
Abstract
The nucleolus is a dynamic sub-nuclear structure integral to the function of a eukaryotic cell. Some of its major roles involve ribosome subunit biogenesis, RNA processing, cell cycle control and responding to cellular stress, such as infection. Our understanding of the relationship between viruses and the nucleolus has moved from a phenomenological approach describing protein localisation to functional studies involving genetic analysis and proteomic approaches. These advances have provided fundamental insights as to how and why the nucleolus is targeted by many different viruses both to usurp normal functioning and to recruit nucleolar proteins to facilitate virus replication. This knowledge has been exploited for therapeutic strategies involving targeted inhibition of virus replication and live-attenuated recombinant vaccines.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- Japanese Encephalitis Virus
- Hepatitis Delta Virus
- Nucleolar Protein
- Japanese Encephalitis Virus
- Tomato Bushy Stunt Virus
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
The nucleolus is a dynamic sub-nuclear structure with roles in ribosome subunit biogenesis, mediation of cell stress responses and regulation of cell growth (Boulon et al. 2010). The proteome and structure of the nucleolus are constantly changing in response to metabolic conditions, and virus infection represents one of the major challenges to nucleolar function (Greco 2009; Hiscox 2002, 2003, 2007; Hiscox et al. 2010). Viruses are obligate intracellular parasites and rely on the host cell for genome replication, protein expression and assembly of new virus particles. During infection there is a constant war between viruses trying to subvert the host cell and host-mediated anti-viral activity and interaction with the nucleolus is likely to be a key stage in this.
Interaction with the nucleolus is a pan-virus phenomenon and evidence suggests that proteins from many different types of viruses, such as those with DNA, RNA or RNA/DNA (e.g. retroviruses) genomes, encode proteins that can localise to the nucleolus during infection (Table 14.1). These examples include viruses with DNA genomes including the poxviruses, which replicate in the cytoplasm, as well as the herpes and adenoviruses, which replicate in the nucleus. HIV-1, perhaps the classic example of a retrovirus, undergoes an initial replication event in the cytoplasm and then further activity in the nucleus. RNA viruses encompass genomes of single-stranded positive and negative polarity and also double-stranded RNA. Established dogma suggests that positive strand-RNA viral genome synthesis and transcription occur in the cytoplasm. Examples of negative strand RNA viruses can be found, which replicate in the cytoplasm (most of the Mononegavirales) and the nucleus (e.g. influenza viruses).
The reason why RNA viruses, and positive-strand RNA viruses in particular, interact with the nucleolus when the site of genome replication is in the cytoplasm is less intuitive. In this latter case, viral proteins that are normally required in the cytoplasm must transit through the nuclear pore complex both to and from the nucleus. This process is crucial for virus biology because if the viral proteins that are required for cytoplasmic functions such as RNA synthesis and encapsidation are sequestered in the nucleolus or nucleus, then progeny virus production will be affected as has been revealed by inhibitor and genetic studies (Lee et al. 2006; Tijms et al. 2002). Viruses may interact with the nucleolus to usurp host cell functions and recruit nucleolar proteins to facilitate virus replication. Investigating the interactions between viruses and the nucleolus may facilitate the design of novel anti-viral therapies both in terms of recombinant vaccines (Pei et al. 2008) and molecular intervention (Rossi et al. 2007), and also contribute to a more detailed understanding of the cell biology of the nucleolus.
For many years our understanding of the interaction of viruses and the nucleolus was phenomenological and focused on identifying viral proteins that localised to this structure, their mechanisms of trafficking and potential interaction with nucleolar proteins (e.g. see Table 14.1). However, recent research capitalising on advances in proteomics, viral genetics and cellular imaging techniques are beginning to increase our understanding of the mechanisms viruses use to subvert host cell nucleoli and facilitate virus biology (Hiscox et al. 2010).
New data are now emerging that support the view that many viruses interact with the nucleus and nucleolus, particularly to facilitate virus replication. One of the best-studied viruses in terms of viral interactions with the nucleolus is HIV-1 and is described in detail in Chap. 17. Although HIV has clearly defined cytoplasmic and nuclear replication strategies, the virus has a positive-sense RNA genome in the sense that the viral capsid contains two copies of positive-sense RNA, but these are reverse transcribed in the cytoplasm and then trafficked to the nucleus, where ultimately the new genome is transcribed and trafficked back to the cytoplasm. Part of the reasoning for the interaction of HIV-1 with the nucleolus is the trafficking of intronless mRNA from the nucleus into the cytoplasm (Michienzi et al. 2000). This is a property shared with herpes viruses and indicated that different viruses have evolved similar strategies involving subversion of nucleolar function for the benefit of virus biology (Boyne and Whitehouse 2006). In the case of HIV-1, this knowledge has also led to the design and implementation of effective genetic therapies against the virus (Unwalla et al. 2008).
2 DNA Virus Interactions with the Nucleolus
A large number of viruses with DNA genomes have been shown to interact with nucleolus, and this perhaps is not surprising as most DNA viruses replicate in the nucleus. A genome-wide screen of three distinct herpesviruses, herpes simplex virus 1 (HSV-1), cytomegalovirus (CMV) and Epstein–Barr virus (EBV), has shown that at least 12 herpesvirus-encoded proteins specifically localise to the nucleolus (Salsman et al. 2008), which are implicated in many aspects of the herpesvirus life cycle. Therefore, a number of proteomic studies are currently being undertaken to study changes, in a global context, within the nucleolar proteome during virus infections, and are discussed later (Lam et al. 2010). Several different herpes virus proteins have been shown to cause the redistribution of nucleolar proteins and hence disruption of the nucleolus. These include herpes simplex virus 1, the major tegument structural protein VP22 (Lopez et al. 2008), and the US11 (Xing et al. 2010) and UL24 proteins (Bertrand and Pearson 2008; Lymberopoulos and Pearson 2007). Such disruption in many cases may have a direct effect on nucleolar function.
A significant area of virus biology that has been investigated is the role of viral proteins that traffic through the nucleolus. For example, a number of HIV proteins that traffic through the nucleolus have been implicated in virus mRNA processing (Dundr et al. 1995). Similar observations have also been made in herpesviruses (Boyne and Whitehouse 2006, 2009; Leenadevi and Dalziel 2009). Initial studies utilising the prototype γ-2 herpesvirus, herpes virus saimiri (HVS), demonstrated that the HVS nucleolar trafficking ORF57 protein induces nucleolar redistribution of the host cell human TREX proteins, which are involved in mRNA nuclear export (Boyne and Whitehouse 2006). Intriguingly, ablating ORF57 nucleolar trafficking led to a failure of ORF57-mediated viral mRNA nuclear export (Boyne and Whitehouse 2006). The precise role of this nucleolar sequestration is yet to be determined, but possible effects on viral mRNA/protein processing and viral ribonucleoprotein particle assembly are currently being investigated.
This property may also be conserved in other ORF57 homologues as recent analysis has shown that the ORF57 protein from Kaposi’s sarcoma associated herpesvirus (KSHV) also dynamically traffics through the nucleolus (Boyne et al. 2008b). Moreover, on the rapid disorganisation of the nucleolus a reduction is observed in virus mRNA nuclear export (Boyne and Whitehouse 2009). The formation of an ORF57-mediated export competent ribonucleoprotein particle within the nucleolus may also have implications for the translation of viral mRNAs. For example, it has recently been demonstrated that the cellular nucleo-cytoplasmic shuttle protein, PYM, which is involved in translation enhancement, is redistributed to the nucleolus in the presence of the KSHV ORF57 protein (Boyne et al. 2010). This interaction effectively enhances the translation of the predominantly intronless transcripts made by KSHV, and draws parallels with potential translation enhancement of positive strand RNA virus genomes through their interaction with the nucleolus (discussed later).
A second area of virus replication where nucleolar proteins are sequestered involves the replication of the virus DNA genome. For example, we (Matthews) and others have observed that nucleolar antigens upstream binding factor (UBF) and nucleophosmin (B23.1) are both sequestered into adenovirus DNA replication centres where they promote viral DNA replication (Hindley et al. 2007; Lawrence et al. 2006; Okuwaki et al. 2001). Similarly, in HSV-1 infected cells, a number of nucleolar proteins including nucleolin and UBF are recruited into viral DNA replication centres (Lymberopoulos and Pearson 2010). These are specific sites where replication and encapsidation of the HSV-1 genome occurs. Evidence suggests that sequestration of UBF is essential for viral DNA replication as overexpression of tagged version of UBF acts in a dominant-negative manner inhibiting virus DNA replication (Stow et al. 2009). Moreover, depletion of nucleolin results in reduced virus gene expression and infectious virion production (Calle et al. 2008; Sagou et al. 2010).
