenic tone does not require KCC3. The hypertension that develops is, apparently, dependent upon increased sympathetic tone.210
14.45 Aldosterone Synthase Overexpression
TG mice overexpressing the terminal enzyme of aldosterone biosynthesis, aldosterone synthase (AS), in cardiac tissue were developed by gene targeting with the α-myosin heavy chain promoter. When compared with WT, TG AS mRNA was increased 100-fold and aldosterone concentrations were increased 1.7-fold. This increased cardiac production of aldosterone-induced major dysfunction in coronary arteries, independent of the endothelium but there were no changes in myocardial function or structure.211
14.46 Cysteine and Glycine-Rich Protein-2 (CSRP-2)
A gene disruption cassette was cloned into exon 4, and a mouse strain lacking functional cysteine and glycine-rich protein-2 (CSRP-2) (CSRP-2−/−) was created. CSRP-2−/− mice have subtle alterations in cardiac ultrastructure including changes in myocardiocyte thickness and hypertrophy.212
14.47 Parathyroid Hormone Type-1 Receptor and PTH/PTH-Related Protein
Transgenic mice that overexpress parathyroid hormone type-1 receptor (PTH1R) in smooth muscle have been used to demonstrate that the PTH/PTH-related protein (PTHrP)/PTH1R system is an important regulator of cardiovascular function. Unfortunately, PTHrP/TTH1R−/− mice are not viable, so this model is unavailable for study. Three weeks after the intravenous administration of either human-PTH1R or hPTH1R tagged with green-fluorescence protein there was generalized expression of both hPTH1R mRNA and protein, particularly in blood vessels, liver, heart, kidney, and CNS. PTH1R overexpression decreased blood pressure and renal tone but also decreased HR. The TG mice demonstrated marked reductions in the production of angiotensinogen in the liver and in circulating levels of both renin activity and angiotensinogen.213
14.48 Vitamin D Receptor
Absence of a functional vitamin D receptor (VDR), or the key activating enzyme, 25-OHD-1-α-hydroxylase (CYP27B1), in mice results in a bone and growth plate phenotype that mimics humans with congenital CYP27B1 mutations or severe vitamin D deficiency. VDR−/− mice also develop increased renin activity resulting in hypertension, myocardial hypertrophy, and increased thrombogenicity.214
14.49 Thromboxane Receptor (Tp)
Tp−/− and WT control mice were exposed to 21 weeks of l-NAME and a 6% salt diet. Hypertension and myocardial hypertrophy were attenuated in the Tp−/− mice but renal hypertrophy and the severity of glomerulosclerosis, tubule vacuolization, and chronic inflammation of the interstitium were enhanced.215
14.50 T and B Cells
Mice lacking T and B cells (RAG-1−/−) exposed to Angio-II infusion- or DOCA-salt-induced hypertension exhibit a blunted hypertensive response and do not develop functional abnormalities of the vascular system. This result is, apparently, due to the lack of T cells.216
14.51 Vanilloid Type-1 Receptors (TRPV-1)
Hypertension was produced by uninephrectomy and DOCA-salt in WT and TRPV-1−/− mice. There was no difference in the level of hypertension produced between the two groups but there were significant increases in the urinary excretion of albumin and 8-isoprostane, glomerulosclerosis, renal cortical tubulointerstitial injury, nuclear antigen-positive cells, and renal monocyte/macrophage infiltration in the TRPV-1−/− animals.217
14.52 Serotonin Transporter (SERT)
Negative results are also important. SERT−/− mice and rats have the same basal arterial pressures as WT controls and the same response to DOCA-salt hypertension. SERT does not seem to be involved in the regulation of arterial blood pressure.218
14.53 CC Chemokine Receptor-2 (CCR-2)
CCR-2−/− and age-matched WT control mice were infused continuously with either Angio-II (5.2 ng/10 g/min) or vehicle for 2 or 4 weeks. Angio-II infusion resulted in similar degrees of hypertension and LV hypertrophy in both strains. CCR-2−/− mice with Angio-II-induced hypertension had significant decreases in oxidative stress, macrophage infiltration, albuminuria, and renal damage. GFR was significantly greater in CCR-2−/− than in the WT controls.219
14.54 Thymosin β-4
Hinkel et al.220 have shown that the retrograde infusion of embryonic eEPCs into zones of myocardial ischemia provides rapid paracrine protection against ischemia-reperfusion injury. Intact pigs were subjected to 60 min of ischemia via percutaneous occlusion of the LAD. After 55 min of ischemia control eEPCs or cells transfected with thymosin β-4 (T-β-4) shRNA or T-β-4 alone were infused retrograde into the anterior interventricular vein. These studies demonstrate that the short-term cardioprotection resulting from retrograde infusion of eEPCs is, at least partially, due to T-β-4.
References
Hoit BD, Kiatchoosakun S, Restivo J, et al. Naturally occurring variation in cardiovascular traits among inbred mouse strains. Genomics. 2002;79:679–685.
Cowan PJ, Shinkel TA, Fisicaro N, et al. Targeting gene expression to endothelium in transgenic animals: A comparison of the human ICAM-2, PECAM-1 and endoglin promoters. Xenotransplantation. 2003;10:223–231.
Singbartl K, Thatte J, Smith ML, Wethmar K, Day K, Ley K. A CD2-green fluorescence protein-transgenic mouse reveals very late antigen-4-dependent CD8+ lymphocyte rolling in inflamed venules. J Immunol. 2001;166:7520–7526.
Placier S, Boffa JJ, Dussaule JC, Chatziantoniou C. Reversal of renal lesions following interruption of nitric oxide synthesis inhibition in transgenic mice. Nephrol Dial Transplant. 2006;21:881–888.
Pichler A, Prior JL, Luker GD, Piwnica-Worms D. Generation of a highly inducible Gal4®Fluc universal reporter mouse for in vivo bioluminescence imaging. Proc Natl Acad Sci USA. 2008;105:15932–15937.
Viswanathan S, Burch JB, Fishman GI, Moskowitz IP, Benson DW. Characterization of sinoatrial node in four conduction system marker mice. J Mol Cell Cardiol. 2007;42:946–953.
Zhai P, Eurell TE, Cotthaus R, Jeffery EH, Bahr JM, Gross DR. Effect of estrogen on global myocardial ischemia-reperfusion injury in female rats. Am J Physiol Heart Circ Physiol. 2000;279:H2766–H2775.
Zhai P, Eurell TE, Cotthaus RP, Jeffery EH, Bahr JM, Gross DR. Effects of dietary phytoestrogen on global myocardial ischemia-reperfusion injury in isolated female rat hearts. Am J Physiol Heart Circ Physiol. 2001;281:H1223–H1232.
Arias-Loza PA, Jazbutyte V, Pelzer T. Genetic and pharmacologic strategies to determine the function of estrogen receptor alpha and estrogen receptor beta in cardiovascular system. Gend Med. 2008;5 Suppl A:S34–S45.
