
Abstract
Several species of Septoria are associated with leaf and fruit spot of pistachio (Pistacia vera), though their identity has always been confused, making identification problematic. The present study elucidates the taxonomy of the Septoria spp. associated with pistachio, and distinguishes four species associated with this host genus. Partial nucleotide sequence data for five gene loci, ITS, LSU, EF-1α, RPB2 and Btub were generated for a subset of isolates. Cylindroseptoria pistaciae, which is associated with leaf spots of Pistacia lentiscus in Spain, is characterised by pycnidial conidiomata that give rise to cylindrical, aseptate conidia. Two species of Septoria s. str. are also recognised on pistachio, S. pistaciarum, and S. pistaciae. The latter is part of the S. protearum species complex, and appears to be a wide host range pathogen occurring on hosts in several different plant families. Septoria pistacina, a major pathogen of pistachio in Turkey, is shown to belong to Pseudocercospora, and not Septoria as earlier suspected. Other than for its pycnidial conidiomata, it is a typical species of Pseudocercospora based on its smooth, pigmented conidiogenous cells and septate conidia. This phenomenon has also been observed in Pallidocercospora, and seriously questions the value of conidiomatal structure at generic level, which has traditionally been used to separate hyphomycetous from coelomycetous ascomycetes. Other than DNA barcodes to facilitate the molecular identification of these taxa occurring on pistachio, a key is also provided to distinguish species based on morphology.
Similar content being viewed by others

Introduction
The genus Pistacia (Anacardiaceae), which is believed to have originated in Central Asia, consists of at least 11 species (Parfitt & Badenes 1997). Among these, Pistacia vera (pistachio), which is native to Western Asia and parts of the Middle East, is the only cultivated and economically important species (Tous & Ferguson 1996). Several important plant pathogens have been recorded from pistachio, causing fruit and root rot, blossom and shoot blight, canker and rust, and other problems (Michailides et al. 1995, https://doi.org/nt.ars-grin.gov/fungaldatabases/). Of these, Septoria leaf spot is one of the more important diseases associated with fruit and leaf spot.
Desmaziéres (1842) published the first description of a Septoria species causing a leaf spot of Pistacia vera in northern France, for which he introduced the name S. pistaciae. In the same year, Léveillé (1842) described and illustrated Dothidea pistaciae causing a leaf spot of a Pistacia sp. in Crimea. Cooke (1884), upon examination of the type material on which Léveillé based D. pistaciae, transferred it to Septoria. Apparently Cooke was unaware of the existence of Desmaziéres’ name. Allescher (1901) proposed the binominal S. pistacina to replace S. pistaciae (Lév.) Cooke 1884 and to differentiate it from S. pistaciae Desm. 1842. Caracciolo (1934) reported a third species from pistachio as causing a serious leaf spot in Sicily, which he subsequently described as S. pistaciarum. Finally, Chitzanidis (1956) reported sexual morphs for two of the three species, namely Mycosphaerella pistacina (for Septoria pistacina) and Mycosphaerella pistaciarum (for Septoria pistaciarum) (Teviotdale et al. 2001).
Septoria pistaciarum is known from the USA, and the eastern Mediterranean and southeast Anatolian regions (Young & Michailides 1989). Septoria pistaciae is known from the USA (Califonia), Asia (Armenia, Republic of Georgia, India, Israel, Kazakhstan, Kirgizstan, Syria, Tadjikistan, Turkey, Turkmenistan and Uzbekistan), Europe (Albania, France, Greece, Italy, Portugal and Ukraine), and Africa (Egypt) (Pantidou 1973, Dudka et al. 2004, Andrianova & Minter 2004, Haggag et al. 2006). Septoria pistacina is known from Greece (Chitzanidis 1956), Syria, Turkey and Iran (Aghajani et al. 2009), and appears to have a more limited distribution.
The application of these Septoria names to the respective diseases that they are associated with has been plagued by confusion ever since they were first introduced. The aim of the present study is to elucidate the taxonomy of the Septoria species associated with fruit and leaf spot diseases of pistachio, and to place them in a phylogenetic context within Mycosphaerellaceae.
Materials and Methods
Isolates
Isolations were made from leaf spots by placing leaves in damp chambers for 1–2 d to enhance sporulation. Single conidial colonies were established from sporulating conidiomata on Petri dishes containing 2 % malt extract agar (MEA) as described earlier (Crous et al. 1991). Additional strains were obtained from the culture collection of the CBS-KNAW Fungal Biodiversity Centre (CBS), Utrecht, The Netherlands. Colonies were subcultured onto potato-dextrose agar (PDA), oatmeal agar (OA), and MEA (Crous et al. 2009c), and incubated at 25 °C under continuous near-ultraviolet light to promote sporulation. Reference strains were deposited at the CBS (Table 1).