In addition to enhancing virus replication, nucleolar proteins are redistributed to alter cellular pathways during infection. For example, the nucleolar targeted HSV-1 US11 protein has been shown to interact with homeodomain-interacting protein kinase 2 (HIPK2), which plays a role in p53-mediated cellular apoptosis and hypoxic response (Calzado et al. 2009) and also participates in the regulation of the cell cycle (Calzado et al. 2007). This interaction alters the sub-cellular localisation of HIPK2 and protects against HIPK2-mediated cell cycle arrest (Giraud et al. 2004). In contrast, the cellular protein, protein interacting with the carboxyl terminus-1 (PICT-1), can sequester the virally encoded apoptosis suppressor protein, KS-Bcl-2 protein, from the mitochondria into the nucleolus to down-regulate its anti-apoptotic activity (Kalt et al. 2010). This is a potential interesting interplay between two sub-cellular structures involved in the viral stress response (Olson 2009), and maybe more common and widespread. For example, bacterial infection has been shown to disrupt the nucleolus through regulating mitochondrial dysfunction (Dean et al. 2010).
3 Interactions of RNA Viruses with the Nucleolus
Although many RNA virus proteins have been shown to localise to the nucleolus, most attention has focused on viral capsid proteins. These are proteins that associate with the viral genome for encapsidation and assembly of new virus particles. These proteins may also modulate replication (and transcription, where appropriate) of the viral genome. Increasingly, capsid proteins have also been shown to have a number of roles in modulating host cell signalling pathways and functions. These capsid proteins are referred to as capsid, nucleoproteins or nucleocapsid proteins, depending on the virus. In many cases, they are phosphorylated (Chen et al. 2005), which can modulate activity (Spencer et al. 2008).
Many examples of these proteins have been shown to localise to the nucleolus both when over-expressed and also in infected cells. These include proteins from positive-strand animal and plant RNA viruses, including the coronavirus nucleocapsid protein (Chen et al. 2002; Hiscox et al. 2001; Wurm et al. 2001), the arterivirus nucleocapsid protein (Rowland et al. 1999), the alphavirus capsid protein (Jakob 1994) and non-structural protein nsP2 (Rikkonen et al. 1992, 1994) and the umbravirus ORF3 protein (Ryabov et al. 2004). Capsid proteins from negative-strand RNA viruses also localise to the nucleolus. These have strain dependent localisation of a number of different influenza virus proteins (Emmott et al. 2010c; Han et al. 2010; Melen et al. 2007; Volmer et al. 2010).
For many years this has followed a phenomenological pattern and viral capsid and RNA-binding proteins might simply localise to the nucleolus because they diffuse through the nuclear pore complex and associate with compartments in the nucleus that have high RNA contents – the nucleolus in particular because it is transcriptionally active. In this case, sub-cellular localisation to the nucleolus would have no physiological consequence for the virus or the cell. However, RNA virus replication is error prone and selection pressure might apply to such a fortuitous localisation (given the ∼4,500+ nucleolar proteins and their diverse roles (Ahmad et al. 2009)), with the concomitant effect that the virus could select for changes that ultimately disrupt nucleolar function and/or recruit nucleolar proteins to aid virus replication.
There is a potential correlation between the nucleolar localisation of a viral protein and the loss of an essential nucleolar function. The molecular mechanisms responsible for this effect are unknown, but the displacement and re-localisation of nucleolar proteins by viral proteins could increase or decrease the nucleolar, nuclear and/or cytoplasmic pool of these proteins. Certainly, the accumulation of viral proteins in the nucleolus could potentially cause volume exclusion or crowding effects, which have been proposed to play a fundamental role in the formation of nuclear compartments including the nucleolus, and can be addressed by proteomic strategies. Therefore, disruption of nucleolar architecture and function might be common in virus-infected cells if viral proteins target the nucleolus or a stage of the virus lifecycle disrupts nucleolar proteins. For example, poliovirus infection results in the selective redistribution of nucleolin from the nucleolus to the cytoplasm (Waggoner and Sarnow 1998) and inactivation of UBF, which shuts off RNA polymerase I transcription in the host cell. The infection of cells with IBV has been shown to disrupt nucleolar architecture (Dove et al. 2006b) and cause arrest of the cell cycle in the G2/M phase and failure of cytokinesis (Dove et al. 2006a). The IBV and arterivirus nucleocapsid proteins associate with nucleolin and fibrillarin, respectively. Similarly, the HIV-1 Rev protein has been shown to localise to the DFC and GC and over-expression of Rev protein alters the nucleolar architecture and is associated with the accumulation of nucleophosmin (Dundr et al. 1995).
4 Trafficking of Virus Proteins to the Nucleolus
Many different virus proteins localise to the nucleolus (Table 14.1). However, predicting viral (and cellular) nucleolar targeting signals has historically been problematic and only recently has bioinformatic software been developed to fascilitate this (Scott et al. 2011). Nucleolar trafficking might be mediated by virtue of the fact that viral proteins that are trafficked to the nucleolus contain motifs that resemble host nucleolar targeting signals, that is, a form of molecular mimicry is used (Rowland and Yoo 2003). The discovery of specific nucleolar trafficking signals in viral proteins has indicated a functional mechanism behind this observed localisation (Lee et al. 2003; Reed et al. 2006; Rowland et al. 2003). Analysis of the different nucleolar trafficking signals identified in viral proteins using dynamic live-cell imaging has certainly demonstrated that different proteins can confer differential trafficking rates and localisation patterns (Emmott et al. 2008). This is very similar to cellular nucleolar proteins (Lechertier et al. 2007).
In some virus proteins, both NLSs and nucleolar targeting signals act in concert to direct a protein to the nucleolus. The arterivirus porcine reproductive and respiratory syndrome virus (PRRSV) nucleocapsid protein localises to the nucleolus and has been shown to contain two potential NLSs, a pat4 and a downstream pat7 motif (Rowland et al. 1999, 2003). Analysis revealed that a 31 amino acid sequence incorporating the pat7 motif could direct the nucleocapsid protein to both the nucleus and nucleolus. The protein also contains a predicted NES, presumably to allow the protein to traffic back into the cytoplasm to contribute to viral function in this compartment. This is common with other similar related proteins. For example, in the avian coronavirus nucleocapsid protein an eight amino acid sequence is necessary and sufficient to target the protein to the nucleolus (Reed et al. 2006) and contains an NES (Reed et al. 2007). Intriguingly, genetic analysis (Lee et al. 2006), dynamic live-cell imaging (You et al. 2008) and use of trafficking inhibitors (Tijms et al. 2002) paint a picture of the requirement of these positive sense RNA virus capsid proteins localising to the nucleolus as soon as they are translated, prior to their involvement in virus replication or assembly. This may be related to subversion of host cell function, protein modification (e.g. phosphorylation) or recruitment of nucleolar proteins.
Viral proteins might also traffic to the nucleolus through association with cellular nucleolar proteins (Yoo et al. 2003). For example, the hepatitis delta antigen has been shown to contain a nucleolar targeting signal that also corresponded to a site that promoted binding to nucleolin (Lee et al. 1998). Mutating this region prevented nucleolin binding to the delta antigen and nucleolar trafficking. By implication, this relates nucleolin binding to nucleolar trafficking (Lee et al. 1998). Certainly, interaction with nucleophosmin and hepatitis delta antigens can modulate viral replication (Huang et al. 2001) and more recently combined proteomic-RNAi screens have revealed many other nucleolar proteins that can be associated with this viral protein (Cao et al. 2009). Trafficking and accumulation of viral proteins to and from the nucleolus, similar to cellular proteins, may also be cell cycle related. For example, the coronavirus nucleocapsid protein localises preferentially to the nucleolus in the G2 phase of the cell cycle (Cawood et al. 2007), as does the human cytomegalovirus protein UL83 in the G1 phase (Arcangeletti et al. 2011). Again these trafficking profiles may be related to the interaction with cellular nucleolar proteins (Emmott and Hiscox 2009).