Seli E, Guzeloglu-Kayisli O, Kayisli UA, Kizilay G, Arici A. Estrogen increases apoptosis in the arterial wall in a murine atherosclerosis model. Fertil Steril. 2007;88:1190–1196.
Simpson ER, Jones ME. Of mice and men: The many guises of estrogens. Ernst Schering Found Symp Proc. 2006;1:45–67.
Sun D, Yan C, Jacobson A, Jiang H, Carroll MA, Huang A. Contribution of epoxyeicosatrienoic acids to flow-induced dilation in arteries of male ERalpha knockout mice: Role of aromatase. Am J Physiol Regul Integr Comp Physiol. 2007;293:R1239–R1246.
Zhai P, Eurell TE, Cooke PS, Lubahn DB, Gross DR. Myocardial ischemia-reperfusion injury in estrogen receptor-alpha knockout and wild-type mice. Am J Physiol Heart Circ Physiol. 2000;278:H1640–H1647.
Huss JM, Imahashi K, Dufour CR, et al. The nuclear receptor ERRα is required for the bioenergetic and functional adaptation to cardiac pressure overload. Cell Metab. 2007;6:25–37.
Corbacho AM, Eiserich JP, Zuniga LA, Valacchi G, Villablanca AC. Compromised aortic vasoreactivity in male estrogen receptor-alpha-deficient mice during acute lipopolysaccharide-induced inflammation. Endocrinology. 2007;148:1403–1411.
Babiker FA, Lips D, Meyer R, et al. Estrogen receptor beta protects the murine heart against left ventricular hypertrophy. Arterioscler Thromb Vasc Biol. 2006;26:1524–1530.
Pedram A, Razandi M, Lubahn D, Liu J, Vannan M, Levin ER. Estrogen inhibits cardiac hypertrophy: Role of estrogen receptor-beta to inhibit calcineurin. Endocrinology. 2008;149:3361–3369.
Foryst-Ludwig A, Clemenz M, Hohmann S, et al Metabolic actions of estrogen receptor beta (ERbeta) are mediated by a negative cross-talk with PPARgamma. PLoS Genet. 2008;4:e1000108.
Wang M, Crisostomo PR, Markel T, Wang Y, Lillemoe KD, Meldrum DR. Estrogen receptor beta mediates acute myocardial protection following ischemia. Surgery. 2008;144:233–238.
Pelzer T, Loza PA, Hu K, et al. Increased mortality and aggravation of heart failure in estrogen receptor-beta knockout mice after myocardial infarction. Circulation. 2005;111:1492–1498.
Ullrich ND, Krust A, Collins P, MacLeod KT. Genomic deletion of estrogen receptors ERalpha and ERbeta does not alter estrogen-mediated inhibition of Ca2+ influx and contraction in murine cardiomyocytes. Am J Physiol Heart Circ Physiol. 2008;294:H2421–H2427.
O’Lone R, Knorr K, Jaffe IZ, et al. Estrogen receptors alpha and beta mediate distinct pathways of vascular gene expression, including genes involved in mitochondrial electron transport and generation of reactive oxygen species. Mol Endocrinol. 2007;21:1281–1296.
Luksha L, Poston L, Gustafsson JA, Aghajanova L, Kublickiene K. Gender-specific alteration of adrenergic responses in small femoral arteries from estrogen receptor-beta knockout mice. Hypertension. 2005;46:1163–1168.
Luksha L, Poston L, Gustafsson JA, Hultenby K, Kublickiene K. The oestrogen receptor beta contributes to sex related differences in endothelial function of murine small arteries via EDHF. J Physiol. 2006;577:945–955.
Cruz MN, Douglas G, Gustafsson JA, Poston L, Kublickiene K. Dilatory responses to estrogenic compounds in small femoral arteries of male and female estrogen receptor-beta knockout mice. Am J Physiol Heart Circ Physiol. 2006;290:H823–H829.
Douglas G, Cruz MN, Poston L, Gustafsson JA, Kublickiene K. Functional characterization and sex differences in small mesenteric arteries of the estrogen receptor-beta knockout mouse. Am J Physiol Regul Integr Comp Physiol. 2008;294:R112–R120.
Tsutsumi S, Zhang X, Takata K, et al. Differential regulation of the inducible nitric oxide synthase geneby estrogen receptors {alpha}and {beta}. J Endocrinol. 2008 Nov; 199(2):267-273.
Umetani M, Domoto H, Gormley AK, et al. 27-Hydroxycholesterol is an endogenous SERM that inhibits the cardiovascular effects of estrogen. Nat Med. 2007;13:1185–1192.
Belo NO, Sairam MR, Dos Reis AM. Impairment of the natriuretic peptide system in follitropin receptor knockout mice and reversal by estradiol: Implications for obesity-associated hypertension in menopause. Endocrinology. 2008;149:1399–1406.
Ebrahimian T, Sairam MR, Schiffrin EL, Touyz RM. Cardiac hypertrophy is associated with altered thioredoxin and ASK1 signaling in a mouse model of menopause. Am J Physiol Heart Circ Physiol. 2008.
Matsui Y, Nakano N, Shao D, et al. Lats2 is a negative regulator of myocyte size in the heart. Circ Res. 2008;103:1309–1318.
Cohn HI, Harris DM, Pesant S, et al. Inhibition of vascular smooth muscle G protein-coupled receptor kinase 2 enhances {alpha}1DAR constriction. Am J Physiol Heart Circ Physiol. 2008;295:H1695–H1704.
Yu X, Huang S, Patterson E, et al. Proteasome degradation of GRK2 during ischemia and ventricular tachyarrhythmias in a canine model of myocardial infarction. Am J Physiol Heart Circ Physiol. 2005;289:H1960–H1967.
Iaccarino G, Rockman HA, Shotwell KF, Tomhave ED, Koch WJ. Myocardial overexpression of GRK3 in transgenic mice: Evidence for in vivo selectivity of GRKs. Am J Physiol. 1998;275:H1298–H1306.
Hirotani S, Zhai P, Tomita H, et al. Inhibition of glycogen synthase kinase 3β during heart failure is protective. Circ Res. 2007;101:1164–1174.
Luckey SW, Mansoori J, Fair K, Antos CL, Olson EN, Leinwand LA. Blocking cardiac growth in hypertrophic cardiomyopathy induces cardiac dysfunction and decreased survival only in males. Am J Physiol Heart Circ Physiol. 2007;292:H838–H845.
Korshunov VA, Daul M, Massett MP, Berk BC. Axl mediates vascular remodeling induced by deoxycorticosterone acetate-salt hypertension. Hypertension. 2007;50:1057–1062.
Rexhepaj R, Boini KM, Huang DY, et al. Role of maternal glucocorticoid inducible kinase SGK1 in fetal programming of blood pressure in response to prenatal diet. Am J Physiol Regul Integr Comp Physiol. 2008;294:R2008–R2013.
