
Abstract
Viruses have evolved elaborate means to regulate diverse cellular pathways in order to create a cellular environment that facilitates viral survival and reproduction. This includes enhancing viral macromolecular synthesis and assembly, as well as preventing antiviral responses, including intrinsic, innate, and adaptive immunity. There are numerous mechanisms by which viruses mediate their effects on the host cell, and this includes targeting various cellular post-translational modification systems, including sumoylation. The wide-ranging impact of sumoylation on cellular processes such as transcriptional regulation, apoptosis, stress response, and cell cycle control makes it an attractive target for viral dysregulation. To date, proteins from both RNA and DNA virus families have been shown to be modified by SUMO conjugation, and this modification appears critical for viral protein function. More interestingly, members of the several viral families have been shown to modulate sumoylation, including papillomaviruses, adenoviruses , herpesviruses, orthomyxoviruses, filoviruses , and picornaviruses . This chapter will focus on mechanisms by which sumoylation both impacts human viruses and is used by viruses to promote viral infection and disease.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
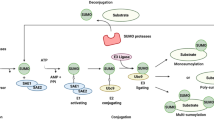
1 Introduction
Viral proteins were among the first defined substrates for sumoylation, beginning with the demonstration in 1999 that the human cytomegalovirus (HCMV) immediate-early 1 protein (IE1) ) was SUMO modified (Muller and Dejean 1999). Among the 7 families of DNA viruses that infect humans, five families (Parvoviridae, Papillomaviridae, Adenoviridae, Herpesviridae, and Poxviridae) have one or more sumoylated proteins, illustrating the widespread utilization of the sumoylation system by nuclear viruses. To date sumoylated proteins have not been reported in the Polyomaviridae and the Hepadnaviridae families, though this may be for lack of investigation and not the absence of substrates for these viruses. Additionally, a number of DNA viral proteins can influence sumoylation globally or for specific substrates, and this is likely to be a critical mechanism to alter the host environment to facilitate the viral life cycle and/or overcome host defenses. DNA viral proteins that are sumoylated and/or which regulate sumoylation are listed in Table 21.1.
Among the RNA viruses infecting humans, sumoylation was first observed for retroviruses which also have a nuclear phase to their life cycle. Specifically, both virion antigens (Gurer et al. 2005; Yueh et al. 2006) and the Tax regulatory protein (Lamsoul et al. 2005) of retroviruses were shown to be substrates for sumoylation. Subsequently, however, sumoylation of viral proteins and/or effects on sumoylation by viral proteins has been observed for nine other human RNA virus families. Many of these RNA viral proteins have no nuclear phase, so cytoplasm ic sumoylation must be occurring as has now been demonstrated for many cellular proteins (Alonso et al. 2015; Hilgarth et al. 2004; Manning Fox et al. 2012). Whether or not sumoylation is relevant for the other families of human RNA viruses remains to be determined, but seems likely given the growing appreciation of the SUMO system as a key target for viral manipulation. RNA viral proteins that are sumoylated or which influence host sumoylation at listed in Table 21.2.
It is clear from numerous studies over the last nearly two decades that sumoylation is an important post-translational process that contributes to successful infection for a wide range of DNA and RNA viruses. In addition to viral proteins being substrates for and regulated by sumoylation, there is now extensive evidence that some viral proteins can act as surrogate sumoylation enzymes and/or modulate the activity of the authentic host sumoylation system. The ability of viral proteins to usurp the sumoylation system is not unexpected as this system contributes to the regulation of many critical cellular pathways and thus provides viruses with mechanisms to exert control over these pathways. Several recent reviews have covered many aspects of viral sumoylation (Everett et al. 2013b; Hannoun et al. 2016; Mattoscio et al. 2013; Varadaraj et al. 2014; Wilson 2012; Wimmer and Schreiner 2015; Wimmer et al. 2012) and should be consulted for more exhaustive coverage of this subject. In this chapter we will provide an overview of the role of sumoylation in human viral families that have been identified to date.
2 DNA Viruses
2.1 Parvoviruses
Parvoviruses have small (approximately 5000 bp), single-stranded DNA genomes with only two genes, one which encodes the capsid proteins and the other which encodes the replication proteins (designated NS or Rep proteins). Adeno associated virus type 2 (AAV-2) is a member of this family whose reproductive cycle is dependent upon co-infection with unrelated helper viruses, typically adenovirus or herpesvirus, and which establishes a latent infection in the absence of the helpers (Berns and Linden 1995). The Rep78 protein of AAV-2 binds the SUMO conjugating enzyme, Ubc9 , and is sumoylated primarily at lysine 84, though other lysines may also serve as weaker acceptor sites (Weger et al. 2004). Converting lysine 84 to the conserved but nonsumoylatable arginine causes a significant decrease in the half-life of Rep78, implicating sumoylation in the control of Rep78 levels. Functionally the effect of sumoylation was not characterized, but sumoylation could be one of the mechanisms that helps regulate Rep to ensure appropriate intracellular levels to maintain the latent state.
AAV based vectors have become widely used in gene therapy due to their ability to infect a wide variety of dividing and nondividing cells (Hastie and Samulski 2015; Lisowski et al. 2015). However, there are numerous impediments to their practical usage, including limited coding capacity, host immune response, and modest transduction efficiency. Recently, Holshcer et al. demonstrated that host sumoylation is one factor that restricts AAV transduction (Holscher et al. 2015). Using a whole genome siRNA screen they found that genes encoding products in the sumoylation pathway, principally SAE1/SAE2 (the activating enzyme subunits) and Ubc9 (the conjugating enzyme), interfered with AAV transduction. The effect appears to be at the level of capsid entry and may involve sumoylation of capsid protein. Most AAV VP1 capsid proteins contain a putative sumoylation motif, though actual modification of VP1 by SUMO was not experimentally tested. Surprisingly, while this effect was observed for several AAV serotypes tested, it was not observed for parvovirus H1. It was also not observed when using AAV pseudovirions comprised of human papillomavirus capsid antigens, so this does not appear to be a general antiviral defense. It remains to be determined if sumoylation of AAV virions actually occurs under natural infection conditions and whether or not this impacts the viral infections process in any significant fashion.
2.2 Papillomaviruses
The human papillomaviruses (HPVs) are important pathogens with over 200 subtypes that cause benign disease (warts) and promote certain epithelial tumors, including cervical cancer and certain oral cancers (Lee et al. 2016; Madkan et al. 2007). HPVs infecting the mucosa can be classified in two types based on their capacity to cause carcinogenesis: low risk and high risk. Reproduction of papillomaviruses is intimately coordinated with the differentiation process of stratified epithelium (Nguyen et al. 2014). Interruption of the normal virus life cycle and persistent expression of the two oncogene proteins, high risk HPV E6 E6 and E7 , underlines the basis for cancer progression (Galloway and Laimins 2015). HPV E7 stimulates the cell cycle by promoting E2F release from pRb , while E6 promotes p53 degradation in an ubiquitin -dependent and independent manner (Doorbar 2006). However, both of these viral proteins are multifunctional and make complex contributions to both oncogenesis and normal viral reproduction. They are also involved in the molecular mechanisms by which HPV hijacks the process of keratinocyte differentiation, leading to the pathology observed with common warts. In addition to E6 and E7, two other viral early proteins, E1 (Bergvall et al. 2013) and E2 (McBride 2013), are essential for normal viral replication.
Currently there are three papillomavirus proteins that have been shown to be sumoylated, two early proteins, E2 (Wu et al. 2007, 2008) and E1 (Rangasamy and Wilson 2000), and the minor capsid protein, L2 (Marusic et al. 2010). The papillomavirus E2 protein is a multifunctional polypeptide with roles in viral DNA replication (Chiang et al. 1992), genome segregation (You et al. 2004), and transcription (Demeret et al. 1997). As a transcription factor, E2 is both an activator and repressor depending on the promoter context (Ledl et al. 2005). E2 has a major sumoylation site at lysine 292, and similar to many transcription factors, E2 modification at this site regulates its transactivation ability (Wu et al. 2008). Furthermore, E2 can interact with a variety of other host cell transcription factors such as Sp1 (Steger et al. 2002), YY1 (Lee et al. 1998), and C/EBP (Hadaschik et al. 2003), all of which are themselves sumoylated (Deng et al. 2007; Kim et al. 2002; Spengler and Brattain 2006). Preliminary results in our lab indicate that E2 reduces the sumoylation of C/EBP in vivo. Because sumoylation of C/EBP negatively regulates transcriptional synergy (Subramanian et al. 2003) the E2 -mediated reduction of C/EBP sumoylation may account for the observed enhancement of C/EBP activity by E2 protein (Hadaschik et al. 2003). Similar effects of E2 on its other sumoylated binding partners could be a general feature of E2 that contributes to its dysregulation of cellular transcription. In addition to being functionally regulated by direct sumoylation, E2 stability is also sumoylation dependent with increased sumoylation resulting in a dramatically extended half-life (Wu et al. 2009). However, this effect is not due to direct sumoylation of E2 as the lysine 292 mutant that cannot be sumoylated remains stabilized by increasing overall cellular sumoylation levels. This indicates that there is an indirect mechanism connecting sumoylation and turnover of E2 which is likely important during the viral life cycle as sumoylation is known to be a dynamic process during keratinocyte differentiation (Deyrieux et al. 2007; Heaton et al. 2012).
Papillomavirus E1 proteins are replicative helicases that bind to the viral origin of replication, catalyze unwinding of the duplex DNA, and recruits host cell replication factors to direct the synthesis of the viral genome (Bergvall et al. 2013). E1 proteins were the second viral protein to be identified as a substrate for sumoylation (Rangasamy et al. 2000). Bovine papillomavirus (BPV ) E1 is sumoylated at lysine 514 (Rangasamy et al. 2000) and human papillomavirus type 11 is sumoylated at the analogous position which is lysine 559 (Fradet-Turcotte et al. 2009); most other HPV E1 proteins have putative sumoylation sites in a similar location which suggests that E1 sumoylation is a widespread and general feature of this protein. PIAS proteins enhance the sumoylation of several tested HPV types which supports their role as a general SUMO ligase for papillomavirus E1 proteins (Rosas-Acosta et al. 2005). Initially, sumoylation was reported to be required for proper nuclear localization of BPV E1 (Rangasamy et al. 2000), but a subsequent investigation failed to detect a nuclear impairment for either BPV or HPV E1 when sumoylation was inhibited (Fradet-Turcotte et al. 2009; Rosas-Acosta and Wilson 2008), so a functional role of sumoylation has not been defined.
In addition to the two early regulatory proteins, E1 and E2 , one of the viral capsid structural proteins, L2 , is also sumoylated (Marusic et al. 2010). Like E2 protein sumoylation increases the stability of L2, though is this case it is a direct effect of SUMO addition at lysine 35 of L2. However, while the sumoylated form of L2 is more stable, it cannot bind to L1 , the major capsid protein. These results suggest that sumoylation may help regulate virion assembly through modulation of L2 levels and incorporation into the L1 virion. Interestingly, L2 also plays a role in viral infection as it accompanies the incoming viral genome to the nuclear PML bodies after virion disassembly. Bund et al. showed that L2 contains a SUMO interacting motif (SIM) that is critical for this localization, presumably through binding to sumoylated proteins in the PML body (Bund et al. 2014). Mutation of the SIM prevented localization and reduced infectivity in a pseudovirion assay, while sumoylation of L2 itself was not required.
Beyond the modification of some papillomavirus proteins by SUMO, another intriguing feature of papillomaviruses is their ability to dysregulate the sumoylation system through the activity of certain viral proteins. The first HPV protein with known effects on sumoylation is the E7 protein. E7 is an early papillomavirus protein that both helps drive the host cell into a proliferative state and counteracts innate immunity (Hebner and Laimins 2006). A major target of E7 protein is the host pRB protein that regulates the activity of the host E2 FE2F transcription factor (Roman and Munger 2013). The papillomavirus E7 protein has an LxCxE motif that binds pRB (Munger et al. 2001), and E7 binding inhibits sumoylation of pRB (Ledl et al. 2005). Since sumoylation appears to reduce the repressive ability of pRB, interaction with E7 is likely to have functional consequences. Additionally, E7 also binds many other cellular targets (Wilson and Rosas-Acosta 2003) and might be able to modulate sumoylation of these proteins to more generally influence cellular protein functions. Interestingly, many of the functions of E7 are inhibited by the cellular tumor suppressor, p14ARF (Pan et al. 2003), which is known to stimulate sumoylation of certain substrates (Rizos et al. 2005; Woods et al. 2004). This suggests that antagonism between the sumoylation inhibition by E7 and the enhancement of sumoylation by p14ARF may be an important determinant of the outcome of infections.
Like E7 , the second viral oncoprotein, E6 , also influences host sumoylation, though in a broader fashion. In a study published in 2006, Dejean’s group showed that high risk HPV-E6, but not low risk E6, was capable of binding to and inhibiting PIASy activity (Bischof et al. 2006). PIASy is a SUMO ligase which enhances sumoylation of specific substrates such as p53 (Schmidt and Muller 2003), and E6 binding blocks sumoylation of PIASy-specific substrates. Functionally, since PIASy act as a promoter of cellular senescence (Bischof et al. 2006), this inhibition of PIASy by E6 may contribute to the ability of E6 to inhibit cellular senescence. Although E6 binds to PIASy and inhibits its ligase activity, it does not target this enzyme for degradation like it does for p53 (Huibregtse et al. 1994). Mechanistically, E6 may be acting simply by sequestering or blocking PIASy in the complex. Whether or not E6 binds to other members of the PIAS family, or to other classes of SUMO ligases has not been reported. Subsequently, work in our lab demonstrated that high risk E6 proteins can also bind to Ubc9 , the SUMO conjugating enzyme (Heaton et al. 2011). Unlike the situation for PIASy, E6 targets Ubc9 for proteasomal degradation via the ubiquitin ligase E6AP which results in a significant perturbation of the overall cellular sumoylation profile. While the consequences of this interaction for viral fitness have not yet been examined, the ability of E6 to dysregulate host sumoylation globally through attacks on PIASy and Ubc9 suggest that this will have important ramifications for the viral life cycle. As will be seen in later sections, this ability of viral proteins to modulate the sumoylation system has now been demonstrated for a number of distinct viruses, and the SUMO system appears to be another important host pathway that is commonly usurped to promote the viral agenda.
2.3 Adenoviruses
Adenoviruses are primarily respiratory pathogens that are associated with a number of human diseases (Lynch et al. 2011). Adenoviruses use complex alternative splicing to make multiple proteins from each of their early genes, and these early gene products, particularly from the E1A and E1B genes, are critical for reshaping the cellular environment to facilitate viral gene expression and replication (Imperiale et al. 1995). To date, only one adenoviral protein has been identified as a substrate for sumoylation, the early gene product E1B-55K protein. The E1B-55K protein interacts with the SUMO conjugation enzyme, Ubc9 (Wimmer et al. 2013) resulting in sumoylation of E1B-55K at lysine 104 (Endter et al. 2001). Sumoylation of E1 B-55K also requires phosphorylation at a C-terminal motif (Wimmer et al. 2013), a site which is likely modified by cellular protein kinase CK2 (Ching et al. 2012). The phosphorylation site on E1B-55K protein is not part of the SUMO recognition motif so addition of the phosphate moiety is not creating a typical phosphorylation-dependent sumoylation motif (Hietakangas et al. 2006). Instead, phosphorylation appears to be providing a cross-talk mechanism between phosphorylation and sumoylation by an as yet undefined mechanism. Recently, a host cell viral restriction factor, KAP1 , was also shown to bind E1B -55K and enhance sumoylation of the viral protein, though again the mechanism is unclear (Burck et al. 2016). Interestingly, adenovirus infection also led to reduced sumoylation of KAP1 itself, resulting in decreased epigenetic silencing, suggesting a complex interplay between E1B-55K and KAP1 that is at least in part involving the sumoylation system.
