
Abstract
Panus natarajanus sp. nov. from the Western Ghats of Karnataka, India, is described, illustrated and discussed along with P. velutinus, P. tephroleucus, P. ciliatus, P. hookerianus, Lentinus fasciatus, L. concentricus, and L. megacystidiatus. Lentinus alpacus is re-allocated to Panus as P. alpacus comb, nov., and contrasted with P. parvus. A short description with illustrations is provided of L. concinnus, which is also newly reported from the Western Ghats. A synoptic key for all species of lentinoid fungi of the Western Ghats is provided.
Similar content being viewed by others

Introduction
The generic demarcation and phylogenetic relationships between Lentinus and Panus and their putative allies Neolentinus, Heliocybe, and Pleurotus has been controversial (Küehner 1980, Corner 1981, Pegler 1983a, Redhead & Ginns 1985, Singer 1975, 1986, Hibbett & Vilgalys 1991, 1993, Rune 1994, Thorn et al. 2000, Garcia-Sandoval et al. 2011). Corner (1981) considered Lentinus and Panus as separate genera on the basis of their hyphal system and included some species placed in Pleurotus by Singer (1975), whereas, Pegler (1983a) treated Panus as a subgenus of Lentinus and included several species of Singer’s Pleurotus in Panus. Singer (1986) included Lentinus crinitus (the widely accepted lectotype species of Lentinus) and Panus conchatus (the conserved type species of Panus) in Panus, and Lentinus was used for Neolentinus lepideus and Lentinula edodes. However, Pegler (1975) separated Lentinula edodes from Lentinus in the basis of the non-decurrent lamellae and a monomitic hyphal system with inflated, sclerified generative hyphae. Many subsequent authors have accepted that Lentinus and Panus are separate genera (Corner 1981, Moser 1978, Singer 1975, 1986, Thorn et al. 2000). Recent studies using RFLP, and cladistic analyses of rDNA sequence data along with morphological characters, support Lentinus subgen. Lentinus sensu Pegler as equivalent to Lentinus sensu Corner. In addition, a large part of the subgenus Lentinus sensu Pegler, Neolentinus and Panus s. str. is monophyletic whereas, the Lentinus sensu Pegler is polyphyletic (Redhead & Ginns 1985, Hibbett & Vilgalys 1991, 1993, Hibbett & Thorn 1994, Hibbett et al. 1993, Grand et al. 2011, Seelan et al. 2015). Further, in a phylogenetic and phylogenomic overview of Polyporales, Lentinus was nested within the core polyporoid clade, while Panus was nested within the residual polyporoid clade (Binder et al. 2013). Lentinus s. str. is characterized by species with radiate, descending, or intermediate tramas with ligative hyphae and hyphal pegs in the hymenium, whereas Panus s. str. is distinguished by the strongly radiate hymenophoral trama with dimitic hyphae and lacking hyphal pegs. The brown rot fungi Neolentinus and Heliocybe were segregated from Lentinus sections Pulverulenti, Cirrhosi, and Squamosi sensu Pegler (1983a) based on the bipolar mating system and type of wood decay (Redhead & Ginns 1985, Hibbett & Vilgalys 1993, Garcia-Sandoval et al. 2011) and included in Gloeophyllales (Binder et al. 2005, Hibbett & Donoghue 2001, Thorn et al. 2000). Rune (1994) considered Heliocybe to be a synonym of Neolentinus and suggested the transfer of Heliocybe species to Neolentinus. However, in a phylogenetic study on Gloeophyllales, Heliocybe sulcata was placed as the sister group to the clade Neolentinus (Garcia-Sandoval et al. 2011), which supported the retention of Heliocybe and Neolentinus as separate genera. As a result of all these studies, Neolentinus and Heliocybe are now placed in Gloeophyllales, while Lentinula is in Agaricales, and both Lentinus s. str. and Panus s. str. are in Polyporales. Lentinus sensu Pelger is dispersed through at least three orders, Agaricales, Polyporales, and Gloeophyllales.
A review of the literature revealed that 15 species belonging to Lentinus and Panus have been reported from the Western Ghats (Sathe & Daniel 1980, Sathe & Deshpande 1980, Sathe & Kulkarni 1980, Manjula 1983, Pegler 1983a, Manimohan et al. 2004, Natarajan et al. 2005, Farook et al. 2013, Senthilarasu 2014). This group is well explored in the Western Ghats of Kerala where six species of Lentinus and five species of Panus are known: L. bambusinus (Kumar & Manimohan 2005), L. patulus (Mohanan 2011), L. polychrous (Manimohan et al. 2004, Mohanan 2011), L. sajor-caju (Natarajan 1978, Sathe & Daniel 1980, Florence 2004, Manimohan et al. 2004, Pradeep & Vrinda 2007, Varghese et al. 2010, Mohanan 2011; Florence 2004, as Pleurotus sajor-caju; Manimohan et al. 2004, Pradeep & Vrinda 2007, Varghese et al. 2010, Mohanan 2011, all as L. dicholamellatus), L. squarrosulus (Sharma et al. 1985, Florence & Yesodharan 2000, Florence 2004, Manimohan et al. 2004, Pradeep & Vrinda 2007, Varghese et al. 2010, Mohanan 2011), L. tigrinus (Mohanan 2011), P. caespiticola (Manimohan et al. 2004, as L. caespiticola; Manimohan & Leelavathy 1995, as L. caespiticola var. asiaticus), P neostrigosus (Manimohan et al. 2004, Mohanan 2011, as L. strigosus), P. hookerianus (Manimohan et al. 2004, as L. hookerianus), P. indicus (Sathe & Daniel 1980, Florence 2004), and P. similis (Manimohan et al. 2004, as L. similis).