DNA isolation, amplification and analyses
Genomic DNA was extracted from fungal colonies growing on MEA using the UltraCleanTM Microbial DNA Isolation Kit (MoBio Laboratories, Solana Beach, CA, USA) according to the manufacturer’s protocol. The primers LSU1Fd (Crous et al. 2009a) and LR5 (Vilgalys & Hester 1990) were used to amplify the partial 28S rRNA gene (LSU), ITS5 and ITS4 (White et al. 1990) were used to amplify the ITS region T1 (O’Donnell & Cigelnik 1997) and b-Sandy-R (Stukenbrock et al. 2012) were used to amplify the partial β-tubulin locus (Btub), EF1-728F (Carbone & Kohn 1999) and EF-2 (O’Donnell et al. 1998) were used to amplify the partial translation elongation factor-1α locus (EF) and fRPB2-5F (Liu et al. 1999) and fRPB2-414R (Quaedvlieg et al. 2011) were used to amplify the partial RNA polymerase II second largest subunit locus (RPB2). A basic alignment of the obtained sequence data was first done using MAFFT v. 7 [(https://doi.org/mafft.cbrc.jp/alignment/server/index.html) (Katoh et al. 2002)] and if necessary, manually improved in BioEdit v. 7.0.5.2 (Hall 1999). To check the congruency of the RPB2 and LSU datasets, a 70 % neighbour-joining (NJ) reciprocal bootstrap was performed (Mason-Gamer & Kellogg 1996, Lombard et al. 2010). A Bayesian analysis (critical value for the topological convergence diagnostic set to 0.01) was performed on the concatenated LSU/RPB2 loci using MrBayes v. 3.2.1 (Huelsenbeck & Ronquist 2001) as described by Crous et al. (2006) using nucleotide substitution models that were selected using MrModeltest v. 2.3 (Nylander 2004). Sequences derived from this study were lodged at GenBank, and the alignment was deposited in TreeBASE (https://doi.org/www.treebase.org/treebase/index.html).
Morphology
Observations were made with a Zeiss V20 Discovery stereo-microscope, and with a Zeiss Axio Imager 2 light microscope using differential interference contrast (DIC) illumination and an AxioCam MRc5 camera and Zen software. Colony characters and pigment production were noted after 2 wk of growth on MEA, PDA and OA incubated at 25 °C. Colony colours (surface and reverse) were rated according to the colour charts of Rayner (1970). Morphological descriptions were based on cultures sporulating on PDA, and taxonomic novelties and metadata were deposited in MycoBank (https://doi.org/www.MycoBank.org; Crous et al. 2004).
Results
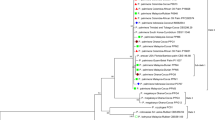
The RPB2 and LSU sequence datasets did not show any conflicts in their tree topology for the 70 % reciprocal bootstrap trees, allowing us to combine them in the multigene analyses. The LSU sequence contained 745 base pairs, of which 99 where unique, the RPB2 sequence contained 317 base pairs, of which 157 where unique. For both datasets, the GTR-I-gamma substitution model (as calculated by MrModeltest) was used during the MrBayes run. During the generation of the tree (Fig. 1), a total of 7 216 trees were generated, and 5 412 (75 %) of them where sampled for the final tree

A Bayesian 50 % majority rule RPB2/LSU consensus tree containing representative isolates belonging to Pseudocercospora and related genera (Mycosphaerellaceae). Bayesian posterior probabilities support values for the respective nodes are displayed in the tree. A stop rule (set to 0.01) for the critical value for the topological convergence diagnostic was used for the Bayesian analysis. The tree was rooted to Zymoseptoria verkleyi (CBS 133618). The scalebar indicates 0.1 expected changes per site.
Taxonomy
Cylindroseptoria pistaciae Quaedvlieg et al., Stud. Mycol. 75: 359 (2013).
(Fig. 2)

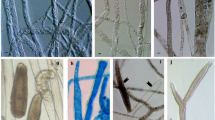
Cylindroseptoria pistaciae (CBS 471.69). A–B. Conidiomata sporulating in culture. C–D. Intercalary chains of chlamydospore-like cells. E–F. Conidiogenous cells. G–H. Conidia. Bars = 10 µm, H applies to C and D.
