
Abstract
The degree to which humans have top-down control over which information they process remains a central debate within the attention literature. Most of the evidence supporting the top-down control of visuospatial attention has come from cueing paradigms in which target stimuli are preceded by cues that are similar or dissimilar from the target. These studies find that the cues similar to targets capture attention, but dissimilar cues do not, suggesting the top-down control of attention. Here, we used a modified cueing paradigm to investigate an alternative possibility that the cue type differences are due to sequential dependency effects occurring between cue and target processing rather than the top-down control of attention. When individuals searched for color targets, we replicated contingent capture effects in RTs, which are susceptible to sequential dependencies, but memory performance was always best at the cued locations, regardless of the cue’s identity. When individuals searched for onset targets, we observed contingent capture in both tasks. These results demonstrate the utility of the memory probe paradigm and suggest an asymmetry between how strongly onsets and color defined cues capture attention.
Similar content being viewed by others

There is a shift in cognitive science to view the mind as a prediction engine rather than something that builds up experience piece-by-piece through sensory experience (Clark, 2013; 2015; Friston & Kiebel, 2009). Using vision as an example, rather than a bottom-up approach to building a visual world, the visual system is hypothesized to predict what the world is like and then detect prediction errors in order to adjust its model. Such predictive processing leads to an empirical prediction for visual cognition paradigms; a given instance of stimulus processing will be strongly influenced by what stimulus was recently processed. This empirical prediction has support from a range of paradigms. For example, in the object preview paradigm (Kahneman, Treisman, & Gibbs, 1992), there is a cost for responding to a placeholder with a given letter inside of it if the placeholder had previously been perceived as containing a different letter (see also Hommel, 1998). Moreover, in spatial cueing, a mismatch between the cue’s color/shape and the target’s color/shape can eliminate cue location validity effects (Huffman, Al-Aidroos, & Pratt, 2017). Indeed, merely reading a color word can cause items of that color to capture attention, even if the word is a 100 % invalid cue of target color (Ansorge & Becker, 2012). Each of these effects can be attributed to a mechanism in the visual system that predicts a stable visual world (an assumption supported by most real-life experience) such that when the prediction is incorrect there is a processing cost. In the current study, we examine whether sequential dependences can account for performance differences between goal consistent and inconsistent cues in a spatial cueing paradigm.
The paradigm in question is the well-known contingent capture paradigm. In contingent capture studies, a spatial cueing paradigm is modified to include cues that are consistent or inconsistent with a visual target’s defining characteristic (Folk, Remington, & Johnston, 1992). The paradigm is meant to study whether top-down control over attentional capture exists. That is, given that an individual is looking for targets defined by color, can they form an attentional control set (ACS) such that color defined cues capture attention while onset defined cues do not? The answer is overwhelming yes; ACS compatible cues typically generate larger cue validity effects than incompatible cues (Born, Kerzel, & Pratt, 2015; Folk, Remington, & Johnston, 1992; Lien, Ruthruff, Goodin, & Remington, 2008)
For our current purposes, it is important to point out a necessary feature of contingent capture paradigms. Namely, in the ACS compatible cue condition it is generally the case that there is non-spatial feature overlap between cues and targets, but in the ACS incompatible condition this typically not the case. This leaves open that possibility that attentional capture effects may exist in the ACS incompatible condition but might be obscured by sequential dependence related costs.Footnote 1 By sequential dependence related cost, we refer to costs related to the cue’s identity on target processingFootnote 2 (such as cost for processing a white target following a red cue). To test this, we looked to replicate the contingent capture effect in response time (RT; which is susceptible to sequential dependence effects) while also intermixing a secondary task (a memory probe) that should be resilient to sequential dependence effects. If the contingent capture effect is related to a true ability to ignore goal irrelevant information, then it should be present in both measures.