Functionally, E1B-55K sumoylation is required for nuclear localization of E1B-55K (Kindsmuller et al. 2007). Sumoylation blocks a CRM -dependent nuclear export sequence (NES), thus helping retain E1B-55K in the nucleus. Subnuclear localization of E1B-55K is also regulated by the viral E4orf6 protein such that the intranuclear distribution of E1B-55K changes as E4orf6 levels increase during infection (Leppard and Everett 1999). As E4orf6 levels increase there is a decrease in E1B-55K sumoylation, suggesting that E4orf6 may in part be regulating E1B-55K through controlling sumoylation levels (Lethbridge et al. 2003). Importantly, nuclear retention and subnuclear localization of E1B-55k also relies on its interaction with the type IV and V isoforms of the PML protein, and the interaction with PML IV requires sumoylation of E1B-55K protein while the interaction with PML V is largely independent of sumoylation (Wimmer et al. 2010). Surprisingly, sumoylation of E1B -55K was not required for its interaction with another PML associated protein, Daxx , but is required for the E1 B-55K-mediated proteasomal degradation of Daxx (Schreiner et al. 2011; Wimmer et al. 2013). A mutant of E1B-55K lacking the SUMO conjugation site was defective for Daxx degradation and for transformation of primary rodent cells, indicating that sumoylation of E1B-55K is important for its transforming activity (Schreiner et al. 2011). Thus, these combined studies indicate that modification of E1B-55K protein by SUMO is clearly important for various biological activities and functions of this viral regulatory protein.
In addition to being a substrate for sumoylation, the E1B -55K protein itself has SUMO E3 ligase ability. A critical host target for E1 B-55K is nuclear p53 (Weitzman and Ornelles 2005). Through direct binding with p53, E1B-55K enhances the sumoylation of p53 (Muller and Dobner 2008). While this study reported that E1B-55K SUMO ligase activity on p53 was not observed in vitro, a subsequent study was able to confirm E3 activity by E1B-55K protein in a reconstituted in vitro sumoylation reaction, thus confirming that E1B-55K itself has intrinsic ligase activity (Pennella et al. 2010). The sumoylation of p53 by E1B-55K also requires that E1B-55K itself be sumoylated (Muller and Dobner 2008). Since E1B-55K, in conjunction with E4orf6 , mediates ubiquitinylation and proteasomal degradation of p53 , this was the first example of a viral protein having dual activity in both the ubiquitin and SUMO pathways. Mutation of the p53 sumoylation site decreased the ability of E1B-55K to repress the transcriptional activity of p53 and to tether p53 in PML bodies (Pennella et al. 2010; Wimmer et al. 2016). These results demonstrate that the E1B-55K protein induced sumoylation is functionally relevant and contributes to the overall abrogation of p53 defenses in the adenoviral infected cell.
Like E1B-55K protein, a second adenoviral protein, the E4orf3 protein, was also shown to affect sumoylation of cellular proteins (Sohn and Hearing 2012). E4orf3 protein induces sumoylation of the host Mre11 and Nbs1 proteins in a process that requires relocalization of these proteins into E4orf3 nuclear tracks and does not require the E1 B-55K SUMO ligase activity. The sumoylation of these two proteins is transient and peaks during early infection. Mre11 is modified by SUMO2 while Nbs1 is modified by either SUMO1 or SUMO2, though the functional significance of these paralog differences is not clear. Interestingly, the adenoviral induced sumoylation of these two proteins is only seen with subgroup C adenoviruses. Both the Mre11 and Nbs1 proteins are components of the MRN complex which functions in DNA damage repair and which must be inactivated by many viruses (Hollingworth and Grand 2015), so presumably the sumoylation is helping subgroup C adenoviruses to defeat this defense process. E4orf 3 from type C adenoviruses also induces relocalization and sumoylation of TFII -I, a general transcription factor that can repress the adenoviral intermediate promoter L4P (Bridges et al. 2016). Sumoylation of TFII -I is followed by ubiquitinylation and degradation of TFII -I, leading to derepression of L4P, exemplifying how adenovirus can modulate viral transcription through the host sumoylation system. Furthermore, E4orf3 does not cause an overall increase in host sumoylation (Sohn et al. 2015), so the effect on these three host proteins represents a specific and targeted role for E4orf3 protein. However, these are not the only targets for E4orf3 as a proteomic analysis found that E4orf3 expression influenced the sumoylation of 51 host proteins, most for them showing increased sumoylation (Sohn et al. 2015). E4orf3 protein was subsequently found to specifically sequester the PIAS3 SUMO E3 ligase into its nuclear tracks while not affecting the localization of other PIAS family members, including PIAS1, PIAS2, and PIAS4 (Higginbotham and O’Shea 2015). This result suggests that PIAS3 may be the primary E3 ligase through which E4orf3 protein mediates its effects on sumoylation of host proteins. The ability of E4orf3 protein to target PIAS3 was not restricted to the subclass type C adenoviruses, suggesting that E4orf3-induced effects on sumoylation will have a more general role in the adenoviral life cycle beyond the regulating the MRN and TFII -I proteins.
Another adenovirus protein that influences host sumoylation is the E1A protein (Ledl et al. 2005). E1A is an early protein that regulates viral genome transcription and contributes to cell transformation (Berk 2005). A major target for E1A is the host pRB protein, a regulatory factor that binds to E2 FE2F and masks the E2F transcriptional activation of S phase genes (Dyson 2016). In addition, pRB can recruit repressive chromatin remodeling factors to E2F bound promoters to further silence gene expression (Berk 2005). Ledl et al. showed that SUMO is attached to pRB at a single residue, lysine 720, in the B-box motif which interacts with LxCxE-motif proteins such as E1A (Ledl et al. 2005). Mutation of lysine 720 results in a pRB protein with increased repressive activity on an E2F -responsive promoter, indicating that sumoylation negatively regulates pRB repressive activity. E1A is known to bind pRB leading to the release and activation of E2F . However, binding of E1 A to pRB also prevents sumoylation of lysine 720, thereby providing another level of control of pRB by this viral protein (Ledl et al. 2005). Ultimately, E1A seeks to activate cellular S-phase and increase transcription of DNA replication related genes to promote viral genome reproduction, and influencing the sumoylation state of pRB may provide fine regulation of E2F activity. Furthermore, E1A binds to a variety of other cellular proteins (Berk 2005), at least two of which are themselves sumoylated, CtBP (Lin et al. 2003) and p300 (Girdwood et al. 2003). This raises the possibility that E1A might also decrease the sumoylation state of binding partners other than pRB if complex formation also blocks their sumoylation. Additionally, E1A also directly binds the SUMO conjugase, Ubc9 (Hateboer et al. 1996). The binding element in E1A is located within conserved region 2 and is comprised of the sequence EVIDLT (Yousef et al. 2010). This E1A motif interacts with the N-terminal region of Ubc9 , a region which also binds SUMO and is involved in polysumoylation (Knipscheer et al. 2007). While E1A binding to Ubc9 did not affect global sumoylation in cultured human cells, this interaction did interfere with polysumoylation in a yeast model, suggesting that subtle alterations in host polysumoylation could be occurring during viral infection (Yousef et al. 2010). For example, Yousef et al. observed that E1 A affected PML localization in an Ubc9 -dependent manner, and PML is known to be polysumoylated (Tatham et al. 2001), so perhaps E1A is disrupting PML polysumoylation through blocking this activity of Ubc9 . Alternatively, since E1A does not block the monosumoylation activity of Ubc9 (Yousef et al. 2010), E1A could be enhancing the sumoylation of specific substrates by redirecting Ubc9 to PML bodies or other E1A targets. Clearly, a great deal of further investigation is required to sort out all the possible ramifications for sumoylation in the adenoviral life cycle.
Lastly, while not a human virus, it is germane to mention the GAM1 protein of the avian type 1 adenovirus known as CELO (Chicken Embryo Lethal Orphan virus) (Chiocca et al. 1996). GAM1 has the most dramatic inhibition of global sumoylation of all known viral proteins, and is well-characterized mechanistically. Moreover, a CELO Gam1 negative mutant is replication-defective, which clearly establishes GAM1 as an important protein for the viral life cycle. Interestingly, this 30 kDa viral protein has no homology to other known anti-apoptotic proteins such as E1B from the adenovirus type 5 or to the Bcl2 and Bax family of eukaryotic proteins. GAM1 is predominantly located in the nucleus, and sumoylation is globally reduced when GAM1 is expressed in a dose dependent manner (Boggio et al. 2004; Colombo et al. 2002). GAM1 was also shown to re-distribute SUMO from the nucleus to the cytoplasm , promoting nuclear transcription factor and HDAC (histone deacetylase ) de-sumoylation, thus positively influencing cellular transcription. Mechanistically, it was established that GAM1 binds to the SUMO activation enzyme complex, SAE1/SAE2 (Boggio et al. 2004) and reduces the half-life of SAE1/SAE2 by recruiting a cellular ubiquitin E3 ligase to the GAM1-SAE1/SAE2 complex that ubiquitinylates SAE1 leading to proteasomal degradation (Boggio et al. 2007). The loss of SAE1 destabilizes both SAE2 and Ubc9 , resulting in reduction in their levels as well. Even though the de-sumoylation process is unaffected by GAM1, the inability to perform new SUMO modification greatly decrease the entire pool of sumoylated subtracts. The net result of this GAM1 effect is an increase in overall cellular transcriptional activity which facilitates viral replication. While GAM1 may be an extreme example, it is likely that other viruses have evolved related strategies to target the sumoylation enzymes in order to enhance the cellular transcriptional environment to favor their replication needs.
2.4 Herpesviruses
The family Herpesviridae is a diverse group with three subfamilies (α, β, and γ) that encompasses eight human viruses termed human herpesviruses (HHV) 1-8, though most have common names such as cytomegalovirus and Epstein-Barr virus (Grinde 2013). A characteristic feature of this family is that all members can establish latent infections, either with or without an acute disease presentation, leading to very complex interactions with their respective host cells. Studies over the last 15 years have revealed that sumoylation plays important roles in the life cycle of the herpesvirus family members, both through sumoylation of viral proteins and through viral manipulation of the host sumoylation process. Of the eight human herpesviruses , five have been shown to have one or more sumoylated viral proteins: cytomegalovirus (CMV) (Hofmann et al. 2000; Muller and Dejean 1999; Sinigalia et al. 2012), Epstein-Barr virus (EBV) (Adamson and Kenney 2001; Chang et al. 2004; Rosendorff et al. 2004), Kaposi’s sarcoma-associated herpesvirus (KSHV) (Izumiya et al. 2005), varicella-zoster virus (VZV) (Stallings and Silverstein 2006), and human herpesvirus 6 (HHV6) (Gravel et al. 2002; Stanton et al. 2002). These five herpesviruses include representatives of the alpha (VZV ), beta (HCMV and HHV6), and gamma (EBV and KSHV ) subgroups, further demonstrating that sumoylation of herpesvirus proteins is a frequent and common event for members of this family. For the remaining three human herpesviruses (herpes simplex types 1, herpes simplex type 2, and human herpesvirus 7) sumoylation of viral proteins has not been reported, though a recent proteomics study suggested that several HSV 1 proteins may be modified by SUMO2 (Sloan et al. 2015). Based on these observations, it is likely that there will be sumoylated proteins identified for the remaining human herpesviruses types as well.
As discussed in previous sections, papillomaviruses and adenoviruses target the sumoylation system to alter the cellular milieu and/or avoid host defenses, and this strategy is used by several herpesviruses as well. For example, while HSV has not yet been definitely shown to have sumoylated viral proteins, it does modulate host sumoylation through the viral ICP0 protein (Boutell et al. 2011). A number of other herpesviruses also manipulate the host sumoylation system through viral proteins that have SUMO ligase activity, SUMO targeted ubiquitin ligase (STUbL ) activity, or undefined mechanisms. In some cases this reflects a global alteration in host sumoylation while in other cases the effects are confined to specific substrates. Several recent reviews examine the role of sumoylation for herpesviruses (Boutell and Everett 2013; Campbell and Izumiya 2012; Chang et al. 2016), and the following sections will provide a brief overview for the individual family members.
2.4.1 Herpes Simplex Virus
The initial observations that viruses could impact host sumoylation were made for an alpha herpes virus, herpes simplex (HSV). The HSV ICP0 immediate early gene product (also known as Vmw110) is not itself sumoylated, but causes loss of high molecular weight isoforms of PML (Everett et al. 1998). These forms were eventually determined to be SUMO-modified versions of PML, a major component of nuclear ND10 bodies (Muller and Dejean 1999). Similarly, ICP0 also decreases the amount of sumoylated Sp100 , another major constituent of ND10s (Everett et al. 1998). ND10 disruption by HSV is necessary for effective lytic replication, and this disruption requires ICP0. Originally, this loss of sumoylated PML and Sp100 was believed to cause disruption of the ND10s, but subsequent studies showed that the SUMO protease , SENP1, could elicit similar loss of the sumoylated forms of PML and Sp100 without affecting ND10 structures (Bailey and O’Hare 2002). ICP0 does not inhibit PML sumoylation nor does it cause desumoylation of PML in vitro (Boutell et al. 2003), though it does recruit SENP1, at least under conditions of transient co-expression, which may contribute to desumoylation of PML components (Bailey and O’Hare 2002). In subsequent studies it became clear that the major effect of ICP0 on ND10 bodies appears to be due to its known ubiquitin ligase activity (Boutell et al. 2002). ND10 bodies are part of an intrinsic anti-viral defense that blocks HSV infection effectively in the absence of ICP0 (Everett et al. 2006, 2008). Several of the components of ND10 bodies, including PML, Sp100 , and Daxx contain SUMO interacting motifs (SIMs) that are critical for recruitment of these proteins to the HSV genome (Cuchet-Lourenco et al. 2011). ICP0 also has SIM motifs that are required for its ability to overcome the anti-viral function of ND10s (Boutell et al. 2011). These SIM motifs help direct ICP0 to sumoylated proteins where it causes their ubiquitinylation and proteasomal degradation. While ICP0 causes a general decrease in cellular sumoylated proteins, sumoylated PML is preferentially degraded (Sloan et al. 2015). One putative SIM in the central region of ICP0 was absolutely required for this general effect on sumoylated host proteins and for PML isoforms other than type 1, while several putative SIMs in the C-terminal region also contributed (Everett et al. 2014). Interestingly, ICP0 can also interact directly with PML type 1 and target it for degradation in a SUMO-independent fashion, suggesting that this isoform may be particularly important for restricting HSV replication (Cuchet-Lourenco et al. 2012). Overall, it appears that ICP0 acts as a SUMO-targeted ubiquitin ligase (STUbL) that is highly active against PML and other components of ND10 bodies, leading to their degradation, and thus overcoming this host defense mechanism sufficiently to allow HSV productive infections (Lanfranca et al. 2014). Since a proteomic analysis of SUMO 2 identified 124 other host proteins whose sumoylation was significantly reduced by ICP0 (Sloan et al. 2015), it will also be of interest to evaluate the biological role of these other proteins in HSV replication.