5 Functional Relevance of Nucleolar Interactions to the Viral Life Cycle
Many different examples now exist to show that the disruption of nuclear or nucleolar trafficking of viral proteins affects viral pathogenesis, and argues against nucleolar localisation as a purely phenomenological observation. For example, the Semliki Forest virus non-structural protein nsP2 can localise to the nucleolus (Peranen et al. 1990; Rikkonen et al. 1992, 1994) and disruption of this localisation through a single amino acid change results in a reduction in neurovirulence (Fazakerley et al. 2002). Such in vitro data has also been backed up by persuasive in vivo data. Mutation of the arterivirus nucleocapsid protein pat7 NLS motif in the context of a full-length clone revealed that this sequence could have a key role in virus pathogenesis in vivo, as animals infected with mutant viruses had shorter viraemia than wild-type viruses (Lee et al. 2006; Pei et al. 2008). Interestingly, reversions occurred in the mutated nucleocapsid gene sequence and although the amino acid sequence of the pat7 motif was altered, its function was not; this new signal was defined as a pat8 motif (Lee et al. 2006). The clear implications of this groundbreaking work is that disruption of nucleolar trafficking of a viral protein proves functional relevance and illustrates the potential of exploiting this knowledge for the generation of growth attenuated recombinant vaccines (Pei et al. 2008; Reed et al. 2006, 2007).
Similarly, point mutations in the Japanese encephalitis virus (JEV) core protein that abolished nuclear and nucleolar localisation resulted in recombinant viruses with impaired replication in mammalian cells, compared to wild type virus (Mori et al. 2005; Tsuda et al. 2006). Curiously, replication of recombinant viruses was not impaired in insect cells, illustrating this could potentially be related to differences in nucleolar architecture and proteomes between these cell types (Thiry and Lafontaine 2005). The JEV core protein has been shown to interact with nucleophosmin and is translocated from the nucleolus to the cytoplasm.
Flaviviruses in general (JEV, Dengue virus and West Nile virus) appear to have a part-nuclear stage to the synthesis of viral RNA and several components of the viral replicase together with newly synthesised RNA have been found in the nucleus of infected cells (Uchil et al. 2006). One intriguing question that has yet to be elucidated is how such viral RNA traffics from the nucleus to the cytoplasm. Most cellular mRNAs are spliced and it is part of the splicing process that signals nuclear export. Certain DNA viruses, such as herpesvirus saimiri, produce intron-less mRNA and these viruses have evolved specific viral proteins (such as herpesvirus saimiri ORF57 (Boyne et al. 2008a)), which interact with the cellular mRNA export machinery (e.g. the mRNA processing and export factor ALY) to traffic viral mRNA from the nucleus to the cytoplasm (Boyne et al. 2008b, 2010; Boyne and Whitehouse 2006) and a similar process might be required by RNA viruses. For example, tomato bushy stunt virus (TBSV) redistributes ALY from the nucleus to the cytoplasm, and this might be a way the virus mediates host cell protein synthesis (Uhrig et al. 2004). In plants RNA silencing, a host defence mechanism targets virus RNAs for degradation in a sequence-specific manner and viruses use several mechanisms to counteract this system (Canto et al. 2006). TBSV encodes a protein, P19, which interferes with this pathway. However, ALY might transport P19 from the cytoplasm to the nucleus or nucleolus and disrupt its silencing suppression activity. Nucleolin has also been shown to be involved in the trafficking of herpes simplex virus type 1 nucleocapsids from the nucleus to the cytoplasm (Sagou et al. 2010), drawing parallels with the involvement of nucleolar proteins in the movement of plant viruses (Kim et al. 2007a, b). Different plant virus proteins involved in long-distance phloem-associated movement of virus particles or with roles in binding to the RNA virus genomes localise to the nucleolus and other sub-nuclear structures (Kim et al. 2007b; Ryabov et al. 2004). This may be mediated by association with nuclear proteins, as is the case with fibrillarin and the ORF3 protein of plant umbraviruses (Kim et al. 2007a).
Hijacking the nucleolus is not exclusive to plant viruses and may also occur with mammalian viruses. Similar to the plant rhabdovirus maize fine streak virus (MFSV), whose nucleocapsid and phosphoproteins localise to the nucleolus (Tsai et al. 2005), the animal negative-stranded RNA virus Borna disease virus has been reported to use the nucleolus as a site for genome replication, and its RNA-binding protein has the appropriate trafficking signals for import to and export from the cytoplasm to the nucleus (Pyper et al. 1998). The hepatitis delta virus genome also has differential synthesis in the nucleus with RNA being transcribed in the nucleolus (Huang et al. 2001); this is similar to the potato spindle tuber viroid where RNAs of opposite polarity are sequestered in different nuclear compartments, with the positive-sense RNA being transported to the nucleolus. Again localisation to different sub-nuclear strcutures may have different roles in the virus life cycle (Li et al. 2006). An intriguing recent discovery has been made showing that adeno-associated virus (AAV) encodes an additional protein called assembly-activating protein (AAP) that localises to the nucleolus and promotes assembly of the viral capsid (Sonntag et al. 2010).
As a result of their limited genomes and coding capacities, recruitment of cellular proteins with defined functions in RNA metabolism would be a logical step to facilitate RNA virus infection. As nucleolar proteins have many crucial functions in cellular RNA biosynthesis, processing and translation, it comes as no surprise that nucleolar proteins are incorporated into the replication and/or translation complexes formed by RNA viruses. Given that some nucleolar proteins have many different functions, the same nucleolar protein might be used by a virus for different aspects of the replication pathway. Studies suggest that the human rhinovirus 3 C protease (3Cpro) pre-cursors, 3CD’ and/or 3CD, localise in the nucleoli of infected cells early in infection and inhibit cellular RNA transcription via proteolytic mechanisms (Amineva et al. 2004). This general property is not restricted to human rhinovirus and in terms of the inhibition of cellular translation has also been described for encephalomyocarditis virus (Aminev et al. 2003a, b), again suggesting roles in translational regulation.
6 Applying Quantitative Proteomics to Study Viral Interactions with the Nucleolar Proteome
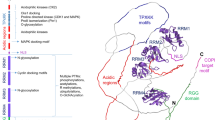
Given the many roles of the nucleolus in the life cycle of the cell, including as stress sensor (Boulon et al. 2010; Mayer and Grummt 2005), it would seem reasonable that comprehensive unbiased analysis of the nucleolar proteome would yield interesting data, particularly, with providing clues as to what cellular nucleolar functions may be altered by virus infection and what mechanisms the nucleolus may use to respond to this. How the nucleolar proteome changes in response to virus-infection has been investigated using stable isotope labelling with amino acids in cell culture (SILAC) coupled to LC-MS/MS and bioinformatics (Fig. 14.1). These studies, led by our laboratories, have analysed purified nucleoli and the nucleus, and have directly stemmed from the pioneering work of the Lamond laboratory in analysing purified nucleoli using quantitative proteomics (Andersen et al. 2005). Viruses investigated so far have included human adenovirus (Lam et al. 2010), avian coronavirus (Emmott et al. 2010a, b), different strains of influenza virus (Emmott et al. 2010c) and human respiratory syncytial virus (Munday et al. 2010). Overall, our data indicates that only a small proportion of nucleolar proteins change in abundance in virus-infected cells, and these tend to be virus-specific. For example, in adenovirus infected cells just 7% of proteins identified show a twofold or greater change compared to almost a third of nucleolar antigens showing a greater than twofold change when cells are treated with ActD which inhibits rRNA synthesis (Lam et al. 2010). What is notable is that direct comparison between the adenovirus data set and the ActD dataset shows no clear correlation (Hiscox et al. 2010; Lam et al. 2010), further supporting the case that adenovirus induces effects on the nucleolus distinct from that of a generalised, non-specific shut down of nucleolar function. This fits well with a previous observation that adenovirus infection does not affect rRNA synthesis even 36 h post-infection (Lawrence et al. 2006). These results were initially surprising given the number of different viral proteins that can localise to this structure and how they interact with nucleolar proteins. This suggests that the nucleolar proteome and architecture is resilient during early stages of infection but may become disrupted as more and more damage accumulates inside cells because of virus activity, as clearly evidenced in live-cell imaging experiments (Bertrand and Pearson 2008; Dove et al. 2006b; Lymberopoulos et al. 2010).

