
Abstract
This contribution provides a synopsis of the presentations and discussions during the SIG session on cryptic speciation in lichen-forming fungi held during IMC9. In several cases, a re-examination of morphology against the background of molecular phylogenetic evidence revealed, sometimes subtle, morphological and/or chemical characters, supporting the distinction of particular clades at species level. However, there are also examples of cryptic species in which no morphological characters could be identified to distinguish between lineages. Several cases were presented in which distinct lineages are correlated with biogeographical patterns. When and how to name cryptic species was debated, and the use of terms such as “complex” or “aggregate” commended where the taxa formed part of a single lineage.
Similar content being viewed by others


Introduction
The presentations at the session demonstrated that current species recognition in lichen forming-fungi vastly underestimates the true number of species. Based on phylogenetic and population studies, many cases were presented showing that numerous distinct lineages are hidden under a single species name. The issues raised can be grouped under the following headings: naming cryptic species, numbers of cryptic species, recognition of cryptic species, supporting species separations, and phylogeographic correlations. Collectively, these presentations provide a synopsis of the current state of knowledge of cryptic speciation in lichen-forming fungi.
Naming Cryptic Species
The recognition and naming of cryptic species from cryptic lineages was discussed and approaches and options were suggested. Hawksworth (2010) examined different groups as foraminifera, plant-pathogenic fungi, insects, and plants. The main species concepts were reviewed, and a pragmatic concept was proposed, defining a species as “groups of individuals separated by inheritable discontinuities and which it is useful to give a species name to” (Hawksworth 1996, 2010). The term cryptic species was circumscribed as “populations which are phylogenetically distinct and able to reproduce themselves, by sexual means or otherwise, but which are distinguished by molecular or other features that are either not evident macroscopically or generally overlooked” (Hawksworth 2010).
An increasing number of lichen-forming species are used as biomonitors or bioindicators of pollutants, environmental disturbance, or ecological continuity. Consequently there was the issue of how to proceed when cryptic species or lineages are found in taxa used in such studies where identifications need to be made quickly during field assessments — and access to a modern molecular laboratory is impractical. An acceptable way of referring to such groups of species was commended by Hawksworth (2010). The term “complex” or “aggregate” was supported as used when the populations are closely related, i.e. have a recent shared common ancestor. This practice is already familiar to and regularly used by botanists, citizen scientists, and ecologists dealing with complexes in plants, for example the Rubus fruticosus aggr. and the Taraxacum officinale aggr.
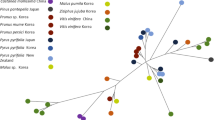
In some situations, however, the option of recognizing subspecies was suggested as perhaps the most appropriate solution, for example in paraphyletic populations (Figs 1 and 2) such as that of Parmelina pastillifera and P. tiliacea s. str. (Núñez-Zapata et al. 2010) (Fig. 1). In contrast, in cases where the cryptic taxa are not closely related but a result of convergence, i.e. they do not either occupy the same clade or have a recent common ancestor, it has to be recognized that the “complex” approach could give a misleading impression of affinity, as in Parmelina cryptotiliacea (Núñez Zapata et al. 2010) or lineages in Parmelia saxatilis (Divakar et al. 2010b).

Phylogenetic tree of Parmelina (Parmeliaceae). Majority rule consensus tree based on 18000 trees from B/MCMC tree sampling procedure from a combined data set of nulTS rDNA and mtLSU rDNA sequences. Posterior probabilities ≥0.95 in the Bayesian analysis are indicated above the branches and MP boostrap values ≥0.75 below branches. Branches with significative support in both analyses are in bold. (AU=Austria, CI=Canary Islands, FR=France, GE=Germany, IN=lndia, IT=ltaly, MO=Morocco, SP=Spain, SV=Slovenia, TK=Turkey, TN=Tunisia, USA=United States of America). Figure provided by Nuñez-Zapata et al.

Parmelina pastillifera (MAF 16473; upper) and P. tiliacea (MAF 16632; lower) both showing isidia, but in P. pastillifera they are peltate while in P. tiliacea are cylindrical. Bars = 5 mm.
