
Abstract
Studies of exogenous covert orienting use peripheral cues (stimuli) that are spatially uninformative about the locations of subsequent targets. When the time course of the cue’s influence on performance is explored (by varying the cue target onset asynchrony; CTOA), a biphasic pattern is usually seen with better performance at the cued location when the CTOA is short (typically attributed to attentional capture) and worse performance at the cued location when the CTOA is long (attributed to inhibition of return). However, while spatially uninformative, these cues (even when a nonaging foreperiod is used) entail a temporal contingency with the subsequent target. Consequently, this so-called capture may reflect an unintended consequence of endogenous allocation of temporal attention. Following Lawrence and Klein (Journal of Experimental Psychology: General, 142(2), 560–572, 2013) we used Rescorla’s (Psychological Review, 74, 71–80, 1967) truly random control condition to ensure that the spatially uninformative peripheral stimuli were temporally completely uninformative. Even such completely uninformative peripheral stimuli generated the prototypical biphasic pattern.
Similar content being viewed by others
In their taxonomy of attention, Klein and Lawrence (2011, 2012) distinguished between the allocation of information processing resources between and within different domains (space, time, and task) and the modes of control of this allocation (endogenous and exogenous). Although we believe that Posner (1980) was the first psychologist to use the terms endogenous and exogenous, the distinction between volitional versus reflexive control that they represent has a long history (LaBerge, 1990). In the domain of space, the distinction between endogenous versus exogenous control of orienting has received considerable study since this distinction was highlighted by Posner (1980). Early behavioral (e.g., Briand & Klein, 1987; Jonides, 1981; Klein, 1994; Müller & Rabbitt, 1989) and neuroscientific (e.g., Chica, Bartolomeo & Lupiáñez, 2013; Hopfinger & West, 2006) studies established the reality and importance of the distinction that has been validated in more recent work (for a review, see Klein, 2009). As noted by Klein and Lawrence (2011), the distinction also applies in the domain of time, but it has taken longer for this to be studied (for recent efforts, see Lawrence & Klein, 2013; Rohenkohl, Coull, & Nobre, 2011). Although the present experiment is concerned with exogenously controlled spatial attention, its rationale is that to elicit truly exogenous orienting, the spatial cues must be truly uninformative: not just spatially uninformative but also temporally uninformative.
A well-established paradigm for studying exogenously controlled spatial attention is Posner’s spatial cuing paradigm (Posner & Cohen, 1984). In this paradigm, a spatially uninformative cue is usually presented in one of two or more peripheral locations where, subsequently, a target stimulus will be presented. Spatially uninformative means that regardless of the location that had been cued, the target will be presented equally often in all of the possible target locations. By convention, when the target appears in the same location as the prior cue, this is called a valid trial, and when the cue and target appear in the different locations, this is called an invalid trial. When the interval between the onsets of the cue and target (cue–target onset asynchrony, or CTOA) is varied, the prototypical (but not universal) pattern of results, first discovered by Posner and Cohen (1984), is early facilitation at the cued location followed by inhibition at the cued location. This biphasic pattern is illustrated in Fig. 1. The early facilitation is typically attributed to capture of attention by the cue. On the assumption that attention is normally disengaged from the cued location (the uninformativeness of the cue makes keeping it there suboptimal), the later inhibition, which was called “inhibition of return” (IOR) by Posner, Rafal, Choate, & Vaughan, 1985), is typically attributed to a reluctance of attention to return to a recently visited location.

Time course of cuing effects from three different reviews of the literature. Dashed line (saccadic responses) is from Klein’s (2000) review of inhibition of return. Solid line (simple manual detection responses) is from Samuel and Kat’s (2003) graphic meta-analysis. Dotted line (nonspatial discrimination responses) is derived from Redden et al. (2016)
In this description, we have carefully qualified “uninformative cues” by the word spatially. Importantly, in the vast majority of studies of exogenous orienting, this qualifier has not been used: Indeed, in the typical methods section, the reader would be told: “Participants were instructed ignore the cues that were uninformative about the upcoming target.” We assert that until the present experiment, participants have always had incentive to pay attention to the peripheral cues because they were temporally informative. A few researchers have been alert to this possibility, and their efforts to make their spatially uninformative cues temporally uninformative will be covered in the Discussion.