Description: Conidiomata pycnidial, erumpent, globose, black, separate, with a black crusty outer layer of cells, to 200 µm diam, with a central ostiole; wall of 3–6 layers of brown textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells phialidic (mostly monophialidic, but a few observed to also be polyphialidic), lining the inner cavity, hyaline, smooth, ampulliform, 5–8 × 3–4 µm, proliferating percurrently (inconspicuous) or with periclinal thickening at apex (also occurring as solitary loci on superficial hyphae surrounding pycnidia). Conidia hyaline, smooth, cylindrical, mostly straight, rarely slightly curved, apex subobtuse, base truncate, guttulate, aseptate, (9−)11−13(−18) × 2.5−3(−3.5) µm (from Quaedvlieg et al. 2013)
Culture characteristics: Colonies on PDA flat, circular, lacking aerial mycelium, surface fuscous-black, reverse olivaceousblack, after 14 d at 24 °C, 3.5 cm diam; on MEA surface fuscous-black, reverse olivaceous-black, after 14 d, 4.5 cm diam; on OA similar to PDA.
Type: Spain: Mallorca: El Arenal, on leaves of Pistacia lentiscus, 25 May 1969, H. A. van der Aa (CBS H-21301 — holotype; culture ex-type CBS 471.69).
Notes: Quaedvlieg et al. (2013) established the genus Cylindroseptoria for taxa having cupulate to pycnidial conidiomata, and phialidic conidiogenous cells with periclinal thickening, that give rise to cylindrical, aseptate conidia. Although Cylindroseptoria pistaciae was introduced as a novel species (from symptomatic leaves of Pistacia lentiscus), no information is available about its potential role as plant pathogen.
Pseudocercospora pistacina (Allesch.) Crous, Quaedvlieg & Sarpkaya, comb. nov.
Basionym: Septoria pistacina Allesch., Rabenh. Krypt.-Fl. 1(6): 830 (1900) [“1899”]
Synonyms: Dothidea pistaciae Lév., in Démidoff, Voy. Russ. Mér. 2: 108 (1842).
Non Pseudocercospora pistaciae (Chupp) Crous & U. Braun, Mycotaxon 78: 338 (2001).
Septoria pistaciae (Lév.) Cooke, Grevillea 13 (66): 45 (1884); nom. illegit., non S. pistaciae Desm. 1842.
Mycosphaerella pistacina Chitzan., Ann. Inst. Phytopath. Benaki 10: 42 (1956).
MycoBank MB805893
(Fig. 3)

Pseudocercospora pistacina (CBS 135840). A. Fruit tree orchard. B–C. Leaf spots. D. Disease symptoms on fruit. E. Conidia cirrhi oozing from immersed pycnidial conidiomata. F–G. Vertical section through pycnidia. H. Conidiogenous cells giving rise to conidia. I. Conidia. Bars: F = 300 µm, G = 150 µm, H−l = 10 µm.
Description: Leaf spots numerous, brown, amphigenous, angular, confined by leaf veins, to 30 mm long, 3–6 mm diam, containing numerous small, aggregated, immersed conidiomata. Fruit spots grey to pale brown, 1–4 mm diam, coalescing to form larger spots, surrounded by a distinct, reddish margin. Conidiomata subepidermal, globose to depressed, to 300 µm diam with a wide central ostiole, to 100 µm diam; wall 10–20 µm thick, of 3–6 layers of brown textura angularis. Conidiophores subcylindrical, pale brown, smooth, 0–3-septate, branched or not, 10–30 × 3–5 µm. Conidiogenous cells terminal and sublateral, pale brown, smooth, subcylindrical to doliiform, 6–15 × 2.5–4 µm; proliferating several times percurrently at the apex. Conidia pale brown, smooth, guttulate, subcylindrical, curved, medianly 1-septate, constricted at the septum, apex obtuse, tapering at the base to a truncate hilum, 1.5–2 µm diam, (32−) 35−42(−50) × (3−)3.5−4(−5) µm.
Chitzanidis (1956) reports ascomata as 90−110 × 80−110 µm, asci as 44.5−54.5 × 13−14.5 µm, and ascospores as 26−40 × 3−5 µm.
Culture characteristics: Colonies after 2 wk at 24 °C reaching 10 mm diam, erumpent with sparse aerial mycelium and even, lobate margins; on OA, MEA and PDA dirty white, remaining sterile; in reverse iron-grey.
Specimens examined: Turkey: Manisa: Selendi, on Pistachio vera, 2010, K. Sarpkaya (CPC 45sln005 = CBS 135840). Gaziantep: Nizip, on P. vera, 2010, K. Sarpkaya (CPC 27NZ080 = CPC 23117). Sanliurfa: Birecik, on P. vera, 2010, K. Sarpkaya (CPC 63br043 = CBS 135841). Aydin: Merkez, on P. vera, 2010, K. Sarpkaya (CPC 09mrk010 = CPC 23118); collection site unknown, on P. vera, 2010, K. Sarpkaya (CPC 21874).