The diversity of lentinoid fungi from the Western Ghats of Tamil Nadu, Maharashtra, and Karnataka is meager. Lentinus crinitus (Natarajan & Raman 1981), L. squarrosulus (Natarajan & Manjula 1978), P. neostrigosus (Pegler 1983a, as L. strigosus), P. velutinus (Pegler 1983a, as L. velutinus), and P. similis (Pegler 1983a, as L. similis) have been reported from the Western Ghats of Tamil Nadu (Natarajan 1978, Natarajan & Manjula 1978, Natarajan & Raman 1981, Manjula 1983, Pegler 1983a). In the Western Ghats of Maharashtra, L. alpacus (Senthilarasu & Singh 2012), L. connatus (Léville 1846, as L. javanicus; Pegler 1983a), L. sajor-caju (Senthilarasu 2014), and L. squarrosulus (Theissen 1911, as L. aff. subnudus; Pegler 1983a) have been reported. Only one species, P. similis (Pegler 1983a, as L. similis) has been authentically reported from the Western Ghats of Karnataka though Usha & Janardhana (2014) in their biodiversity study, listed L. cladopus and L. sajor-caju from the Western Ghats of Chickmagalur and Hassan.
Here, the generic concept of Corner (1981) is followed for Lentinus and Panus. A new species, Panus natarajanus that differs morphotaxonomically from other reported species is described, illustrated and discussed. Panus alpacus is proposed as a new combination for L. alpacus. A short description with illustrations is provided for L. concinnus, reported here for the first time from Western Ghats, to provide a clear demarcation from the common and widely distributed L. tigrinus. Panus similis is also reported from the Kadamakkal reserve forest of the Western Ghats of Karnataka. In addition, a synoptic key to the Lentinus and Panus species of the Western Ghats and its foothills is provided.
Materials and Methods
Collections were made from different forest regions of the Western Ghats of Maharashtra and Karnataka. The colour terminologies used are from Kornerup & Wanscher (1978). Thin handmade sections were made from dried specimens rehydrated with alcohol. The microscopic observations were made in 10% KOH, stained in 3% phloxine and Cotton blue. The range of spores with extreme values in parentheses precedes average spore dimensions in parenthesis. The spore quotient (Q) represents mean length by mean width of 50 basidiospores measured. The holotype specimens were deposited at Ajrekar Mycological Herbarium (AMH), MACS’ Agharkar Research Institute, Pune, India, and isotypes are preserved in personal collections (Macrofungal Collection of India (MCI) at SRM Research Institute, SRM University, Kattankulathur, Tamil Nadu, India. The key for the lentinoid fungi of Western Ghats is prepared on the basis of published reports. Index Fungorum (www.indexfungorum.org) and Species Fungorum (www.speciesfungorum.org) are followed for nomenclature and currently accepted names, excluding some species as mentioned in the discussion.
Taxonomy
Panus natarajanus Senthil., sp. nov.
MycoBank MB805095

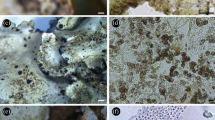
A. Lentinus concinnus in its natural habitat, surface view (AMH 9568 paratype). B. L. concinnus gills view (AMH 9568 paratype). C. Panus natarajanus in its natural habitat, gills view (AMH 9470, holotype). D. P. natarajanus surface view (AMH 9470, holotype).

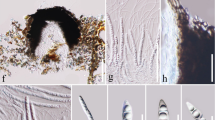
A. Panus natarajanus (AMH 9470, holotype). a. Basidiospores. b. Basidia. c. Cheilocystidia. d. Sclerocystidia. e. Epicuticular generative hyphae. f. Skeletal hyphae. g. Generative hyphae with clamp-connections. B. Lentinus concinnus (AMH 9568, paratype). a. Basidiospores. b. Basidia. c. Cheilocystidia. d. Skeletal hyphae. e. Skeleto-ligative hyphae. f. Inflated generative hyphae. Bars = 10 µm.
Etymology: ‘natarajanus’, in honourof the late Krishnamoorthy Natarajan (1942–2008), an eminent mycologist with incredible knowledge on the fleshy fungi of India.
Diagnosis: Differs from Panus velutinus in the concentrically zonate pileus, shorter and slenderer stipe, and the slightly smallerspores.
Type: India: Karnataka state: Sirsi, 14°27′15.0″N 74°41′60.6″E, 11 Oct. 2012, G. Senthilarasu (AMH 9470 — holotype; MCI 966 — isotype).