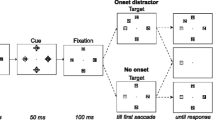
To the best of our knowledge, all experiments on exogenous spatial orienting have utilized a trial structure characterized by a fixation period, followed by a cue, or the absence of a cue, followed by a target (or the absence of a target—a catch trial). It is our belief that regardless of the stimulus-onset asynchrony (SOAs) used, the distribution of SOAs, or the frequency of catch trials, this trial structure generates an incentive to attend to the cue because it signals that a target is likely to follow (at some point) before the next trial begins. Following in the footsteps of Lawrence and Klein (2013), in the present study we used a method borrowed from the animal learning literature, Rescorla’s (1967) “truly random control procedure,” to make the spatially uninformative peripheral cues also temporally uninformative and thus to eliminate this incentive. As applied to studies of human performance, this procedure does away with the typical trial structure of an experiment because the two events (cuesFootnote 1 and targets) are presented completely randomly and independently from each other, as illustrated in Fig. 2. Importantly, in this procedure there is no contingency between cues and targets, and the phenomenological experience is one of two completely independent sequences of events.

Schematic illustration of the relative timing of targets (row of filled circles above the timeline) and cues (checkerboard circles below the timeline) by independently sampling from an exponential distribution
Possible outcomes
The effects (early facilitation and later IOR) of spatially uninformative peripheral cues upon performance in response to validly and invalidly cued targets have been shown to be sensitive to featural (Folk, Remington, & Johnston, 1992) and spatial (Hilchey, Klein, & Ivanoff, 2012; Wang & Klein, 2012) attentional control settings. The simple questions we are asking here is whether, when spatially uninformative cues are also made temporally uninformative, will they still generate early facilitation and IOR, or will either or both of these effects be eliminated?
Method
Participants
Forty participants (ages 18 to 23 years, 36 right-handed, 34 female), undergraduate students who received extra credit for their participation in the study, contributed the results that are described below. A sample size of 20 participants was initially targeted on the basis of our informal assessment that the phenomena of interest have relatively large effect sizes, with published sample sizes typically ranging from 6 to 24. Preliminary analysis after collection of 20 participants was consistent with those reported below in terms of the point estimates of effects (which in turn were also consistent with the literature), but we found the uncertainty thereon unacceptably large (e.g., 95% confidence intervals spanning up to 80 ms) and thus doubled data collection to a total of 40.Footnote 2
Apparatus and stimuli
The experiment was implemented on an iMac computer (2.8 GHz) with a 27-inch screen. Stimulus presentation and data collection were carried out by a program written in Python by MAL. Participants used a gaming controller to respond. The experiment room was normally lighted.
There were two possible target stimuli, either a full black or full white circle with a diameter of 2° presented in a gray background. Targets were centered 10° to the left or right of the center of the screen and presented for 100 milliseconds. There was one type of peripheral cue, consisting of a checkered black and white ring with a diameter of 5°. When presented (for a duration of 50 ms), a cue was centered 10° to the left or right of the center of the screen. The timing of both targets and cues was randomly generated by independent sampling from a shifted and truncated exponential function with a minimum of 3 s, mean of 4 s, and maximum of 10 s. Cues and targets were randomly assigned to the left or right locations.
Procedure
After reading and signing the informed consent letter, participants were seated at a distance of approximately 60 cm from the computer monitor. They were also instructed to ignore the cues and to respond to the color of the targets irrespective of their location by pushing the right and left triggers, for the white and black targets, respectively. Participants were instructed keep their gaze focused on the central fixation stimuli and to respond as rapidly and as accurately as possible. Following every response, a number appeared at the center of the screen that indicated participant’s reaction time. To provide feedback on their responses, the color of this number was consistent with participant’s response (white for the right trigger and black for the left). The experiment began with a short block of practice and was followed by two experimental blocks. Each experimental block was composed of eight subblocks, each of which included of 29 targets.