Diagram of a “classic” SILAC experiment. This technology allows high-throughput quantitative proteomics and has been readily applied to the nucleolus, especially when coupled with dynamic live-cell imaging (Andersen et al. 2005). The ability to simultaneously compare up to three different conditions through selection of the appropriate isotope label has enabled the recent studies of how the nucleolar proteome changes in virus-infected cells (Emmott et al. 2010a; Emmott et al. 2010b; Emmott et al. 2010c; Hiscox et al. 2010; Lam et al. 2010)
7 Future Research Directions
Coupling quantitative proteomic analysis of the nucleolus and deep sequencing throughout infection in time-course experiments of lytic, latent, acute and persistent viruses would reveal valuable insights into the response of the nucleolus to virus infection. Likewise, being able to move from studying cell culture-adapted laboratory strains into clinical isolates replicating in primary cells would yield more biologically relevant information, particularly with regard to the severity of disease and nucleolar changes. These technologies could also be applied to large-scale analysis of viral proteins that traffic to the nucleolus and the cellular nucleolar proteins that they associate with (e.g. using SILAC and EGFP-traps (Trinkle-Mulcahy et al. 2008)), thus generating and integrating interactome networks with the nucleolar proteome during infection.
References
Ahmad Y, Boisvert FM, Gregor P, Cobley A, Lamond AI (2009) NOPdb: nucleolar proteome database–2008 update. Nucleic Acids Res 37:D181–D184
Aminev AG, Amineva SP, Palmenberg AC (2003a) Encephalomyocarditis viral protein 2A localizes to nucleoli and inhibits cap-dependent mRNA translation. Virus Research 95:45–57
Aminev AG, Amineva SP, Palmenberg AC (2003b) Encephalomyocarditis virus (EMCV) proteins 2A and 3BCD localize to nuclei and inhibit cellular mRNA transcription but not rRNA transcription. Vir Res 95:59–73
Amineva SP, Aminev AG, Palmenberg AC, Gern JE (2004) Rhinovirus 3 C protease precursors 3CD and 3CD’ localize to the nuclei of infected cells. J Gen Virol 85:2969–2979
Andersen JS, Lam YW, Leung AK, Ong SE, Lyon CE, Lamond AI, Mann M (2005) Nucleolar proteome dynamics. Nature 433:77–83
Arcangeletti MC, De Conto F, Ferraglia F, Pinardi F, Gatti R, Orlandini G, Calderaro A, Motta F, Medici MC, Martinelli M, Valcavi P, Razin SV, Chezzi C, Dettori G (2003) Human cytomegalovirus proteins PP65 and IEP72 are targeted to distinct compartments in nuclei and nuclear matrices of infected human embryo fibroblasts. J Cell Biochem 90:1056–1067
Arcangeletti MC, Rodighiero I, Mirandola P, De Conto F, Covan S, Germini D, Razin S, Dettori G, Chezzi C (2011) Cell-cycle-dependent localization of human cytomegalovirus UL83 phosphoprotein in the nucleolus and modulation of viral gene expression in human embryo fibroblasts in vitro. J Cell Biochem 112:307–317
Armbruester V, Sauter M, Roemer K, Best B, Hahn S, Nty A, Schmid A, Philipp S, Mueller A, Mueller-Lantzsch N (2004) Np9 protein of human endogenous retrovirus K interacts with ligand of numb protein X. J Virol 78:10310–10319
Bartoe JT, Albrecht B, Collins ND, Robek MD, Ratner L, Green PL, Lairmore MD (2000) Functional role of pX open reading frame II of human T-lymphotropic virus type 1 in maintenance of viral loads in vivo. J Virol 74:1094–1100
Baunoch DA, Das P, Browning ME, Hari V (1991) A temporal study of the expression of the capsid, cytoplasmic inclusion and nuclear inclusion proteins of tobacco etch potyvirus in infected plants. J Gen Virol 72:487–492
Beauchemin C, Boutet N, Laliberte JF (2007) Visualization of the interaction between the precursors of VPg, the viral protein linked to the genome of turnip mosaic virus, and the translation eukaryotic initiation factor iso 4E in Planta. J Virol 81:775–782
Bertrand L, Pearson A (2008) The conserved N-terminal domain of herpes simplex virus 1 UL24 protein is sufficient to induce the spatial redistribution of nucleolin. J Gen Virol 89:1142–1151
Blanie S, Mortier J, Delverdier M, Bertagnoli S, Camus-Bouclainville C (2009) M148R and M149R are two virulence factors for myxoma virus pathogenesis in the European rabbit. Vet Res 40:11
Boulon S, Westman BJ, Hutten S, Boisvert FM, Lamond AI (2010) The nucleolus under stress. Mol Cell 40:216–227
Boyne JR, Whitehouse A (2006) Nucleolar trafficking is essential for nuclear export of intronless herpesvirus mRNA. Proc Natl Acad Sci USA 103:15190–15195
Boyne JR, Whitehouse A (2009) Nucleolar disruption impairs Kaposi’s sarcoma-associated herpesvirus ORF57-mediated nuclear export of intronless viral mRNAs. FEBS Lett 583:3549–3556
Boyne JR, Colgan KJ, Whitehouse A (2008a) Herpesvirus saimiri ORF57: a post-transcriptional regulatory protein. Front Biosci 13:2928–2938
Boyne JR, Colgan KJ, Whitehouse A (2008b) Recruitment of the complete hTREX complex is required for Kaposi’s sarcoma-associated herpesvirus intronless mRNA nuclear export and virus replication. PLoS Pathog 4:e1000194
Boyne JR, Jackson BR, Taylor A, Macnab SA, Whitehouse A (2010) Kaposi’s sarcoma-associated herpesvirus ORF57 protein interacts with PYM to enhance translation of viral intronless mRNAs. EMBO J 29:1851–1864
Calle A, Ugrinova I, Epstein AL, Bouvet P, Diaz JJ, Greco A (2008) Nucleolin is required for an efficient herpes simplex virus type 1 infection. J Virol 82:4762–4773
Calzado MA, Renner F, Roscic A, Schmitz ML (2007) HIPK2: a versatile switchboard regulating the transcription machinery and cell death. Cell Cycle 6:139–143
Calzado MA, De La Vega L, Munoz E, Schmitz ML (2009) From top to bottom: the two faces of HIPK2 for regulation of the hypoxic response. Cell Cycle 8:1659–1664
Canto T, Uhrig JF, Swanson M, Wright KM, MacFarlane SA (2006) Translocation of Tomato bushy stunt virus P19 protein into the nucleus by ALY proteins compromises its silencing suppressor activity. J Virol 80:9064–9072
Cao D, Haussecker D, Huang Y, Kay MA (2009) Combined proteomic-RNAi screen for host factors involved in human hepatitis delta virus replication. RNA 15:1971–1979
Caporale M, Arnaud F, Mura M, Golder M, Murgia C, Palmarini M (2009) The signal peptide of a simple retrovirus envelope functions as a posttranscriptional regulator of viral gene expression. J Virol 83:4591–4604
Cawood R, Harrison SM, Dove BK, Reed ML, Hiscox JA (2007) Cell cycle dependent nucleolar localization of the coronavirus nucleocapsid protein. Cell Cycle 6:863–867
Chen H, Wurm T, Britton P, Brooks G, Hiscox JA (2002) Interaction of the coronavirus nucleoprotein with nucleolar antigens and the host cell. J Virol 76:5233–5250
Chen H, Gill A, Dove BK, Emmett SR, Kemp FC, Ritchie MA, Dee M, Hiscox JA (2005) Mass spectroscopic characterisation of the coronavirus infectious bronchitis virus nucleoprotein and elucidation of the role of phosphorylation in RNA binding using surface plasmon resonance. J Virol 79:1164–1179
Cheng G, Brett ME, He B (2002) Signals that dictate nuclear, nucleolar, and cytoplasmic shuttling of the gamma(1)34.5 protein of herpes simplex virus type 1. J Virol 76:9434–9445
Cochrane AW, Perkins A, Rosen CA (1990) Identification of sequences important in the nucleolar localization of human immunodeficiency virus Rev: relevance of nucleolar localization to function. J Virol 64:881–885