To test for sequential dependency effects , we modified the contingent capture paradigm to include memory probe trials. This addition is based on the recent work of Gaspelin, Leonard, and Luck (2015; see also Kim & Cave, 1995), who adapted the additional singleton search task (Theeuwes, 1992) to include a memory probe component on a subset of trials. Specifically, on 70 % of trials the participants completed a visual search for a shape singleton and reported the location of a dot within the shape singleton. In the other 30 % of trials, they briefly presented letters in each of the search items. Furthermore, on one half of the trials there was an additional color singleton. The question at hand was – what would memory performance be like at the additional singleton’s location? Consistent with the additional singleton capturing attention, the data showed increased memory at that location relative to neutral distactors. Further work indicated that this was true when the task could be completed using singleton search mode, but not when feature search mode was required, in which case memory was supressed at the additional singleton location despite the distractor causing no changes in RT.
The purposes of the present study, the critical point from Gaspelin et al. (2015), is that the allocation of attention was reflected by fluctuations in memory, and the memory performance fluctuations were informative regarding changes in processing in a condition where no RT differences were observed. In other words, in the absence of sequential dependency effects, attentional allocation could be infered from memory performance. Given that, for the current study, sequential dependency effects should affect RTs due to the close relationship between cues and targets within a trial, but not the visual working memory for stimuli that are highly different from the preceding cue stimuli and require a different response type that was unspeeded, factors that make sequential dependency effects less likely (see Hommel, 1998; Huffman & Pratt, 2017, Experiment 1). Thus, if sequential dependency effects are masking attentional capture by goal-inconsistent cues, we predict that we will replicate the contingent capture effect in RTs, but that memory performance will be better at cued locations regardless of the cue’s goal consistency. If goal-inconsistent cues do not capture attention, however, memory performance will not be affected by the goal-inconsistent cue’s location.
Experiment 1
To examine the role of sequential dependencies in contingent capture we adapted the typical contingent capture cueing procedure to include memory probe trials. If the lack of cue validity effects in these paradigms is related to individuals’ attention not being captured by goal incompatible cues, then contingent capture will be evident in the memory data. If both cues capture attention equally, however, the cue validity effect in the memory probe task will be unaffected by cue type.
Method
Participants
Eighteen University of Toronto undergraduate students participated in exchange for course credit. All students reported normal or corrected-to-normal vision, were naïve to the study’s purpose, and provided informed consent.
Stimuli and apparatus
Participants completed the experiment using a PC connected a CRT monitor (screen resolution: 1,024 × 768; refresh rate: 85 Hz). We used Matlab by Mathworks with the Psychophysics toolbox (Kleiner, Brainard, Pelli, Ingling, Murray, & Broussard, 2007) for stimulus presentation. We presented stimuli on a black background. Stimuli consisted of a gray (CIE: luminance=53.6 cd/m2, x=.003, y=-.006) circular (0.4° visual angle) fixation point, two gray square placeholders (4° visual angle), one white (CIE: luminance=85.2 cd/m2, x=.005, y=.010) onset cue or two four-dot cues (0.4° visual angle per dot) with one dot appearing on each of the placeholder’s four sides, 0.4° visual angle red (CIE: luminance=17.4 cd/m2, x=.590, y=.324) or green (CIE: luminance=17.1 cd/m2, x=.296, y=.545) target dots, and white capital letters from the entire English alphabet draw in 35 font size. In the search condition, participants responded with their left hands using the ‘z’ and ‘x’ keys on a standard QWERTY keyboard. In the memory probe condition, participants responded with their right hands using the left and right buttons on a computer mouse.