Design and methods of analysis
Because the experimental procedure entailed random generation of the times of cues and targets, a variety of rulesFootnote 3 were applied to each target to ensure that we could unambiguously attribute responses to unambiguously cued targets. The application of these rules excluded 46% of the targets and yielded a distribution of SOAs, including some negative ones for which a cue was presented after a target but prior to the participant’s response to that target. We then excluded SOAs below −200 ms and above 1,200 ms, as the distribution of SOAs is approximately uniform from −200 ms to 1,200 ms, but begins to taper off thereafter (i.e., SOAs become increasingly less frequent as they increase beyond about 1,200 ms), and prior literature indicates we should expect to observe the phenomena of interest in this SOA range. This trimming removed 37% of the remaining cue–target–response triads. Additional exclusions included responses for which more than one trigger was pressed (including presses below response threshold; 3.3%), responses for which RTs were faster than 200 ms (0.2% of trials), or slower than 1,000 ms (0.9% of trials). After all the exclusions described above, a total 7,429 responses contributed to the analyses described below.
Following Lawrence and Klein (2013), analysis of performance variables (reaction time and accuracy) was achieved by a generalized additive model with subjects contributing a random effect on the intercept.Footnote 4 Error data were modeled as binomial and transformed to the percentage and, consistent with the results shown in Fig. 3, a significant interaction between the two factors, F(7, 243) = 2.26, p < .01, scale for reporting purposes. Only accurate trials were included in the analysis of RTs, and RTs were log-transformed prior to analysis, then transformed back to the millisecond scale for reporting purposes.

Upper panel: Reaction time as a function of the spatial relation (valid/invalid) and SOA between cues and targets. Lower panel: Cuing effects as a function of SOA. Dashed circles represent findings from the literature as described in the Discussion. In both panels, ribbons reflect 95% confidence intervals
Results
As illustrated in Fig. 3, reaction times revealed cue-generated early facilitation followed by a later inhibitory effect. The time of peak facilitation occurred around 127 ms (25–198 ms) with a magnitude of 18 ms (11–26 ms), and the time of peak IOR occurred around 620 ms (496–751 ms) with a magnitude of 18 ms (10–25 ms). Accuracies did not vary substantially as a function of SOA. The error rate in the Invalid condition was 6.8% (6.3%–7.4%), and in the valid condition, it was 6.0% (5.5%–6.5%). The overall cueing effect on error rate was 0.8% (−0.2%–1.9%), and the 95% CI included zero.
Although it was not a focus, the design of our experiment permits us to explore whether completely uninformative peripheral cues affect the magnitude of the Simon effect (Simon, 1990). The Simon effect refers to the relatively ubiquitous finding (e.g., Klein, Dove, Ivanoff, & Eskes, 2006; Urcuioli, Vu, & Proctor, 2005) that responses are faster and tend to be more accurate when there is spatial congruence between the target and response locations than when they do not correspond. As illustrated in a Supplement to this article, a robust Simon effect was obtained. In reaction time, this was relatively constant over the −200 to 1,200 ms time course, whereas in accuracy the effect decreased as a function of increasing foreperiod. Importantly, the biphasic pattern of cuing effects in RT that is illustrated in Fig. 3 was equally manifest for Simon congruent and Simon incongruent trials.
Discussion
In the present study, we employed Rescorla’s truly random procedure to eliminate any temporal contingencies between the spatially uninformative peripheral cues and the targets.Footnote 5 Our results replicated the prototypical biphasic pattern of early facilitation followed by later IOR. That we obtained this pattern even though our cues were both spatially and temporally uninformative strongly suggests that neither phase of this biphasic pattern has been generated in previous studies because the spatially uninformative peripheral cues were, in these studies, temporally informative.