Description: Pileus 8–80 mm diam, thin, coriaceous, broadly, deeply infundibuliform, expanding to cyathiform; surface often with pale purplish tints when young, becoming different shades of reddish brown (8D5, 8D6, 8E8, 9E6, 9E7), darker towards the centre and covered by concentrically arranged, dense squamules, <1 mm long, more towards the centre, short hispid, <1 mm long, towards the margin, uniformly hispid when young, often with pale to dark concentric zones especially towards margin; margin at first strongly involute, becoming reflexed, not striate, not ciliate, lobed. Lamellae arcuate, short to deeply decurrent, Venetian red (8D8) to reddish brown (8E8), becoming dark reddish brown (9E8) on drying, often with vinaceous tints when young, narrow, <1mm wide, densely crowded with lamellulae of different lengths; edge entire. Stipe 20–40 × 1–5 mm, slender, cylindric, equal, expanding slightly at the base and apex, solid; surface concolorous with the pileus, short hispid with the indumentums extending into the bases of the lamellae, arising from a pseudosclerotium. Pileal context to 1 mm thick at the centre, white. Basidiospores 4.5–5(−5.5) × 2.5–3 (4.81 ±0.21 × 2.78 ±0.13) µm, Q = 1.73, oblong to cylindric, hyaline, thin-walled, with few visible contents. Basidia 15.5–20 × 3.5–5 µm, narrowly clavate-cylindric, bearing four short, thick sterigmata. Lamella edge sterile with scattered cheilocystidia, soon collapsing. Cheilocystidia 25–30 × 7–9 µm, cylindric-clavate, often sinuous, thin walled. Sclerocystidia abundant on sides of lamellae to scattered on lamellae edge, 22–60 × 4.5-8 µm, initially clavate with refractive gloeo-contents, soon developing a thickened wall up to 1 µm thick, hyaline to brownish, scarcely projecting 15 µm beyond the basidia. Hymenophoral trama irregular, of radiate construction, hyaline, consisting of generative hyphae and skeletal hyphae. Subhymenial layer narrow, irregular. Hypodermium subgelatinized, to 250 µm wide, consisting of hyaline, generative hyphae, to 3 µm diam. Pileal context consisting of a dimitic hyphal system with skeletal and non-inflated generative hyphae, hyaline, thin-walled with abundant clamp-connections; skeletal hyphae 1.5–4 µm diam, thick-walled, wall to 1.5 µm thick, hyaline; generative hyphae to 2.5 µm diam, hyaline, branched, thin-walled with abundant clamp-connections. Pileal surface an indefinite epicutis producing erect, loose fascicles, to 700 µm long, of brown, generative hyphae, 3–7 µm diam, thick-walled, to 2 µm thick, with obtusely rounded apex and abundant clamp-connections.
Habitat: Lignicolous, on an unidentified wood, solitary, scattered to gregarious, in Vateria indica/ Dipterocarpus indicus dominant dipterocarp forest. Known only from the type locality.
Notes: The pantropical Panus velutinus (Pegler 1983a, as L. velutinus) differs from P. natarajanus in having larger basidiomes (pileus 20–175 vs 8–80 mm diam), velutinate, distinctly not zonate or obscurely zonate pileus, elongated, thick stipe (20–250 × 2–10 vs 20–40 × 1–5 mm) and larger spores (5–8 × 3–3.5 vs 4.5–5.5 × 2.5–3 µm). Panus tephroleucus (Pegler 1983a, as L. tephroleucus) differs from P. natarajanus in having a smaller pileus (10–40 vs 8–80 mm diam), yellowish to greyish brown surface, moderately, distantly spaced lamellae, larger basidiospores (6–8 vs 4.5–5.5 µm), and smaller sclerocystidia (22–28 vs 22–60 µm). Panus natarajanus differs from P. ciliatus (Pegler 1983a, as L. ciliatus) in having smaller (8–80 vs 2–150 mm), non striate pileus lacking ciliate margin, shorter and slenderer stipe (20–40 × 1–5 vs 3–130 × 3–15 mm), and larger sclerocystidia (22–60 vs 19–30 µm).
Panus natarajanus closely resembles the Himalayan P. hookerianus (Pegler 1983a, as L. hookerianus) in having similar sized and coloured basidiomes with a hispid pileus and stipe. However, P. natarajanus differs in having shorter hispid hairs, (≤700 µm vs 1–2 mm long), concentrically zonate pileus, slightly smaller spores (4.5–5.5 × 2.5–3 vs 5–6.5 × 2.7–3.5 µm), larger cheilocystidia (25–30 × 7–9 vs 14–19 × 3–4 µm) and sclerocystidia (22–60 × 4.5–8 vs 17–25 × 6–7.5 µm). Lentinus fasciatus (Pegler 1983a) differs from P. natarajanus in having longer hispid hairs (1–3 mm) and spores (6–8.5 × 3–4 µm).
Panus natarajanus morphologically also closely resembles L. concentricus (Karunarathna et al. 2011a) described from Thailand in having somewhat similar sized basidiomes and a concentrically zonate pileus. However, L. concentricus differs morphologically in the yellowish brown to yellowish brown clay pileus, Pompeian yellow lamellae, and microscopically in the slightly larger basidiospores (5–7 × 2.5–4 vs 4.5–5.5 × 2.5–3 µm) and absence of sclerocystidia. AnotherThailand species, L. megacystidiatus (Karunarathna et al. 2011a) differs from P. natarajanus in the greyish orange, smaller pileus (2–20 vs 8–80 mm diam), with densely ciliate margin, greyish orange lamellae, shorter and slenderer stipe (2–14 × 2–4 vs 20–40 × 1–5 mm) and larger basidiospores (6–8 × 3–4 vs 4.5–5.5 × 2.5–3 µm). Both L. concentricus and L. megacystidiatus have a greater resemblance to Panus rather than Lentinus on the basis of morphotaxonomic characters.
Panus alpacus (Senthil. & S.K. Singh) Senthil., comb. nov.
MycoBank MB805096
Basionym: Lentinus alpacus Senthil. &S.K. Singh, Mycotaxon 121: 70 (2013) [“2012”].
Type: India: Maharashtra state: Pune, Pune University Campus, 18°31′18.4″N 73°49′53.6″E, on decaying twigs, solitary, 21 Jul. 2011, G. Senthilarasu (AMH 9442 — holotype).
Additional material examined: India: Maharashtrastate: Pune, Pune University Campus, 18°31′18.4″N 73°49′53.6″E, on an unidentified twig, solitary to scattered, 3 Aug. 2009, G. Senthilarasu (AMH 9457); loc. cit, 26 July 2012, G. Senthilarasu (AMH 9526), 1 Aug. 2012 G. Senthilarasu (AMH 9527, MCI 951).
Notes: Senthilarasu & Singh (2012) described this species as Lentinus alpacus from the Western Ghats of Maharashtra and it is known only from the type locality. However, the distinguishing morphotaxonomic characters of skeletal hyphae in the context, the lack of hyphal pegs, and the presence of metuloids in the hymenium, agree with the genus Panus sensu Corner, and hence the new combination Panus alpacus is made here.