Numbers of Cryptic Species
There is a growing body of evidence that the approach to current species recognition in lichenized fungi, which is largely based on morphology and chemistry, vastly underestimates the number of phylogenetic species. Phylogenetic studies repeatedly indicate that numerous distinct lineages can be hidden under a single species name (Arguello et al. 2007, Baloch & Grube 2009, Grube & Kroken 2000, Kroken & Taylor 2001, Molina et al. 2004, Wirtz et al. 2008). In a number of cases, morphological or chemical differences have been interpreted as intraspecific variability. Re-examination of morphology against the background of a molecular phylogeny often reveals, sometimes subtle, and previously overlooked or viewed as unimportant, morphological and/or chemical characters, supporting the distinction of these clades at species level (Arguello et al. 2007, Divakar et al. 2005a, 2005b, Molina et al. 2004, Wirtz et al. 2008). However, there are also cases of cryptic species in which no morphological characters have yet been identified to distinguish distinct lineages. In several cases, distinct lineages are correlated with distinct biogeographical patterns (Arguello et al. 2007, Crespo et al. 2010, Molina et al. 2004, Wirtz et al. 2008). Phylogenetic studies identified distinct lineages that occur in different geographic regions, such as continents.
Recognition of Cryptic Species
The large and increasing number of cryptic lineages detected in fungi means that the recognition of these lineages as separate taxa is a major issue of current fungal taxonomy (Crespo & Pérez-Ortega 2009, Hawksworth 2001). However, cryptic species in lichen-forming fungi may be compared to fungi with other biologies where morphological characters are almost absent, thus the pertinence of using this concept in lichens was discussed (Hawksworth 2010, Pérez-Ortega & Printzen 2010). Unlike many microscopic fungi, some groups of lichens form distinctive macroscopic structures, frequently with a foliose or fruticose form, and with easily observable phenotypical differences. Despite these structures, the plasticity of morphological and chemical characters in these fungi results in a relatively high number of lichens, species or genera, being “difficult” for identification, often accompanied by a frequent lack of generative characters (Divakar et al. 2010b) or the frequency of homoplasy and convergence of characters (Grube & Hawksworth 2007, Muggia 2010, Muggia et al. 2010, Parnmen et al. 2010, Divakar et al. 2010b).
Although only relatively few lichens have yet been identified as comprising cryptic species using molecular data (Grube & Kroken 2000, Kroken & Taylor 2001, Crespo et al. 2002, Feuerer & Thell 2002, Printzen et al. 2003, Molina et al. 2004, Argüello et al. 2007, Wirtz et al. 2008, Fehrer et al. 2009, Divakar et al. 2010a), assemblages of morphologically similar species where identification remains dubious due to variability or ambiguity of key characters used to distinguish those taxa are common. Thus, morphological identification of a lichen-forming species, sometimes even a genus, can be difficult. Therefore, cryptic taxa have been recognised historically in lichens, although not necessarily by that term. “The recognition and characterization of cryptic species is a burgeoning and exciting activity in current systematics, and a major challenge for mycologists of all kinds, not least lichenologists” (Hawksworth 2010). Suggestions for when to formally recognise species within cryptic lineages that are found in molecular studies were discussed (Muggia 2010, Pérez-Ortega & Printzen 2010), and a consensus of the session was to recognise species formally when the phylogeny was unequivocal and other evidence supported their separation, whether ultramicroscopic, “new” morphological, ecological (Muggia 2010) or geographical (Parnmen et al. 2010) were discussed as examples for complementary evidence.
Supporting Species Separations
Recent molecular phylogenies have supported some species separations that were previously based on subtle characters: for example, Parmelina carporrhizans and P. quercina (Argüello et al. 2007, Divakar et al. 2010b), Caloplaca alociza and C. albopruinosa (Muggia 2010). It is also frequently found that distantly related major lineages show a surprising degree of morphological convergence. Examples of this phenomenon can be found within large families such as Parmeliaceae. For example, Parmelina and Austroparmelina were recently separated as independent genera based on geography and phylogeny. However, all species of Austroparmelina were previously included in concept of the genus Parmelina (Crespo et al. 2010, Divakar et al. 2010b). Also there are examples in microlichens, as in Capnodiales where the morphologically similar genera Racodium and Cystocoleus belong to independent lineages in recent phylogenetic studies (Muggia et al. 2008, Muggia 2010).