Mayr M, Metzler B, Chung YL, et al. Ischemic preconditioning exaggerates cardiac damage in PKC-delta null mice. Am J Physiol Heart Circ Physiol. 2004;287:H946–H956.
Huang J, Shelton JM, Richardson JA, Kamm KE, Stull JT. Myosin regulatory light chain phosphorylation attenuates cardiac hypertrophy. J Biol Chem. 2008;283:19748–19756.
Ofir M, Arad M, Porat E, et al. Increased glycogen stores due to gamma-AMPK overexpression protects against ischemia and reperfusion damage. Biochem Pharmacol. 2008;75:1482–1491.
Tsukada S, Iwai M, Nishiu J, et al. Inhibition of experimental intimal thickening in mice lacking a novel G-protein-coupled receptor. Circulation. 2003;107:313–319.
McMullen JR, Amirahmadi F, Woodcock EA, et al. Protective effects of exercise and phosphoinositide 3-kinase(p110alpha) signaling in dilated and hypertrophic cardiomyopathy. Proc Natl Acad Sci USA. 2007;104:612–617.
Fujii A, Nakano D, Katsuragi M, et al. Role of gp91phox-containing NADPH oxidase in the deoxycorticosterone acetate-salt-induced hypertension. Eur J Pharmacol. 2006;552:131–134.
Salguero G, Akin E, Templin C, et al. Renovascular hypertension by two-kidney one-clip enhances endothelial progenitor cell mobilization in a p47phox-dependent manner. J Hypertens. 2008;26:257–268.
Frantz S, Brandes RP, Hu K, et al. Left ventricular remodeling after myocardial infarction in mice with targeted deletion of the NADPH oxidase subunit gp91PHOX. Basic Res Cardiol. 2006;101:127–132.
Wiesel P, Patel AP, Carvajal IM, et al. Exacerbation of chronic renovascular hypertension and acute renal failure in heme oxygenase-1-deficient mice. Circ Res. 2001;88:1088–1094.
Nath KA, d’Uscio LV, Juncos JP, et al. An analysis of the DOCA-salt model of hypertension in HO-1−/− mice and the Gunn rat. Am J Physiol Heart Circ Physiol. 2007;293:H333–H342.
Yaar R, Jones MR, Chen JF, Ravid K. Animal models for the study of adenosine receptor function. J Cell Physiol. 2005;202:9–20.
Tikh EI, Fenton RA, Dobson JG, Jr. Contractile effects of adenosine A1 and A2A receptors in isolated murine hearts. Am J Physiol Heart Circ Physiol. 2006;290:H348–H356.
Tang T, Lai NC, Roth DM, et al. Adenylyl cyclase type V deletion increases basal left ventricular function and reduces left ventricular contractile responsiveness to beta-adrenergic stimulation. Basic Res Cardiol. 2006;101:117–126.
Egli RE, Kash TL, Choo K, et al. Norepinephrine modulates glutamatergic transmission in the bed nucleus of the stria terminalis. Neuropsychopharmacology. 2005;30:657–668.
Ihalainen JA, Tanila H. In vivo regulation of dopamine and noradrenaline release by alpha2A-adrenoceptors in the mouse nucleus accumbens. J Neurochem. 2004;91:49–56.
Zhu QM, Lesnick JD, Jasper JR, et al. Cardiovascular effects of rilmenidine, moxonidine and clonidine in conscious wild-type and D79N alpha2A-adrenoceptor transgenic mice. Br J Pharmacol. 1999;126:1522–1530.
Sallinen J, Haapalinna A, MacDonald E, et al. Genetic alteration of the alpha2-adrenoceptor subtype c in mice affects the development of behavioral despair and stress-induced increases in plasma corticosterone levels. Mol Psychiatry. 1999;4:443–452.
Rockman HA, Choi DJ, Akhter SA, et al. Control of myocardial contractile function by the level of beta-adrenergic receptor kinase 1 in gene-targeted mice. J Biol Chem. 1998;273:18180–18184.
Cristina C, Diaz-Torga G, Baldi A, et al. Increased pituitary vascular endothelial growth factor-a in dopaminergic D2 receptor knockout female mice. Endocrinology. 2005;146:2952–2962.
Tavernier G, Jimenez M, Giacobino JP, et al. Norepinephrine induces lipolysis in beta1/beta2/beta3-adrenoceptor knockout mice. Mol Pharmacol. 2005;68:793–799.
Kurihara N, Alfie ME, Sigmon DH, Rhaleb NE, Shesely EG, Carretero OA. Role of nNOS in blood pressure regulation in eNOS null mutant mice. Hypertension. 1998;32:856–861.
Duplain H, Burcelin R, Sartori C, et al. Insulin resistance, hyperlipidemia, and hypertension in mice lacking endothelial nitric oxide synthase. Circulation. 2001;104:342–345.
Grasselli A, Nanni S, Colussi C, et al. Estrogen receptor-alpha and endothelial nitric oxide synthase nuclear complex regulates transcription of human telomerase. Circ Res. 2008;103:34–42.
Billon A, Lehoux S, Lam Shang Leen L, et al. The estrogen effects on endothelial repair and mitogen-activated protein kinase activation are abolished in endothelial nitric-oxide (NO) synthase knockout mice, but not by NO synthase inhibition by N-nitro-l-arginine methyl ester. Am J Pathol. 2008;172:830–838.
Takenaka K, Nishimura Y, Nishiuma T, et al. Ventilator-induced lung injury is reduced in transgenic mice that overexpress endothelial nitric oxide synthase. Am J Physiol Lung Cell Mol Physiol. 2006;290:L1078–L1086.
Gava AL, Peotta VA, Cabral AM, Vasquez EC, Meyrelles SS. Overexpression of eNOS prevents the development of renovascular hypertension in mice. Can J Physiol Pharmacol. 2008;86:458–464.
Hao YH, Yong HY, Murphy CN, et al. Production of endothelial nitric oxide synthase (eNOS) over-expressing piglets. Transgenic Res. 2006;15:739–750.
Kadowaki T, Yamauchi T. Adiponectin and adiponectin receptors. Endocr Rev. 2005;26:439–451.
Otabe S, Yuan X, Fukutani T, et al. Overexpression of human adiponectin in transgenic mice results in suppression of fat accumulation and prevention of premature death by high-calorie diet. Am J Physiol Endocrinol Metab. 2007;293:E210–E218.
Ohashi K, Iwatani H, Kihara S, et al. Exacerbation of albuminuria and renal fibrosis in subtotal renal ablation model of adiponectin-knockout mice. Arterioscler Thromb Vasc Biol. 2007;27:1910–1917.
Bauche IB, El Mkadem SA, Pottier AM, et al. Overexpression of adiponectin targeted to adipose tissue in transgenic mice: Impaired adipocyte differentiation. Endocrinology. 2007;148:1539–1549.