Notes: Because of the pycnidial conidiomata and pigmented conidia, Pseudocercospora pistacina can be confused with Phaeophloeospora or Kirramyces (syn. Teratosphaeria; Crous et al. 2009a, b), though it is phylogenetically unrelated to these genera. Pseudocercospora pistacina clusters basally within Pseudocercospora, but based on the genes studied here, could not be recognised as a separate genus. The genus Pseudocercospora was recently circumscribed as having species with conidiophores that are solitary, fasciculate, synnematal, or arranged in sporodochia, giving rise to conidia that are pigmented, have unthickened or slightly thickened and darkened scars (Crous et al. 2013a). By including Septoria pistacina in Pseudocercospora, we are expanding the generic circumscription of the latter to also include taxa with well-defined pycnidial conidiomata (on host and in culture). Conidiomatal structure has to date been paramount in identifying taxa with enclosed conidiomata (Sutton 1980, Nag Raj 1993), and thus P. pistacina is rather atypical within Pseudocercospora s. str.
Septoria pistaciae Desm., Annls Sci. Nat., Bot., sér. 2 17: 112 (1842).
Synonyms: Phloeospora pistaciae (Desm.) Petr., Annls mycol. 20: 18 (1922).
Cylindrosporium pistaciae (Desm.) Vassil., Fungi Imp. Paras. 2: 510 (1950).
(Fig. 4)

Septoria pistaciae (PC 0142143). A. Herbarium specimen. B–F. Conidiogenous cells giving rise to conidia. G. Conidia. Bars = 10 µm, D applies to E–G.
Description: Leaf spots initially small, orbicular or oblong, scattered, brown to dark brown, 1–2 mm diam, becoming irregular, 5–10 mm, covering large portions of leaf, becoming greyish brown with distinct, narrow brown margin. Conidiomata pycnidial, amphigenous, separate or densely aggregated in the centre of leaf spots, immersed, becoming erumpent, brown to dark brown, globose to pyriform, (40−) 70−90(−120) µm diam, with central ostiole, 15–20 µm diam; wall of 3–4 layers of brown textura angularis. Conidiophores reduced to conidiogenous cells, or up to 4-septate, subcylindrical with lateral and terminal conidiogenous cells, 5−25 × 3−4 µm. Conidiogenous cells hyaline, smooth, ampulliform to subcylindrical, 5−10 × 3−4 µm, lining inner layer of conidiomatal cavity, proliferating sympodially, rarely percurrently. Conidia hyaline, smooth, 0–3-septate, (9−)13−22(−25) × (1.5−)2(−3) µm, obclavate to narrowly subcylindrical, apex subobtuse, base obconically truncate with flattened scar. Spermatial state occurring in conidiomata along with conidia. Spermatogenous cells hyaline, smooth, ampulliform, 4−6 × 3−5 µm. Spermatia hyaline, smooth, ellipsoid to subcylindrical, with obtuse ends, 2.5−3.5 × 1.5 µm.
Culture characteristics: Colonies after 2 wk at 25 °C reaching 40 mm diam on OA; surface sienna, smooth, with even margins, lacking aerial mycelium; culture sterile on OA, PDA, MEA and on barley leaves placed on synthetic nutrient-poor agar (Crous et al. 2009c).
Specimens examined: France: on leaves of Pistacia vera, 1842, Desmazière [Pl. Crypt. Nord Fr., fasc. 24, no. 1181] (PC0142144 — holotype; authentic specimen from general herbarium (PC) no. 1181, PC0142143). — Italy: on leaves of P. vera, June 1951, G. Goidánich (CBS 420.51; culture sterile).
Notes: Septoria pistaciae is part of the species complex for which Verkley et al. (2013) adopted the oldest name, S. protearum, which has an ex-type culture. Isolates in this complex could not be robustly distinguished based on a seven-gene phylogeny, and represent collections with a range of hosts covering six different plant families. Whether this is one plurivorous taxon that can undergo host jumping (Crous & Groenewald 2005), or several closely related taxa that cannot be distinguished based on the set of genes employed by Verkley et al. (2013), awaits further study and inoculation trials.
Andrianova & Minter (2004) described conidia of S. pistaciae as 1–3-septate, (20−)22−25(−34) × 1.5(−2) µm, obclavate to narrowly subcylindrical (based on type material, LE 42353). Our measurements from type material are considerably smaller, namely 0–3-septate, (9−)13−22(−25) × (1.5−)2(−3) µm (PC 0142144). Type material of S. protearum has conidia that are (0−)1−3(−4)-septate, (6−)12−22(-30) × 1.5−2 µm, obclavate to narrowly subcylindrical (Swart et al. 1998, Crous et al. 2013b). If these two taxa are eventually shown to be synonymous, the name S. pistaciae (1842) predates that of S. protearum (1998), but even that may not be the oldest epithet for this taxon. The single isolate available to us for study (CBS 420.51) proved to be sterile, so its morphology could not be confirmed.