Phylogeographic Correlations
A number of lichen-forming species were historically thought to have wide distributions, including cosmopolitan and pantropical species. However, while that may be so for some species, molecular analyses have repeatedly demonstrated that many lineages can be hidden under a similar morphology. Several examples were discussed in the symposium (Divakar 2010, Muggia 2010, Parnmen et al. 2010). Divakar et al. (2010) also found a correlation between reproductive modes and distribution patterns. In fertile species, cryptic lineages were frequently found, and geographically disjunct populations were discovered to represent different lineages (Divakar et al. 2010a). Several examples of this type were presented, including Melanelixia glabra and Parmelina quercina, two species distributed in areas with winter rain (Mediterranean climate) in North Africa, Europe and North America (Argüello et al. 2007, Divakar et al. 2010a, b). In sorediate taxa, cryptic lineages have also been found, but in this case the lineages can include specimens from different geographical regions; examples include Flavoparmelia caperata, Parmotrema reticulatum, and P. tinctorum (Divakar et al. 2005, 2010).
References
Argüello A, del Prado R, Cubas P, Crespo A (2007) Parmelia quercina (Parmeliaceae, Lecanorales) includes four phylogenetically supported morphospecies. Biological Journal of the Linnean Society 91: 455–467.
Baloch E, Grube M (2009) Pronounced genetic diversity in tropical epiphyllous lichen fungi. Molecular Ecology 18: 2185–2197.
Crespo A, Molina MC, Blanco O, Schroeter B, Sancho LG, Hawksworth DL (2002) rDNA ITS and ß-tubulin gene sequence analyses reveal two monophyletic groups within the cosmopolitan lichen Parmelia saxatilis. Mycological Research 106: 788–795.
Crespo A, Pérez-Ortega S (2009) Cryptic species and species pairs in lichens: A discussion on the relationship between molecular phylogenies and morphological characters. Anales del Jardin Botànico de Madrid 66 (S1): 71–81.
Crespo A, Ferencova Z, Pérez-Ortega S, Elix JA, Divakar PK (2010) Austroparmelina, a new Australasian lineage in parmelioid lichens (Parmeliaceae, Ascomycota). Systematics and Biodiversity 8: 209–221.
Divakar PK, Molina MC, Lumbsch HT, Crespo A (2005a) Parmelia barrenoae, a new lichen species related to Parmelia sulcata (Parmeliaceae) based on molecular and morphological data. Lichenologist 37: 37–46.
Divakar PK, Blanco O, Hawksworth DL, Crespo A (2005b) Molecular phylogenetic studies on the Parmotrema reticulatum (syn. Rimelia reticulata) complex, including the confirmation of P. pseudoreticulatum as a distinct species. Lichenologist 37: 55–65.
Divakar PK, Figueras G, Hladun N, Crespo A (2010a) Morphological versus phylogenetic species: An example from Melanelixia glabra (Parmeliaceae, Ascomycota). Fungal Diversity 42: 47–55.
Divakar PK, Cubas P, Blanco O, Del-Prado R, Nühez-Zapata J, Roca-Valiente B, Lumbsch HT, Crespo A (2010b) An overview on hidden diversity in lichens: Parmeliaceae. <https://doi.org/www.imc9.info/prog_sig3_detail_divakar.htm>
Fehrer J, Slavikové-Bayerové S, Orange A (2008) Large genetic divergence of new, morphologically similar species of sterile lichens from Europe (Lepraria, Stereocaulaceae, Ascomycota): concordance of DNA sequence data with secondary metabolites. Cladistics 24: 443–458.
Feuerer T, Thell A (2002) Parmelia ernstiae — a new macrolichen from Germany. Mitteilungen aus dem Institut für Allgemeine Botanik in Hamburg 30–32: 49–60.