Shinmura K, Tamaki K, Saito K, Nakano Y, Tobe T, Bolli R. Cardioprotective effects of short-term caloric restriction are mediated by adiponectin via activation of AMP-activated protein kinase. Circulation. 2007;116:2809–2817.
Lin HV, Kim JY, Pocai A, et al. Adiponectin resistance exacerbates insulin resistance in insulin receptor transgenic/knockout mice. Diabetes. 2007;56:1969–1976.
Carvalho E, Kotani K, Peroni OD, Kahn BB. Adipose-specific overexpression of GLUT4 reverses insulin resistance and diabetes in mice lacking GLUT4 selectively in muscle. Am J Physiol Endocrinol Metab. 2005;289:E551–E561.
Hertzel AV, Smith LA, Berg AH, et al. Lipid metabolism and adipokine levels in fatty acid-binding protein null and transgenic mice. Am J Physiol Endocrinol Metab. 2006;290:E814–E823.
Jurczak MJ, Danos AM, Rehrmann VR, Allison MB, Greenberg CC, Brady MJ. Transgenic overexpression of protein targeting to glycogen markedly increases adipocytic glycogen storage in mice. Am J Physiol Endocrinol Metab. 2007;292:E952–E963.
Yamaguchi M, Nakagawa T. Change in lipid components in the adipose and liver tissues of regucalcin transgenic rats with increasing age: Suppression of leptin and adiponectin gene expression. Int J Mol Med. 2007;20:323–328.
Huang J, Gabrielsen JS, Cooksey RC, et al. Increased glucose disposal and AMP-dependent kinase signaling in a mouse model of hemochromatosis. J Biol Chem. 2007;282:37501–37507.
Al-Aly Z, Shao JS, Lai CF, et al. Aortic Msx2-wnt calcification cascade is regulated by TNF-alpha-dependent signals in diabetic ldlr−/− mice. Arterioscler Thromb Vasc Biol. 2007;27:2589–2596.
Ye Z, Ahmed KA, Hao S, et al. Active CD4+ helper T cells directly stimulate CD8+ cytotoxic T lymphocyte responses in wild-type and MHC II gene knockout C57BL/6 mice and transgenic RIP-mOVA mice expressing islet beta-cell ovalbumin antigen leading to diabetes. Autoimmunity. 2008;41:501–511.
Costa C, Zhao L, Burton WV, et al. Transgenic pigs designed to express human CD59 and H-transferase to avoid humoral xenograft rejection. Xenotransplantation. 2002;9:45–57.
Smolenski RT, Forni M, Maccherini M, et al. Reduction of hyperacute rejection and protection of metabolism and function in hearts of human decay accelerating factor (hDAF)-expressing pigs. Cardiovasc Res. 2007;73:143–152.
Lee JM, Tu CF, Tai HC, et al. The hDAF exogene protects swine endothelial and peripheral blood mononuclear cells from xenoreactive antibody mediated cytotoxicity in hDAF transgenic pigs. Transplant Proc. 2006;38:2270–2272.
Brandl U, Erhardt M, Michel S, et al. Soluble galalpha(1,3)gal conjugate combined with hDAF preserves morphology and improves function of cardiac xenografts. Xenotransplantation. 2007;14:323–332.
Brandl U, Jockle H, Erhardt M, et al. Reduced fibrin deposition and intravascular thrombosis in hDAF transgenic pig hearts perfused with tirofiban. Transplantation. 2007;84:1667–1676.
Warnecke G, Severson SR, Ugurlu MM, et al. Endothelial function in pigs transgenic for human complement regulating factor. Transplantation. 2002;73:1060–1067.
Zhou CY, McInnes E, Parsons N, et al. Production and characterization of a pig line transgenic for human membrane cofactor protein. Xenotransplantation. 2002;9:183–190.
Grabie N, Hsieh DT, Buono C, et al. Neutrophils sustain pathogenic CD8+ T cell responses in the heart. Am J Pathol. 2003;163:2413–2420.
Adachi O, Yamato E, Kawamoto S, et al. High-level expression of viral interleukin-10 in cardiac allografts fails to prolong graft survival. Transplantation. 2002;74:1603–1608.
Klugewitz K, Blumenthal-Barby F, Schrage A, Knolle PA, Hamann A, Crispe IN. Immunomodulatory effects of the liver: Deletion of activated CD4+ effector cells and suppression of IFN-gamma-producing cells after intravenous protein immunization. J Immunol. 2002;169:2407–2413.
Fischbein MP, Yun J, Laks H, et al. Regulated interleukin-10 expression prevents chronic rejection of transplanted hearts. J Thorac Cardiovasc Surg. 2003;126:216–223.
Takeuchi T, Ueki T, Sunaga S, et al. Murine interleukin 4 transgenic heart allograft survival prolonged with down-regulation of the Th1 cytokine mRNA in grafts. Transplantation. 1997;64:152–157.
Rothermel AL, Wang Y, Schechner J, et al. Endothelial cells present antigens in vivo. BMC Immunol. 2004;5:5.
Reuter H, Philipson KD. Sodium-calcium exchanger overexpression in the heart - Insights from a transgenic mouse model. Basic Res Cardiol. 2002;97 Suppl 1:131–135.
Muller JG, Isomatsu Y, Koushik SV, et al. Cardiac-specific expression and hypertrophic upregulation of the feline Na(+)-Ca(2+) exchanger gene H1-promoter in a transgenic mouse model. Circ Res. 2002;90:158–164.
Maxwell K, Scott J, Omelchenko A, et al. Functional role of ionic regulation of Na+/Ca2+ exchange assessed in transgenic mouse hearts. Am J Physiol. 1999;277:H2212–H2221.
Weber CR, Ginsburg KS, Philipson KD, Shannon TR, Bers DM. Allosteric regulation of Na/Ca exchange current by cytosolic Ca in intact cardiac myocytes. J Gen Physiol. 2001;117:119–131.
Yao A, Su Z, Nonaka A, et al. Effects of overexpression of the Na+-Ca2+ exchanger on [Ca2+]i transients in murine ventricular myocytes. Circ Res. 1998;82:657–665.
Cho CH, Lee SY, Shin HS, Philipson KD, Lee CO. Partial rescue of the Na+-Ca2+ exchanger (NCX1) knock-out mouse by transgenic expression of NCX1. Exp Mol Med. 2003;35:125–135.
Karashima E, Nishimura J, Iwamoto T, et al. Involvement of Na+-Ca2+ exchanger in cAMP-mediated relaxation in mice aorta: Evaluation using transgenic mice. Br J Pharmacol. 2007;150:434–444.
Su Z, Bridge JH, Philipson KD, Spitzer KW, Barry WH. Quantitation of Na/Ca exchanger function in single ventricular myocytes. J Mol Cell Cardiol. 1999;31:1125–1135.