Panus alpacus has already been compared with L. courtetianus, P. tephroleucus, and P. ciliatus (Senthilarasu & Singh 2012). Panus alpacus morphologically also differs from another small (10–20 mm pileal diam) species, Panus parvus (Drechsler-Santos et al. 2012) described from Brazil, in having a reddish brown pileus with shorter squamules (≤840 µm long) more scattered towards the disc rather than blackish, spiniform, longer squamules (to 1 mm long) on the pileus, more towards the margin (ciliate), and longer and slenderer stipe (14–25 × 1–1.5 vs 5–15 × 1–4 mm). In addition, P. parvus differs microscopically from P. alpacus in the larger gloeocystidia (40–75 × 8–17 vs 18–45 × 5–11 µm) and metuloids (30–80 × 9–13 vs 19–25 × 7–11.5 µm).
Panus similis (Berk. & Broome) T.W. May & A.E. Wood, Mycotaxon 54: 148 (1995).
Material examined: India: Karnataka state: Kadamakkal Reserve Forest, Manadukka, 12°30′N 79°39′E, on wood, solitary to scattered, 10 Aug. 2010, G. Senthilarasu (MCI 1002); Uppangala forest, (12°30′N 79°39′E), 12 Aug. 2010, G. Senthilarasu (MCI 1005).
Notes: Panus similis differs from P. velutinus, P. ciliatus, and P. hookerianus in the strongly plicate striate pileus. Panus similis is a common species that was earlier reported as L. similis from the Nagarhole wildlife sanctuary in the Coorg district of Karnataka (Pegler 1983a). The morphotaxonomic characters of the present collections agree with the description of L. similis provided by Pegler (1983a). The basidiospore measurements of our collections are 4.5–6 × 2–3 µm.
Sathe & Kulkarni (1980) reported L. velutinus from Sampaje in the Western Ghats of Karnataka. The original description and illustrations of this species indicate the pileal surface is sulcate striate which is the distinguished character of P. similis. Their report may represent P. similis in having a sulcate striate pileus, shorter stipe (30–67 × 5–7 mm) and smaller basidiospores (4.5–6 × 2.3–4 µm). However, the precise identity cannot be determined on the basis of presence of metuloidal cystidia in the hymenium.
Lentinus concinnus Pat., Bull. Soc. mycol. Fr. 8(2): 47 (1892).
Description: Pileus 10–40 mm diam, convex, becoming broadly convex to plane, narrowly, shallowly to deeply depressed; surface initially with dark violaceous tints covered by light brown squamules (7D5), squamules becoming yellowish white (4A2) to pale yellow (4A3) to greyish yellow (4B3), covered by light brown (7D5) to dark brown (9F6) to reddish brown (9F7) to violet brown (10F6–10F8), appressed, superficial, squamules, more numerous towards the disc, pellucid to tuberculate striate; margin initially incurved, becoming decurved to plane, faintly to distinctly striate, ciliate when young. Gills subdecurrent, light orange (5A4) to greyish orange (5B4), becoming yellowish white (4A2) to pale yellow (4A3) to yellowish white (2A2), rarely furcated, regularly intervenose, moderately close; margin serrulate. Stipe 12–22 × 1–3 mm, central to eccentric; surface yellowish white (4A2) to greyish yellow (4B3) to greyish orange (5B3), covered by loose, scattered, superficial, light brown (7D5) squamules, leathery, solid becoming stuffed. Basidiospores (5.5−)6.5–7.5(−9.5) × 3–3.5(−4.0) (7.11 ±0.54 × 3.31 ±0.21) µm, Q =2.14, cylindrical. Basidia 20–28 × 4–6 µm, narrowly clavate. Lamella edge sterile. Cheilocystidia 13.5–33 × 4.5–6.5 µm, clavate to fusoid, often sinuous-nodulose. Hyphal pegs scattered, 38–70 × 20–50 µm, projecting 40–60 µm beyond the hymenial layer. Pileal context trimitic, with skeletal, skeleto-ligative and generative hyphae; generative hyphae inflated to 20 µm diam.
Material examined: India: Maharashtra state: Sinhgad, 18°31′18.4″N 73°49′53.6″E, solitary to caespitose, on an unidentified log, 30 June 2012, G. Senthilarasu (AMH 9568).
Notes: The morphotaxonomic characters of the present collection agree with the description of L. concinnus provided by Pegler (1983a). However, it differs in having slightly larger spores and basidia and more inflated generative hyphae. Lentinus concinnus is easily confused with L. tigrinus. The former is clearly distinguished from L. tigrinus in the field by the smaller basidiomes, whitish to cream pileus, covered by reddish brown to violet brown squamules, regularly interveined, moderately close lamellae, and shorter, slenderer stipe. Lentinus tigrinus has larger basidiomes (≤10 cm diam pileus), a whitish to cream pileus with greyish brown to blackish squamules, is occasionally slightly interveined, crowded lamellae and has an elongated, thick stipe (≤10 mm). Although this species has already been reported from Chennai, Tamil Nadu (Pegler 1983a), this is the first record from the Western Ghats region.