Further evidence for the importance of sumoylation in the HSV life cycle comes from the recent observations that the PIAS SUMO ligases contribute to the intrinsic anti-viral response to this virus. PIAS1 is localized to ND10 bodies in a SIM -dependent fashion (Brown et al. 2016), likely through interaction with sumoylated PML . PIAS1 causes SUMO1 accumulation at sites of HSV nuclear entry, and helps inhibit HSV replication by a mechanism that is additive to PML. Like PML, the restrictive effect of PIAS1 is overcome by ICP0 . Similarly, PIAS4 also associates with HSV nuclear entry sites in a SIM -dependent process and cooperates with PML to restrict HSV (Conn et al. 2016). ICP0 can reduce this accumulation and overcome the restriction imposed by PIAS4. These combined results strongly support an anti-herpes simplex role for PIAS1 and PIAS 4 that is functionally reduced by ICP0.
2.4.2 Varicella-Zoster Virus
The second human alpha herpesvirus is varicella-zoster virus, the etiological agent of chickenpox and the reactivation disease known as shingles (Gilden et al. 2015). Sumoylation in this member of the alpha subgroup has not been extensively studied, and there are only two reports concerning sumoylation and this virus. It was reported that one of the viral early proteins, ORF29p, is sumoylated, but the role of sumoylation in the function of this protein was not investigated (Stallings and Silverstein 2006). A second viral protein, ORF61, contains three SIMs that facilitate binding to SUMO1 (Wang et al. 2011). These SIMs are required for ORF61 to interact with and disperse PML in ND10 bodies. During skin infection, the number of ND10 bodies is reduced, but this is not observed in ORF61 SIM mutants. Also, these mutants did not demonstrate typical skin lesions and had reduced viral spread. There results suggest that the SUMO binding function of ORF61 is critical for overcoming the anti-viral effects of the PML bodies, similar to ICP0 of HSV. ORF61 is homologous to ICP0 (Moriuchi et al. 1992) and has ubiquitin ligase (Everett et al. 2010), but whether or not ORF61 has STUbL activity has not been determined.
2.4.3 Cytomegalovirus
The family of beta herpesviruses includes three human virus members, cytomegalovirus (CMV), human herpes virus 6 (HHV6), and human herpes virus 7 (HHV7). There is not yet a report on sumoylation in HHV7, but sumoylation plays a role in the life cycle of both HHV6 and CMV. The immediate early IE1 protein is sumoylated for both CMV (Muller and Dejean 1999) and HHV6 (Gravel et al. 2002). Additionally, IE2-p86 (Hofmann et al. 2000), the UL44 protein (Sinigalia et al. 2012), and the pp71 tegument protein (Hwang and Kalejta 2009) for CMV are sumoylated. Multiple groups identified lysine 450 as the SUMO acceptor site in IE1 (Lee et al. 2004; Nevels et al. 2004; Sadanari et al. 2005; Spengler et al. 2002; Xu et al. 2001), but the role of SUMO modification in IE1 function remains poorly understood. Like HSV ICP0 , the CMV IE1 disrupts ND10s and reduces the level of sumoylated forms of PML and Sp100 (Everett et al. 2013a; Muller and Dejean 1999; Tavalai et al. 2011), but unlike ICP0 this disruption is proteasome independent (Lee et al. 2004; Xu et al. 2001). IE1 also does not cause any global decrease in sumoylated proteins so it does not appear to be a STUbL like ICP0 (Scherer et al. 2013). Furthermore, IE1 mutants with the SUMO acceptor site lysine changed are still capable of disrupting PML ND10 bodies and overcoming the anti-viral activity of these elements, indicating that sumoylation of IE1 itself is not involved in this process (Lee et al. 2004; Spengler et al. 2002; Xu et al. 2001). IE1 also does not have an intrinsic SUMO protease activity, and it has been suggested that the desumoylation of PML seen with IE1 expression may be due to disruption of PML aggregation (Kang et al. 2006).
Since sumoylation of IE1 is not related to its function in disrupting ND10 bodies, the actual role of this modification remains vague, though a K450R mutant virus grows more slowly and with reduced yield, suggesting that sumoylation is important for a fully robust virus (Nevels et al. 2004). Conflicting reports exist for a role for sumoylation in IE1 intracellular localization with Nevels et al. finding no sumoylation requirement for proper localization of IE1 (Nevels et al. 2004), while a second report indicated that sumoylated and unsumoylated IE1 exhibited different cellular fraction properties which could reflect different intracellular locations (Sadanari et al. 2005). In other studies, IE1 mutants with the SUMO acceptor lysine altered were unaffected for protein stability and transactivation activity on several promoters, so sumoylation is unlikely to contribute generally to those functions (Spengler et al. 2002). However, while transactivation at several promoters is unaffected by the SUMO site mutation, the mutant exhibited decreased IE2 transcripts and protein expression, so specific regulation of IE1’s activity on the IE2 promoter may be a critical function of SUMO modification (Nevels et al. 2004).
To date, the only other reported effect for IE1 sumoylation is on its interaction with the host STAT2 protein (Huh et al. 2008). STAT2 is anti-viral in that it induces interferon -stimulated genes, but this function is antagonized by unmodified IE1 which reduces STAT2 binding to target promoters. Sumoylation of IE1 prevents its binding to STAT2, suggesting that sumoylation is actually a negative regulator of IE1 activity in this pathway. Thus, sumoylation of IE1 appears to have both a positive role in IE2 expression and a negative role in preventing IE1 inhibition of the anti-viral STAT2 pathway.
In contrast to IE1 , sumoylation of IE2-p86 protein has more clearly defined functional effects. IE2-p86 is a transactivator of both viral and cellular promoters, and it plays a role in both lytic and latent infections. There are two sumoylation sites in IE2-p86, at lysines 175 and 180, and the protein is effectively modified by either SUMO1 or SUMO2 (Ahn et al. 2001; Hofmann et al. 2000). While targeting of IE2-p86 to ND10 bodies was not dependent upon sumoylation, mutational inactivation strongly decreased the ability of IE2-p86 to activate viral early promoters (Hofmann et al. 2000). Conversely, over-expression of SUMO1 increased the transactivation ability of IE2-p86 (Ahn et al. 2001), so the combined results are consistent with sumoylation of IE2-p86 being a positive regulator of transactivation. In the context of viral infections, this effect of sumoylation on IE2-p86 appears to be biologically important. While it was initially reported that mutation of the IE2-p86 sumoylation sites had no effect on growth of the CMV Towne strain (Lee and Ahn 2004), subsequent studies with other CMV strains, including a clinical isolate, showed a major impact on viral growth for the sumoylation minus IE2-p86 protein (Berndt et al. 2009; Kim et al. 2010). Viruses with sumoylation defective IE2-p86 had reduced levels of immediate-early gene products and much lower levels of viral DNA replication (Berndt et al. 2009), consistent with IE2-p86 sumoylation being critical for the normal viral replicative cycle, except perhaps in the Towne strain.
In addition to being sumoylated, IE2-p86 noncovalently binds SUMO (Ahn et al. 2001; Hofmann et al. 2000), leading to the identification of a SIM motif at amino acid 200 (Berndt et al. 2009). SIM minus mutants showed reduced sumoylation of IE2-p86 leading to replication impairment similar to SUMO site mutants (Berndt et al. 2009; Kim et al. 2010). Additionally, SIM mutants showed reductions in IE2 association with viral promoters, formation of viral transcription domains, and late protein expression (Kim et al. 2010). The SIM motif in IE2-p86 was also necessary for interaction with the host sumoylated form of the TATA-binding protein (TBP) , an interaction which enhanced IE2’s transactivation ability (Kim et al. 2010). Given that many other transcription associated factors are known to be sumoylated, the potential for IE2-p86 to recruit numerous other host proteins through SIM-SUMO interaction is intriguing. Lastly, a recent study found that IE2 inhibited IE1 sumoylation (Kim et al. 2014). This negative regulation was found to be mediated through PIAS1 binding by IE2. PIAS1 stimulates sumoylation of IE1, so binding of PIAS1 by IE2 may sequester it and prevent PIAS1-mediated sumoylation of IE1 . Consistent with this model, levels of IE1 sumoylation peaked early in infection and decreased in late phase where IE2 levels were highest. Thus, a complex interplay between IE1, IE2 , and the SUMO system appears to contribute to the fine regulation of viral protein activities.
The third CMV viral protein known to be sumoylated is the UL44 protein (Sinigalia et al. 2012). UL44 is a subunit of the viral DNA polymerase that confers processivity through binding to the viral DNA template. UL44 both binds Ubc9 and is extensively sumoylated with at least 16 SUMO attachment sites identified by mass spectrometry , leading to modification of approximately 50% of the protein late in infection. Sumoylation with either SUMO1 or SUMO2/3 enhanced DNA binding by UL44, but SUMO over-expression during infection led to a decrease in UL44 association with viral replication centers, suggesting that sumoylation may actually be a negative regulator of UL44 function for viral replication. Paradoxically, over-expression of sumoylation increased levels of viral DNA and resulted in two to three fold higher virus titers, though this effect could be due to sumoylation of the other viral or host proteins, so the biological consequences of UL44 sumoylation remain unclear.
The final CMV protein known to be involved in sumoylation is the pp71 tegument protein (Hwang and Kalejta 2009). Tegument proteins are components of the virion and are delivered to the host cell upon uptake of the virion. Pp71 acts to overcome the repressive effects of the Daxx host protein on viral early promoters by mediating degradation of Daxx. Hwang and Kalejta showed that expression of pp71 causes an increase in the sumoylation of cellular Daxx protein (Hwang and Kalejta 2009). This stimulation of Daxx sumoylation required direct interaction between pp71 and Daxx, but was not required for pp71-induced degradation of Daxx. Furthermore, a functional effect of this pp71-induced sumoylation of Daxx on viral immediate early promoters was not observed, so like UL44 the significance of this pp71 effect is not yet defined.
2.4.4 Human Herpesvirus 6
Like CMV, human herpesvirus 6 (HHV6) is a betaherpesviruses and expresses an IE1 protein, though the CMV IE1 and the HHV6 IE1 lack significant identity at the protein level (Gravel et al. 2002). Even without much relatedness to CMV IE1, the HHV6 IE1 still possess a transcriptional activating function and is sumoylated (Gravel et al. 2002; Stanton et al. 2002). There is a single SUMO attachment site in IE1 at lysine 802, and SUMOs 1–3 could each be conjugated (Gravel et al. 2004). Polysumoylation at this site was also observed.
Like CMV IE1 , HHV6 IE1localizes to ND10 bodies (Gravel et al. 2002; Stanton et al. 2002). Also like CMV IE1, sumoylation of HHV6 IE1 is not required for localization to ND10s (Gravel et al. 2004). Surprisingly, HHV6 IE1 does not cause disruption of ND10s when transiently expressed alone. Even during viral infection, ND10s did not disperse and instead condensed into a smaller number of larger bodies (Gravel et al. 2002). Furthermore, it has not yet been reported that HHV6 IE1 affects the sumoylation of PML or any other cellular proteins. These results indicate that HHV6 does impact ND10 structure, though whether or not is functionally inactivates ND10s is unknown. It was shown that over-expression of SUMO increases IE1 levels (Gravel et al. 2004; Stanton et al. 2002), however, this does not require an intact sumoylation site (Gravel et al. 2004) and is apparently an indirect effect whose mechanism in unknown.
There was also no indication that sumoylation affected the transactivation ability of HHV6 IE1 , so the functional consequences of IE1 sumoylation, if any, remain to be discovered.
HHV6 also expresses an IE2 immediate early protein that is a transcriptional transactivator . Unlike the CMV IE2 protein, no sumoylation of HHV6 IE2 has been detected (Tomoiu et al. 2006). Interestingly though HHV6 IE2 does bind to Ubc9 and this binding represses transactivation activity. The repression did not require catalytically active Ubc9 so appears not to involve sumoylation, and both the mechanism and functional consequences for viral infection are uncharacterized.
2.4.5 Epstein-Barr Virus
The third herpesvirus subgroup, gamma, contains two human virus members, Epstein-Barr virus (EBV) and Kaposi’s sarcoma herpes virus (KSHV) . ). EBV is the etiological agent of infectious mononucleosis and is also associated with several diverse cancer s (Hislop 2015). Sumoylation plays an important role in the life cycle of this virus with four viral proteins known to be sumoylated (BZLF 1, Rta, EBNA 3B, and EBNA3C), three viral proteins that modulate sumoylation (LF2, LMP, and BGLF 4), several other viral proteins that have putative SIM motifs (Li et al. 2012a), and one report suggesting that viral microRNAs may modulate the sumoylation system (Callegari et al. 2014). The first reported EBV protein identified as a SUMO substrate is the BZLF1 protein (also known as Z protein or ZTA), one of the EBV immediate early gene products (Adamson and Kenney 2001). Two groups demonstrated that BZLF1 has a single SUMO acceptor lysine at position 12, and that BZLF 1 can be modified by SUMOs 1-3 (Hagemeier et al. 2010; Murata et al. 2010). Sumoylation of BZLF1 can be regulated by the viral protein kinase (EBV-PK, also known as BGLF 4) whose expression decreases sumoylation of BZLF1 (Hagemeier et al. 2010). This effect on sumoylation of BZLF1 does not require phosphorylation of BZLF1 itself so the actual mechanism is unknown. Functionally, the consensus is that sumoylation represses that transactivation ability of BZLF1 (Adamson 2005; Hagemeier et al. 2010; Murata et al. 2010) as mutation of the SUMO acceptor site increases transactivation activity as does desumoylation of BZLF1 with exogenously expressed SENP . At least part of the repressive effect appears to be due to preferential interaction of cellular histone deacetylase 3 (HDAC3) with the sumoylated form of BZLF1 (Murata et al. 2010). Similarly, the cellular scaffolding protein, RanBPM, was shown to enhance BZLF1 transactivation by binding BZLF1 and inhibiting its sumoylation (Yang et al. 2015b). Consistent with the repressive effect of sumoylation on BZLF1, a nonsumoylatable lysine 12 to alanine mutation exhibited two-fold increased viral production (Hagemeier et al. 2010).
In contrast to the repression of transactivation activity, sumoylation of BZLF1 has no effect on its intracellular localization or stability (Adamson 2005; Hagemeier et al. 2010), though it does contribute to ND10 body disruption by EBV (Adamson and Kenney 2001; Hagemeier et al. 2010). BZLF1 protein expressed alone is sufficient to disrupt ND10 bodies, in part because it reduces the sumoylation of PML in ND10 bodies apparently by competing for limited SUMO1 (Adamson and Kenney 2001). However, a SUMO site mutation in BZLF1 still shows partial ability to disrupt ND10 bodies, so reduction of PML sumoylation by competition cannot be the only mechanism by which BZLF1 causes ND10 dispersion (Hagemeier et al. 2010). Whether or not BZLF 1 affects the sumoylation of other proteins has not been examined, but would be predicted based on the proposed competition mechanism.