To provide a firm basis for comparison with the literature, we focused on studies that were previously reviewed by Redden, Hilchey, and Klein (2016). These studies, like ours, had used a discrimination task and reported both RT and accuracy for each condition. Redden et al. were specifically interested in IOR, and hence only cue–target SOAs of 700 ms or longer. Some of the studies they reviewed only tested at one long CTOA. For present purposes we focused on studies from Redden et al. in which a range of SOAs was used (both short and long) and for which, in the resulting data base, SOAs were represented by at least three independent observations. Cuing effects from these studies (which are listed in the Appendix) are plotted in Fig. 3, bottom panel, as dashed circles (and in Fig. 1, as the dotted line).
One can see here that there is quite good agreement between the biphasic pattern we have obtained using Rescorla’s method for removing temporal contingencies between cues and targets and the pattern from studies that used multiple foreperiods in mixed block designs. If there are any differences worth noting here (and, because the cues and targets used in the present study are relatively unique, the differences might not be noteworthy), they are that in the present study, IOR may have an earlier onset, and, at the longest, SOA may not have as great a magnitude. An earlier onset in our experiment could be easily predicted by Klein’s (2000) suggestion that when there is incentive for participants to pay attention to the cue, it may take longer for attention to disengage from it, thereby delaying the appearance of IOR (see also, Milliken, Lupiáñez, Roberts, & Stevanovski, 2003; Wang, Satel, Hilchey, & Klein, 2012).
There have been several previous attempts to explore the interaction of temporal expectancies and the time course of exogenous spatial cueing effects. Milliken et al. (2003), for example, manipulated whether the three foreperiods (100, 500, and 900 ms) that were mixed in a block were equiprobable or whether there was a strong bias toward the early or late foreperiod. This manipulation had little effect on the time course of cuing effects when they used a detection task, which showed robust IOR at the longer SOAs regardless of the bias condition. When they used a discrimination task, there was greater facilitation and greater IOR when the 100 ms CTOA was likely than in the other two conditions. Importantly, all the bias conditions showed a numerical biphasic pattern with early facilitation and later IOR.
Tipper and Kingstone (2005) conducted a study whose rational was very similar to ours. They note, that in a typical exogenous cuing experiment: “While it is true that the cue does not predict where a target will appear, the cue is highly predictive as to when a target will appear. In other words, although there is no reason to attend to the cue based on its unreliable spatial information, there is good reason to attend to the cue because of its reliable temporal information” (p. B56). They found that increasing the probability of catch trials decreased, but did not eliminate, IOR. We do not dispute this finding, but their conclusion that this effect is about temporal attention is far from established. Their manipulation of catch trial probability was based on their belief that when the probability of catch trials was high (25%) “participants [would choose] to ignore a cue’s temporal information” p. B60. But, there is no evidence from their study that would allow any inferences about the degree to which their participants were or were not using the temporal information their cues provided. A simpler interpretation is that the magnitude of IOR is positively related to the probability of the response (see Ivanoff & Klein, 2004, where such an effect is established).
Gabay and Henik (2008, 2010) used a procedure (called nonaging foreperiodsFootnote 6) in which expectancy effects are supposed to be eliminated, because the momentary probability of the presentation of a target at each CTOA remains constant as each possible CTOA elapses without presentation of the target. When the task was simple detection (Gabay & Henik, 2008), the nonaging condition greatly reduced the overall foreperiod effect (relative to the standard, aging, condition, and one with accelerated aging), but the biphasic pattern was robust regardless how the distribution of foreperiods was manipulated. In striking contrast, when the task was discrimination (Gabay & Henik, 2010), the biphasic pattern was not obtained in the nonaging foreperiod condition; rather, facilitation was present for all the CTOAs. This finding remains anomalous. It suggests that the cue captured attention and then, despite the fact that the cue was spatially uninformative, the participants failed to disengage attention from it. Perhaps, as time elapsed during a trial, participants continued to attend the cue while trying to understand how the probabilities were changing. Regardless how one interprets the absence of IOR in this experiment with its nonspatial discrimination task, the dramatic difference in the pattern of results when compared to the same authors’ 2008 experiment strongly implies, in agreement with Milliken et al. (2003), that the type of task (detection versus discrimination) matters for the time course of cuing effects in a paradigm with a cue-target trial structure. In this context, we believe that repeating the present application of Rescorla’s truly random control condition, which does not have a cue–target trial structure, while changing the task to be performed with the target to simple detection and saccadic localization (see Fig. 1) would be very interesting.