Davey J, Colman A, Dimmock NJ (1985) Location of influenza virus M, NP and NS1 proteins in microinjected cells. J Gen Virol 66:2319–2334
Dean P, Scott JA, Knox AA, Quitard S, Watkins NJ, Kenny B (2010) The enteropathogenic E. coli effector EspF targets and disrupts the nucleolus by a process regulated by mitochondrial dysfunction. PLoS Pathog 6:e1000961
Dillon PJ, Nelbock P, Perkins A, Rosen CA (1991) Structural and functional analysis of the human immunodeficiency virus type 2 Rev protein. J Virol 65:445–449
Dove B, Brooks G, Bicknell K, Wurm T, Hiscox JA (2006a) Cell cycle perturbations induced by infection with the coronavirus infectious bronchitis virus and their effect on virus replication. J Virol 80:4147–4156
Dove BK, You JH, Reed ML, Emmett SR, Brooks G, Hiscox JA (2006b) Changes in nucleolar morphology and proteins during infection with the coronavirus infectious bronchitis virus. Cell Microbiol 8:1147–1157
Dundr M, Leno GH, Hammarskjold ML, Rekosh D, Helga-Maria C, Olson MO (1995) The roles of nucleolar structure and function in the subcellular location of the HIV-1 Rev protein. J Cell Sci 108:2811–2823
Emmott E, Hiscox JA (2009) Nucleolar targetting: the hub of the matter. EMBO Reports 10:231–238
Emmott E, Dove BK, Howell G, Chappell LA, Reed ML, Boyne JR, You JH, Brooks G, Whitehouse A, Hiscox JA (2008) Viral nucleolar localisation signals determine dynamic trafficking within the nucleolus. Virology 380:191–202
Emmott E, Rodgers MA, Macdonald A, McCrory S, Ajuh P, Hiscox JA (2010a) Quantitative proteomics using stable isotope labeling with amino acids in cell culture reveals changes in the cytoplasmic, nuclear, and nucleolar proteomes in vero cells infected with the coronavirus infectious bronchitis virus. Mol Cell Proteomics 9:1920–1936
Emmott E, Smith C, Emmett SR, Dove BK, Hiscox JA (2010b) Elucidation of the avian nucleolar proteome by quantitative proteomics using stable isotope labeling with amino acids in cell culture (SILAC) and alteration in the coronavirus infectious bronchitis virus infected cells. Proteomics 10:3558–3562
Emmott E, Wise H, Loucaides EM, Matthews DA, Digard P, Hiscox JA (2010c) Quantitative proteomics using SILAC coupled to LC-MS/MS reveals changes in the nucleolar proteome in influenza A virus infected cells. J Proteome Res 9:5335–5345
Falcon V, Acosta-Rivero N, Chinea G, de la Rosa MC, Menendez I, Duenas-Carrera S, Gra B, Rodriguez A, Tsutsumi V, Shibayama M, Luna-Munoz J, Miranda-Sanchez MM, Morales-Grillo J, Kouri J (2003) Nuclear localization of nucleocapsid-like particles and HCV core protein in hepatocytes of a chronically HCV-infected patient. Biochem Biophys Res Commun 310:54–58
Fazakerley JK, Boyd A, Mikkola ML, Kaariainen L (2002) A single amino acid change in the nuclear localization sequence of the nsP2 protein affects the neurovirulence of semliki forest virus. J Virol 76:392–396
Finsterbusch T, Steinfeldt T, Caliskan R, Mankertz A (2005) Analysis of the subcellular localization of the proteins Rep, Rep’ and Cap of porcine circovirus type 1. Virology 343:36–46
Fong SE, Greenwood JD, Williamson JC, Derse D, Pallansch LA, Copeland T, Rasmussen L, Mentzer A, Nagashima K, Tobin G, Gonda MA (1997) Bovine immunodeficiency virus tat gene: cloning of two distinct cDNAs and identification, characterization, and immunolocalization of the tat gene products. Virology 233:339–357
Ghorbel S, Sinha-Datta U, Dundr M, Brown M, Franchini G, Nicot C (2006) Human T-cell leukemia virus type I p30 nuclear/nucleolar retention is mediated through interactions with RNA and a constituent of the 60 S ribosomal subunit. J Biol Chem 281:37150–37158
Giraud S, Diaz-Latoud C, Hacot S, Textoris J, Bourette RP, Diaz JJ (2004) US11 of herpes simplex virus type 1 interacts with HIPK2 and antagonizes HIPK2-induced cell growth arrest. J Virol 78:2984–2993
Goatley LC, Marron MB, Jacobs SC, Hammond JM, Miskin JE, Abrams CC, Smith GL, Dixon LK (1999) Nuclear and nucleolar localization of an African swine fever virus protein, I14L, that is similar to the herpes simplex virus-encoded virulence factor ICP34.5. J Gen Virol 80:525–535
Goic B, Bustamante J, Miquel A, Alvarez M, Vera MI, Valenzuela PD, Burzio LO (2008) The nucleoprotein and the viral RNA of infectious salmon anemia virus (ISAV) are localized in the nucleolus of infected cells. Virology 379:55–63
Gomez Corredor A, Archambault D (2009) The bovine immunodeficiency virus rev protein: identification of a novel lentiviral bipartite nuclear localization signal harboring an atypical spacer sequence. J Virol 83:12842–12853
Gonzalez I, Martinez L, Rakitina DV, Lewsey MG, Atencio FA, Llave C, Kalinina NO, Carr JP, Palukaitis P, Canto T (2010) Cucumber mosaic virus 2b protein subcellular targets and interactions: their significance to RNA silencing suppressor activity. Mol Plant Microbe Interact 23:294–303
Greco A (2009) Involvement of the nucleolus in replication of human viruses. Rev Med Virol 19:201–214
Guo YX, Dallmann K, Kwang J (2003) Identification of nucleolus localization signal of betanodavirus GGNNV protein alpha. Virology 306:225–235
Guo H, Ding Q, Lin F, Pan W, Lin J, Zheng AC (2009) Characterization of the nuclear and nucleolar localization signals of bovine herpesvirus-1 infected cell protein 27. Virus Res 145:312–320
Han H, Cui ZQ, Wang W, Zhang ZP, Wei HP, Zhou YF, Zhang XE (2010) New regulatory mechanisms for the intracellular localization and trafficking of influenza A virus NS1 protein revealed by comparative analysis of A/PR/8/34 and A/Sydney/5/97. J Gen Virol 91:2907–2917
Harders J, Lukacs N, Robert-Nicoud M, Jovin TM, Riesner D (1989) Imaging of viroids in nuclei from tomato leaf tissue by in situ hybridization and confocal laser scanning microscopy. EMBO J 8:3941–3949
Harms JS, Ren X, Oliveira SC, Splitter GA (2000) Distinctions between bovine herpesvirus 1 and herpes simplex virus type 1 VP22 tegument protein subcellular associations. J Virol 74:3301–3312
Haupt S, Stroganova T, Ryabov E, Kim SH, Fraser G, Duncan G, Mayo MA, Barker H, Taliansky M (2005) Nucleolar localization of potato leafroll virus capsid proteins. J Gen Virol 86:2891–2896
Hindley CE, Davidson AD, Matthews DA (2007) Relationship between adenovirus DNA replication proteins and nucleolar proteins B23.1 and B23.2. J Gen Virol 88:3244–3248
Hirano M, Kaneko S, Yamashita T, Luo H, Qin W, Shirota Y, Nomura T, Kobayashi K, Murakami S (2003) Direct interaction between nucleolin and hepatitis C virus NS5B. J Biol Chem 278:5109–5115
Hiscox JA (2002) Brief review: the nucleolus - a gateway to viral infection? Arch Virol 147:1077–1089
Hiscox JA (2003) The interaction of animal cytoplasmic RNA viruses with the nucleus to facilitate replication. Vir Res 95:13–22
Hiscox JA (2007) RNA viruses: hijacking the dynamic nucleolus. Nat Rev Microbiol 5:119–127
Hiscox JA, Wurm T, Wilson L, Cavanagh D, Britton P, Brooks G (2001) The coronavirus infectious bronchitis virus nucleoprotein localizes to the nucleolus. J Virol 75:506–512
Hiscox JA, Whitehouse A, Matthews DA (2010) Nucleolar proteomics and viral infection. Proteomics. doi:10.1002/pmic.201000251