A second immediate early protein for EBV is the BRLF 1 protein which is also known as Rta. Like BZLF1, Rta is sumoylated, though for this protein there are three sumoylation sites at lysines 19, 213, and 517 (Chang et al. 2004). Members of the PIAS family act as SUMO ligases for Rta as PIAS1 and PIASx (alpha and beta) interact with Rta and stimulate its sumoylation (Liu et al. 2006). In contrast to BZLF1, it was initially reported that the transactivation activity of Rta is enhanced by sumoylation (Chang et al. 2004; Liu et al. 2006). However, a later study demonstrated that sumoylation was not needed for transactivation (Calderwood et al. 2008), so a functional role for Rta sumoylation in transactivation remains questionable. Interestingly, there is additional complex regulation of Rta function, some of which involves sumoylation. Two cellular proteins, RanBPM and RNF4 , and one viral protein, LF2, have been shown to influence Rta and affect sumoylation. RanBPM, the nucleocytoplasmic scaffolding protein that inhibits BZLF 1 sumoylation has the opposite effect on Rta as RanBPM binds Rta and enhances its sumoylation leading to increased activity on several promoters (Chang et al. 2008). However, sumoylation of Rta promotes its association with a cellular STUbL known as RNF4 through the SIM motifs in RNF4 (Yang et al. 2013). Thus, Rta sumoylation may enhances its activity, but it also leads to its proteasomal degradation, perhaps providing a check and balance possibly to prevent excessive Rta activity. Rta activity is also negatively regulated by the viral LF2 protein (Calderwood et al. 2008). LF2 expression induces Rta modification by SUMO2 and SUMO3 at four additional residues, lysines 426, 446, 517, and 530 (Heilmann et al. 2010). Paradoxically, these SUMO modifications are not required for the negative regulation by LF2 which is due to LF2 binding and relocalization of Rta to the extranuclear cytoskeleton . Consequently, neither endogenous sumoylation nor LF2-induced sumoylation events have a clearly defined effect on Rta and understanding the biological role of SUMO modification for this protein awaits further investigation.
The last known sumoylated EBV proteins are the related EBNA 3B and 3C proteins (Rosendorff et al. 2004). EBNA3B and 3C, along with a third related gene product, EBNA3A are transcriptional regulatory factors with both overlapping and distinct functions (Robertson et al. 1996). Both EBNA3B and 3C are sumoylated with sumoylation being much more prominent for EBNA3C; EBNA3A was not seen to be sumoylated (Rosendorff et al. 2004). Mutants of EBNA3C that could not be sumoylated were more diffusely distributed rather than accumulating in nuclear dots. Nonetheless, these mutants were still wild-type in their ability to transactivate the LMP1 promoter in conjunction with EBNA2, so this function does not appear to require sumoylation. Instead of covalent modification by SUMO, one or more SIM-like motifs in EBNA3C are important for LMP1 promoter activation (Lin et al. 2002; Rosendorff et al. 2004). EBNA3C interacts with p300/CBP at the LMP1 promoter, but a p300 mutant that was not sumoylated could still be co-activated by EBNA3C, so p300 is not the target for the EBNA3C SIM motif(s) (Rosendorff et al. 2004). This led Rosendorff et al. to propose that binding of EBNA3C to an unidentified sumoylated repressor counteracts that repressive effect to allow LMP1 promoter activation. The potential role of EBNA3B sumoylation was not investigated.
As mentioned above, the EBV protein kinase (BGLF4) is able to inhibit sumoylation of the BZLF1 protein (Hagemeier et al. 2010). BGLF 4 was subsequently shown to have two SIM motifs, one in the N-terminal region and one in the C-terminal region adjacent to a nuclear export sequence (Li et al. 2012a). Both SIM motifs contribute to nuclear accumulation of BGLF4 by blocking nuclear export. SUMO binding by BGLF4 was also required for its ability to inhibit sumoylation of BZLF1 , as was the kinase activity of BGLF4. Since it was previously shown that phosphorylation of BZLF1 itself by BGLF4 is not required for inhibition of sumoylation (Hagemeier et al. 2010), the requisite target for the kinase activity in this process is not known. Interestingly, BGLF4 could also globally reduced cellular sumoylation in a SIM - and kinase-dependent fashion, though what, if any, biological role this has in the viral life cycle was not tested (Li et al. 2012a). Lastly, BGLF 4 also contributes to several other viral functions, including ND10 dispersion, DNA damage response induction, and production of extracellular virus, and all these activities required the intact SIM s. Thus, while targets and mechanistic pathways for all of these BGLF4-related functions are relatively undefined, it is likely that the all involve targeting of BGLF4 through interaction with SUMO moieties conjugated to various host and/or viral proteins.
Lastly, a very important effect on sumoylation has been elucidated for latent membrane protein 1 (LMP1). LMP1 is the primary viral oncoprotein and it plays a key role in maintaining the latent state (Li and Chang 2003). While not itself sumoylated, LMP1 interacts with Ubc9 through sequences in its C-terminal activating region 3 (CTAR3) and causes a general increase in host protein sumoylation (Bentz et al. 2011). Subsequently, two specific cellular targets of LMP1-mediated sumoylation were identified, IRF7 (Bentz et al. 2012) and KAP1 (Bentz et al. 2015). IRF7 is an interferon regulatory factor involved in innate immunity. LMP1 promotes sumoylation of IRF7 at lysine 452, resulting in decreased IRF7 function which likely helps abrogate the innate response during EBV latency (Bentz et al. 2012). KAP1 (KRAB-associated protein 1) also is believed to have antiviral activity, likely through its transcriptional repressive activity (Iyengar and Farnham 2011). KAP1 was shown to bind to EBV early promoters and the lytic origin of replication (OriLyt) in a sumoylation-dependent fashion (Bentz et al. 2015). These results suggested that LMP1-mediated sumoylation of KAP1 helps to repress the lytic phase of EBV and promote maintenance of the latent state. It will be of interest to see if there are other targets of LMP1-mediated sumoylation that also contribute to the viral life cycle.
2.4.6 Kaposi’s Sarcoma-Associated Herpes Virus
Like the other human virus member of the gamma herpesvirus subgroup, sumoylation is also highly important of Kaposi’s sarcoma-associated herpes virus (KSHV ) (Campbell and Izumiya 2012; Chang and Kung 2014). Four KSHV viral proteins, K-bZIP , K-Rta, LANA1 (ORF73), and LANA2 (vIRF3) modulate sumoylation of host and/or viral proteins. K-bZIP was the first KSHV protein identified as a SUMO substrate with modification occurring at lysine 158 (Izumiya et al. 2005). Sumoylation of K-bZIP is negatively regulated by phosphorylation at threonine 111 catalyzed by the viral ORF36 protein kinase (Izumiya et al. 2007). However, it was later reported that K-bZIP splice variants exist, and that 2 of the variants lack the lysine 158 and are sumoylated on newly generated lysine, residue 207 (Lefort et al. 2010). What, if any, different properties the variants have has not been extensively investigated.
Functionally, K-bZIP is a transcriptional repressor, and sumoylation of K-bZIP enhances its repressive activity (Izumiya et al. 2005). Additionally, K-bZIP binds Ubc9 and recruits it to viral promoters where it likely sumoylates and inactivates other transcription factors to facilitate repression. One pathway that has been characterized for K-bZIP is interferon -stimulated gene expression where K-bZIP represses target genes (Lefort et al. 2010). A SUMO site mutant of K-bZIP is impaired for the ability to repress these target genes, indicating that sumoylation is a positive regulator for this activity of K-bZIP. Subsequently it was shown that K-bZIP is a viral SUMO E3 ligase that can catalyze its own sumoylation (Chang et al. 2010). K-bZIP contains a SIM motif the preferentially binds SUMO2/3 and not SUMO1, and its ligase function also utilizes primarily SUMO2/3. Using this ligase activity K-bZIP can sumoylate known binding partners such as p53 and pRB , and thus regulate their activities. More globally, during KSHV reactivation there is an increase of SUMO2/3 association with promoter regions of both cellular (Chang et al. 2013) and viral genomes (Yang et al. 2015a). For the cellular genome, the viral factor(s) responsible were not examined, but K-bZIP would be a likely candidate. Consistent with this speculation, K-bZIP was required for the increased SUMO2/3 association with viral DNA as a mutant lacking the SUMO ligase activity showed no increase (Yang et al. 2015a). Importantly, the SUMO ligase defective mutant of K-bZIP showed increased virus production during infection, and the same results were seen with a SUMO2/3 knockdown. These results indicated that K-bZIP induced sumoylation is repressing viral activation and may function to help maintain the latent state. Similarly, enhanced SUMO2/3 modification at cellular promoters is likely to help prevent viral reactivation and/or to repress cellular innate immune responses to the virus.
While K-bZIP has SUMO E3 ligase activity that enhances sumoylation of viral and cellular proteins, the K-Rta protein is a STUbL that targets sumoylated proteins for proteasomal degradation (Izumiya et al. 2013). K-Rta has multiple SIM motifs that provide strong binding to multimerized SUMO, with a preference for SUMO2/3. PML is one of the specific targets for K-Rta leading to degradation of PML and contributing to the dispersal of the ND10 bodies by this virus. K-Rta can also target sumoylated K-bZIP for degradation, suggesting a mechanism for counteracting the repressive function of K-bZIP. Mutations in the SIM motifs of K-Rta prevent PML degradation, decrease transcriptional activation by K-Rta, and reduce viral replication, which is strongly consistent with the STUbL activity being important for all these functions. Further characterization of the opposing functions of K-bZIP and K-Rta will likely provide greater insight into the regulatory balance between viral reproduction and latency.
KSHV has two latency associated nuclear antigens, LANA1 and LANA2 , that both contribute to modulation of substrate sumoylation status. LANA1 contains a SIM motif that is specific for SUMO2 and which facilitates the sumoylation of LANA1 at lysine 1140 (Cai et al. 2013). Through this SIM element, LANA1 recruits poly-sumoylated host KAP1 protein which is part of a transcriptional repressive complex that also includes Sin3A. Deletion of the SIM motif prevents association of LANA1 with this complex and results in loss of viral episomal genome maintenance and loss of repression of K-Rta expression, implicating this SIM as a critical part of the mechanism by which LANA1 helps to establish and maintain latency. LANA1 has been shown to enhance sumoylation of Sp100 which promotes its accumulation into nuclear aggregates which are likely ND10 bodies, and it was proposed the ND10-induced restrictions may be necessary for establishment of viral latency (Gunther et al. 2014). Similarly, LANA1 enhances sumoylation of histones in a SIM -dependent which is correlated with repression of viral genes and again may contribute to establishment of latency (Campbell and Izumiya 2012). Interestingly, a broader proteomic study identified 151 host proteins that interacted with the LANA1 SIM (Gan et al. 2015). These results suggest that LANA1 could be modifying and regulating a large and complex protein network to facilitate its latency function for KSHV .
LANA2 is another latency associated protein that modulates the sumoylation state of partner proteins. LANA2 increases sumoylation of PML which ultimately leads to disruption of ND10 bodies due to proteolytic degradation (Marcos-Villar et al. 2009, 2011). Both an intact SIM motif in LANA2 and sumoylation of LANA2 at multiple sites were important for the ND10 dispersion. In contrast to the enhancement of PML sumoylation, LANA2 binding to the pocket proteins (pRB , p107, and p130) (Marcos-Villar et al. 2014) or p53 (Laura et al. 2015) inhibits their sumoylation. For the pocket protein, this effect requires binding via an LxCxE motif in LANA2 that is typical of proteins that interact with pRB and the other pocket proteins (Marcos-Villar et al. 2014). For p53 , inhibition of sumoylation requires both the LANA2 SIM motif and sumoylation of LANA2 (Laura et al. 2015). The ability of LANA2 to target and influence the sumoylation of two key cellular regulatory proteins, pRB and p53 , strongly implies that modulating these pathways is critical in the viral life cycle.
2.5 Poxviruses
Poxviruses are complicated pathogens with complex virions, large genomes, and cytoplasmic genome replication, which is distinct from the typical nuclear location for genome replication other DNA viruses (Lefkowitz et al. 2006). The primary human pathogen was variola, the agent of smallpox , though most studies in recent decades have been conducted on the vaccine strain known as vaccinia virus (Roberts and Smith 2008; Voigt et al. 2016). To date, only two the vaccinia virus proteins, A40R (Palacios et al. 2005) and E3 (Gonzalez-Santamaria et al. 2011) are known to be targets for sumoylation. E3 was originally shown to interact with SUMO1 in a yeast two hybrid assay (Rogan and Heaphy 2000). A subsequent study confirmed the interaction with SUMO and demonstrated that a SUMO interacting motif (SIM) was required for this association (Gonzalez-Santamaria et al. 2011). Interestingly, the SIM motif in E3 was required for protein stability, nuclear localization, and efficient sumoylation of E3. With the SIM motif intact, E3 could be modified by either SUMO1 or SUMO2, with SUMO addition sites mapped to lysines 40 and 99. Sumoylation of E3 occurred both in the context of viral infection and with transfected E3 expression, indicating that no viral components were required for E3 modification. Functionally, the SUMO modification repressed the transcriptional activation activity of E3 on two tested target genes, which suggests that this post-translational modification is likely to be biologically relevant to the viral life cycle .
The other known sumoylated vaccinia protein is the A40R gene product which is modified by SUMO as a single site, lysine 95 (Palacios et al. 2005). Like E3, modification of A40R does not require any viral products and is intrinsic to A40R in the presence of the sumoylation system. A unique feature of A40R is that its sumoylation is nearly quantitative, in contrast to most proteins for which only a small fraction exists in the sumoylated form at any time. The need for extensive sumoylation is likely explained by the observation that A40R is insoluble unless sumoylated. A40R lacking the SUMO acceptor site self-aggregates in the cytoplasm forming rod-shaped bodies rather than localizing to the viral mini-nuclei where viral DNA replication occurs. Nonetheless, because nonsumoylated A40R associates with the cytosolic side of the endoplasmic reticulum (ER), the authors speculated that during viral infection a small fraction of nonsumoylated A40R may function to bring ER membranes together and assist with membrane fusion around the viral replication foci .
3 RNA Viruses
Sumoylation was originally thought to be an exclusively nuclear event, so its role in virology was initially examined only for viruses with a nuclear phase (most DNA viruses and a few RNA viruses). As appreciation grew that sumoylation also occurs in the cytoplasm and various membrane compartments, greater attention was placed on searching for contributions of this post-translational modification system to human RNA virus biology. There are now numerous examples of sumoylated proteins from RNA viruses, both positive and negative stranded, as well as examples of RNA virus proteins that influence the sumoylation of other viral and/or host proteins. Two RNA viruses with prominent nuclear aspects, influenza and retroviruses , are the most intensively investigated for sumoylation, while for most other RNA viruses there are currently only limited reports concerning sumoylation. Consequently, detailed mechanistic and functional characterization of the ability of RNA viruses to use and/or manipulate the sumoylation system is still lacking for most RNA viruses. This section will review the current knowledge about the burgeoning list of RNA viruses with a sumoylation connection.