Key to species of Lentinus and Panus occurring in Western Ghats
-
1
Context having thick walled branched skeleto-ligative hyphae; hyphal pegs present (rarely absent in one species); hymenium lacking gloeocystidia and thick-walled metuloids or sclerocystidia ..................................................... 2
Context having thickwalled, unbranched skeletal hyphae; hyphal pegs never present; hymenium having conspicuous gloeocystidia and thick-walled metuloids or sclerocystidia ..................................................... 8
-
2
(1) Pileus often strigose, glabrescent at the center; surface yellowish brown to reddish brown, radially covered by concolorous, fibrillose hairs; margin not involute, distinctly ciliate; cilia cream to yellowish brown; stipe paler or concolorous with the pileus, covered by scattered, white to yellowish brown, scurfy squamules ............. L. crinitus Pileus surface not strigose, often squamulose, squamules appressed ..................................................... 3
-
3
(2) Generative hyphae inflated, ≤ 20 µm diam; lamellae edge dentate ..................................................... 4
Generative hyphae not inflated, ≤6 µm diam; lamellae entire or denticulate ............................................... 5
-
4
(3) Pileus small to large, 10–100 mm diam; surface initially greyish to blackish brown, becoming pale ochraceus to cream to white, covered by greyish brown to blackish brown squamules; lamellae occasionally, slightly interveining; stipe 20–100 × 2–10 mm ..................................................... L. tigrinus Pileus small, 10–40 mm diam, surface initially with violaceous tints, becoming yellowish white to pale yellow to greyish yellow, covered by light brown to dark brown to reddish brown to violet brown squamules; lamellae rarely furcated, regularly intervenose; stipe short and slender, 12–22 × 1–3 mm ............................................... L. concinnus
-
5
(3) Hyphal pegs absent; pileal surface white to cream, glabrous to minute, fugacious, brown to blackish squamulose; margin sulcate striate, finely eroded; lamellae edge entire .................................................. L. patulus
Hyphal pegs present, abundant; pileal surface glabrous to squamulose to squamose; margin not sulcate striate; lamellae finely denticulate .......................................................... 6
-
6
(5) Pileal surface variable in colour, at first whitish, becoming yellowish white to pale yellow greyish yellow or pale orange to brownish orange, glabrous with appressed reddish brown squamules especially towards the centre, finely radially striate to translucent striate, entire, becoming rimose; lamellae whitish; annulus present, fugacious ..................................................... L. sajor-caju
Pileus surface tomentose to appressed to recurved fibrillose squamulose .................................................. 7
-
7
(6) Pileal surface white to cream to pale yellow brown, becoming yellowish, covered by appressed to recurved, concolorous to greyish to fuscous brown squamules; lamellae white to pale yellowish brown ........................................................... L. squarrosulus
Pileal surface ochraceous-cream to blond to greyish yellow to fuscous brown, covered by concentrically arranged, fine tomentum with recurved, fibrillose squamules; lamellae pale greyish black to greyish brown to reddish brown with reddish to purplish tints ........................................................... L. polychrous
-
8
(1) Hymenium having conspicuous refractive gloeocystidia and or thick walled metuloids .......................................... 9
Gloeocystidia and or thick walled metuloids absent; small sclerocystidia occasionally present ..................... 13
-
9
(8) Pileus villose to hispid strigose or squamulose ..................................................... 10
Pileus fibrillose squamulose to tomentose, becoming glabrous ..................................................... 12
-
10
(9) Pileus flabelliform, small (10–30 mm diam); surface pale brown, becoming darkerwith age, covered by long concolorous, tuft of hairs; lamellae creamish white, becoming brown on drying; stipe reduced (5–10 × 2–3 mm), lateral ..................................................... P. indicus
Pileus infundibuliform, small to large (6–100 mm diam); stipe well developed, central to eccentric ............................... 11
-
11
(10) Basidiomes mostly central, rarely eccentric, sometimes stipe branched, bearing two pilei; pileus very small (6–25 mm diam), surface Venetian red to Persian red to reddish brown bearing reddish brown squamules, more towards the disc;stipe short and slender (14–25 × 1–1.5 mm); gloeocystidia and metuloids present ..................................................... P. alpacus
Basidiomes mostly eccentric to lateral, rarely central; pileus small to large (20–100 mm diam), surface white to yellowish brown, covered by villose to hispid tomentose, more towards the margin; stipe to 10 mm thick; gloeocystidia absent, metuloids present .......................... P. neostrigosus (as L. strigosus)
-
12
(9) Basidiomes often tufted with dead roots and bamboo rhizomes; pileus small to large, to 200 mm diam, surface initially reddish grey, becoming white to yellowish white to pale yellow, finely appressed squamulose, becoming glabrous; lamellae dichotomously furcate; stipe glabrous, becoming tomentose to strigose ..................................................... L. bambusinus
Basidiomes always found to be associated with dead grass stems and roots; pileus small, to 30 mm diam, surface orange white to pale ochre, finely tomentose, becoming glabrous; lamellae not forked; stipe finely pruinose .................................... P. caespiticola (as L. caespiticola and L. caespiticola var. asiaticus)
-
13
(8) Basidiomes robust, often caespitose, centrally stipitate to eccentric to lateral; pileal surface white to yellowish brown, becoming darker towards center, initially tomentose fibrillose, becoming glabrescent; margin neither plicate nor striate, not ciliate; lamellae white, becoming yellowish brown; stipe whitish to cream, becoming yellowish brown to dark brown ..................................................... L. connatus
Pileal surface densely hispid strigose or squamulose to velutinate or if glabrescent then strongly plicate striate ................................................................. 14
-
14
(13) Basidiomes medium to large; pileus 20–45(−80) mm diam; surface pale brown to cinnamon brown to dark brown, covered by hispid strigose with short erect hairs (0.5–2 mm); margin neither striate to sulcate striate nor zonate, densely ciliate; stipe concolorous with the pileus, velutinate to short hispid ................................................................ P. hookerianus (as L. hookerianus)
Pileus almost smooth or squamulose to short hispid; margin either striate to sulcate striate or concentrically zonate .................................................................. 15
-
15
(14) Pileus 20–80 mm diam; surface initially with pinkish tints, becoming uniformly, pale brown to cinnamon brown, initially velutinate, becoming almost smooth, strongly plicate striate, striae extending up to the disc; margin concentrically not zonate; lamellae whitish; stipe central; surface concolorous with the pileus, felty; basidiospores 4.5–6 × 2–3 µm .................................... P. similis (as L. similis)
Pileus velutinate to hispid to squamulose, neither smooth nor plicate striate; margin zonate or not zonate .......................................................................... 16
-
16
(15) Pileus 8–80 mm diam; surface reddish brown, darker at the center, initially short hispid, becoming squamulose at the center, to short hispid towards margin, often with pale and dark concentric zones near margin; margin not ciliate; lamellae reddish brown, often with vinaceous tints; stipe short and slender (20–40 × 1–5 mm); surface concolorous with the pielus, short hispid; basidiospores small, 4.5–5.5 × 2.5–3 µm ......................................................................... P. natarajanus
Pileus 20–175 mm diam; surface pale brown to orange brown, reddish brown to reddish yellow to brownish yellow, velutinate to short hispid or subsquamulose, not zonate or obscurely zonate; margin densely ciliate; stipe very long and thick (20–250 × 2–10 mm); surface concolorous with the pileus, velutinate; basidiospores 5–8 × 3–3.5 µm ......................................... P. velutinus (as L. velutinus)
Excluded Species
Lentinus dicholamellatus Manim., Mycotaxon 90: 312 (2004).