Septoria pistaciarum Caracc., Boll. Stud. Inform. R. Giard Colon Palermo 13: 10 [extr.] (1934).
Synonym: Mycosphaerella pistaciarum Chitzan., Ann. Inst. Phytopath. Benaki 10: 42 (1956).
(Fig. 5)

Septoria pistaciarum (CBS 135838). A. Colony sporulating on SNA with sterile barley leaves. B. Colony on OA. C–E. Conidiogenous cells giving rise to conidia. F–G. Conidia. Bars: A–B = 200 µm, C, F–G = 10 µm, C applies to D and E.
Description: Leaf spots angular, brown, amphigenous, 1–2 mm diam, coalescing to become larger leaf spots, confined by leaf veins. Conidiomata pycnidial, erumpent, brown, globose, to 200 µm diam, with central ostiole, exuding a crystalline cirrhus of conidia; wall of 3–6 layers of brown textura angularis. Conidiophores reduced to conidiogenous cells, or one supporting cell which can be branched at the base. Conidiogenous cells lining the inner cavity, hyaline, smooth, subcylindrical to ampulliform or doliiform, 5−15 × 2.5−4 µm, proliferating percurrently near apex, or sympodially. Conidia solitary, hyaline, smooth, guttulate, straight to curved, narrowly obclavate to subcylindrical, (1–)3-septate, apex subobtuse, base obconically truncate, 2 µm diam with minute marginal frill, (45−)55−65(−75) × (2.5−)3(−3.5) µm.
Chitzanidis (1956) reports ascomata as 95−130 × 85−120 µm, asci 47.5−60.5 × 8−12 µm, and ascospores 18−30 × 3–5 µm.
Culture characteristics: Colonies after 2 wk at 24 °C reaching 30 mm diam. Colonies erumpent, folded with feathery, lobate margins. On PDA surface olivaceous grey with patches of pale olivaceous grey and dirty white, reverse olivaceous grey. On OA surface greyish sepia with patches of dirty white, and an umber pigment diffusing into agar. On MEA surface pale olivaceous grey with patches of dirty white; olivaceous grey in reverse.
Specimens examined: Turkey: Hatay: Merkez, on Pistacia terebinthus, 2012, K. Sarpkaya (CPC 002B = CPC 23115). Sakarya: Geyve, on P. vera, 2012, K. Sarpkaya (CPC 003C = CPC 23114). Kutahya: Emet, on P. vera, 2012, K. Sarpkaya (CPC 001A = CBS 135839). Manisa: Selendi, on P. vera, 2012, K. Sarpkaya (CPC 45sln034 = CBS 135838); Demirci, on P. vera, 2012, K. Sarpkaya (CPC 5DMR032 = CPC 23116).
Notes: Septoria pistaciarum is morphologically distinct from the other species occurring on pistachio, in having much larger conidia (45−75 × 2.5−3.5 µm). In the field it can also be disnguished on diseased host plants in causing more angular leafspots, confined by leafveins.
Discussion
The aim of the present study was to clarify which species of Septoria occur on pistachio, and to place them in a phylogenetic context within Mycosphaerellaceae, as recently circumscribed (Quaedvlieg et al. 2013, Verkley et al. 2013). From results obtained, it is clear that up to four septoria-like taxa occur on pistachio, of which two belong to other genera, namely Cylindroseptoria pistaciae and Pseudocercospora pistacina. The remaining two species represent true species of Septoria, namely S. pistaciae and S. pistaciarum. Because of discrepancies in previously published literature, much confusion arose regarding how to distinguish these taxa. In the present study we have been able to compile a key to facilitate identification of these taxa (see below). Sexual morphs have also been described for two of these taxa, namely Septoria pistacina (i.e. Mycosphaerella pistacina) and Septoria pistaciarum (i.e. Mycosphaerella pistaciarum) (Chitzanidis 1956, Teviotdale et al. 2001). However, because the genus Mycosphaerella is restricted to Ramularia (Verkley et al. 2004, Crous et al. 2009a, b, Koike et al. 2011), in moving towards a single nomenclature for fungi (see Hawksworth et al. 2011, Wingfield et al. 2012), the use of Mycosphaerella should be avoided for the mycosphaerella-like sexual morphs linked to Septoria.