As noted by Lawrence and Klein (2013, p. 561), in any experiment that uses a trial structure (each trial consists of a cue followed by target or catch trial), regardless of the distribution of foreperiods, “the signal always precedes the target, thereby manifesting a temporal contingency.” This contingency is not overcome by the use of a nonaging foreperiod or variation in the probability of catch trials. But it is overcome through the use of Rescorla’s “truly random control” condition as we have done here. Therefore, we believe that what we have observed here (see Fig. 3) is the true time course of exogenous cuing when the peripheral cues are truly uninformative—that is, both spatially and temporally uninformative.
Notes
The normal meaning of the word cue makes its use here somewhat misleading because cues should be informative about something, and our method seeks to make them entirely uninformative. That noted, for present purposes, it is important to preserve the connection to the literature on exogenous orienting, which, till now, has relied upon spatially uninformative cues. This footnote reminds the reader that we believe we have made our cues completely uninformative.
In a research program seeking to apply null-hypothesis testing to achieve one-off decisions with regard to the presence/absence of an effect, a flexible stopping-rule would induce inflation of the Type I error rate. Although our decision to double the N from 20 to 40 to reduce the 95% CI is not such a flexible stopping rule, it might increase the Type I error rate. That noted, we are not proposing any such one-off decisions, but instead seek to contribute to the cumulative evidence of the scientific process. Those seeking such decisions may consider the current report exploratory rather than confirmatory.
First, a target was excluded if there was no response between the onset of this target and the onset of the next target. We then found the nearest response to a target and excluded the target if this response occurred before the target or was closer to a different target. We then found the nearest cue to the target in question. We excluded the target if this cue was presented after the nearest response or if this cue was closer to a different target.
We also conducted a more traditional analysis of variance on LogRT. First we binned the target responses into 200 ms bins centered on SOAs of −200, 0, 200, 400, 600, 800, 1,000, and 1,200 ms. In this analysis SOA and cue–target spatial relation were the factors. Analysis revealed a significant main effect for SOA, F(7, 243) = 4.850, p = .001 and, consistent with the results shown in Figure 3, a significant interaction between the two factors, F(7, 243) = 2.26, p < 0.01.
An anonymous reviewer pointed out that in learning paradigms using Rescorla’s truly random-control-condition animals may develop a weak association between the rewards (USs) and the conditioned stimuli (CSs) (e.g., Rescorla, 2000). Whereas this does not challenge our use of this paradigm to eliminate the temporal informativeness of the cues, it does raise the possibility that in our experiment, participants might begin to associate cues with targets.
When foreperiods are equiprobably mixed in a block, as each foreperiod elapses without presentation of the target, the probability of the target being presented at each foreperiod increases. This is true even when there are catch trials. Imagine an experiment in which there are three equiprobable (25%) foreperiods and 25% catch trials. At the start of the trial each foreperiod has a 25% chance of target presentation. After the shortest foreperiod elapses without presentation of a target, the remaining foreperiods have a 33% chance of target presentation; and when the middle foreperiod elapses, the longest one has a 50% chance of target presentation. This growth in the momentary probability of the target has been referred to as “aging foreperiods.” Nickerson and Burnham (1969) proposed a method, nonaging foreperiods, to overcome this change in momentary probability. With nonaging foreperiods, at the beginning of a trial the early foreperiods are more probable and probability decreases exponentially. In the previous example, there would be 12.5% catch trials, 12.5% longest foreperiod; 25% middle foreperiod; 50% shortest foreperiod. With this method, the momentary probability of a target being presented at any foreperiod, given it has not yet been presented yet, is 50%.
References
Briand, K., & Klein, R. M. (1987). Is Posner's beam the same as Treisman's glue?: on the relationship between visual orienting and feature integration theory. Journal of Experimental Psychology: Human Perception & Performance, 13(2), 228–247.