Notes: Manimohan et al. (2004) described this species, Lentinus dicholamellatus, from Kerala and placed it in Lentinus subgen. Lentinus sect. Dicholamellatae, sensu Pegler, on the basis of the robust basidiomes having repeatedly furcate lamellae, with few or absence of lamellulae, presence of skeleto-ligative hyphae, scattered hyphal pegs and uninflated generative hyphae in the hymenium. The morphotaxonomic characters of this species are distinguished from L. badius, L. araucariae and L. brunneofloccosus that are placed in the same section (Pegler 1983a). Subsequently, L. dicholamellatus was reported from different regions of Kerala (Pradeep & Vrinda 2007, Varghese et al. 2010, Mohanan 2011). However, the ITS sequences generated from L. dicholamellatus confirms that this falls morphologically within the highly variable L. sajor-caju (Seelan et al. 2015).
Lentinus giganteus Berk., London J. Bot. 6: 493 [bis] (1847).
Notes: The taxonomic position of L. giganteus, described from Sri Lanka, is controversial. Corner (1981) treated L. giganteus as type of his new subgenus Gigantopanus of Panus, and Pegler (1983a) as type of section Gigantopanus of subgenus Panus. The morphotaxonomic characters of L. giganteus shared with Lentinus are a dimitic hyphal system with a radial construction of the hymenophoral trama, generative hyphae being much narrower than the skeletal hyphae, and spores with an oil guttule. These characters are also similar to Pleurotus in having wider, well seperated lamellae, elongate ellipsoid to ellipsoid rather than cylindric spores and conspicuous lecythiform cheilocystidia at the gill edge. However, in a revision of L. giganteus from Sri Lanka and Thailand, L. giganteus was nested within the Pleurotus clade and consequently the species has been transferred to Pleurotus (Karunarathna et al. 2011b). This species, which has been reported from different regions of the Western Ghats of Kerala (Joseph et al. 1995, Manimohan et al. 2004, Pradeep & Vrinda 2007, Mohanan 2011) is therefore excluded from the key.
Lentinus tuber-regium (Fr.) Fr., Syn. Gen. Lentinus: 3 (1836).
Notes: Corner (1981) placed the nematode destroying species Pleurotus tuber-regium and Pl. levis in Panus, whereas, Pegler (1983a) placed these species in Lentinus. Singer (1975, 1986) and Kühner (1980) treated these species in Pleurotus. However, in a phylogenetic study, the nematotoxic compound producing L. tuber-regium was placed in the Pleurotus clade, supporting a placement in Pleurotus (Thorn et al. 2000). This fungus is therefore excluded from the key, but has been reported from the Western Ghats of Kerala (Geetha et al. 2002) and of Tamil Nadu (Johnsy et al. 2011, Davidson et al. 2012, Kumar & Kaviyarasan 2012). This is a strictly terrestrial species arising from a subterranean sclerotium.
Panus torulosus Fr., Epicr. Syst. Mycol.: 397 (1838).
Notes: Sathe & Kulkarni (1980) reported Panus torulosus from Sampaje in the Western Ghats of Maharashtra. The presence of hyphal pegs in the hymenophore and context consisting of skeleto-ligative hyphae confirms that this represents a species of Lentinus. Further, the large pileus (7–170 mm diam) with a short stipe (22–25 × 12–17 mm), presence of lamellae and lamellulae which are not furcated, and the non-inflated generative hyphae in the hyphal system place this species in of Lentinus sect. Tigrini sensu Pegler(1983a).
Discussion
The xeromorphic lentinoid species, Lentinus and Panus belonging to Polyporaceae, are widely distributed through tropical, subtropical, temperate, and boreal regions (Pegler 1983a, b, c, Corner 1981). The genera are well represented in India, with 20 species of Lentinus and eight of Panus (including P. alpacus and P. natarajanus) distributed in different regions (Bilgrami et al. 1979, 1991, Sathe & Daniel 1980, Sathe & Deshpande 1980, Sathe & Kulkarni 1980, Manjula 1983, Pegler 1983a, Manimohan et al. 2004, Natarajan et al. 2005, Farook et al. 2013, Senthilarasu 2014). Of the 17 species known in the Western Ghats, four species (L. bambusinus, P. indicus, P. alpacus, and P. natarajanus) are new to science. However, the morphotaxonomic characters of L. bambusinus agree with Panus sensu Corner. The lentinoid fungal diversity of the Western Ghats mainly comprises species familiar in south-east Asia as well as in other tropical regions, notably East Africa and the Lesser Antilles are present (Pegler 1977, 1983c, 1986). The most common species widely distributed in almost all the regions are L. crinitus, L. polychrous, L. squarrosulus, L. sajor-caju, L. ciliatus, L. connatus, P. neostrigosus, P. velutinus, and P. similis. However, all the lentinoid species newly described from the Western Ghats appear to be confined to that region. Although this study confirms that the Western Ghats has a high species diversity of lentinoid fungi, the study of their diversity is confined to a part of Western Ghats of Kerala, Maharashtra, Tamil Nadu, and Karnataka. Large areas of the Western Ghats do, however, remain completely unexplored or underexplored for this widely distributed but often misunderstood tropical assemblage of macromycetes.