Nakamura TY, Iwata Y, Arai Y, Komamura K, Wakabayashi S. Activation of Na+/H+ exchanger 1 is sufficient to generate Ca2+ signals that induce cardiac hypertrophy and heart failure. Circ Res. 2008;103:891–899.
Dibbs ZI, Diwan A, Nemoto S, et al. Targeted overexpression of transmembrane tumor necrosis factor provokes a concentric cardiac hypertrophic phenotype. Circulation. 2003;108:1002–1008.
Huber SA, Feldman AM, Sartini D. Coxsackievirus B3 induces T regulatory cells, which inhibit cardiomyopathy in tumor necrosis factor-alpha transgenic mice. Circ Res. 2006;99:1109–1116.
Mori S, Gibson G, McTiernan CF. Differential expression of MMPs and TIMPs in moderate and severe heart failure in a transgenic model. J Card Fail. 2006;12:314–325.
Sawaya SE, Rajawat YS, Rami TG, et al. Downregulation of connexin40 and increased prevalence of atrial arrhythmias in transgenic mice with cardiac-restricted overexpression of tumor necrosis factor. Am J Physiol Heart Circ Physiol. 2007;292:H1561–H1567.
Sekiguchi K, Tian Q, Ishiyama M, et al. Inhibition of PPAR-alpha activity in mice with cardiac-restricted expression of tumor necrosis factor: Potential role of TGF-beta/Smad3. Am J Physiol Heart Circ Physiol. 2007;292:H1443–H1451.
Panagopoulou P, Davos CH, Milner DJ, et al. Desmin mediates TNF-alpha-induced aggregate formation and intercalated disk reorganization in heart failure. J Cell Biol. 2008;181:761–775.
Sakata Y, Chancey AL, Divakaran VG, Sekiguchi K, Sivasubramanian N, Mann DL. Transforming growth factor-beta receptor antagonism attenuates myocardial fibrosis in mice with cardiac-restricted overexpression of tumor necrosis factor. Basic Res Cardiol. 2008;103:60–68.
Pinderski Oslund LJ, Hedrick CC, Olvera T, et al. Interleukin-10 blocks atherosclerotic events in vitro and in vivo. Arterioscler Thromb Vasc Biol. 1999;19:2847–2853.
Pinderski LJ, Fischbein MP, Subbanagounder G, et al. Overexpression of interleukin-10 by activated T lymphocytes inhibits atherosclerosis in LDL receptor-deficient mice by altering lymphocyte and macrophage phenotypes. Circ Res. 2002;90:1064–1071.
Willuweit A, Sass G, Schoneberg A, Eisel U, Tiegs G, Clauss M. Chronic inflammation and protection from acute hepatitis in transgenic mice expressing TNF in endothelial cells. J Immunol. 2001;167:3944–3952.
Reifenberg K, Lehr HA, Torzewski M, et al. Interferon-gamma induces chronic active myocarditis and cardiomyopathy in transgenic mice. Am J Pathol. 2007;171:463–472.
Arsenijevic D, Garcia I, Vesin C, et al. Differential roles of tumor necrosis factor-alpha and interferon-gamma in mouse hypermetabolic and anorectic responses induced by LPS. Eur Cytokine Netw. 2000;11:662–668.
Monraats PS, Pires NM, Schepers A, et al. Tumor necrosis factor-alpha plays an important role in restenosis development. FASEB J. 2005;19:1998–2004.
Metzler B, Mair J, Lercher A, et al. Mouse model of myocardial remodelling after ischemia: Role of intercellular adhesion molecule-1. Cardiovasc Res. 2001;49:399–407.
Metzler B, Haubner B, Conci E, et al. Myocardial ischaemia-reperfusion injury in haematopoietic cell-restricted {beta}1 integrin knockout mice. Exp Physiol. 2008;93:825–833.
Son NH, Park TS, Yamashita H, et al. Cardiomyocyte expression of PPARgamma leads to cardiac dysfunction in mice. J Clin Invest. 2007;117:2791–2801.
Cresci S, Jones PG, Sucharov CC, et al. Interaction between PPARA genotype and beta-blocker treatment influences clinical outcomes following acute coronary syndromes. Pharmacogenomics. 2008;9:1403–1417.
Marionneau C, Aimond F, Brunet S, et al. PPARalpha-mediated remodeling of repolarizing voltage-gated K+ (kv) channels in a mouse model of metabolic cardiomyopathy. J Mol Cell Cardiol. 2008;44:1002–1015.
Zhou Y, Luo P, Chang HH, et al. Colfibrate attenuates blood pressure and sodium retention in DOCA-salt hypertension. Kidney Int. 2008;74:1040–1048.
Obih P, Oyekan AO. Regulation of blood pressure, natriuresis and renal thiazide/amiloride sensitivity in PPARalpha null mice. Blood Press. 2008;17:55–63.
Odegaard JI, Ricardo-Gonzalez RR, Goforth MH, et al. Macrophage-specific PPARgamma controls alternative activation and improves insulin resistance. Nature. 2007;447:1116–1120.
Murtaza I, Wang HX, Feng X, et al. Down-regulation of catalase and oxidative modification of protein kinase CK2 lead to the failure of apoptosis repressor with caspase recruitment domain to inhibit cardiomyocyte hypertrophy. J Biol Chem. 2008;283:5996–6004.
Liu F, Wei CC, Wu SJ, et al. Apocynin attenuates tubular apoptosis and tubulointerstitial fibrosis in transgenic mice independent of hypertension. Kidney Int. 2009;75:156–166.
Park JK, Fischer R, Dechend R, et al. p38 Mitogen-activated protein kinase inhibition ameliorates angiotensin II-induced target organ damage. Hypertension. 2007;49:481–489.
Cervenka L, Horacek V, Vaneckova I, et al. Essential role of AT1A receptor in the development of 2K1C hypertension. Hypertension. 2002;40:735–741.
Cervenka L, Vaneckova I, Huskova Z, et al. Pivotal role of angiotensin II receptor subtype 1A in the development of two-kidney, one-clip hypertension: Study in angiotensin II receptor subtype 1A knockout mice. J Hypertens. 2008;26:1379–1389.
Osorio JC, Cheema FH, Martens TP, et al. Simvastatin reverses cardiac hypertrophy caused by disruption of the bradykinin 2 receptor. Can J Physiol Pharmacol. 2008;86:633–642.
Rhaleb NE, Peng H, Alfie ME, Shesely EG, Carretero OA. Effect of ACE inhibitor on DOCA-salt- and aortic coarctation-induced hypertension in mice: Do kinin B2 receptors play a role? Hypertension. 1999;33:329–334.
Madeddu P, Milia AF, Salis MB, et al. Renovascular hypertension in bradykinin B2-receptor knockout mice. Hypertension. 1998;32:503–509.