3.1 Retrovirus
Sumoylation has a role in the life cycle of two human retroviruses of the lentivirus subgroup, HIV and HTLV . Two HIV proteins, GAG (Jaber et al. 2009) and integrase (Zamborlini et al. 2011), interact with the sumoylation system. One of the GAG proteolytic products, p6, is sumoylated and this seems to be a negative regulatory event early in infection, perhaps as a host defense (Gurer et al. 2005). Alternatively, later in infection the full-length GAG binds Ubc9 , and reduction of Ubc9 by RNAi resulted in decreased incorporation of the ENV glycoprotein into released virions (Jaber et al. 2009). While not affecting total number of virions produced, the resulting virions were eight- to ten-fold less infectious. This effect did not require catalytically active Ubc9 so is unlikely to involve sumoylation of GAG. In contrast, integrase is sumoylated, but the functional consequence is unresolved as two groups have reported different effects. Zamborlini et al. reported that integrase has three SUMO consensus motifs and is multiply sumoylated (Zamborlini et al. 2011). They found that a triple mutant at the three consensus sites had normal stability and intracellular localization, but was replication defective at a step after reverse transcription but before integration, suggesting that sumoylation was necessary for this undefined step. However, Li et al. subsequently reported that overexpression of sumoylation caused integrase to accumulated in nuclear punctate bodies rather than be diffuse (Li et al. 2012b). They also observed that the degree of viral genome integration was inversely correlated with the levels of cellular sumoylation, leading to their suggestion that sumoylation is antiviral for HIV . While their studies did not directly confirm that these effects of sumoylation were mediated through integrase, it does raise questions about what sumoylation actually does for integrase activity. Interestingly, a recent report on a SENP inhibitor supports the antiviral nature of integrase sumoylation (Madu et al. 2015). Treatment of HIV infected cells with this inhibitor reduces the infectivity of progeny virions as the virions had defective integration during subsequent infection . The inhibitor effect could be abrogated by mutation of the SUMO acceptor sites in integrase, implying that sumoylation of integrase is a negative regulator and that the only active integrase molecules are ones that have been desumoylated by SENPs prior to virion packaging. These are clearly intriguing results that will need further study to clarify the precise mechanisms.
A second human lentivirus, human T-lymphotropic virus (HTLV) has one known sumoylated protein, Tax . Tax is a key regulator of the NF-ĸB pathway and is known to be modified by both ubiquitin and acetyl groups. Tax has also been reported as sumoylated for both HTLV1 (Lamsoul et al. 2005) and HTLV2 (Turci et al. 2009). Several groups have shown that Tax sumoylation is involved with its ability to accumulate in nuclear bodies (Kfoury et al. 2011; Lamsoul et al. 2005; Nasr et al. 2006). Sumoylation of Tax also appears to regulate its targeting of NEMO , a Tax partner, to centrosomes (Kfoury et al. 2011), and to be required for binding of the STUbL, RNF4 , , RNF4, which ubiquitinylates Tax to relocalize it from the nucleus to the cytoplasm (Fryrear et al. 2012). Thus, it appears that regulation of Tax intracellular localization is a complex process reflecting the contributions of several post-translation modification events. Unfortunately, the functional consequences of this complex intracellular regulation are still uncertain as initial reports that sumoylation is required for NF-ĸB activation (Lamsoul et al. 2005; Nasr et al. 2006) have been refuted in later studies (Bonnet et al. 2012; Pene et al. 2014). So as for HIV integrase, it is likely the sumoylation of Tax is important, but our understanding of the relevant mechanism is poor. A similar situation exists for Tax2 from the related HTLV-2B where Tax2 is sumoylated but the functional role of sumoylation is not clear (Journo et al. 2013; Turci et al. 2009, 2012).
3.2 Orthomyxovirus
A second RNA virus with significant impact by the sumoylation system is the orthomyxovirus, influenza A virus (IAV) . Five IAV proteins (NS1 , PB1, NP, M1, and NS2 ) are modified by sumoylation (Pal et al. 2011), and at least some functional characterization has been performed to evaluate the role of sumoylation on four of the five targets. The first identified IAV SUMO target was the NS1 protein (Pal et al. 2010). Two groups showed that sumoylation of NS1 was important for viral multiplication and the loss of sumoylation resulted in delayed growth (Santos et al. 2013; Xu et al. 2011). Both groups agreed that lysine 221 is a major SUMO acceptor site, but Santos et al. also found that lysine 70 was a SUMO site (Santos et al. 2013). The groups differed in that Xu et al. found that sumoylation stabilized NS1 (Xu et al. 2011) while Santos et al. found no effect on NS1 stability though they did report an effect on NS1 multimerization (Santos et al. 2013). The difference in the stability results may reflect that Santos et al. examined a double mutant of NS1 with both SUMO sites altered while Xu et al. only eliminated sumoylation at the major SUMO site. Lastly, Santos et al. also reported that sumoylated NS1 was greatly reduced in its ability to prevent interferon production during infection, and this might account for the delay replication (Santos et al. 2013).
For the remaining four sumoylated IAV proteins, there is modest information available about the role of sumoylation in their function, though three of the four are interrelated in their functions so sumoylation may have coordinating effects. The M1 protein is sumoylated at lysine 242, and this modification is required for effective interaction between M1 and the viral ribonucleoprotein (NP) (Wu et al. 2011). Without sumoylation of M1 and formation of this complex, viral maturation and assembly is greatly reduced which produces a strong reduction in viral yield. As cited above, NP is itself sumoylated with acceptor sites mapping to lysines 4 and 7; the lysine residue 7 is highly conserved and is critical for viral reproduction, so it may be the primary SUMO site (Han et al. 2014). The sumoylation deficient mutant of NP exhibits altered intracellular localization which may prevent effective interaction with M1 and contribute to its functional defect in viral replication. Further regulation of this process is mediated by the viral NS2 protein (Gao et al. 2015). NS2 interacts with the cellular AIMP2 protein, and the NS2-AIMP2 complex switches the modification of M1 lysine 242 from ubiquitin to SUMO which would facilitate the M1-NP interaction. Whether or not this requires the reported sumoylation of NS2 itself is unknown.
It is also of interest to note the influenza infection leads to a general dysregulation of cellular sumoylation (Pal et al. 2011). A detailed proteomic study found that influenza infection specifically targeted 63 host proteins for sumoylation while generally promoting global desumoylation (Domingues et al. 2015). The pattern of changes was distinct from other stress responses and involved at least ten potential antiviral factors, so likely reflects a specific influenza-induced process that reflects the battle between host containment of viral infections and viral attempts to interdict these defenses .
3.3 Filovirus
Among the human filoviruses, sumoylation has only been examined for the infamous Ebola virus (Chang et al. 2009). The viral VP35 protein binds Ubc9 , PIAS1, and IRF7. IRF7 is a transcription factor required for interferon transcription , and VP35 enhances the sumoylation of IRF7 which reduces its transactivation capacity and results in lower levels of interferon production. In this example, the virus appears to be exploiting the host sumoylation system to reduce the innate immune response .
3.4 Paramyxovirus
Parainfluenza virus commonly causes respiratory infections and is a frequent cause of childhood illness. The P protein of this virus is sumoylated at lysine 254 (Sun et al. 2011). This protein is a co-factor of viral RNA-dependent RNA polymerase , and mutation of the SUMO acceptor site produced a virus that grew with lower levels of viral RNA and reduced titers. However, the mutant P protein remained wild-type in most activities that were tested, so no biochemical or molecular mechanism to explain the effect of sumoylation was reported.
3.5 Rhabdovirus
This family includes the highly dangerous rabies virus as well as the more benign vesicular stomatitis virus (VSV). While sumoylation of rhabdoviral proteins has not yet been demonstrated, there is a recent report that sumoylation is involved in regulating both rabies and VSV infection (Maarifi et al. 2016). Stable exogenous SUMO expression blocked VSV viral mRNA synthesis, thus inhibiting all the downstream steps needed for viral reproduction. The host MxA protein is a known inhibitor of VSV transcription, and blocking MxA prevents the effect of increased SUMO expression. MxA protein becomes highly stable and accumulates under SUMO overexpression conditions, thus it appears that sumoylation is a host defense that restricts VSV by increasing the levels of this host protein that inhibits early steps in the VSV life cycle. In contrast to VSV, exogenous SUMO expression led to higher titers of rabies virus , an effect that was specific for SUMO3 and not SUMO1. It is known that MxA is not inhibitory for rabies virus, but SUMO3 expression did increase sumoylation of IRF3 which decreases its transcriptional activity and reduces the viral induction of interferon . Thus, for rabies virus increasing sumoylation has a positive effect on viral production, possibly through reduction of the interferon response. The opposing effects of sumoylation on two different viruses in the same family illustrate the complexity of the viral-host interplay and the diversity with which different viruses have evolved to interact with the sumoylation system.
3.6 Coronavirus
Human coronavirus are typically associated with mild respiratory diseases, but the SARS coronavirus is a highly lethal agent. There is only one report involving SARS and the sumoylation system, with Fan et al. demonstrating that the SARS nucleocapsid protein (N) bound to and co-localized with Ubc9 (Fan et al. 2006). Unfortunately, there was little functional characterization of this interaction and no follow up studies have been reported.
3.7 Flavivirus
Two members of this family, dengue virus and hepatitis C virus (HCV) , have known interactions with the sumoylation system. The dengue envelope protein binds Ubc9 and co-localize with Ubc9 on the cytoplasm ic side of the nuclear membrane (Chiu et al. 2007). Since overexpression of Ubc9 lead to reduced plaque formation, Ubc9 may be acting as part of the antiviral defense to restrict dengue infection at early stages. Conversely, silencing Ubc9 expression also reduced the replication of dengue, suggesting that the sumoylation system may have both positive and negative effects on dengue (Su et al. 2016). In this case the effect was related to sumoylation of the viral NS5 protein. Sumoylation of NS5 was required for viral RNA replication and suppression of the interferon production. Sumoylation of NS2 required a SIM motif in NS2 and led to increased stability which could account for both the observed effects on replication and interferon. For HCV , it was observed that SUMO1 is upregulated during infection and that knockdown of SUMO reduced viral replication (Akil et al. 2016). No specific viral target was identified, but the results suggest that sumoylation is a positive factor for this virus also .
3.8 Picornavirus
The encephalomyocarditis virus (EMCV) can cause disease in many mammals, likely including humans. The EMCV 3C protease enhances sumoylation of PML leading to association of PML with ND10s and subsequent proteasomal degradation of PML (El Mchichi et al. 2010). While the mechanism of 3C action on PML sumoylation is undetermined, this study does suggest that EMCV has evolved anti-PML activities as have many of the DNA viruses. A second picornavirus , enterovirus 71 is a more definite human pathogen that can cause serious central nervous system infections. EV71 also has a 3C protease, and in this case 3C was shown to bind Ubc9 and be sumoylated at lysine 52 (Chen et al. 2011). Sumoylation decreases protease activity of 3C and promotes 3C degradation. The K52R mutant of 3C that could not be sumoylated had increased viral levels during infection of cultured cells, consistent with sumoylation being a negative cellular regulatory event against EV71 .
3.9 Reovirus
Rotaviruses are important human pathogens of the reovirus family and are the causative agents of acute gastroenteritis. Campagna et al. found that sumoylation was a positive regulator for rotavirus and that five viral proteins were sumoylated, VP1, VP2, NSP2, VP6, and NSP5 (Campagna et al. 2013). VP1, VP2, and NSP2 were also able to bind SUMO noncovalently and likely have SIM motifs. The NSP5 protein was further investigated and two putative sumoylation sites were identified, lysines 19 and 82. A mutant NSP5 protein that was not sumoylated was mostly wild-type in its activities, but did display hyperphosphorylation and a defect in viroplasm formation when co-expressed with VP2. However, this defect was not sufficient to reduce viral replication so the biological importance of this modification to the overall viral life cycle is unclear .
3.10 Deltavirus
The hepatitis delta virus (HDV) is a serious pathogen that is always found in conjunction with hepatitis B virus (HBV) and which exacerbates HBV infections (Sureau and Negro 2016). HDV expresses a nuclear antigen (HDAg) that occurs in a short (S-HDAg) and long (L-HDAg) form. Multiple lysines in the short form are sumoylated, and this modification enhances production of viral genomic RNA and mRNA (Tseng et al. 2010).
4 Conclusion
The sumoylation system has been shown to be an important player in many biological processes, such as cellular differentiation, transcriptional regulation, and cell growth (Chymkowitch et al. 2015; Eifler and Vertegaal 2015). Perturbing sumoylation changes cellular response to diverse signaling pathways (Sharrocks 2006), so this system has huge potential as a target for pathogens to use to manipulate the host environment. Sumoylation also helps regulate host defense systems and appears in many cases to contribute to an “antiviral state” (Hannoun et al. 2016), so intracellular pathogens such as viruses may need to alter sumoylation to interfere with these defenses. The large body of evidence presented here indicates that SUMO and its enzymatic pathway play an important role in viral-host interactions for a large and growing number of DNA and RNA viruses. Numerous viral proteins are substrates for sumoylation with example of both positive and negative regulation of their activities. Thus, as with other types of host cell post-translational modification, viruses have clearly adapted to use sumoylation to regulate aspects of their life cycle. Additionally, there is an expanding list of viral proteins that can manipulate sumoylation, either a globally or in a substrate-specific manner. Given the wide range of cellular pathways affected by sumoylation, determining how these changes impact overall viral fitness require extensive experimentation, so in most reported cases the overall effect is still poorly understood. It is likely that many additional viral proteins that impact sumoylation await to be discovered. Identifying and understanding these global and specific viral effects on sumoylation will provide new insight into viral-host interactions and may highlight new targets for therapeutic treatment of viral infections.