References
Bilgrami KS, Jamalludin, Rizwi AM (1979) Fungi of India. New Delhi: Today and Tomorrow’s Printers and Publishers.
Bilgrami KS, Jamalludin, Rizwi AM (1991) Fungi of India. New Delhi: Today and Tomorrow’s Printers and Publishers.
Binder M, Hibett DS, Larsson KH, Larsson E, Langer E, et al. (2005) The phylogenetic distribution of resupinate forms across the major clades of mushroom-forming fungi (homobasidiomycetes). Systematics and Biodiversity 3: 113–157.
Binder M, Justo A, Riley R, Salamov A, Lopez-Giraldez F, et al. (2013) Phylogenetic and phylogenomic overview of the Polyporales. Mycologia 105: 1350–1373.
Corner EJH (1981) The agaric genera Lentinus, Panus, and Pleurotus with particular reference to Malaysian species. Nova Hedwigia 69: 1–169.
Davidson S, Johnsy G, Samuel SA, Kaviyarasan V (2012) Mushrooms in the food culture of Kaani tribes of Kanyakumari District. Indian Journal of Traditional Knowledge 11: 150–153.
Drechsler-Santos ER, Wartchow F, Coimbra VRM, Gibertoni TB, Cavalcanti MAQ (2012) Studies on lentinoid fungi (Lentinus and Panus) from the semi-arid region of Brazil. Journal of the Torrey Botanical Society 139: 437–446.
Farook AV, Khan SS, Manimohan P (2013) A checklist of agarics (gilled mushrooms) of Kerala State, India. Mycosphere 4: 97–131.
Florence EJM (2004) Biodiversity Documentation for Kerala. Part 2: Microorganisms (Fungi). [KFRI Handbook no.17.] Kerala: Kerala Forest Research Institute.
Florence EJM, Yesodharan K (2000) Macrofungal Flora of Peechi-Vazhani Wildlife Sanctuary. [KFRI Research Report no. 191.] Kerala: Kerala Forest Research Institute.
Garcia-Sandoval R, Wang Z, Binder M, Hibbett DS (2011) Molecular phylogenetics of the Gloeophyllales and relative ages of clades of Agaricomycotina producing a brown rot. Mycologia 103: 510–524.
Geetha D, Sherin AS, Gokulapalan C, Suhrban M (2002) Pleurotus tuber-regium—a new promising mushroom for the tropics. Mushroom Research 11: 73–76.
Grand EA, Hughes KW, Petersen RH (2011) Relationships within Lentinus subg. Lentinus (Polyporales, Agaricomycetes) with emphasis on sects. Lentinus and Tigrini. Mycological Progress 10: 399–413.
Hibbett DS, Donoghue MJ (2001) Analysis of character correlations among wood decay mechanisms, mating systems, and substrate ranges in homobasidiomycetes. Systematic Biology 50: 215–242.
Hibbett DS, Thorn RG (1994) Nematode-trapping in Pleurotus tuberregium. Mycologia 86: 696–699.
Hibbett DS, Vilgalys R (1991) Evolutionary relationships of Lentinus to the Polyporaceae: evidence from restriction analysis of enzymatically amplified ribosomal DNA. Mycologia 83: 425–439.
Hibbett DS, Vilgalys R (1993) Phylogenetic relationships of Lentinus (Basidiomycotina) inferred from molecular and morphological characters. Systematic Botany 18: 409–433.
Hibbett DS, Murakami S, Tsuneda A (1993) Sporocarp ontogeny in Panus (Basidiomycotina): Evolution and Classification. American Journal of Botany 80: 1336–1348.
Johnsy G, Davidson S, Dinesh MG, Kaviyarasan V (2011) Nutritive value of edible wild mushrooms collected from the Western Ghats of Kanyakumari District. Botany Research International A: 69–74.
Joseph AV, Abraham TK, Vrinda KB, Pradeep CK (1995) New agarics from southern India. Mushroom Research 4: 1–6.
Karunathrana SC, Yang ZL, Zhao RL, Vellinga EC, Bahkali AH, et al. (2011a) Three new species of Lentinus from northern Thailand. Mycological Progress 10: 389–398.
Karunarathna SC, Yang ZL, Raspe O, Ko Ko TW, Vellinga EC, et al. (2011b) Lentinus giganteus revisited: new collections from Sri Lanka and Thailand. Mycotaxon 118: 57–71.
Kornerup A, Wanscher JH (1978) Methuen Handbook of Colour. 3rd edn. London: Methuen.
Küehner R (1980) Les hymenomycetes agaricoides. Bulletin de la Société Linnéenne de Lyon 49: 1–1027.
Kumar M, Kaviyarasan V (2012) Distribution of Lentinus tuberregium (Fr), an indigenous edible medicinal mushroom in Tamil Nadu, South India. Journal of Academia and Industrial Research 1: 296–300.
Kumar TKA, Manimohan P (2005) A new species of Lentinus from India. Mycotaxon 92: 119–123.
Lévillé JH (1846) Description des champignons de I’ herbier de Museum de Paris. Annates des Sciences Naturelles Botanique, ser. 3 5: 111–167. 249–305.