Procedure
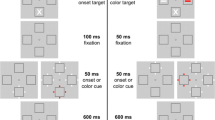
See Fig. 1 for an example of the experimental procedure. Every trial began with the central fixation dot and two placeholders placed to either side of fixation (5° visual angle, center-to-center). Following 1,000 ms, the cue display appeared. In the onset condition, four white dots appeared around one of the placeholders. In the color cue condition, four dots appeared around both placeholders, but one of the four dot cues was red or green. These remained for 100 ms before offsetting. After 50 ms, the search display appeared. The search display consisted of a dot appearing on the left or right side of each placeholder (the target’s location within a placeholder was independent of which side of the screen in appeared on). One of the dots was white and one was red or green. The red or green dot was designated the target. In the search condition, we asked participants to report whether the target was on the placeholder’s left or right side as quickly as they could without sacrificing accuracy. In the memory probe condition, a letter appeared in each placeholder (letters were never repeated) at the same time as the search display would have and remained for 150 ms. The letters were followed with a mask composed of five randomly chosen letters appearing in a random location in a circular area with a diameter of 1° centered around the memory display letters’ locations. None of the masking letters could match the letters that appeared within the memory display and the mask remained for 300 ms. Next, a letter appeared in the center of the display. The letter was equally likely be one of the two letters that appeared previously or a letter that was absent from the memory probe display. Participants reported whether the letter was present or absent from the memory display.

Stimulus presentation for the search and probe tasks. In the search task, participants looked for the red or green dot and reported its location. In the probe task, they attempted to remember the two letters. They then had to report whether the centrally presented letter was present in the memory display
Design
All combinations of factors cue type (onset or color), cue location, target location (left or right placeholder), target side (left or right side of the placeholder), target color, and trial type [search (two-thirds of trials) or memory probe (one-third of trials)] were counterbalanced, repeated eight times each, and presented in random order. This resulted in 512 search trials and 256 memory probe trials. Within the search trials, we determined cue color randomly. Within the memory probe trials, we randomly determined which letters appeared in each placeholder (with the constraint that the same two letters could not appear) and whether the cued location letter, uncued location letter, or absent letter was the probe letter.
Results
Search condition
We trimmed trials where errors were made or RTs were below 100 ms as anticipatory responses or 2.5 SDs above the grand mean as attentional lapses (9.1 % of trials). We analyzed the search condition trials using a 2 (cue type: onset or color) × (cue spatial validity: cued or uncued), repeated measures ANOVA with RTs as the dependent variable (Fig. 2a). Cue type did not affect RTs, F(1,17)<1, but RTs were faster when the cue validly indicated the target’s location (M±SE=577±23 ms) than when it did not (607±25 ms), F(1,17)=50.002, p<.001, \( {\eta}_p^2 \)=.746. Importantly, replicating the contingent capture effect, the cue validity was larger in the color condition (41±7 ms) than in the onset condition (25±3 ms), resulting in a significant interaction, F(1,17)=12.707, p =.002, \( {\eta}_p^2 \)=.428 (Fig. 3).

Performance in the search (a) and memory probe (b) tasks as a function of cue location and cue type for Experiment 1. The error bars represent the 95 % confidence intervals of the mean cued-uncued difference

Performance in the search (a) and memory probe (b) tasks as a function of cue location and cue type for Experiment 2. The error bars represent the 95 % confidence intervals of the mean cued-uncued difference
The same ANOVA ran with error rate as the dependent measure indicated no effect of cue type on error rate, F(1,17)<1, but showed significantly fewer errors were made in the valid (5.21±0.7 %) compared to invalid (9.86±0.94 %) cued conditions, F(1,17)=8.843, p=.009, \( {\eta}_p^2 \)=.342. In addition, cue type and cue validity interacted, F(1,17)=5.227, p=.035, \( {\eta}_p^2 \)=.235. The cue validity effect and the interaction were in the same direction as the RT results so do not indicative of a speed-accuracy tradeoff.
Memory probe condition
We analyzed memory probe performance using a 2 (cue type) × 2 (memory probe location: cued or uncued) repeated measures ANOVA with d’ as the dependent variable. We used z(proportion of hits) – z(proportion of false alarms) to calculate d’ with higher d’ indicating increased memory performance. A hit was the proportion of trials in which the probe letter was one of the items that was present in the memory display and the participants reported that it was present. A false alarm was the proportion of trials in which the probe letter was not present in the memory display, but participants reported that it was. Within each cue type condition, we collapsed across cue validity conditions as cue validity loses its meaning when the probe item was absent from the display. Cue type did not affect memory performance, F(1,17)=2.352, p=.143, \( {\eta}_p^2 \)=.122, but memory was better at the cued (1.72±0.14) compared to the uncued location (1.42±0.13), F(1,17)=20.652, p<.001, \( {\eta}_p^2 \)=.548. Critically, there was no interaction between memory probe location and cue type, F(1,11)<1.Footnote 3 In addition, we calculated a single false alarm rate that was independent of cue type and used that for both validity conditions. This was because validity loses its meaning when the probe item was not in the search display.