References
Adamson AL (2005) Effects of SUMO-1 upon Epstein-Barr virus BZLF1 function and BMRF1 expression. Biochem Biophys Res Commun 336:22–28
Adamson AL, Kenney S (2001) Epstein-Barr virus immediate-early protein BZLF1 is SUMO-1 modified and disrupts promyelocytic leukemia bodies. J Virol 75:2388–2399
Ahn J-H, Xu Y, Jang W-J, Matunis MJ, Hayward GS (2001) Evaluation of interactions of human cytomegalovirus immediate-early IE2 regulatory protein with small ubiquitin-like modifiers and their conjugation enzyme Ubc9. J Virol 75:3859–3872
Akil A, Wedeh G, Zahid Mustafa M, Gassama-Diagne A (2016) SUMO1 depletion prevents lipid droplet accumulation and HCV replication. Arch Virol 161:141–148
Alonso A, Greenlee M, Matts J, Kline J, Davis KJ, Miller RK (2015) Emerging roles of sumoylation in the regulation of actin, microtubules, intermediate filaments, and septins. Cytoskeleton 72:305–339
Bailey D, O’Hare P (2002) Herpes simplex virus 1 ICPO co-localizes with a SUMO-specific protease. J Gen Virol 83:2951–2964
Bentz GL, Whitehurst CB, Pagano JS (2011) Epstein-Barr virus latent membrane protein 1 (LMP1) C-terminal-activating region 3 contributes to LMP1-mediated cellular migration via its interaction with Ubc9. J Virol 85:10144–10153
Bentz GL, Shackelford J, Pagano JS (2012) Epstein-Barr virus latent membrane protein 1 regulates the function of interferon regulatory factor 7 by inducing its sumoylation. J Virol 86:12251–12261
Bentz GL, Moss CR 2nd, Whitehurst CB, Moody CA, Pagano JS (2015) LMP1-induced sumoylation influences the maintenance of Epstein-Barr virus latency through KAP1. J Virol 89:7465–7477
Bergvall M, Melendy T, Archambault J (2013) The E1 proteins. Virology 445:35–56
Berk AJ (2005) Recent lessons in gene expression, cell cycle control, and cell biology from adenovirus. Oncogene 24:7673–7685
Berndt A, Hofmann-Winkler H, Tavalai N, Hahn G, Stamminger T (2009) Importance of covalent and noncovalent SUMO interactions with the major human cytomegalovirus transactivator IE2p86 for viral infection. J Virol 83:12881–12894
Berns KI, Linden RM (1995) The cryptic life style of adeno-associated virus. BioEssays 17:237–245
Bischof O, Schwamborn K, Martin N, Werner A, Sustmann C, Grosschedl R, Dejean A (2006) The E3 SUMO ligase PIASy is a regulator of cellular senescence and apoptosis. Mol Cell 22:783–794
Boggio R, Colombo R, Hay RT, Draetta GF, Chiocca S (2004) A mechanism for inhibiting the SUMO pathway. Mol Cell 16:549–561
Boggio R, Passafaro A, Chiocca S (2007) Targeting SUMO E1 to ubiquitin ligases – a viral strategy to counteract sumoylation. J Biol Chem 282:15376–15382
Bonnet A, Randrianarison-Huetz V, Nzounza P, Nedelec M, Chazal M, Waast L, Pene S, Bazarbachi A, Mahieux R, Benit L, Pique C (2012) Low nuclear body formation and Tax SUMOylation do not prevent NF-kappaB promoter activation. Retrovirology 9:77
Boutell C, Everett RD (2013) Regulation of alphaherpesvirus infections by the ICP0 family of proteins. J Gen Virol 94:465–481
Boutell C, Sadis S, Everett RD (2002) Herpes simplex virus type 1 immediate-early protein ICP0 and its isolated RING finger domain act as ubiquitin E3 ligases in vitro. J Virol 76:841–850
Boutell C, Orr A, Everett RD (2003) PML residue lysine 160 is required for the degradation of PML induced by herpes simplex virus type 1 regulatory protein ICP0. J Virol 77:8686–8694
Boutell C, Cuchet-Lourenco D, Vanni E, Orr A, Glass M, McFarlane S, Everett RD (2011) A viral ubiquitin ligase has substrate preferential SUMO targeted ubiquitin ligase activity that counteracts intrinsic antiviral defence. PLoS Pathog 7:e1002245
Bridges RG, Sohn SY, Wright J, Leppard KN, Hearing P (2016) The adenovirus E4-ORF3 protein stimulates SUMOylation of general transcription factor TFII-I to direct proteasomal degradation. mBio 7:e02184–e02115
Brown JR, Conn KL, Wasson P, Charman M, Tong L, Grant K, McFarlane S, Boutell C (2016) SUMO ligase protein inhibitor of activated STAT1 (PIAS1) is a constituent promyelocytic leukemia nuclear body protein that contributes to the intrinsic antiviral immune response to herpes simplex virus 1. J Virol 90:5939–5952
Bund T, Spoden GA, Koynov K, Hellmann N, Boukhallouk F, Arnold P, Hinderberger D, Florin L (2014) An L2 SUMO interacting motif is important for PML localization and infection of human papillomavirus type 16. Cell Microbiol 16:1179–1200
Burck C, Mund A, Berscheminski J, Kieweg L, Muncheberg S, Dobner T, Schreiner S (2016) KAP1 Is a host restriction factor that promotes human adenovirus E1B-55K SUMO modification. J Virol 90:930–946
Cai Q, Cai S, Zhu C, Verma SC, Choi JY, Robertson ES (2013) A unique SUMO-2-interacting motif within LANA is essential for KSHV latency. PLoS Pathog 9:e1003750
Calderwood MA, Holthaus AM, Johannsen E (2008) The Epstein-Barr virus LF2 protein inhibits viral replication. J Virol 82:8509–8519
Callegari S, Gastaldello S, Faridani OR, Masucci MG (2014) Epstein-Barr virus encoded microRNAs target SUMO-regulated cellular functions. FEBS J 281:4935–4950
Campagna M, Marcos-Villar L, Arnoldi F, de la Cruz-Herrera CF, Gallego P, Gonzalez-Santamaria J, Gonzalez D, Lopitz-Otsoa F, Rodriguez MS, Burrone OR, Rivas C (2013) Rotavirus viroplasm proteins interact with the cellular SUMOylation system: implications for viroplasm-like structure formation. J Virol 87:807–817
Campbell M, Izumiya Y (2012) Post-translational modifications of Kaposi’s sarcoma-associated herpesvirus regulatory proteins – SUMO and KSHV. Front Microbiol 3:31
Chang PC, Kung HJ (2014) SUMO and KSHV replication. Cancers 6:1905–1924
Chang LK, Lee YH, Cheng TS, Hong YR, Lu PJ, Wang JJ, Wang WH, Kuo CW, Li SSL, Liu ST (2004) Post-translational modification of Rta of Epstein-Barr virus by SUMO-1. J Biol Chem 279:38803–38812
Chang LK, Liu ST, Kuo CW, Wang WH, Chuang JY, Bianchi E, Hong YR (2008) Enhancement of transactivation activity of Rta of Epstein-Barr virus by RanBPM. J Mol Biol 379:231–242
Chang TH, Kubota T, Matsuoka M, Jones S, Bradfute SB, Bray M, Ozato K (2009) Ebola Zaire virus blocks type I interferon production by exploiting the host SUMO modification machinery. PLoS Pathog 5:493–493
Chang PC, Izumiya Y, Wu CY, Fitzgerald LD, Campbell M, Ellison TJ, Lam KS, Luciw PA, Kung HJ (2010) Kaposi’s sarcoma-associated herpesvirus (KSHV) encodes a SUMO E3 ligase that is SIM-dependent and SUMO-2/3-specific. J Biol Chem 285:5266–5273
Chang PC, Cheng CY, Campbell M, Yang YC, Hsu HW, Chang TY, Chu CH, Lee YW, Hung CL, Lai SM, Tepper CG, Hsieh WP, Wang HW, Tang CY, Wang WC, Kung HJ (2013) The chromatin modification by SUMO-2/3 but not SUMO-1 prevents the epigenetic activation of key immune-related genes during Kaposi’s sarcoma associated herpesvirus reactivation. BMC Genomics 14:824
Chang PC, Campbell M, Robertson ES (2016) Human oncogenic herpesvirus and post-translational modifications – phosphorylation and SUMOylation. Front Microbiol 7:962
Chen SC, Chang LY, Wang YW, Chen YC, Weng KF, Shih SR, Shih HM (2011) Sumoylation-promoted Enterovirus 71 3C Degradation correlates with a reduction in viral replication and cell apoptosis. J Biol Chem 286:31373–31384
Chiang CM, Dong G, Broker TR, Chow LT (1992) Control of human papillomavirus type 11 origin of replication by the E2 family of transcription regulatory proteins. J Virol 66:5224–5231
Ching W, Dobner T, Koyuncu E (2012) The human adenovirus type 5 E1B 55-kilodalton protein is phosphorylated by protein kinase CK2. J Virol 86:2400–2415
Chiocca S, Kurzbauer R, Schaffner G, Baker A, Mautner V, Cotten M (1996) The complete DNA sequence and genomic organization of the avian adenovirus CELO. J Virol 70:2939–2949
Chiu MW, Shih HM, Yang TH, Yang YL (2007) The type 2 dengue virus envelope protein interacts with small ubiquitin-like modifier-1 (SUMO-1) conjugating enzyme 9 (Ubc9). J Biomed Sci 14:429–444
Chymkowitch P, Nguea PA, Enserink JM (2015) SUMO-regulated transcription: challenging the dogma. BioEssays 37:1095–1105
Colombo R, Boggio R, Seiser C, Draetta GF, Chiocca S (2002) The adenovirus protein Gam1 interferes with sumoylation of histone deacetylase 1. EMBO Rep 3:1062–1068
Conn KL, Wasson P, McFarlane S, Tong L, Brown JR, Grant KG, Domingues P, Boutell C (2016) Novel role for protein inhibitor of activated STAT 4 (PIAS4) in the restriction of herpes simplex virus 1 by the cellular intrinsic antiviral immune response. J Virol 90:4807–4826
Cuchet-Lourenco D, Boutell C, Lukashchuk V, Grant K, Sykes A, Murray J, Orr A, Everett RD (2011) SUMO pathway dependent recruitment of cellular repressors to herpes simplex virus type 1 genomes. PLoS Pathog 7:e1002123
Cuchet-Lourenco D, Vanni E, Glass M, Orr A, Everett RD (2012) Herpes simplex virus 1 ubiquitin ligase ICP0 interacts with PML isoform I and induces its SUMO-independent degradation. J Virol 86:11209–11222
Demeret C, Desaintes C, Yaniv M, Thierry F (1997) Different mechanisms contribute to the E2-mediated transcriptional repression of human papillomavirus type 18 viral oncogenes. J Virol 71:9343–9349
Deng ZY, Wan MM, Sui GC (2007) PIASy-mediated sumoylation of Yin Yang 1 depends on their interaction but not the RING finger. Mol Cell Biol 27:3780–3792
Deyrieux AF, Rosas-Acosta G, Ozbun MA, Wilson VG (2007) Sumoylation dynamics during keratinocyte differentiation. J Cell Sci 120:125–136
Domingues P, Golebiowski F, Tatham MH, Lopes AM, Taggart A, Hay RT, Hale BG (2015) Global reprogramming of host SUMOylation during influenza virus infection. Cell Rep 13:1467–1480
Doorbar J (2006) Molecular biology of human papillomavirus infection and cervical cancer. Clin Sci 110:525–541
Dyson NJ (2016) RB1: a prototype tumor suppressor and an enigma. Genes Dev 30:1492–1502
Eifler K, Vertegaal AC (2015) SUMOylation-mediated regulation of cell cycle progression and cancer. Trends Biochem Sci 40:779–793
El Mchichi B, Regad T, Maroui MA, Rodriguez MS, Aminev A, Gerbaud S, Escriou N, Dianoux L, Chelbi-Alix MK (2010) SUMOylation promotes PML degradation during encephalomyocarditis virus infection. J Virol 84:11634–11645
Endter C, Kzhyshkowska J, Stauber R, Dobner T (2001) SUMO-1 modification required for transformation by adenovirus type 5 early region 1B 55-kDa oncoprotein. Proc Natl Acad Sci U S A 98:11312–11317
Everett RD, Freemont P, Saitoh H, Dasso M, Orr A, Kathoria M, Parkinson J (1998) The disruption of ND10 during herpes simplex virus infection correlates with the Vmw110- and proteasome-dependent loss of several PML isoforms. J Virol 72:6581–6591
Everett RD, Rechter S, Papior P, Tavalai N, Stamminger T, Orr A (2006) PML contributes to a cellular mechanism of repression of herpes simplex virus type 1 infection that is inactivated by ICP0. J Virol 80:7995–8005
Everett RD, Parada C, Gripon P, Sirma H, Orr A (2008) Replication of ICP0-null mutant herpes simplex virus type 1 is restricted by both PML and Sp100. J Virol 82:2661–2672
Everett RD, Boutell C, McNair C, Grant L, Orr A (2010) Comparison of the biological and biochemical activities of several members of the alphaherpesvirus ICP0 family of proteins. J Virol 84:3476–3487
Everett RD, Bell AJ, Lu Y, Orr A (2013a) The replication defect of ICP0-null mutant herpes simplex virus 1 can be largely complemented by the combined activities of human cytomegalovirus proteins IE1 and pp71. J Virol 87:978–990
Everett RD, Boutell C, Hale BG (2013b) Interplay between viruses and host sumoylation pathways. Nat Rev Microbiol 11:400–411
Everett RD, Boutell C, Pheasant K, Cuchet-Lourenco D, Orr A (2014) Sequences related to SUMO interaction motifs in herpes simplex virus 1 protein ICP0 act cooperatively to stimulate virus infection. J Virol 88:2763–2774
Fan Z, Zhuo Y, Tan XY, Zhou Z, Yuan JG, Qiang BQ, Yan JH, Peng XZ, Gao GF (2006) SARS-CoV nucleocapsid protein binds to hUbc9, a ubiquitin conjugating enzyme of the sumoylation system. J Med Virol 78:1365–1373
Fradet-Turcotte A, Brault K, Titolo S, Howley PM, Archambault J (2009) Characterization of papillomavirus E1 helicase mutants defective for interaction with the SUMO-conjugating enzyme Ubc9. Virology 395:190–201
Fryrear KA, Guo X, Kerscher O, Semmes OJ (2012) The Sumo-targeted ubiquitin ligase RNF4 regulates the localization and function of the HTLV-1 oncoprotein Tax. Blood 119:1173–1181
Galloway DA, Laimins LA (2015) Human papillomaviruses: shared and distinct pathways for pathogenesis. Curr Opin Virol 14:87–92
Gan J, Wang C, Jin Y, Guo Y, Xu F, Zhu Q, Ding L, Shang H, Wang J, Wei F, Cai Q, Robertson ES (2015) Proteomic profiling identifies the SIM-associated complex of KSHV-encoded LANA. Proteomics 15:2023–2037
Gao S, Wu J, Liu RY, Li J, Song L, Teng Y, Sheng C, Liu D, Yao C, Chen H, Jiang W, Chen S, Huang W (2015) Interaction of NS2 with AIMP2 facilitates the switch from ubiquitination to SUMOylation of M1 in influenza A virus-infected cells. J Virol 89:300–311
Gilden D., Nagel, M., Cohrs, R., Mahalingam, R. Baird, N. 2015 Varicella zoster virus in the nervous system. F1000Research 4(F1000 Faculty Rev):1356. doi:10.12688/f1000research.7153.1
Girdwood D, Bumpass D, Vaughan OA, Thain A, Anderson LA, Snowden AW, Garcia-Wilson E, Perkins ND, Hay RT (2003) p300 transcriptional repression is mediated by SUMO modification. Mol Cell 11:1043–1054
Gonzalez-Santamaria J, Campagna M, Garcia MA, Marcos-Villar L, Gonzalez D, Gallego P, Lopitz-Otsoa F, Guerra S, Rodriguez MS, Esteban M, Rivas C (2011) Regulation of vaccinia virus E3 protein by small ubiquitin-like modifier proteins. J Virol 85:12890–12900
Gravel A, Gosselin J, Flamand L (2002) Human herpesvirus 6 immediate-early 1 protein is a sumoylated nuclear phosphoprotein colocalizing with promyelocytic leukemia protein-associated nuclear bodies. J Biol Chem 277:19679–19687