Chica, A. B., Bartolomeo, P., & Lupiáñez, J. (2013). Two cognitive and neural systems for endogenous and exogenous spatial attention. Behavioural Brain Research, 237, 107–123. https://doi.org/10.1016/j.bbr.2012.09.027
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance, 18, 1030–1044.
Gabay, S., & Henik, A. (2008). The effects of expectancy on inhibition of return. Cognition, 106(3), 1478–1486.
Gabay, S., & Henik, A. (2010). Temporal expectancy modulates inhibition of return in a discrimination task. Psychonomic Bulletin & Review, 17(1), 47–51.
Hilchey, M. D., Klein, R. M., & Ivanoff, J. (2012) Perceptual and motor IOR: Components or flavours? Attention, Perception, & Psychophysics, 74(7), 1416–1429. https://doi.org/10.3758/s13414-012-0332-x
Hopfinger, J. B., & West, V. M. (2006). Interactions between endogenous and exogenous attention on cortical visual processing. NeuroImage, 31(2), 774–789. https://doi.org/10.1016/j.neuroimage.2005.12.049
Ivanoff, J., & Klein, R. M. (2004). Stimulus-response probability and inhibition of return. Psychonomic Bulletin & Review, 11(3), 542–550.
Jonides, J. (1981). Voluntary versus automatic control over the mind’s eye movements. In A. D. Long (Ed.), Attention and performance IX ( pp. 187–203). Hillsdale, NJ: Erlbaum.
Klein, R. M. (1994). Perceptual-motor expectancies interact with covert visual orienting under endogenous but not exogenous control. Canadian Journal of Experimental Psychology, 48, 151–166.
Klein, R. M. (2000). Inhibition of return. Trends in Cognitive Sciences, 4(4), 138–147.
Klein, R. M. (2009). On the control of attention. Canadian Journal of Experimental Psychology, 63, 240–252. https://doi.org/10.1037/a0015807
Klein, R. M., Dove, M., Ivanoff, J., & Eskes, G. A. (2006). Parametric exploration of the Simon effect across visual space. Canadian Journal of Experimental Psychology/Revue canadienne de psychologie expérimentale, 60(2), 112.
Klein, R. M., & Lawrence, M. A. (2011). On the modes and domains of attention. In M. I. Posner (Ed.), Cognitive neuroscience of attention (2nd ed., pp. 11–28). New York, NY: Guilford Press.
Klein, R. M. & Lawrence, M. A. (2012). Pursuing a productive taxonomy of attention. Presented at a symposium: on the future of attention research and practice: defining, measuring and repairing the networks of attention: an international symposium (Halifax, Canada)
LaBerge, D. L. (1990) Attention. Psychological Science, 1(3), 158–162.
Lawrence, M. A., & Klein, R. M. (2013). Isolating exogenous and endogenous mechanisms of temporal attention. Journal of Experimental Psychology: General, 142(2), 560–572. https://doi.org/10.1037/a0029023
Milliken, B., Lupiáñez, J., Roberts, M., & Stevanovski, B. (2003). Orienting in space and time: Joint contributions to exogenous spatial cuing effects. Psychonomic Bulletin & Review, 10(4), 877–883.
Müller, H. J., & Rabbitt, P. M. (1989). Reflexive and voluntary orienting of visual attention: time course of activation and resistance to interruption. Journal of Experimental psychology: Human perception and performance, 15(2), 315–330.
Nickerson, R., & Burnham, D. (1969). Response times with nonaging foreperiods. Journal of Experimental Psychology, 79, 452–457.
Posner, M. I. (1980). Orienting of attention. Quarterly Journal of Experimental Psychology 32, 3–25.
Posner, M. I., & Cohen, Y. (1984). Components of visual orienting. Attention and Performance X, 32, 531–556.
Posner, M. I., Rafal, R. D., Choate, L. S., & Vaughan, J. (1985). Inhibition of return: Neural basis and function. Cognitive Neuropsychology, 2, 211–228.
Redden, R. S., Hilchey, M. D., & Klein, R. M. (2016) Peripheral stimuli generate different forms of inhibition of return when participants make prosaccades versus antisaccades to them. Attention, Perception, & Psychophysics, 78(8), 2283–2291.