Grube M, Hawksworth DL (2007) Trouble with liehen: the re-evaluation and re-interpretation of thallus form and fruit body types in the molecular era. Mycological Research 111: 1116–1132.
Grube M, Kraken S (2000) Molecular approaches and the concept ofspecies and species complexes in lichenized fungi. Mycological Research 104: 1284–1294.
Hawksworth DL (1991) The fungal dimension of biodiversity: magnitude, significance, and conservation. Mycological Research 95: 641–655.
Hawksworth DL (1996) Microbial collections as a tool in biodiversity and biosystematic research. In Culture Collections to Improve the Quality of Life (Samson RA, Stalpers JA, Mei D van der, Stouthamer AH, eds): 26–35. Baarn: Centraalbureau voorSchimmelcultures.
Hawksworth DL (2001) The magnitude of fungal diversity: the 1.5 million species estimate revisited. Mycological Research 105: 1422–1432.
Hawksworth DL (2010) Cryptic speciation: how common is it and how should it be handled taxonomically? <https://doi.org/www.imc9.info/prog_sig3_detail_hawksworth.htm>
Kraken S, Taylor JW (2001) A gene genealogical approach to recognize phylogentic species boundaries in the lichenizedfungus Letharia. Mycologia 93: 38–53.
Molina MC, Crespo A, Blanco O, Lumbsch HT, Hawksworth DL (2004) Phylogenetic relationships and species concepts in Parmelia s.str. (Parmeliaceae) inferred from nuclear ITS rDNA and ß-tubulin sequences. Lichenologist 36: 37–54.
Muggia L (2010) The Implications of Morphological and PhylogeneticSpecies Concepts in Lichens. <https://doi.org/www.imc9.info/prog_sig3_detail_lucia.htm>
Muggia L, Hafellner J, Wirtz N, Hawksworth DL, Grube M (2008) The sterile microfilamentous lichenized fungi Cystocoleus ebeneus and Racodium rupestre are relatives of plant pathogens and clinically important dothidealean fungi. Mycological Research 112: 51–57.
Nühez-Zapata J, Divakar P, Del-Prado R, Cubas P, Crespo A (2010) Cryptic species in the lichen forming fungus Parmelina tiliacea (Ascomycota). <https://doi.org/www.imc9.info/prog_sig3_detail_nunez.htm>
Parnmen S, Rangsiruji A, Mongkolsuk P, Boonpragob K, Lumbsch HT (2010) Species Delimitations and Evolutionary History of the Cladia aggregata aggregate (Lecanorales, Ascomycota). <https://doi.org/www.imc9.info/prog_sig3_detail_parnmen.htm>
Pérez-Ortega S, Printzen C (2010) Species delimitation in lichens: using morphology and molecular markers to find species boundaries. <https://doi.org/www.imc9.info/prog_sig3_detail_ortega.htm>
Vinuesa M, Sanches-Puelles JM, Tibell L (2001) Intraspecificvariation in Mycocalicium subtile (Mycocaliciaceae) elucidated by morphology and the sequences of the ITS1-5.8S-ITS2 regionof rDNA. Mycological Research 105: 323–330.
Wirtz N, Printzen C, Lumbsch HT (2008) The delimitation of Antarctic and bipolar species of neuropogonoid Usnea (Ascomycota, Lecanorales): a cohesion approach of species recognition for the Usnea perpusilla complex. Mycological Research 112: 472–484.
Acknowledgments
We acknowledge the participation of David L. Hawksworth (University Complutense of Madrid, Spain) in finalizing this article; his revision enriched the manuscript with important suggestions improving the text. This work was supported by the Spanish Ministerio de Ciencia e lnnovación (CGL 2010-21646/BOS, CGL2007-64652/BOS) and a grant of the National Science Foundation to H.T.L. (“Hidden diversity in parmelioid lichens”, DEB-0949147).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by-nc/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article
Cite this article
Crespo, A., Lumbsch, H.T. Cryptic species in lichen-forming fungi. IMA Fungus 1, 167–170 (2010). https://doi.org/10.5598/imafungus.2010.01.02.09
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.5598/imafungus.2010.01.02.09