Gravel A, Dion V, Cloutier N, Gosselin J, Flamand L (2004) Characterization of human herpesvirus 6 variant B immediate-early 1 protein modifications by small ubiquitin-related modifiers. J Gen Virol 85:1319–1328
Grinde B (2013) Herpesviruses: latency and reactivation – viral strategies and host response. J Oral Microbiol 5. doi:10.3402/jom.v5i0.22766
Gunther T, Schreiner S, Dobner T, Tessmer U, Grundhoff A (2014) Influence of ND10 components on epigenetic determinants of early KSHV latency establishment. PLoS Pathog 10:e1004274
Gurer C, Berthoux L, Luban J (2005) Covalent modification of human immunodeficiency virus type 1 p6 by SUMO-1. J Virol 79:910–917
Hadaschik D, Hinterkeuser K, Oldak M, Pfister HJ, Smola-Hess S (2003) The papillomavirus E2 protein binds to and synergizes with C/EBP factors involved in keratinocyte differentiation. J Virol 77:5253–5265
Hagemeier SR, Dickerson SJ, Meng Q, Yu XM, Mertz JE, Kenney SC (2010) Sumoylation of the Epstein-Barr virus BZLF1 protein inhibits its transcriptional activity and is regulated by the virus-encoded protein kinase. J Virol 84:4383–4394
Han Q, Chang C, Li L, Klenk C, Cheng J, Chen Y, Xia N, Shu Y, Chen Z, Gabriel G, Sun B, Xu K (2014) Sumoylation of influenza A virus nucleoprotein is essential for intracellular trafficking and virus growth. J Virol 88:9379–9390
Hannoun Z, Maarifi G, Chelbi-Alix MK (2016) The implication of SUMO in intrinsic and innate immunity. Cytokine Growth Factor Rev 29:3–16
Hastie E, Samulski RJ (2015) Adeno-associated virus at 50: a golden anniversary of discovery, research, and gene therapy success--a personal perspective. Hum Gene Ther 26:257–265
Hateboer G, Hijmans EM, Nooij JB, Schlenker S, Jentsch S, Bernards R (1996) mUBC9, a novel adenovirus E1A-interacting protein that complements a yeast cell cycle defect. J Biol Chem 271:25906–25911
Heaton PR, Deyrieux AF, Bian XL, Wilson VG (2011) HPV E6 proteins target Ubc9, the SUMO conjugating enzyme. Virus Res 158:199–208
Heaton PR, Santos A, Rosas-Acosta G, Wilson VG (2012) Analysis of global sumoylation changes occurring during keratinocyte differentiation. PLoS One 7:e30165
Hebner CM, Laimins LA (2006) Human papillomaviruses: basic mechanisms of pathogenesis and oncogenicity. Rev Med Virol 16:83–97
Heilmann AM, Calderwood MA, Johannsen E (2010) Epstein-Barr virus LF2 protein regulates viral replication by altering Rta subcellular localization. J Virol 84:9920–9931
Hietakangas V, Anckar J, Blomster HA, Fujimoto M, Palvimo JJ, Nakai A, Sistonen L (2006) PDSM, a motif for phosphorylation-dependent SUMO modification. Proc Natl Acad Sci U S A 103:45–50
Higginbotham JM, O’Shea CC (2015) Adenovirus E4-ORF3 targets PIAS3 and together with E1B-55K remodels SUMO interactions in the nucleus and at virus genome replication domains. J Virol 89:10260–10272
Hilgarth RS, Murphy LA, Skaggs HS, Wilkerson DC, Xing HY, Sarge KD (2004) Regulation and function of SUMO modification. J Biol Chem 279:53899–53902
Hislop AD (2015) Early virological and immunological events in Epstein-Barr virus infection. Curr Opin Virol 15:75–79
Hofmann H, Floss S, Stamminger T (2000) Covalent modification of the transactivator protein IE2-p86 of human cytomegalovirus by conjugation to the ubiquitin-homologous proteins SUMO-1 and hSMT3b. J Virol 74:2510–2524
Hollingworth R, Grand RJ (2015) Modulation of DNA damage and repair pathways by human tumour viruses. Viruses 7:2542–2591
Holscher C, Sonntag F, Henrich K, Chen Q, Beneke J, Matula P, Rohr K, Kaderali L, Beil N, Erfle H, Kleinschmidt JA, Muller M (2015) The SUMOylation pathway restricts gene transduction by adeno-associated viruses. PLoS Pathog 11:e1005281
Huh YH, Kim YE, Kim ET, Park JJ, Song MJ, Zhu H, Hayward GS, Ahn JH (2008) Binding STAT2 by the acidic domain of human cytomegalovirus IE1 promotes viral growth and is negatively regulated by SUMO. J Virol 82:10444–10454
Huibregtse JM, Scheffner M, Howley PM (1994) E6-AP directs the HPV E6-dependent inactivation of p53 and is representative of a family of structurally and functionally related proteins. Cold Spring Harb Symp Quant Biol 59:237–246
Hwang J, Kalejta RF (2009) Human cytomegalovirus protein pp71 induces Daxx SUMOylation. J Virol 83:6591–6598
Imperiale MJ, Akusjnarvi G, Leppard KN (1995) Post-transcriptional control of adenovirus gene expression. Curr Top Microbiol Immunol 199:139–171
Iyengar S, Farnham PJ (2011) KAP1 protein: an enigmatic master regulator of the genome. J Biol Chem 286:26267–26276
Izumiya Y, Ellison TJ, Yeh ETH, Jung JU, Luciw PA, Kung HJ (2005) Kaposi’s sarcoma-associated herpesvirus K-bZIP represses gene transcription via SUMO modification. J Virol 79:9912–9925
Izumiya Y, Izumiya C, Van Geelen A, Wang DH, Lam KS, Luciw PA, Kung HJ (2007) Kaposi’s sarcoma-associated herpesvirus-encoded protein kinase and its interaction with K-bZIP. J Virol 81:1072–1082
Izumiya Y, Kobayashi K, Kim KY, Pochampalli M, Izumiya C, Shevchenko B, Wang DH, Huerta SB, Martinez A, Campbell M, Kung HJ (2013) Kaposi’s sarcoma-associated herpesvirus K-Rta exhibits SUMO-targeting ubiquitin ligase (STUbL) like activity and is essential for viral reactivation. PLoS Pathog 9:e1003506
Jaber T, Bohl CR, Lewis GL, Wood C, West JT, Weldon RA (2009) Human Ubc9 contributes to production of fully infectious human immunodeficiency virus type 1 virions. J Virol 83:10448–10459
Journo C, Bonnet A, Favre-Bonvin A, Turpin J, Vinera J, Cote E, Chevalier SA, Kfoury Y, Bazarbachi A, Pique C, Mahieux R (2013) Human T cell leukemia virus type 2 tax-mediated NF-kappaB activation involves a mechanism independent of Tax conjugation to ubiquitin and SUMO. J Virol 87:1123–1136
Kang H, Kim ET, Lee HR, Park JJ, Go YY, Choi CY, Ahn JH (2006) Inhibition of SUMO-independent PML oligomerization by the human cytomegalovirus IE1 protein. J Gen Virol 87:2181–2190
Kfoury Y, Setterblad N, El-Sabban M, Zamborlini A, Dassouki Z, El Hajj H, Hermine O, Pique C, De The H, Saib A, Bazarbachi A (2011) Tax ubiquitylation and SUMOylation control the dynamic shuttling of Tax and NEMO between Ubc9 nuclear bodies and the centrosome. Blood 117:190–199
Kim J, Cantwell CA, Johnson PF, Pfarr CM, Williams SC (2002) Transcriptional activity of CCAAT/enhancer-binding proteins is controlled by a conserved inhibitory domain that is a target for sumoylation. J Biol Chem 277:38037–38044
Kim ET, Kim YE, Huh YH, Ahn JH (2010) Role of noncovalent SUMO binding by the human cytomegalovirus IE2 transactivator in lytic growth. J Virol 84:8111–8123
Kim ET, Kim YE, Kim YJ, Lee MK, Hayward GS, Ahn JH (2014) Analysis of human cytomegalovirus-encoded SUMO targets and temporal regulation of SUMOylation of the immediate-early proteins IE1 and IE2 during infection. PLoS One 9:e103308
Kindsmuller K, Groitl P, Hartl B, Blanchette P, Hauber J, Dobner T (2007) Intranuclear targeting and nuclear export of the adenovirus E1B-55K protein are regulated by SUMO1 conjugation. Proc Natl Acad Sci U S A 104:6684–6689
Knipscheer P, van Dijk WJ, Olsen JV, Mann M, Sixma TK (2007) Noncovalent interaction between Ubc9 and SUMO promotes SUMO chain formation. EMBO J 26:2797–2807
Lamsoul I, Lodewick J, Lebrun S, Brasseur R, Burny A, Gaynor RB, Bex F (2005) Exclusive ubiquitination and sumoylation on overlapping lysine residues mediate NF-kappa B activation by the human T-cell leukemia virus tax oncoprotein. Mol Cell Biol 25:10391–10406
Lanfranca MP, Mostafa HH, Davido DJ (2014) HSV-1 ICP0: an E3 ubiquitin ligase that counteracts host intrinsic and innate immunity. Cells 3:438–454
Laura MV, de la Cruz-Herrera CF, Ferreiros A, Baz-Martinez M, Lang V, Vidal A, Munoz-Fontela C, Rodriguez MS, Collado M, Rivas C (2015) KSHV latent protein LANA2 inhibits sumo2 modification of p53. Cell Cycle 14:277–282
Ledl A, Schmidt D, Muller S (2005) Viral oncoproteins E1A and E7 and cellular LxCxE proteins repress SUMO modification of the retinoblastoma tumor suppressor. Oncogene 24:3810–3818
Lee HR, Ahn JH (2004) Sumoylation of the major immediate-early IE2 protein of human cytomegalovirus Towne strain is not required for virus growth in cultured human fibroblasts. J Gen Virol 85:2149–2154
Lee KY, Broker TR, Chow LT (1998) Transcription factor YY1 represses cell-free replication from human papillomavirus origins. J Virol 72:4911–4917
Lee HR, Kim DJ, Lee JM, Choi CY, Ahn BY, Hayward GS, Ahn JH (2004) Ability of the human cytomegalovirus IE1 protein to modulate sumoylation of PML correlates with its functional activities in transcriptional regulation and infectivity in cultured fibroblast cells. J Virol 78:6527–6542
Lee SJ, Yang A, Wu TC, Hung CF (2016) Immunotherapy for human papillomavirus-associated disease and cervical cancer: review of clinical and translational research. J Gynecol Oncol 27:e51
Lefkowitz EJ, Wang C, Upton C (2006) Poxviruses: past, present and future. Virus Res 117:105–118
Lefort S, Gravel A, Flamand L (2010) Repression of interferon-alpha stimulated genes expression by Kaposi’s sarcoma-associated herpesvirus K-bZIP protein. Virology 408:14–30
Leppard KN, Everett RD (1999) The adenovirus type 5 E1b 55K and E4 Orf3 proteins associate in infected cells and affect ND10 components. J Gen Virol 80:997–1008
Lethbridge KJ, Scott GE, Leppard KN (2003) Nuclear matrix localization and SUMO-1 modification of adenovirus type 5 E1b 55K protein are controlled by E4 Orf6 protein. J Gen Virol 84:259–268
Li HP, Chang YS (2003) Epstein-Barr virus latent membrane protein 1: structure and functions. J Biomed Sci 10:490–504
Li RW, Wang LY, Liao GL, Guzzo CM, Matunis MJ, Zhu H, Hayward SD (2012a) SUMO binding by the Epstein-Barr virus protein kinase BGLF4 is crucial for BGLF4 function. J Virol 86:5412–5421
Li Z, Wu S, Wang J, Li W, Lin Y, Ji C, Xue J, Chen J (2012b) Evaluation of the interactions of HIV-1 integrase with small ubiquitin-like modifiers and their conjugation enzyme Ubc9. Int J Mol Med 30:1053–1060
Lin J, Johannsen E, Robertson E, Kieff E (2002) Epstein-Barr virus nuclear antigen 3C putative repression domain mediates coactivation of the LMP1 promoter with EBNA-2. J Virol 76:232–242
Lin X, Sun BH, Liang M, Liang YY, Gast A, Hildebrand J, Brunicardi FC, Melchior F, Feng XH (2003) Opposed regulation of corepressor CtBP by SUMOylation and PDZ binding. Mol Cell 11:1389–1396
Lisowski L, Tay SS, Alexander IE (2015) Adeno-associated virus serotypes for gene therapeutics. Curr Opin Pharmacol 24:59–67
Liu ST, Wang WH, Hong YR, Chuang JY, Lu PJ, Chang LK (2006) Sumoylation of Rta of Epstein-Barr virus is preferentially enhanced by PIASx beta. Virus Res 119:163–170
Lynch JP 3rd, Fishbein M, Echavarria M (2011) Adenovirus. Semin Respir Crit Care Med 32:494–511
Maarifi G, Hannoun Z, Geoffroy MC, El Asmi F, Zarrouk K, Nisole S, Blondel D, Chelbi-Alix MK (2016) MxA mediates SUMO-induced resistance to vesicular stomatitis virus. J Virol 90:6598–6610
Madkan VK, Cook-Norris RH, Steadman MC, Arora A, Mendoza N, Tyring SK (2007) The oncogenic potential of human papillomaviruses: a review on the role of host genetics and environmental cofactors. Br J Dermatol 157:228–241
Madu IG, Li S, Li B, Li H, Chang T, Li YJ, Vega R, Rossi J, Yee JK, Zaia J, Chen Y (2015) A novel class of HIV-1 antiviral agents targeting HIV via a SUMOylation-dependent mechanism. Sci Report 5:17808
Manning Fox JE, Hajmrle C, Macdonald PE (2012) Novel roles of SUMO in pancreatic beta-cells: thinking outside the nucleus. Can J Physiol Pharmacol 90:765–770
Marcos-Villar L, Lopitz-Otsoa F, Gallego P, Munoz-Fontela C, Gonzalez-Santamaria J, Campagna M, Shou-Jiang G, Rodriguez MS, Rivas C (2009) Kaposi’s sarcoma-associated herpesvirus protein LANA2 disrupts PML oncogenic domains and inhibits PML-mediated transcriptional repression of the survivin gene. J Virol 83:8849–8858
Marcos-Villar L, Campagna M, Lopitz-Otsoa F, Gallego P, Gonzalez-Santamaria J, Gonzalez D, Rodriguez MS, Rivas C (2011) Covalent modification by SUMO is required for efficient disruption of PML oncogenic domains by Kaposi’s sarcoma-associated herpesvirus latent protein LANA2. J Gen Virol 92:188–194
Marcos-Villar L, Gallego P, Munoz-Fontela C, de la Cruz-Herrera CF, Campagna M, Gonzalez D, Lopitz-Otsoa F, Rodriguez MS, Rivas C (2014) Kaposi’s sarcoma-associated herpesvirus lana2 protein interacts with the pocket proteins and inhibits their sumoylation. Oncogene 33:495–503
Marusic MB, Mencin N, Licen M, Banks L, Grm HST (2010) Modification of human papillomavirus minor capsid protein L2 by sumoylation. J Virol 84:11585–11589
Mattoscio D, Segre CV, Chiocca S (2013) Viral manipulation of cellular protein conjugation pathways: the SUMO lesson. World J Virol 2:79–90
McBride AA (2013) The papillomavirus E2 proteins. Virology 445:57–79
Moriuchi H, Moriuchi M, Smith HA, Straus SE, Cohen JI (1992) Varicella-zoster virus open reading frame 61 protein is functionally homologous to herpes simplex virus type 1 ICP0. J Virol 66:7303–7308
Muller S, Dejean A (1999) Viral immediate-early proteins abrogate the modification by SUMO-1 of PML and Sp100 proteins, correlating with nuclear body disruption. J Virol 73:5137–5143
Muller S, Dobner T (2008) The adenovirus E1B-55K oncoprotein induces SUMO modification of p53. Cell Cycle 7:754–758
Munger K, Basile JR, Duensing S, Eichten A, Gonzalez SL, Grace M, Zacny VL (2001) Biological activities and molecular targets of the human papillomavirus E7 oncoprotein. Oncogene 20:7888–7898
Murata T, Hotta N, Toyama S, Nakayama S, Chiba S, Isomura H, Ohshima T, Kanda T, Tsurumi T (2010) Transcriptional repression by sumoylation of Epstein-Barr virus BZLF1 protein correlates with association of histone deacetylase. J Biol Chem 285:23925–23935