Manimohan P, Leelavathy KM (1995) A new variety of Lentinus caespiticola from southern India. Mycological Research 99: 451–452.
Manimohan P, Divya N, Kumar TKA, Vrinda KB, Pradeep CK (2004) The genus Lentinus in Kerala State. Mycotaxon 90: 311–318.
Manjula B (1983) A revised list of the agaricoid and boletoid basidiomycetes from India and Nepal. Proceedings of the Indian Academy of Sciences, Plant Science 92: 81–213.
Mohanan C (2011) Macrofungi of Kerala. [KFRI Handbook no. 27.] Kerala: Kerala Forest Research Institute.
Moser M (1978) Keys to Agarics and Boleti. London: R. Philips.
Natarajan K (1978) South Indian Agaricales VI. Kavaka 6: 65–70.
Natarajan K, Manjula B (1978) Studies on Lentinus squarrosulus Lev. In: Indian Mushrooms Science J (Atal C, Bhat BK, Kaul TN, eds) 1: 451–456. USA: Indo-American Literature House.
Natarajan K, Raman N (1981) South Indian Agaricales. Nova Hedwigia 34: 163–176.
Natarajan K, Kumaresan V, Narayanan K (2005) A checklist of Indian agarics and boletes (1984–2002). Kavaka 33: 61–128.
Pegler DN (1975) The classification of the genus Lentinus Fr. (Basidiomycota). Kavaka 3: 11–20.
Pegler DN (1977) A preliminary agaric flora of East Africa. Kew Bulletin, Additional Series 6: 1–615.
Pegler DN (1983a) The genus Lentinus: a world monograph. Kew Bulletin, Additional Series 10: 1–281.
Pegler DN (1983b) Lentinus araucariae, an Australasian member of the Lentinus badius-complex. Cryptogamie. Mycologie 4: 123–128.
Pegler DN (1983c) The agaric flora of the Lesser Antilles. Kew Bulletin, Additional Series 9: 1–668.
Pegler DN (1986) Agaric flora of Sri Lanka. Kew Bulletin, Additional Series 12: 1–519.
Pradeep CK, Vrinda KB (2007) Some noteworthy agarics from Western Ghats of Kerala. Journal of Mycopathological Research 1: 1–14.
Redhead SA, Ginns JH (1985) A reappraisal of agaric genera associated with brown rots of wood. Transactions of Mycological Society of Japan 26: 349–381.
Rune F (1994) Neolentinus — a well founded genus in Pleurotaceae that includes Heliocybe. Mycological Research 98: 542–544.
Sathe AV, Daniel J (1980) Agaricales (mushrooms) of Kerala State — Part III. In: Agaricales (Mushrooms) of South West India: 75–108. [MACS Monograph no, 1.] Pune: Maharashtra Association for the Cultivation of Science.
Sathe AV, Deshpande S (1980) Agaricales (mushrooms) of Maharashtra State — Part I. In: Agaricales (Mushrooms) of South West India: 9–42. [MACS Monograph no, 1.] Pune: Maharashtra Association for the Cultivation of Science.
Sathe AV, Kulkarni SM (1980) Agaricales (mushrooms) of Karnataka State — Part II. In: Agaricales (Mushrooms) of South West India: 43–73. [MACS Monograph no, 1.] Pune: Maharashtra Association for the Cultivation of Science.
Seelan SJ, Justo A, Nagy LG, Grand EA, Redhead SA et al. (2015) Phylogenetic relationships and morphological evolution in Lentinus, Polyporellus and Neofavolus, emphasizing southeastern Asian taxa. Mycologia 107: in press.
Senthilarasu G (2014) Diversity of agarics (gilled mushrooms) of Maharashtra, India. Current Research in Environmental & Applied Mycology 4: 58–78.
Senthilarasu G, Singh SK (2012) A new species of Lentinus from India. Mycotaxon 121: 69–74.
Sharma JK, Mohanan C, Florence EJM (1985) Disease survey in nurseries and plantations of forest tree species grown in Kerala. KFRI Research Report 36: 1–268.
Singer R (1975) The Agaricales in Modern Taxonomy. 3rd edn. Vaduz: J. Cramer.
Singer R (1986) The Agaricales in Modern Taxonomy. 4th edn. Kbnigstein: Sven Koeltz Scientific Books.
Theissen F (1911) Fungi aliquot Bombayensis, a revised edition, Blatter Collections, Annates Mycologica 9: 153–159.
Thorn RG, Moncalvo J-M, Reddy CA, Vilgalys R (2000) Phylogenetic analyses and the distribution of nematophagy support a monophyletic Pleurotaceae within the polyphyletic pleurotoid-lentinoid fungi. Mycologia 92: 241–252.
Usha N, Janardhana GR (2014) Diversity of macrofungi in the Western Ghats of Karnataka (India). Indian Forester 140: 531–536.
Varghese SP, Pradeep CK, Vrinda KB (2010) Mushrooms of tribal importance in Wayanad area of Kerala. Journal of Mycopathological Research 48: 311–320.
Acknowledgments
I particularly thank David J. Hibbett for his valuable comments and suggestions on a draft version of this manuscript. Sincere thanks are also extended to David L. Hawksworth and Pedro W. Crous for their critical remarks to improve the quality of the paper. I thank P. Manimohan, V. Kumaresan, and H. Raja for providing valuable literature. Paromita Chakraborty is also appreciated for her kind support. Thanks are due to the Director, SRM Research Institute, SRM University for providing laboratory facilities.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by-nc/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Senthilarasu, G. The lentinoid fungi (Lentinus and Panus) from Western Ghats, India. IMA Fungus 6, 119–128 (2015). https://doi.org/10.5598/imafungus.2015.06.01.06
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.5598/imafungus.2015.06.01.06