Nogueira BV, Peotta VA, Meyrelles SS, Vasquez EC. Evaluation of aortic remodeling in apolipoprotein E-deficient mice and renovascular hypertensive mice. Arch Med Res. 2007;38:816–821.
Heo HJ, Yun MR, Jung KH, et al. Endogenous angiotensin II enhances atherogenesis in apoprotein E-deficient mice with renovascular hypertension through activation of vascular smooth muscle cells. Life Sci. 2007;80:1057–1063.
Mazzolai L, Korber M, Bouzourene K, et al. Severe hyperlipidemia causes impaired renin-angiotensin system function in apolipoprotein E deficient mice. Atherosclerosis. 2006;186:86–91.
Basso F, Amar MJ, Wagner EM, et al. Enhanced ABCG1 expression increases atherosclerosis in LDLr-KO mice on a western diet. Biochem Biophys Res Commun. 2006;351:398–404.
Watari Y, Yamamoto Y, Brydun A, et al. Ablation of the bach1 gene leads to the suppression of atherosclerosis in bach1 and apolipoprotein E double knockout mice. Hypertens Res. 2008;31:783–792.
Liu H, Zheng F, Li Z, et al. Reduced acute vascular injury and atherosclerosis in hyperlipidemic mice transgenic for lysozyme. Am J Pathol. 2006;169:303–313.
Oyama J, Blais C, Jr, Liu X, et al. Reduced myocardial ischemia-reperfusion injury in toll-like receptor 4-deficient mice. Circulation. 2004;109:784–789.
Riad A, Jager S, Sobirey M, et al. Toll-like receptor-4 modulates survival by induction of left ventricular remodeling after myocardial infarction in mice. J Immunol. 2008;180:6954–6961.
Shishido T, Nozaki N, Yamaguchi S, et al. Toll-like receptor-2 modulates ventricular remodeling after myocardial infarction. Circulation. 2003;108:2905–2910.
Shishido T, Nozaki N, Takahashi H, et al. Central role of endogenous toll-like receptor-2 activation in regulating inflammation, reactive oxygen species production, and subsequent neointimal formation after vascular injury. Biochem Biophys Res Commun. 2006;345:1446–1453.
Papanikolaou A, Papafotika A, Murphy C, et al. Cholesterol-dependent lipid assemblies regulate the activity of the ecto-nucleotidase CD39. J Biol Chem. 2005;280:26406–26414.
Bauer PM, Yu J, Chen Y, et al. Endothelial-specific expression of caveolin-1 impairs microvascular permeability and angiogenesis. Proc Natl Acad Sci USA. 2005;102:204–209.
Odening KE, Hyder O, Chaves L, et al. Pharmacogenomics of anesthetic drugs in transgenic LQT1 and LQT2 rabbits reveal genotype-specific differential effects on cardiac repolarization. Am J Physiol Heart Circ Physiol. 2008;295:H2264–H2272.
Brunner M, Peng X, Liu GX, et al. Mechanisms of cardiac arrhythmias and sudden death in transgenic rabbits with long QT syndrome. J Clin Invest. 2008;118:2246–2259.
Koren G. Electrical remodeling and arrhythmias in long-QT syndrome: Lessons from genetic models in mice. Ann Med. 2004;36 Suppl 1:22–27.
Killeen MJ, Thomas G, Sabir IN, Grace AA, Huang CL. Mouse models of human arrhythmia syndromes. Acta Physiol (Oxf). 2008;192:455–469.
Tillmanns J, Carlsen H, Blomhoff R, et al. Caught in the act: In vivo molecular imaging of the transcription factor NF-kappaB after myocardial infarction. Biochem Biophys Res Commun. 2006;342:773–774.
Frantz S, Hu K, Bayer B, et al. Absence of NF-kappaB subunit p50 improves heart failure after myocardial infarction. FASEB J. 2006;20:1918–1920.
Frantz S, Tillmanns J, Kuhlencordt PJ, et al. Tissue-specific effects of the nuclear factor kappaB subunit p50 on myocardial ischemia-reperfusion injury. Am J Pathol. 2007;171:507–512.
Ramana KV, Willis MS, White MD, et al. Endotoxin-induced cardiomyopathy and systemic inflammation in mice is prevented by aldose reductase inhibition. Circulation. 2006;114:1838–1846.
Henke N, Schmidt-Ullrich R, Dechend R, et al. Vascular endothelial cell-specific NF-kappaB suppression attenuates hypertension-induced renal damage. Circ Res. 2007;101:268–276.
Li HL, Zhuo ML, Wang D, et al. Targeted cardiac overexpression of A20 improves left ventricular performance and reduces compensatory hypertrophy after myocardial infarction. Circulation. 2007;115:1885–1894.
Dufour CR, Wilson BJ, Huss JM, et al. Genome-wide orchestration of cardiac functions by the orphan nuclear receptors ERRalpha and gamma. Cell Metab. 2007;5:345–356.
Alaynick WA, Kondo RP, Xie W, et al. ERRgamma directs and maintains the transition to oxidative metabolism in the postnatal heart. Cell Metab. 2007;6:13–24.
Nosek TM, Brotto MA, Jin JP. Troponin T isoforms alter the tolerance of transgenic mouse cardiac muscle to acidosis. Arch Biochem Biophys. 2004;430:178–184.
Takimoto E, Soergel DG, Janssen PM, Stull LB, Kass DA, Murphy AM. Frequency- and afterload-dependent cardiac modulation in vivo by troponin I with constitutively active protein kinase A phosphorylation sites. Circ Res. 2004;94:496–504.
Pena JR, Wolska BM. Troponin I phosphorylation plays an important role in the relaxant effect of beta-adrenergic stimulation in mouse hearts. Cardiovasc Res. 2004;61:756–763.
Angelone T, Quintieri AM, Brar BK, et al. The antihypertensive chromogranin A-derived peptide catestatin as a novel endocrine/paracrine modulator of cardiac function: Inotropic and lusitropic actions on the rat heart. Endocrinology. 2008 Oct; 149(10):4780-4793.
Mahapatra NR. Catestatin is a novel endogenous peptide that regulates cardiac function and blood pressure. Cardiovasc Res. 2008;80:330–338.
Hu C, Dandapat A, Sun L, et al. Modulation of angiotensin II-mediated hypertension and cardiac remodeling by lectin-like oxidized low-density lipoprotein receptor-1 deletion. Hypertension. 2008;52:556–562.
Adamek A, Jung S, Dienesch C, et al. Role of 5-lipoxygenase in myocardial ischemia-reperfusion injury in mice. Eur J Pharmacol. 2007;571:51–54.
Kirchhefer U, Baba HA, Hanske G, et al. Age-dependent biochemical and contractile properties in atrium of transgenic mice overexpressing junctin. Am J Physiol Heart Circ Physiol. 2004;287:H2216–H2225.