Discussion
In Experiment 1, we observed a contingent capture effect in RTs such that the cue validity effect was larger following color cues than following onset cues. In contrast, however, in the memory probe data we observed a cue validity effect that was unaffected by cue type. But, before making any conclusions it is noteworthy that while onset cues were goal inconsistent for the search task, they were arguably consistent with the memory probe task in which the two items were both white onsets. Therefore, to further test for the roles of sequential dependencies in contingent capture paradigms, we conducted Experiment 2 in which we made onsets goal consistent.
Experiment 2
Experiment 2 replicated Experiment 1, except that targets were now defined as onsets rather than the color item. If the data pattern matches that of Experiment 1 (contingent capture effects for RT, but not for memory), there would be strong evidence for sequential dependencies playing a role in generating RT contingent capture effects.
Method
Participants
Eighteen University of Toronto undergraduate students participated in exchange for course credit. All students reported normal or corrected-to-normal vision, were naïve to the study’s purpose, and provided informed consent.
Stimuli and apparatus
The stimuli were the same as Experiment 1 except that the target display now consisted of only the white dot and were presented on an LCD monitor (screen resolution: 2,560 × 1,440; refresh rate: 144 Hz).
Procedure
The procedure was the same as Experiment 1 except that participants now reported the location of a white dot.
Design
The design was the same as Experiment 1.
Results
Search condition
We used the same trimming (9.1 % of trials discarded) procedure and analyses as in Experiment 1. A 2 (cue type × (cue spatial validity), repeated measures ANOVA with RTs as the dependent variable indicated no main effect of cue type, F(1,17)=1.156, p=.297, \( {\eta}_p^2 \)=.064. The cue validity main effect was significant, F(1,17)=17.699, p=.001, \( {\eta}_p^2 \)=.510, with faster RTs in the valid (560±20 ms) than in the invalid (581±20 ms) condition. Importantly, replicating the contingent capture effect, in the onset cue condition there was a positive validity effect (56±7 ms), but in the color cue condition there was a negative validity effect (-15±5 ms), resulting in a significant interaction, F(1,17)=79.963, p<.001, \( {\eta}_p^2 \)=.825.
In the corresponding error rate analysis, there was a main effect of cue type, F(1,17)=9.726, p=.006, \( {\eta}_p^2 \)=.364, with a lower error rate in onset (4.8±0.8 %) than the color condition (6.5±1.0 %). The main effect of cue validity was non-significant, F(1,17)=2.099, p=.166, \( {\eta}_p^2 \)=.110. Finally, the interaction was significant, F(1,17)=8.866, p=.008, \( {\eta}_p^2 \)=.343. This reflected a larger cue validity effect in the onset (3.9±0.9 %) than in the color condition (-1.5±1.3 %), thus did not represent a speed-accuracy tradeoff.
Memory probe condition
The 2 (cue type) × 2 (memory probe location: cued or uncued) repeated measures ANOVA with d’ as the dependent variable indicated to main effect of cue validity, F(1,17)=2.330, p=.145, \( {\eta}_p^2 \)=.121, or cue type, F(1,17)=3.873, p=.066, \( {\eta}_p^2 \)=.186. Critically, consistent with contingent capture, in the onset cue condition there was a positive validity effect (0.32±0.11), but in the color cue condition there was a negative validity effect (-0.12±0.07 ms), resulting in a significant interaction, F(1,17) = 79.963, p < .001, \( {\eta}_p^2 \) = .825.