Nasr R, Chiari E, El-Sabban M, Mahieux R, Kfoury Y, Abdulhay M, Yazbeck V, Hermine O, de The H, Pique C, Bazarbachi A (2006) Tax ubiquitylation and sumoylation control critical cytoplasmic and nuclear steps of NF-kappa B activation. Blood 107:4021–4029
Nevels M, Brune W, Shenk T (2004) SUMOylation of the human cytomegalovirus 72-kilodalton IE1 protein facilitates expression of the 86-kilodalton IE2 protein and promotes viral replication. J Virol 78:7803–7812
Nguyen HP, Ramirez-Fort MK, Rady PL (2014) The biology of human papillomaviruses. Curr Probl Dermatol 45:19–32
Pal S, Rosas JM, Rosas-Acosta G (2010) Identification of the non-structural influenza A viral protein NS1A as a bona fide target of the Small Ubiquitin-like MOdifier by the use of dicistronic expression constructs. J Virol Methods 163:498–504
Pal S, Santos A, Rosas JM, Ortiz-Guzman J, Rosas-Acosta G (2011) Influenza A virus interacts extensively with the cellular SUMOylation system during infection. Virus Res 158:12–27
Palacios S, Perez LH, Welsch S, Schleich S, Chmielarska K, Melchior F, Locker JK (2005) Quantitative SUMO-1 modification of a vaccinia virus protein is required for its specific localization and prevents its self-association. Mol Biol Cell 16:2822–2835
Pan W, Datta A, Adami GR, Raychaudhuri P, Bagchi S (2003) P19ARF inhibits the functions of the HPV16 E7 oncoprotein. Oncogene 22:5496–5503
Pene S, Waast L, Bonnet A, Benit L, Pique C (2014) A non-SUMOylated tax protein is still functional for NF-kappaB pathway activation. J Virol 88:10655–10661
Pennella MA, Liu Y, Woo JL, Kim CA, Berk AJ (2010) Adenovirus E1B 55-kilodalton protein is a p53-SUMO1 E3 ligase that represses p53 and stimulates its nuclear export through interactions with promyelocytic leukemia nuclear bodies. J Virol 84:12210–12225
Rangasamy D, Wilson VG (2000) Bovine papillomavirus E1 protein is sumoylated by the host cell Ubc9 protein. J Biol Chem 275:30487–30495
Rangasamy D, Woytek K, Khan SA, Wilson VG (2000) SUMO-1 modification of bovine papillomavirus E1 protein is required for intranuclear accumulation. J Biol Chem 275:37999–38004
Rizos H, Woodruff S, Kefford RF (2005) p14ARF interacts with the SUMO-conjugating enzyme Ubc9 and promotes the Sumoylation of its binding partners. Cell Cycle 4:597–603
Roberts KL, Smith GL (2008) Vaccinia virus morphogenesis and dissemination. Trends Microbiol 16:472–479
Robertson ES, Lin J, Kieff E (1996) The amino-terminal domains of Epstein-Barr virus nuclear proteins 3A, 3B, and 3C interact with RBPJ(kappa). J Virol 70:3068–3074
Rogan S, Heaphy S (2000) The vaccinia virus E3L protein interacts with SUMO-1 and ribosomal protein L23a in a yeast two hybrid assay. Virus Genes 21:193–195
Roman A, Munger K (2013) The papillomavirus E7 proteins. Virology 445:138–168
Rosas-Acosta G, Wilson VG (2008) Identification of a nuclear export signal sequence for bovine papillomavirus E1 protein. Virology 373:149–162
Rosas-Acosta G, Langereis MA, Deyrieux A, Wilson VG (2005) Proteins of the PIAS family enhance the sumoylation of the papillomavirus E1 protein. Virology 331:190–203
Rosendorff A, Illanes D, David G, Lin J, Kieff E, Johannsen E (2004) EBNA3C coactivation with EBNA2 requires a SUMO homology domain. J Virol 78:367–377
Sadanari H, Yamada R, Ohnishi K, Matsubara K, Tanaka J (2005) SUMO-1 modification of the major immediate-early (IE) 1 and 2 proteins of human cytomegalovirus is regulated by different mechanisms and modulates the intracellular localization of the IE1, but not IE2, protein. Arch Virol 150:1763–1782
Santos A, Pal S, Chacon J, Meraz K, Gonzalez J, Prieto K, Rosas-Acosta G (2013) SUMOylation affects the interferon blocking activity of the influenza A nonstructural protein NS1 without affecting its stability or cellular localization. J Virol 87:5602–5620
Scherer M, Reuter N, Wagenknecht N, Otto V, Sticht H, Stamminger T (2013) Small ubiquitin-related modifier (SUMO) pathway-mediated enhancement of human cytomegalovirus replication correlates with a recruitment of SUMO-1/3 proteins to viral replication compartments. J Gen Virol 94:1373–1384
Schmidt D, Muller S (2003) PIAS/SUMO: new partners in transcriptional regulation. Cell Mol Life Sci 60:2561–2574
Schreiner S, Wimmer P, Groitl P, Chen SY, Blanchette P, Branton PE, Dobner T (2011) Adenovirus type 5 early region 1B 55K oncoprotein-dependent degradation of cellular factor Daxx is required for efficient transformation of primary rodent cells. J Virol 85:8752–8765
Sharrocks AD (2006) PIAS proteins and transcriptional regulation – more than just SUMO E3 ligases? Genes Dev 20:754–758
Sinigalia E, Alvisi G, Segre CV, Mercorelli B, Muratore G, Winkler M, Hsiao HH, Urlaub H, Ripalti A, Chiocca S, Palu G, Loregian A (2012) The human cytomegalovirus DNA polymerase processivity factor UL44 is modified by SUMO in a DNA-dependent manner. PLoS One 7:e49630
Sloan E, Tatham MH, Groslambert M, Glass M, Orr A, Hay RT, Everett RD (2015) Analysis of the SUMO2 proteome during HSV-1 infection. PLoS Pathog 11:e1005059
Sohn SY, Hearing P (2012) Adenovirus regulates sumoylation of Mre11-Rad50-Nbs1 components through a paralog-specific mechanism. J Virol 86:9656–9665
Sohn SY, Bridges RG, Hearing P (2015) Proteomic analysis of ubiquitin-like posttranslational modifications induced by the adenovirus E4-ORF3 protein. J Virol 89:1744–1755
Spengler ML, Brattain MG (2006) Sumoylation inhibits cleavage of Sp1 N-terminal negative regulatory domain and inhibits Sp1-dependent transcription. J Biol Chem 281:5567–5574
Spengler ML, Kurapatwinski K, Black AR, Azizkhan-Clifford J (2002) SUMO-1 modification of human cytomegalovirus IE1/IE72. J Virol 76:2990–2996
Stallings CL, Silverstein SJ (2006) Posttranslational modification and cell type-specific degradation of varicella-zoster virus ORF29p. J Virol 80:10836–10846
Stanton R, Fox JD, Caswell R, Sherratt E, Wilkinson GWG (2002) Analysis of the human herpesvirus-6 immediate-early 1 protein. J Gen Virol 83:2811–2820
Steger G, Schnabel C, Schmidt HM (2002) The hinge region of the human papillomavirus type 8 E2 protein activates the human p21(WAF1/CIP1) promoter via interaction with Sp1. J Gen Virol 83:503–510
Su CI, Tseng CH, Yu CY, Lai MM (2016) SUMO modification stabilizes dengue virus nonstructural protein 5 to support virus replication. J Virol 90:4308–4319
Subramanian L, Benson MD, Iniguez-Lluhi JA (2003) A synergy control motif within the attenuator domain of CCAAT/enhancer-binding protein alpha inhibits transcriptional synergy through its PIASy-enhanced modification by SUMO-1 or SUMO-3. J Biol Chem 278:9134–9141
Sun DY, Xu P, He B (2011) Sumoylation of the P protein at K254 plays an important role in growth of parainfluenza virus 5. J Virol 85:10261–10268
Sureau C, Negro F (2016) The hepatitis delta virus: replication and pathogenesis. J Hepatol 64:S102–S116
Tatham MH, Jaffray E, Vaughan OA, Desterro JMP, Botting CH, Naismith JH, Hay RT (2001) Polymeric chains of SUMO-2 and SUMO-3 are conjugated to protein substrates by SAE1/SAE2 and Ubc9. J Biol Chem 276:35368–35374
Tavalai N, Adler M, Scherer M, Riedl Y, Stamminger T (2011) Evidence for a dual antiviral role of the major nuclear domain 10 component Sp100 during the immediate-early and late phases of the human cytomegalovirus replication cycle. J Virol 85:9447–9458
Tomoiu A, Gravel A, Tanguay RM, Flamand L (2006) Functional interaction between human herpesvirus 6 immediate-early 2 protein and ubiquitin-conjugating enzyme 9 in the absence of sumoylation. J Virol 80:10218–10228
Tseng CH, Cheng TS, Shu CY, Jeng KS, Lai MMC (2010) Modification of small hepatitis delta virus antigen by SUMO protein. J Virol 84:918–927
Turci M, Lodewick J, Righi P, Polania A, Romanelli MG, Bex F, Bertazzoni U (2009) HTLV-2B Tax oncoprotein is modified by ubiquitination and sumoylation and displays intracellular localization similar to its homologue HTLV-1 Tax. Virology 386:6–11
Turci M, Lodewick J, Di Gennaro G, Rinaldi AS, Marin O, Diani E, Sampaio C, Bex F, Bertazzoni U, Romanelli MG (2012) Ubiquitination and sumoylation of the HTLV-2 Tax-2B protein regulate its NF-kappaB activity: a comparative study with the HTLV-1 Tax-1 protein. Retrovirology 9:102
Varadaraj A, Mattoscio D, Chiocca S (2014) SUMO Ubc9 enzyme as a viral target. IUBMB Life 66:27–33
Voigt EA, Kennedy RB Poland GA, 2016 Defending against smallpox: a focus on vaccines. Expert Rev Vaccines 1–15
Wang L, Oliver SL, Sommer M, Rajamani J, Reichelt M, Arvin AM (2011) Disruption of PML nuclear bodies is mediated by ORF61 SUMO-interacting motifs and required for varicella-zoster virus pathogenesis in skin. PLoS Pathog 7:e1002157
Weger S, Hammer E, Heilbronn R (2004) SUMO-1 modification regulates the protein stability of the large regulatory protein Rep78 of adeno associated virus type 2 (AAV-2). Virology 330:284–294
Weitzman MD, Ornelles DA (2005) Inactivating intracellular antiviral responses during adenovirus infection. Oncogene 24:7686–7696
Wilson VG (2012) SUMOylation at the host-pathogen interface. Biomolecules 2:203–227
Wilson VG, Rosas-Acosta G (2003) Molecular targets for papillomavirus therapy. Curr Drug Targets Infect Disord 3:221–239
Wimmer P, Schreiner S (2015) Viral Mimicry to usurp ubiquitin and SUMO host pathways. Viruses 7:4854–4872
Wimmer P, Schreiner S, Everett RD, Sirma H, Groitl P, Dobner T (2010) SUMO modification of E1B-55K oncoprotein regulates isoform-specific binding to the tumour suppressor protein PML. Oncogene 29:5511–5522
Wimmer P, Schreiner S, Dobner T (2012) Human pathogens and the host cell SUMOylation system. J Virol 86:642–654
Wimmer P, Blanchette P, Schreiner S, Ching W, Groitl P, Berscheminski J, Branton PE, Will H, Dobner T (2013) Cross-talk between phosphorylation and SUMOylation regulates transforming activities of an adenoviral oncoprotein. Oncogene 32:1626–1637
Wimmer P, Berscheminski J, Blanchette P, Groitl P, Branton PE, Hay RT, Dobner T, Schreiner S (2016) PML isoforms IV and V contribute to adenovirus-mediated oncogenic transformation by functionally inhibiting the tumor-suppressor p53. Oncogene 35:69–82
Woods YL, Xirodimas DP, Prescott AR, Sparks A, Lane DP, Saville MK (2004) p14 Arf promotes small ubiquitin-like modifier conjugation of Werners helicase. J Biol Chem 279:50157–50166
Wu YC, Deyrieux AF, Wilson VG (2007) Papillomaviruses and the host SUMOylation system. Biochem Soc Trans 35:1433–1435
Wu YC, Roark AA, Bian XL, Wilson VG (2008) Modification of papillomavirus E2 proteins by the small ubiquitin-like modifier family members (SUMOs). Virology 378:329–338
Wu YC, Bian XL, Heaton PH, Deyrieux AF, Wilson VG (2009) Host cell sumoylation level influences papillomavirus E2 protein stability. Virology 387:176–183
Wu CY, Jeng KS, Lai MMC (2011) The SUMOylation of matrix protein M1 Modulates the assembly and morphogenesis of influenza A virus. J Virol 85:6618–6628
Xu YX, Ahn JH, Cheng MF, Ap Rhys CM, Chiou CJ, Zong JH, Matunis MJ, Hayward GS (2001) Proteasome-independent disruption of PML oncogenic domains (PODs), but not covalent modification by SUMO-1, is required for human cytomegalovirus immediate-early protein IE1 to inhibit PML-mediated transcriptional repression. J Virol 75:10683–10695
Xu K, Klenk C, Liu B, Keiner B, Cheng JK, Zheng BJ, Li L, Han QL, Wang C, Li TX, Chen Z, Shu YL, Liu JH, Klenk HD, Sun B (2011) Modification of nonstructural protein 1 of influenza A virus by SUMO1. J Virol 85:1086–1098
Yang YC, Yoshikai Y, Hsu SW, Saitoh H, Chang LK (2013) Role of RNF4 in the ubiquitination of Rta of Epstein-Barr virus. J Biol Chem 288:12866–12879
Yang WS, Hsu HW, Campbell M, Cheng CY, Chang PC (2015a) K-bZIP Mediated SUMO-2/3 Specific modification on the KSHV genome negatively regulates lytic gene expression and viral reactivation. PLoS Pathog 11:e1005051
Yang YC, Feng TH, Chen TY, Huang HH, Hung CC, Liu ST, Chang LK (2015b) RanBPM regulates Zta-mediated transcriptional activity in Epstein-Barr virus. J Gen Virol 96:2336–2348
You J, Croyle JL, Nishimura A, Ozato K, Howley PM (2004) Interaction of the bovine papillomavirus E2 protein with Brd4 tethers the viral DNA to host mitotic chromosomes. Cell 117:349–360
Yousef AF, Fonseca GJ, Pelka P, Ablack JNG, Walsh C, Dick FA, Bazett-Jones DP, Shaw GS, Mymryk JS (2010) Identification of a molecular recognition feature in the E1A oncoprotein that binds the SUMO conjugase UBC9 and likely interferes with polySUMOylation. Oncogene 29:4693–4704
Yueh A, Leung J, Bhattacharyya S, Perrone LA, Pu SY, Goff SP (2006) Interaction of Moloney murine leukemia virus capsid with Ubc9 and PIASy mediates SUMO-1 addition required early in infection. J Virol 80:342–352
Zamborlini A, Coiffic A, Beauclair G, Delelis O, Paris J, Koh Y, Magne F, Giron ML, Tobaly-Tapiero J, Deprez E, Emiliani S, Engelman A, de The H, Saib A (2011) Impairment of human immunodeficiency virus type-1 integrase SUMOylation correlates with an early replication defect. J Biol Chem 286:21013–21022
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2017 Springer International Publishing AG
About this chapter
Cite this chapter
Wilson, V.G. (2017). Viral Interplay with the Host Sumoylation System. In: Wilson, V. (eds) SUMO Regulation of Cellular Processes. Advances in Experimental Medicine and Biology, vol 963. Springer, Cham. https://doi.org/10.1007/978-3-319-50044-7_21
Download citation
DOI: https://doi.org/10.1007/978-3-319-50044-7_21
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-319-50043-0
Online ISBN: 978-3-319-50044-7
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)