Hivin P, Basbous J, Raymond F, Henaff D, Arpin-Andre C, Robert-Hebmann V, Barbeau B, Mesnard JM (2007) The HBZ-SP1 isoform of human T-cell leukemia virus type I represses JunB activity by sequestration into nuclear bodies. Retrovirology 4:14
Hoch-Marchaim H, Hasson T, Rorman E, Cohen S, Hochman J (1998) Nucleolar localization of mouse mammary tumor virus proteins in T-cell lymphomas. Virology 242:246–254
Hong-Yan Z, Murata T, Goshima F, Takakuwa H, Koshizuka T, Yamauchi Y, Nishiyama Y (2001) Identification and characterization of the UL24 gene product of herpes simplex virus type 2. Virus Genes 22:321–327
Huang WH, Yung BY, Syu WJ, Lee YH (2001) The nucleolar phosphoprotein B23 interacts with hepatitis delta antigens and modulates the hepatitis delta virus RNA replication. J Biol Chem 276:25166–25175
Hutzinger R, Feederle R, Mrazek J, Schiefermeier N, Balwierz PJ, Zavolan M, Polacek N, Delecluse HJ, Huttenhofer A (2009) Expression and processing of a small nucleolar RNA from the Epstein-Barr virus genome. PLoS Pathog 5:e1000547
Indik S, Gunzburg WH, Salmons B, Rouault F (2005) A novel, mouse mammary tumor virus encoded protein with Rev-like properties. Virology 337:1–6
Jakob R (1994) Nucleolar accumulation of Semliki Forest virus nucleocapsid C protein: influence of metabolic status, cytoskeleton and receptors. J Med Microbiol 40:389–392
Kalt I, Borodianskiy-Shteinberg T, Schachor A, Sarid R (2010) GLTSCR2/PICT-1, a putative tumor suppressor gene product, induces the nucleolar targeting of the Kaposi’s sarcoma-associated herpesvirus KS-Bcl-2 protein. J Virol 84:2935–2945
Kim SH, Macfarlane S, Kalinina NO, Rakitina DV, Ryabov EV, Gillespie T, Haupt S, Brown JW, Taliansky M (2007a) Interaction of a plant virus-encoded protein with the major nucleolar protein fibrillarin is required for systemic virus infection. Proc Natl Acad Sci USA 104:11115–11120
Kim SH, Ryabov EV, Kalinina NO, Rakitina DV, Gillespie T, MacFarlane S, Haupt S, Brown JW, Taliansky M (2007b) Cajal bodies and the nucleolus are required for a plant virus systemic infection. EMBO J 26:2169–2179
Kojima T, Callea F, Desmyter J, Shikata T, Desmet VJ (1989) Electron microscopy of ribonucleic acid in nuclear particulate aggregates of hepatitis D using nuclease-gold complexes. J Med Virol 28:183–188
Kubota S, Siomi H, Satoh T, Endo S, Maki M, Hatanaka M (1989) Functional similarity of HIV-I rev and HTLV-I rex proteins: identification of a new nucleolar-targeting signal in rev protein. Biochem Biophys Res Commun 162:963–970
Kubota S, Hatanaka M, Pomerantz RJ (1996) Nucleo-cytoplasmic redistribution of the HTLV-I Rex protein: alterations by coexpression of the HTLV-I p21x protein. Virology 220:502–507
Lam YW, Evans VC, Heesom KJ, Lamond AI, Matthews DA (2010) Proteomics analysis of the nucleolus in adenovirus-infected cells. Mol Cell Proteomics 9:117–130
Langenberg WG, Zhang L (1997) Immunocytology shows the presence of tobacco etch virus P3 protein in nuclear inclusions. J Struct Biol 118:243–247
Lawrence FJ, McStay B, Matthews DA (2006) Nucleolar protein upstream binding factor is sequestered into adenovirus DNA replication centres during infection without affecting RNA polymerase I location or ablating rRNA synthesis. J Cell Sci 119:2621–2631
Lechertier T, Sirri V, Hernandez-Verdun D, Roussel P (2007) A B23-interacting sequence as a tool to visualize protein interactions in a cellular context. J Cell Sci 120:265–275
Lee CH, Chang SC, Chen CJ, Chang MF (1998) The nucleolin binding activity of hepatitis delta antigen is associated with nucleolus targeting. J Biol Chem 273:7650–7656
Lee TW, Blair GE, Matthews DA (2003) Adenovirus core protein VII contains distinct sequences that mediate targeting to the nucleus and nucleolus, and colocalization with human chromosomes. J Gen Virol 84:3423–3428
Lee TW, Lawrence FJ, Dauksaite V, Akusjarvi G, Blair GE, Matthews DA (2004) Precursor of human adenovirus core polypeptide Mu targets the nucleolus and modulates the expression of E2 proteins. J Gen Virol 85:185–196
Lee C, Hodgins D, Calvert JG, Welch SK, Jolie R, Yoo D (2006) Mutations within the nuclear localization signal of the porcine reproductive and respiratory syndrome virus nucleocapsid protein attenuate virus replication. Virology 346:238–250
Leenadevi T, Dalziel RG (2009) The alcelaphine herpesvirus-1 ORF 57 encodes a nuclear shuttling protein. Vet Res Commun 33:409–419
Leopardi R, Roizman B (1996) Functional interaction and colocalization of the herpes simplex virus 1 major regulatory protein ICP4 with EAP, a nucleolar-ribosomal protein. Proc Natl Acad Sci USA 93:4572–4576
Li YJ, Macnaughton T, Gao L, Lai MM (2006) RNA-templated replication of hepatitis delta virus: genomic and antigenomic RNAs associate with different nuclear bodies. J Virol 80:6478–6486
Lin NS, Hsieh CE, Hsu YH (1996) Capsid protein of cucumber mosaic virus accumulates in the nuclei and at the periphery of the nucleoli in infected cells. Arch Virol 141:727–732
Liu JL, Lee LF, Ye Y, Qian Z, Kung HJ (1997) Nucleolar and nuclear localization properties of a herpesvirus bZIP oncoprotein, MEQ. J Virol 71:3188–3196
Lopez MR, Schlegel EF, Wintersteller S, Blaho JA (2008) The major tegument structural protein VP22 targets areas of dispersed nucleolin and marginalized chromatin during productive herpes simplex virus 1 infection. Virus Res 136:175–188
Lower R, Tonjes RR, Korbmacher C, Kurth R, Lower J (1995) Identification of a Rev-related protein by analysis of spliced transcripts of the human endogenous retroviruses HTDV/HERV-K. J Virol 69:141–149
Lutz P, Kedinger C (1996) Properties of the adenovirus IVa2 gene product, an effector of late-phase-dependent activation of the major late promoter. J Virol 70:1396–1405
Lymberopoulos MH, Pearson A (2007) Involvement of UL24 in herpes-simplex-virus-1-induced dispersal of nucleolin. Virology 363:397–409
Lymberopoulos MH, Pearson A (2010) Relocalization of upstream binding factor to viral replication compartments is UL24 independent and follows the onset of herpes simplex virus 1 DNA synthesis. J Virol 84:4810–4815
Lymberopoulos MH, Bourget A, Abdeljelil NB, Pearson A (2010) Involvement of the UL24 protein in herpes simplex virus 1-induced dispersal of B23 and in nuclear egress. Virology 412:341–348
MacLean CA, Rixon FJ, Marsden HS (1987) The products of gene US11 of herpes simplex virus type 1 are DNA-binding and localize to the nucleoli of infected cells. J Gen Virol 68:1921–1937
Macnaughton TB, Gowans EJ, Reinboth B, Jilbert AR, Burrell CJ (1990) Stable expression of hepatitis delta virus antigen in a eukaryotic cell line. J Gen Virol 71:1339–1345
Matthews DA, Russell WC (1998) Adenovirus core protein V is delivered by the invading virus to the nucleus of the infected cell and later in infection is associated with nucleoli. J Gen Virol 79:1671–1675
Mayer C, Grummt I (2005) Cellular stress and nucleolar function. Cell Cycle 4:1036–1038
Mears WE, Lam V, Rice SA (1995) Identification of nuclear and nucleolar localization signals in the herpes simplex virus regulatory protein ICP27. J Virol 69:935–947
Melen K, Kinnunen L, Fagerlund R, Ikonen N, Twu KY, Krug RM, Julkunen I (2007) Nuclear and nucleolar targeting of influenza A virus NS1 protein: striking differences between different virus subtypes. J Virol 81:5995–6006