The placement of Septoria pistacina in Pseudocercospora is somewhat controvertial, as it has a typical pycnidial conidioma, rather than superficial fascicles or synnemata encountered in Pseudocercospora. Phylogenetically, however, there is no support for recognising S. pistacina as a separate genus based on it being a “pigmented Septoria”. Morphologically, the conidiogenous cells and conidia fit the circumscription of Pseudocercospora, but the conidiomatal anatomy does not. Although species of Capnodiales are known to have synasexual morphs with closed and open conidiomata (Crous et al. 2007, 2009a, b), this is the first example of a taxon with a pycnidial conidioma that clusters among species with fasciculate conidiomata. In addition to S. pistacina, we are also aware of a second as yet undescribed species of “pigmented Septoria” (Crous et al., unpubl.), which again clusters in Pallidocercospora (Crous et al. 2013a). Another example of a genus reported to have acervuli, but observed to have superficial conidiomatal fascicles, is Ciferiella, which has also been reduced to synonymy with Pseudocercospora (Quaedvlieg et al. 2013). These findings support the view that conidiomatal morphology in Pseudocercospora is a continuum from sporulating superficially (fascicles, synnemata, sporodochia), via acervuli, to sporulation in an enclosed structure (pycnidia).
In spite of being morphologically distinct, that two of the reported septoria-like taxa on pistachio represent different genera is rather suprising. Although the pathological relevance of Pseudocercospora pistacina (as S. pistacina), S. pistaciae, and S. pistaciarum on Pistacia vera is well-documented (Michailides 2005), nothing is known about that of Cylindroseptoria pistaciae, other than it was associated with leaf spots of Pistacia lentiscus in Spain. Furthermore, it also appears that the importance of certain diseases of pistachio differs between regions or countries. Approximately 85 % of the world’s pistachio production presently comes from Iran, the USA, and Turkey (https://doi.org/faostat.fao.org/site/339/default.aspx). However, pistachio is irrigated in Iran and the USA, where more rounded fruit bearing cultivars are grown. In contrast, irrigation is not practiced in Turkey, and more elongated fruit cultivars are commonly grown there. These differences in cultivation practices also lead to differences in phytopatological problems. In the USA, the main pathogens are reported to be Botryosphaeria dothidea, Botrytis cinerea, and Alternaria alternata. In Turkey, however, Pseudocercospora pistacina is the most common disease of pistachio (Dinç 1983, Michailides et al. 1995), leading to reports of 3–100 % yield loss in epidemic years by this pathogen (Dinç et al. 1979).
Key to septoria-like species occurring on pistachio
-
1
Conidiaaseptate,9−18 × 2.5−3.5 µm .................................................................... Cylindroseptoria pistaciae
Conidia septate .............................................................................................................................................. 2
-
2 (1)
Conidia pale brown, medianly 1-septate, 32−50 × 3−5 µm ............................... Pseudocercospora pistacina
Conidia hyaline, 1–3-septate ......................................................................................................................... 3
-
3 (2)
Conidia 9−34 × 1.5−3 µm .................................................................................................. Septoria pistaciae
Conidia 45−75 × 2.5−3.5 µm ........................................................................................ Septoria pistaciarum
References
Aghajani MA, Aghapour B, Michailides TJ (2009) First report of Septoria leaf spot of pistachio in Iran. Australasian Plant Disease Notes 4: 29–31.
Allescher A (1901) Fungi Imperfecti: Hyalin-sporige Sphaerioideen. Dr L. Rabenhorst’s Kryptogamen-Flora von Deutschland, Oestereich und der Schweiz 1(6): 961–1016.
Andrianova TV, Minter DW (2004) Septoria pistaciae. IMI Descrpitions of Fungi and Bacteria 1586: 1–2.
Bremer H, Karel G, B0iy0ikoğlu K, Göksel N, Petrak F (1952) Beiträge zur Kenntnis der parasitischen Pilze der Türkei. VI. Revue de la Faculté des Sciences de l’Université d’Istanbul, Série B, 17: 260–261.
Bremer H (1954) Bahçe Kültürleri Hastal0iklar0i. Türkiye Fitopatolojisi 295: 109–110.
Caracciolo F (1934) Una grave septoriosi del pistacchio. Bolle Di Studi ed Informazione del R. Giardino di Palermo 13: 66–73.
Carbone I, Kohn LM (1999) A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 91: 553–556.
Chitzanidis A (1956) Species of Septoria on the leaves of Pistacia vera and their perfect states. Annales de l’Institut Phytopathologique Benaki 10: 29–44.
Cooke MC (1884) Synopsis Pyrenomycetum (continued). Grevillea 13: 41–45.
Crous PW, Braun U, Groenewald JZ (2007) Mycosphaerella is polyphyletic. Studies in Mycology 58: 1–32.
Crous PW, Braun U, Hunter GC, Wingfield MJ, Verkley GJM, Shin H-D, Nakashima C, Groenewald JZ (2013a) Phylogenetic lineages in Pseudocercospora. Studies in Mycology 75: 37–114.