Kirchhefer U, Hanske G, Jones LR, et al. Overexpression of junctin causes adaptive changes in cardiac myocyte Ca(2+) signaling. Cell Calcium. 2006;39:131–142.
Zhang L, Franzini-Armstrong C, Ramesh V, Jones LR. Structural alterations in cardiac calcium release units resulting from overexpression of junctin. J Mol Cell Cardiol. 2001;33:233–247.
Kirchhefer U, Klimas J, Baba HA, et al. Triadin is a critical determinant of cellular Ca cycling and contractility in the heart. Am J Physiol Heart Circ Physiol. 2007;293:H3165–H3174.
Hagendorff A, Schumacher B, Kirchhoff S, Luderitz B, Willecke K. Conduction disturbances and increased atrial vulnerability in Connexin40-deficient mice analyzed by transesophageal stimulation. Circulation. 1999;99:1508–1515.
Hagendorff A, Kirchhoff S, Kruger O, et al. Electrophysiological characterization of connexin 40 deficient hearts - In vivo studies in mice. Z Kardiol. 2001;90:898–905.
Hanner F, Schnichels M, Zheng-Fischhofer Q, et al. Connexin 30.3 is expressed in the kidney but not regulated by dietary salt or high blood pressure. Cell Commun Adhes. 2008;15:219–230.
Hanner F, von Maltzahn J, Maxeiner S, et al. Connexin45 is expressed in the juxtaglomerular apparatus and is involved in the regulation of renin secretion and blood pressure. Am J Physiol Regul Integr Comp Physiol. 2008;295:R371–R380.
Wagner C, de Wit C, Gerl M, Kurtz A, Hocherl K. Increased expression of cyclooxygenase 2 contributes to aberrant renin production in connexin 40-deficient kidneys. Am J Physiol Regul Integr Comp Physiol. 2007;293:R1781–R1786.
Krattinger N, Capponi A, Mazzolai L, et al. Connexin40 regulates renin production and blood pressure. Kidney Int. 2007;72:814–822.
Winterhager E, Pielensticker N, Freyer J, et al. Replacement of connexin43 by connexin26 in transgenic mice leads to dysfunctional reproductive organs and slowed ventricular conduction in the heart. BMC Dev Biol. 2007;7:26.
Hattori K, Nakamura K, Hisatomi Y, et al. Arrhythmia induced by spatiotemporal overexpression of calreticulin in the heart. Mol Genet Metab. 2007;91:285–293.
Pattison JS, Waggoner JR, James J, et al. Phospholamban overexpression in transgenic rabbits. Transgenic Res. 2008;17:157–170.
Sag CM, Dybkova N, Neef S, Maier LS. Effects on recovery during acidosis in cardiac myocytes overexpressing CaMKII. J Mol Cell Cardiol. 2007;43:696–709.
Purevjav E, Nelson DP, Varela JJ, et al. Myocardial fas ligand expression increases susceptibility to AZT-induced cardiomyopathy. Cardiovasc Toxicol. 2007;7:255–263.
Niu J, Azfer A, Kolattukudy PE. Protection against lipopolysaccharide-induced myocardial dysfunction in mice by cardiac-specific expression of soluble fas. J Mol Cell Cardiol. 2008;44:160–169.
Niu J, Azfer A, Kolattukudy PE. Monocyte-specific bcl-2 expression attenuates inflammation and heart failure in monocyte chemoattractant protein-1 (MCP-1)-induced cardiomyopathy. Cardiovasc Res. 2006;71:139–148.
Niu J, Azfer A, Deucher MF, Goldschmidt-Clermont PJ, Kolattukudy PE. Targeted cardiac expression of soluble fas prevents the development of heart failure in mice with cardiac-specific expression of MCP-1. J Mol Cell Cardiol. 2006;40:810–820.
Barnes JA, Singh S, Gomes AV. Protease activated receptors in cardiovascular function and disease. Mol Cell Biochem. 2004;263:227–239.
Hamilton JR, Cornelissen I, Coughlin SR. Impaired hemostasis and protection against thrombosis in protease-activated receptor 4-deficient mice is due to lack of thrombin signaling in platelets. J Thromb Haemost. 2004;2:1429–1435.
McGuire JJ, Van Vliet BN, Halfyard SJ. Blood pressures, heart rate and locomotor activity during salt loading and angiotensin II infusion in protease-activated receptor 2 (PAR2) knockout mice. BMC Physiol. 2008;8:20.
Busso N, Chobaz-Peclat V, Hamilton J, Spee P, Wagtmann N, So A. Essential role of platelet activation via protease activated receptor 4 in tissue factor-initiated inflammation. Arthritis Res Ther. 2008;10:R42.
Wu Q, Xu-Cai YO, Chen S, Wang W. Corin: New insights into the natriuretic peptide system. Kidney Int. 2009;75:142–146.
Foronjy RF, Sun J, Lemaitre V, D’Armiento JM. Transgenic expression of matrix metalloproteinase-1 inhibits myocardial fibrosis and prevents the transition to heart failure in a pressure overload mouse model. Hypertens Res. 2008;31:725–735.
Yamada S, Wang KY, Tanimoto A, et al. Matrix metalloproteinase 12 accelerates the initiation of atherosclerosis and stimulates the progression of fatty streaks to fibrous plaques in transgenic rabbits. Am J Pathol. 2008;172:1419–1429.
Janssen A, Hoellenriegel J, Fogarasi M, et al. Abnormal vessel formation in the choroid of mice lacking tissue inhibitor of metalloprotease-3. Invest Ophthalmol Vis Sci. 2008;49:2812–2822.
Holdt LM, Thiery J, Breslow JL, Teupser D. Increased ADAM17 mRNA expression and activity is associated with atherosclerosis resistance in LDL-receptor deficient mice. Arterioscler Thromb Vasc Biol. 2008;28:1097–1103.
Qin Z, Gongora MC, Ozumi K, et al Role of menkes ATPase in angiotensin II-induced hypertension. A key modulator for extracellular superoxide dismutase function. Hypertension. 2008;52:945–951.
Yang A, Sonin D, Jones L, Barry WH, Liang BT. A beneficial role of cardiac P2X4 receptors in heart failure: Rescue of the calsequestrin overexpression model of cardiomyopathy. Am J Physiol Heart Circ Physiol. 2004;287:H1096–H1103.
Angelis E, Tse MY, Adams MA, Pang SC. Effect of AT2 blockade on cardiac hypertrophy as induced by high dietary salt in the proatrial natriuretic peptide (ANP) gene-disrupted mouse. Can J Physiol Pharmacol. 2006;84:625–634.
De Ciuceis C, Amiri F, Brassard P, Endemann DH, Touyz RM, Schiffrin EL. Reduced vascular remodeling, endothelial dysfunction, and oxidative stress in resistance arteries of angiotensin II-infused macrophage colony-stimulating factor-deficient mice: Evidence for a role in inflammation in angiotensin-induced vascular injury. Arterioscler Thromb Vasc Biol. 2005;25:2106–2113.