Rescorla, R. A. (1967). Pavlovian conditioning and its proper control procedures. Psychological Review, 74, 71–80.
Rescorla, R. A. (2000). Associative changes with a random CS–US relationship. The Quarterly Journal of Experimental Psychology Section B, 53(4b), 325–340.
Rohenkohl, G., Coull, J. T., & Nobre, A. C. (2011). Behavioural dissociation between exogenous and endogenous temporal orienting of attention. PLoS ONE, 6(1), 1–5. https://doi.org/10.1371/journal.pone.0014620
Samuel, A. G., & Kat, D. (2003). Inhibition of return: A graphical meta-analysis of its time course, and an empirical test of its temporal and spatial properties. Psychonomic Bulletin & Review, 10, 897–906.
Simon, J. R. (1990). The effects of an irrelevant directional cue on human information processing. In R. W. Proctor & T. G. Reeve (Eds.), Stimulus-response compatibility: An integrated perspective (pp. 31–86). Amsterdam, The Netherlands: North-Holland.
Tipper, C., & Kingstone, A. (2005). Is inhibition of return a reflexive effect?. Cognition, 97(3), B55–B62.
Urcuioli, P. J., Vu, K. P. L., & Proctor, R. W. (2005). A Simon effect in pigeons. Journal of Experimental Psychology: General, 134(1), 93–107.
Wang, Z., & Klein, R. M. (2012) Focal spatial attention eliminates inhibition of return. Psychonomic Bulletin & Review, 19, 462–469.
Wang, Z., Satel, J., Hilchey, M. D., & Klein, R. M. (2012). Averaging saccades are repelled by prior uninformative cues at both short and long intervals. Visual Cognition, 20(7), 825–847.
Author note
The research described here was supported by an NSERC Discovery Grant to Raymond Klein. The instructions, stimuli, program, data files, and analysis scripts associated with the research presented here are posted at https://osf.io/cz48s.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
ESM 1
(PDF 204 kb)
Appendix: Reference list for data from the literature plotted in Figure 3
Appendix: Reference list for data from the literature plotted in Figure 3
López-Ramón, M. F., Chica, A. B., Bartolomeo, P., & Lupiáñez, J. (2011). Attentional orienting and awareness: Evidence from a discrimination task. Consciousness and Cognition, 20(3), 745–755.
Lupiáñez, J., Milán, E. G., Tornay, F. J., Madrid, E., & Tudela, P. (1997). Does IOR occur in discrimination tasks? Yes, it does, but later. Attention, Perception, & Psychophysics, 59(8), 1241–1254.
Lupiáñez, J., & Milliken, B. (1999). Inhibition of return and the attentional set for integrating versus differentiating information. The Journal of General Psychology, 126(4), 392–418.
Lupiáñez, J., Milliken, B., Solano, C., Weaver, B., & Tipper, S. P. (2001). On the strategic modulation of the time course of facilitation and inhibition of return. The Quarterly Journal of Experimental Psychology: Section A, 54(3), 753–773.
Lupiáñez, J., Weaver, B., Tipper, S. P., & Madrid, E. (2001). The effects of practice on cueing in detection and discrimination tasks. Psicológica, 22(1).
Mathôt, S., Dalmaijer, E., Grainger, J., & Van der Stigchel, S. (2014). The pupillary light response reflects exogenous attention and inhibition of return. Journal of Vision, 14(14), 7–7.
Sanabria, D., Morales, E., Luque, A., Gálvez, G., Huertas, F., & Lupiañez, J. (2011). Effects of acute aerobic exercise on exogenous spatial attention. Psychology of Sport and Exercise, 12(5), 570–574.
Rights and permissions
About this article

Cite this article
Habibnezhad, M., Lawrence, M.A. & Klein, R.M. Using Rescorla’s truly random control condition to measure truly exogenous covert orienting. Psychon Bull Rev 26, 569–575 (2019). https://doi.org/10.3758/s13423-018-1544-4
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-018-1544-4