Crous PW, Denman S, Taylor JE, Swart L, Bezuidenhout CM, Hoffman L, Palm ME, Groenewald JZ (2013b) Cultivation and diseases of Proteaceae: Leucadendron, Leucospermum and Protea. 2nd edn. [CBS Biodiversity Series no. 13.] Utrecht: CBSKNAW Fungal Biodiversity Centre.
Crous PW, Gams W, Stalpers JA, Robert V, Stegehuis G (2004) MycoBank: an online initiative to launch mycology into the 21st century. Studies in Mycology 50: 19–22.
Crous PW, Groenewald JZ (2005) Hosts, species and genotypes: opinions versus data. Australasian Plant Pathology 34: 463–470.
Crous PW, Schoch CL, Hyde KD, Wood AR, Gueidan C, et al. (2009a) Phylogenetic lineages in the Capnodiales. Studies in Mycology 64: 17–47.
Crous PW, Slippers B, Wingfield MJ, Rheeder J, Marasas WFO, et al. (2006) Phylogenetic lineages in the Botryosphaeriaceae. Studies in Mycology 55: 235–253.
Crous PW, Summerell BA, Carnegie AJ, Wingfield MJ, Hunter GC, et al. (2009b) Unravelling Mycosphaerella: do you believe in genera? Persoonia 23: 99–118.
Crous PW, Verkley GJM, Groenewald JZ, Samson RA (eds) (2009c) Fungal Biodiversity. [CBS Laboratory Manual Series no.1.]. Utrecht: CBS-KNAW Fungal Biodiversity Centre.
Crous PW, Wingfield MJ, Park RF (1991) Mycosphaerella nubilosa a synonym of M. molleriana. Mycological Research 95: 628–632.
Desmaziéres JB (1842) Neuviéme notice sur quelques plantes cryptogames. Annales des Sciences Naturelles, Botanique, série 2 17: 91–118.
Dinç N (1983) Antepf0ist0iğ0i Hastal0iklar0i ve Mücadele Usulleri. Mesleki Eserler Serisi 2: 19–34.
Dinç N, Göksedef O, Turan K (1979) Gaziantep ili antepf0ist0iklar0inda zarar yapan Karazenk hastal0iğ0i (Septoria pistacina All.) n0in Bio-ekolojisi ve korunma olanaklar0i üzerine araşt0irmalar. Bitki Koruma Bülteni 19: 16–38.
Dudka IO, Heluta VP, Tykhonenko YY, Andrianova TV, Hayova VP, et al. (2004) Fungi of the Crimean Peninsula. Kiev: M.G. Kholodny Institute of Botany, National, Academy of Sciences of Ukraine.
Haggag WM, Abou Rayya MSM, Kasim NE (2006) First report of Septoria pistaciae causing leaf spot of pistachio in Egypt. Plant Disease 90: 1553.
Hall TA (1999) BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series 41: 95–98.
Hawksworth DL, Crous PW, Redhead SA, Reynolds DR, Samson RA, et al. (2011) The Amsterdam Declaration on Fungal Nomenclature. IMA Fungus 2: 105–112.
Hoog GS de, Gerrits van den Ende AHG (1998) Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses 41: 183–189.
Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17: 754–755.
Katoh K, Misawa K, Kuma K, Miyata T (2002) MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Research 30: 3059–3066.
Koike SK, Baameur A, Groenewald JZ, Crous PW. (2011). Cercosporoid leaf pathogens from whorled milkweed and spineless safflower in California. IMA Fungus 2: 7–12.
Lombard L, Crous PW, Wingfield BD, Wingfield MJ (2010) Species concepts in Calonectria (Cylindrocladium). Studies in Mycology 66: 1–13.
Leveillé JH (1842) Obsérvations médicales et énumérations des plantes récueillies en Tauride. In: Voyage dans la Russie meridionale et la Crimée (Démidoff A, ed.) 2: 80–135. Paris: Ernes Boudier.
Liu Y, Whelen S, Hall B (1999) Phylogenetic relationships among ascomycetes: evidence from an RNA polymerse II subunit. Molecular Biology and Evolution 16: 1799–1808.
Mason-Gamer RJ, Kellogg EA (1996) Testing for phylogenetic conflict among molecular data sets in the tribe Triticeae (Gramineae). Systematic Biology 45: 524–545.
Michailides TJ (2005) Pest, disease, and physiological disorders management: above ground fungal diseases. In: Pistachio Production Manual (Beede RH, Freeman MW, Haviland DR, Holtz BA, Kallsen CE, eds): 214–232. Davis, CA: Fruit and Nut Research and Information Center, Department of Plant Sciences, University of California Davis.