Shiba Y, Takahashi M, Yoshioka T, et al. M-CSF accelerates neointimal formation in the early phase after vascular injury in mice: The critical role of the SDF-1-CXCR4 system. Arterioscler Thromb Vasc Biol. 2007;27:283–289.
Amiri F, Virdis A, Neves MF, et al. Endothelium-restricted overexpression of human endothelin-1 causes vascular remodeling and endothelial dysfunction. Circulation. 2004;110:2233–2240.
Lund AK, Agbor LN, Zhang N, et al. Loss of the aryl hydrocarbon receptor induces hypoxemia, endothelin-1, and systemic hypertension at modest altitude. Hypertension. 2008;51:803–809.
Dietz HC, Mecham RP. Mouse models of genetic diseases resulting from mutations in elastic fiber proteins. Matrix Biol. 2000;19:481–488.
Faury G, Pezet M, Knutsen RH, et al. Developmental adaptation of the mouse cardiovascular system to elastin haploinsufficiency. J Clin Invest. 2003;112:1419–1428.
Hirano E, Knutsen RH, Sugitani H, Ciliberto CH, Mecham RP. Functional rescue of elastin insufficiency in mice by the human elastin gene: Implications for mouse models of human disease. Circ Res. 2007;101:523–531.
Hou Y, Okada K, Okamoto C, Ueshima S, Matsuo O. Alpha2-antiplasmin is a critical regulator of angiotensin II-mediated vascular remodeling. Arterioscler Thromb Vasc Biol. 2008;28:1257–1262.
Huggins GS, Lepore JJ, Greytak S, et al. The CREB leucine zipper regulates CREB phosphorylation, cardiomyopathy, and lethality in a transgenic model of heart failure. Am J Physiol Heart Circ Physiol. 2007;293:H1877–H1882.
Koonen DP, Febbraio M, Bonnet S, et al. CD36 expression contributes to age-induced cardiomyopathy in mice. Circulation. 2007;116:2139–2147.
Nahrendorf M, Hu K, Frantz S, et al. Factor XIII deficiency causes cardiac rupture, impairs wound healing, and aggravates cardiac remodeling in mice with myocardial infarction. Circulation. 2006;113:1196–1202.
Kuba K, Zhang L, Imai Y, et al Impaired heart contractility in apelin gene-deficient mice associated with aging and pressure overload. Circ Res. 2007;101:e32–e42.
Hatcher CJ, Kim MS, Mah CS, et al. TBX5 transcription factor regulates cell proliferation during cardiogenesis. Dev Biol. 2001;230:177–188.
Isenberg JS, Frazier WA, Krishna MC, Wink DA, Roberts DD. Enhancing cardiovascular dynamics by inhibition of thrombospondin-1/CD47 signaling. Curr Drug Targets. 2008;9:833–841.
Wang YA, Zheng JW, Fei ZL, et al. A novel transgenic mice model for venous malformation. Transgenic Res. 2009 Apr; 18(2):193–201.
Tiedt R, Coers J, Ziegler S, et al. Pronounced thrombocytosis in transgenic mice expressing reduced levels of mpl in platelets and terminally differentiated megakaryocytes. Blood. 2009;113:1768–1777.
van Eeden PE, Tee LB, Lukehurst S, et al. Early vascular and neuronal changes in a VEGF transgenic mouse model of retinal neovascularization. Invest Ophthalmol Vis Sci. 2006;47:4638–4645.
Isoda K, Kamezawa Y, Ayaori M, Kusuhara M, Tada N, Ohsuzu F. Osteopontin transgenic mice fed a high-cholesterol diet develop early fatty-streak lesions. Circulation. 2003;107:679–681.
Ohashi R, Mu H, Wang X, Yao Q, Chen C. Reverse cholesterol transport and cholesterol efflux in atherosclerosis. QJM. 2005;98:845–856.
Rust MB, Faulhaber J, Budack MK, et al. Neurogenic mechanisms contribute to hypertension in mice with disruption of the K-cl cotransporter KCC3. Circ Res. 2006;98:549–556.
Garnier A, Bendall JK, Fuchs S, et al. Cardiac specific increase in aldosterone production induces coronary dysfunction in aldosterone synthase-transgenic mice. Circulation. 2004;110:1819–1825.
Sagave JF, Moser M, Ehler E, et al. Targeted disruption of the mouse Csrp2 gene encoding the cysteine- and glycine-rich LIM domain protein CRP2 result in subtle alteration of cardiac ultrastructure. BMC Dev Biol. 2008;8:80.
Fritsch S, Lindner V, Welsch S, et al. Intravenous delivery of PTH/PTHrP type 1 receptor cDNA to rats decreases heart rate, blood pressure, renal tone, renin angiotensin system, and stress-induced cardiovascular responses. J Am Soc Nephrol. 2004;15:2588–2600.
Bouillon R, Carmeliet G, Verlinden L, et al. Vitamin D and human health: Lessons from vitamin D receptor null mice. Endocr Rev. 2008;29:726–776.
Francois H, Makhanova N, Ruiz P, et al. A role for the thromboxane (tp) receptor in l-NAME hypertension. Am J Physiol Renal Physiol. 2008;295:F1096–F1102.
Guzik TJ, Hoch NE, Brown KA, et al. Role of the T cell in the genesis of angiotensin II induced hypertension and vascular dysfunction. J Exp Med. 2007;204:2449–2460.
Wang Y, Babankova D, Huang J, Swain GM, Wang DH. Deletion of transient receptor potential vanilloid type 1 receptors exaggerates renal damage in deoxycorticosterone acetate-salt hypertension. Hypertension. 2008;52:264–270.
Ni W, Zhou H, Diaz J, Murphy DL, Haywood JR, Watts SW. Lack of the serotonin transporter does not prevent mineralocorticoid hypertension in rat and mouse. Eur J Pharmacol. 2008;589:225–227.
Liao TD, Yang XP, Liu YH, et al. Role of inflammation in the development of renal damage and dysfunction in angiotensin II-induced hypertension. Hypertension. 2008;52:256–263.
Hinkel R, El-Aouni C, Olson T, et al. Thymosin beta4 is an essential paracrine factor of embryonic endothelial progenitor cell-mediated cardioprotection. Circulation. 2008;117:2232–2240.
Author information
Authors and Affiliations
Rights and permissions
Copyright information
© 2009 Springer Science+Business Media, LLC
About this chapter
Cite this chapter
Gross, D.R. (2009). Other Transgenic Animal Models Used in Cardiovascular Studies. In: Animal Models in Cardiovascular Research. Springer, New York, NY. https://doi.org/10.1007/978-0-387-95962-7_14
Download citation
DOI: https://doi.org/10.1007/978-0-387-95962-7_14
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-0-387-95961-0
Online ISBN: 978-0-387-95962-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)