Michel MR, Elgizoli M, Dai Y, Jakob R, Koblet H, Arrigo AP (1990) Karyophilic properties of Semliki Forest virus nucleocapsid protein. J Virol 64:5123–5131
Michienzi A, Cagnon L, Bahner I, Rossi JJ (2000) Ribozyme-mediated inhibition of HIV 1 suggests nucleolar trafficking of HIV-1 RNA. Proc Natl Acad Sci USA 97:8955–8960
Miron MJ, Blanchette P, Groitl P, Dallaire F, Teodoro JG, Li S, Dobner T, Branton PE (2009) Localization and importance of the adenovirus E4orf4 protein during lytic infection. J Virol 83:1689–1699
Mohammadi H, Sharif S, Rowland RR, Yoo D (2009) The lactate dehydrogenase-elevating virus capsid protein is a nuclear-cytoplasmic protein. Arch Virol 154:1071–1080
Morency E, Coute Y, Thomas J, Texier P, Lomonte P (2005) The protein ICP0 of herpes simplex virus type 1 is targeted to nucleoli of infected cells. Brief report. Arch Virol 150:2387–2395
Mori Y, Okabayashi T, Yamashita T, Zhao Z, Wakita T, Yasui K, Hasebe F, Tadano M, Konishi E, Moriishi K, Matsuura Y (2005) Nuclear localization of Japanese encephalitis virus core protein enhances viral replication. J Virol 79:3448–3458
Munday DC, Emmott E, Surtees R, Lardeau CH, Wu W, Duprex WP, Dove BK, Barr JN, Hiscox JA (2010) Quantitative proteomic analysis of A549 cells infected with human respiratory syncytial virus. Mol Cell Proteomics 9:2438–2459
Narayan M, Younis I, D’Agostino DM, Green PL (2003) Functional domain structure of human T-cell leukemia virus type 2 rex. J Virol 77:12829–12840
Ning B, Shih C (2004) Nucleolar localization of human hepatitis B virus capsid protein. J Virol 78:13653–13668
Nosaka T, Siomi H, Adachi Y, Ishibashi M, Kubota S, Maki M, Hatanaka M (1989) Nucleolar targeting signal of human T-cell leukemia virus type I rex-encoded protein is essential for cytoplasmic accumulation of unspliced viral mRNA. Proc Natl Acad Sci USA 86:9798–9802
Okuwaki M, Iwamatsu A, Tsujimoto M, Nagata K (2001) Identification of nucleophosmin/B23, an acidic nucleolar protein, as a stimulatory factor for in vitro replication of adenovirus DNA complexed with viral basic core proteins. J Mol Biol 311:41–55
Olson MO (2009) Induction of apoptosis by viruses: what role does the nucleolus play? Cell Cycle 8:3452–3453
Orsini MJ, Debouck CM (1996) Inhibition of human immunodeficiency virus type 1 and type 2 Tat function by transdominant Tat protein localized to both the nucleus and cytoplasm. J Virol 70:8055–8063
Peeples ME, Wang C, Gupta KC, Coleman N (1992) Nuclear entry and nucleolar localization of the Newcastle disease virus (NDV) matrix protein occur early in infection and do not require other NDV proteins. J Virol 66:3263–3269
Pei Y, Hodgins DC, Lee C, Calvert JG, Welch SK, Jolie R, Keith M, Yoo D (2008) Functional mapping of the porcine reproductive and respiratory syndrome virus capsid protein nuclear localization signal and its pathogenic association. Virus Res 135:107–114
Peranen J, Rikkonen M, Liljestrom P, Kaariainen L (1990) Nuclear localization of Semliki Forest virus-specific nonstructural protein nsP2. J Virol 64:1888–1896
Pyper JM, Clements JE, Zink MC (1998) The nucleolus is the site of Borna disease virus RNA transcription and replication. J Virol 72:7697–7702
Qi D, Omarov RT, Scholthof KB (2008) The complex subcellular distribution of satellite panicum mosaic virus capsid protein reflects its multifunctional role during infection. Virology 376:154–164
Rajamaki ML, Valkonen JP (2009) Control of nuclear and nucleolar localization of nuclear inclusion protein a of picorna-like Potato virus A in Nicotiana species. Plant Cell 21:2485–2502
Realdon S, Gerotto M, Dal Pero F, Marin O, Granato A, Basso G, Muraca M, Alberti A (2004) Proapoptotic effect of hepatitis C virus CORE protein in transiently transfected cells is enhanced by nuclear localization and is dependent on PKR activation. J Hepatol 40:77–85
Reed ML, Dove BK, Jackson RM, Collins R, Brooks G, Hiscox JA (2006) Delineation and modelling of a nucleolar retention signal in the coronavirus nucleocapsid protein. Traffic 7:833–848
Reed ML, Howell G, Harrison SM, Spencer KA, Hiscox JA (2007) Characterization of the nuclear export signal in the coronavirus infectious bronchitis virus nucleocapsid protein. J Virol 81:4298–4304
Rikkonen M, Peranen J, Kaariainen L (1992) Nuclear and nucleolar targeting signals of Semliki Forest virus nonstructural protein nsP2. Virology 189:462–473
Rikkonen M, Peranen J, Kaariainen L (1994) Nuclear targeting of Semliki Forest virus nsP2. Arch Virol Suppl 9:369–377
Risco C, Menendez-Arias L, Copeland TD, Pinto da Silva P, Oroszlan S (1995) Intracellular transport of the murine leukemia virus during acute infection of NIH 3T3 cells: nuclear import of nucleocapsid protein and integrase. J Cell Sci 108:3039–3050
Rojas MR, Jiang H, Salati R, Xoconostle-Cazares B, Sudarshana MR, Lucas WJ, Gilbertson RL, (2001) Functional analysis of proteins involved in movement of the monopartite begomovirus, Tomato yellow leaf curl virus. Virology 291:110–125
Rossi JJ, June CH, Kohn DB (2007) Genetic therapies against HIV. Nat Biotechnol 25:1444–1454
Rowland RRR, Yoo D (2003) Nucleolar-cytoplasmic shuttling of PRRSV nucleocapsid protein: a simple case of molecular mimicry or the complex regulation by nuclear import, nucleolar localization and nuclear export signal sequences. Vir Res 95:23–33
Rowland RR, Kerwin R, Kuckleburg C, Sperlich A, Benfield DA (1999) The localisation of porcine reproductive and respiratory syndrome virus nucleocapsid protein to the nucleolus of infected cells and identification of a potential nucleolar localization signal sequence. Vir Res 64:1–12
Rowland RRR, Schneider P, Fang Y, Wootton S, Yoo D, Benfield DA (2003) Peptide domains involved in the localization of the porcine reproductive and respiratory syndrome virus nucleocapsid protein to the nucleolus. Virology 316:135–145
Ruben S, Perkins A, Purcell R, Joung K, Sia R, Burghoff R, Haseltine WA, Rosen CA (1989) Structural and functional characterization of human immunodeficiency virus tat protein. J Virol 63:1–8
Ruggieri A, Maldener E, Sauter M, Mueller-Lantzsch N, Meese E, Fackler OT, Mayer J (2009) Human endogenous retrovirus HERV-K(HML-2) encodes a stable signal peptide with biological properties distinct from Rec. Retrovirology 6:17
Ryabov EV, Oparka KJ, Santa Cruz S, Robinson DJ, Taliansky ME (1998) Intracellular location of two groundnut rosette umbravirus proteins delivered by PVX and TMV vectors. Virology 242, 303–313
Ryabov EV, Kim SH, Taliansky M (2004) Identification of a nuclear localization signal and nuclear export signal of the umbraviral long-distance RNA movement protein. J Gen Virol 85:1329–1333
Sagou K, Uema M, Kawaguchi Y (2010) Nucleolin is required for efficient nuclear egress of herpes simplex virus type 1 nucleocapsids. J Virol 84:2110–2121
Salsman J, Zimmerman N, Chen T, Domagala M, Frappier L (2008) Genome-wide screen of three herpesviruses for protein subcellular localization and alteration of PML nuclear bodies. PLoS Pathog 4:e1000100
Saltarelli MJ, Schoborg R, Pavlakis GN, Clements JE (1994) Identification of the caprine arthritis encephalitis virus Rev protein and its cis-acting Rev-responsive element. Virology 199:47–55