Michailides TJ, Morgan DP, Doster MA (1995) Diseases of pistachio in California and their significance. Acta Horticulturae 419: 337–343.
Nag Raj TR (1993) Coelomycetous anamorphs with appendagebearing conidia. Waterloo, ON: Mycologue Publications.
Nylander JAA (2004) MrModeltest. Version 2. Uppsala: Distributed by the author. Evolutionary Biology Centre, Uppsala, University.
O’Donnell K, Cigelnik E (1997) Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Molecular Phylogenetics and Evolution 7: 103–116.
O’Donnell K, Kistler HC, Cigelnik E, Ploetz RC (1998) Multiple evolutionary origins of the fungus causing Panama disease of banana: concordant evidence from nuclear and mitochondrial gene genealogies. Proceedings of the National Academy of Sciences, USA 95: 2044–2049.
Pantidou ML (1973) Fungus-Host Index for Greece. Athens: Benaki Phytopathological Institute.
Parfitt DE, Badenes ML (1997) Phylogeny of the genus Pistacia as determined from analysis of the chloroplast genome. Proceedings of the National Academy of Sciences, USA 94: 7987–7992.
Quaedvlieg W, Kema GHJ, Groenewald JZ, Verkley GJM, Seifbarghi S, Razavi M, Gohari AM, Mehrabi R, Crous PW (2011) Zymoseptoria gen. nov.: a new genus to accommodate Septorialike species occurring on graminicolous hosts. Persoonia 26: 57–69.
Quaedvlieg W, Verkley GJM, Shin H-D, Barreto RW, Alfenas AC, et al. (2013) Sizing up Septoria. Studies in Mycology 75: 307–390.
Rayner RW (1970) A Mycological Colour Chart. Kew: Commonwealth Mycological Institute.
Stukenbrock EH, Quaedvlieg W, Javan-Nikhah M, Zala M, Crous PW, McDonald BA (2012) Zymoseptoria ardabilia and Z. pseudotritici, two progenitor species of the septoria tritici leaf blotch fungus Z. tritici (synonym: Mycosphaerella graminicola). Mycologia 104: 1397–1407.
Sutton BC (1980) The Coelomycetes: fungi imperfecti with pycnidia, acervuli, and stromata. Kew: Commonwealth Mycological Institute.
Swart L, Crous PW, Denman S, Palm ME (1998) Fungi occurring on Proteaceae. I. South African Journal of Botany 64: 137–145.
Teviotdale BL, Michailides TJ, MacDonald J (2001) Diseases of Pistachio (Pistacia vera L.). Common Names of Plant Diseases. St Paul, MN: American Phytopathological Society Press.
Tous J, Ferguson L (1996) Mediterranean fruits. In: Progress in New Crops (Janick J ed.): 416–430. Arlington, VA: American Society for Horticultural Science Press.
Vassiljevsky NI, Karakulin BP (1950) Parazitnye Nesovershennye Griby. Vol. 2. Melanconiales. Moscow: Izdatel’stvo, AN SSSR.
Verkley GJM, Crous PW, Groenewald JZ, Braun U, Aptroot A (2004) Mycosphaerella punctiformis revisited: morphology, phylogeny, and epitypification of the type species of the genus Mycosphaerella (Dothideales, Ascomycota). Mycological Research 108: 1271–1282.
Verkley GJM, Quaedvlieg W, Shin HD, Crous PW (2013) A new approach to species delimitation in Septoria. Studies in Mycology 75: 213–305.
Vilgalys R, Hester M (1990) Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. Journal of Bacteriology 172: 4238–4246.
White TJ, Bruns T, Lee J, Taylor SB (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: PCR Protocols: a guide to methods and applications (Innis MA, Gelfand DH, Sninsky JJ, White TJ eds): 315–322. San Diego: Academic Press.
Wingfield MJ, De Beer ZW, Slippers B, Wingfield BD, Groenewald JZ, et al. (2012) One fungus, one name promotes progressive plant pathology. Molecular Plant Pathology 13: 604–613.
Young DJ, Michailides TJ (1989) First report of Septoria leaf spot of pistachio in Arizona. Plant Disease 73: 775.
Acknowledgements
We thank the technical staff, Arien van Iperen (cultures), Marjan Vermaas (photographic plates), and Mieke Starink-Willemse (DNA isolation, amplifcation and sequencing) for their invaluable assistance.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by-nc/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article
Cite this article
Crous, P.W., Quaedvlieg, W., Sarpkaya, K. et al. Septoria-like pathogens causing leaf and fruit spot of pistachio. IMA Fungus 4, 187–199 (2013). https://doi.org/10.5598/imafungus.2013.04.02.04
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.5598/imafungus.2013.04.02.04