Schoborg RV, Clements JE (1994) The Rev protein of visna virus is localized to the nucleus of infected cells. Virology 202:485–490
Scott MS, Boisvert FM, Lamond AI, Barton GJ (2011) PNAC: a protein nucleolar association classifier. BMC Genomics 12:74
Sharma P, Ikegami M (2009) Characterization of signals that dictate nuclear/nucleolar and cytoplasmic shuttling of the capsid protein of Tomato leaf curl Java virus associated with DNA beta satellite. Virus Res 144:145–153
Siomi H, Shida H, Nam SH, Nosaka T, Maki M, Hatanaka M (1988) Sequence requirements for nucleolar localization of human T cell leukemia virus type I pX protein, which regulates viral RNA processing. Cell 55:197–209
Sonntag F, Schmidt K, Kleinschmidt JA (2010) A viral assembly factor promotes AAV2 capsid formation in the nucleolus. Proc Natl Acad Sci USA 107:10220–10225
Spencer KA, Dee M, Britton P, Hiscox JA (2008) Role of phosphorylation clusters in the biology of the coronavirus infectious bronchitis virus nucleocapsid protein. Virology 370:373–381
Stow ND, Evans VC, Matthews DA (2009) Upstream-binding factor is sequestered into herpes simplex virus type 1 replication compartments. J Gen Virol 90:69–73
Strang BL, Boulant S, Coen DM (2010) Nucleolin associates with the human cytomegalovirus DNA polymerase accessory subunit UL44 and is necessary for efficient viral replication. J Virol 84:1771–1784
Szekely L, Jiang WQ, Pokrovskaja K, Wiman KG, Klein G, Ringertz N (1995) Reversible nucleolar translocation of Epstein-Barr virus-encoded EBNA-5 and hsp70 proteins after exposure to heat shock or cell density congestion. J Gen Virol 76:2423–2432
Thiry M, Lafontaine DL (2005) Birth of a nucleolus: the evolution of nucleolar compartments. Trends Cell Biol 15:194–199
Tijms MA, van der Meer Y, Snijder EJ (2002) Nuclear localization of non-structural protein 1 and nucleocapsid protein of equine arteritis virus. J Gen Virol 83:795–800
Tollefson AE, Ying B, Doronin K, Sidor PD, Wold WS (2007) Identification of a new human adenovirus protein encoded by a novel late l-strand transcription unit. J Virol 81:12918–12926
Trinkle-Mulcahy L, Boulon S, Lam YW, Urcia R, Boisvert FM, Vandermoere F, Morrice NA, Swift S, Rothbauer U, Leonhardt H, Lamond A (2008) Identifying specific protein interaction partners using quantitative mass spectrometry and bead proteomes. J Cell Biol 183:223–239
Tsai CW, Redinbaugh MG, Willie KJ, Reed S, Goodin M, Hogenhout SA (2005) Complete genome sequence and in planta subcellular localization of maize fine streak virus proteins. J Virol 79:5304–5314
Tsuda Y, Mori Y, Abe T, Yamashita T, Okamoto T, Ichimura T, Moriishi K, Matsuura Y (2006) Nucleolar protein B23 interacts with Japanese encephalitis virus core protein and participates in viral replication. Microbiol Immunol 50:225–234
Uchil PD, Kumar AV, Satchidanandam V (2006) Nuclear localization of flavivirus RNA synthesis in infected cells. J Virol 80:5451–5464
Uhrig JF, Canto T, Marshall D, MacFarlane SA (2004) Relocalization of nuclear ALY proteins to the cytoplasm by the tomato bushy stunt virus P19 pathogenicity protein. Plant Physiol 135:2411–2423
Unwalla HJ, Li H, Li SY, Abad D, Rossi JJ (2008) Use of a U16 snoRNA-containing ribozyme library to identify ribozyme targets in HIV-1. Mol Ther 16:1113–1119
Vazquez-Iglesias L, Lostale-Seijo I, Martinez-Costas J, Benavente J (2009) Avian reovirus sigmaA localizes to the nucleolus and enters the nucleus by a nonclassical energy- and carrier-independent pathway. J Virol 83:10163–10175
Volmer R, Mazel-Sanchez B, Volmer C, Soubies SM, Guerin JL (2010) Nucleolar localization of influenza A NS1: striking differences between mammalian and avian cells. Virol J 7:63
Waggoner S, Sarnow P (1998) Viral ribonucleoprotein complex formation and nucleolar-cytoplasmic relocalization of nucleolin in poliovirus-infected cells. J Virol 72:6699–6709
Walton TH, Moen PT, Jr, Fox E, Bodnar JW (1989) Interactions of minute virus of mice and adenovirus with host nucleoli. J Virol 63:3651–3660
Wang SH, Syu WJ, Huang KJ, Lei HY, Yao CW, King CC, Hu ST (2002) Intracellular localization and determination of a nuclear localization signal of the core protein of dengue virus. J Gen Virol 83:3093–3102
Westaway EG, Khromykh AA, Kenney MT, Mackenzie JM, Jones MK (1997) Proteins C and NS4B of the flavivirus Kunjin translocate independently into the nucleus. Virology 234:31–41
Wistuba A, Kern A, Weger S, Grimm D, Kleinschmidt JA (1997) Subcellular compartmentalization of adeno-associated virus type 2 assembly. J Virol 71:1341–1352
Wright KM, Cowan GH, Lukhovitskaya NI, Tilsner J, Roberts AG, Savenkov EI, Torrance L (2010) The N-terminal domain of PMTV TGB1 movement protein is required for nucleolar localization, microtubule association, and long-distance movement. Mol Plant Microbe Interact 23:1486–1497
Wurm T, Chen H, Britton P, Brooks G, Hiscox JA (2001) Localisation to the nucleolus is a common feature of coronavirus nucleoproteins and the protein may disrupt host cell division. J Virol 75:9345–9356
Xing J, Wu F, Pan W, Zheng C (2010) Molecular anatomy of subcellular localization of HSV-1 tegument protein US11 in living cells. Virus Res 153:71–81
Yamada H, Jiang YM, Zhu HY, Inagaki-Ohara K, Nishiyama Y (1999) Nucleolar localization of the UL3 protein of herpes simplex virus type 2. J Gen Virol 80:2157–2164
Yoo D, Wootton SK, Li G, Song C, Rowland RR (2003) Colocalization and interaction of the porcine arterivirus nucleocapsid protein with the small nucleolar RNA-associated protein fibrillarin. J Virol 77:12173–12183
You J, Dove BK, Enjuanes L, DeDiego ML, Alvarez E, Howell G, Heinen P, Zambon M, Hiscox JA (2005) Subcellular localization of the severe acute respiratory syndrome coronavirus nucleocapsid protein. J Gen Virol 86:3303–3310
You JH, Howell G, Pattnaik AK, Osorio FA, Hiscox JA (2008) A model for the dynamic nuclear/nucleolar/cytoplasmic trafficking of the porcine reproductive and respiratory syndrome virus (PRRSV) nucleocapsid protein based on live cell imaging. Virology 378:34–47
Yuan X, Shan Y, Zhao Z, Chen J, Cong Y (2005) G0/G1 arrest and apoptosis induced by SARS-CoV 3b protein in transfected cells. Virol J 2:66
Zhu HY, Yamada H, Jiang YM, Yamada M, Nishiyama Y (1999) Intracellular localization of the UL31 protein of herpes simplex virus type 2. Arch Virol 144:1923–1935
Acknowledgements
DAM and JAH would like to acknowledge their co-workers and collaborators over the years for developing viral interactions with the nucleolus. DAM’s research on the nucleolus is supported by the Wellcome Trust and JAH’s by the BBSRC and a Leverhulme Trust Research Fellowship. EE is supported by a BBSRC Astbury DTG studentship.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2011 Springer Science+Business Media, LLC
About this chapter
Cite this chapter
Matthews, D., Emmott, E., Hiscox, J. (2011). Viruses and the Nucleolus. In: Olson, M. (eds) The Nucleolus. Protein Reviews, vol 15. Springer, New York, NY. https://doi.org/10.1007/978-1-4614-0514-6_14
Download citation
DOI: https://doi.org/10.1007/978-1-4614-0514-6_14
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4614-0513-9
Online ISBN: 978-1-4614-0514-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)