Discussion
In Experiment 2, we observed data patterns consistent with contingent capture in both the search task and the memory probe task. On the one hand, in both tasks onset cues generated large validity effects in both the search and memory conditions. On the other hand, color cues generated a significant negative cue validity effect in the search task which was numerically reflected in the memory task. This same location cost previously been attributed to sequential dependencies (Carmel & Lamy, 2014). As for the differences between Experiments 1 and 2, we will discuss further in the General discussion .
General discussion
In both experiments we found typical contingent capture effects in RTs with the search tasks. This was not, however, the case with the memory probe data. Specifically, there was no contingent capture effect within the memory data when targets were color-based (Experiment 1), but it was present when targets were onset-based (Experiment 2). In addition, we also demonstrated that cueing effects on memory remain while using a partial rather than a full report method (Gaspelin, Leonard, & Luck, 2015). This is important given that range of issues that prohibit one from making strong claims regarding memory from full report data (e.g., Sperling, 1960). Theoretically, the data have implications regarding sequential dependencies within the cueing paradigm as well as an asymmetry between how strongly onsets and color defined cues capture attention.
The search data from both experiments are consistent sequential dependencies playing a role in cueing paradigms. This led to a reduced capture effect in first experiment and a same-location cost at the cued location following a ACS incompatible cue in the second experiment. We suggest that these effects are, at least partially, attributable to sequential dependencies between the cue and the partially mismatching target appearing at that location. Indeed, that cues partially mismatching targets can lead to RT costs has been documented before (Carmel & Lamy, 2014; Huffman et al., 2017; Klein, Wang, Dukewich, He, & Hu, 2015). Given that this pattern of RTs is also predicted by contingent capture accounts, we look to the memory probe data to differentiate between the two possibilities.
The memory data from Experiment 1 clearly suggest that both cues were capturing attention equally as the validity effect in memory was unchanged by cue type. The data from Experiment 2, however, complicate matters. Rather than replicating the results from the color task, we observed contingent capture effects in memory in the onset task. In conjunction with the search data this pattern suggests one of two possibilities. One possibility is that there is an asymmetry between how strongly onsets and color cues capture attention. In particular, it is possible that the more salient onset cues are more likely to capture attention regardless of an individual’s goals. In Experiment 1, both the ACS compatible color cue and ACS incompatible, but highly salient, onset cues capture attention, but the validity effect caused by onset cues is reduced due to object-updating costs (Carmel & Lamy, 2014; Huffman et al., 2017). In constrast, in Experiment 2, the less salient color cues do not capture attention leading to no validity effect in either RT or memory. Furthermore, while color cues do not lead to a validity effect in the search task, they do lead to lead to a same-location cost, which can be explained as an object-updating cost (Carmel & Lamy, 2014).
That ACS incompatible onset cues capture attention is also consistent with recent work by Gaspelin et al. (2016). They found that whether or not a cue validity effect is found in such conditions depends on the difficulty of the subsequent search display. If the search is easy, the cued distractor is quickly rejected and the target is selected next such that small or no validity effects our found. If the search is hard, attention dwells on each selected distractor longer, leading to larger validity effects. As our study was not designed to differentiate between this attentional dwelling account and our sequential dependency account more research will be needed to do so. We also note, however, the fact the findings from Experiment 1 are consistent with their findings adds support for the memory probe method as a valid measure of attentional capture independent of post-capture effects such as sequential dependencies or attentional dwell.
A second possibility is that our findings are due to sequential dependencies in both the search and memory tasks. According to this interpretation, the RT data is interpreted the same as before. The interpretation of the memory data, however, changes. Specifically, in Experiment 1, one could assume that color cues were more likely to capture attention, but the cue validity effect in memory was reduced due to an object-updating cost between the color cue and the white memory stimulus. In Experiment 2 there was no longer any inconguence between the ACS compatible onset cues and the memory items leading to object-updating costs such that the memory data is consistent with contingent capture. We note, however, that such an interpretation would predict a same-location cost in the memory data following color cues as object-updating between the color cue and the memory item would be necessary. Furthermore, there is a theoretical reason why this possibility is less likely. In particular, a perceived task or context change can eliminate feature integration effects where they would be predicted otherwise (Halvorson, Wagschal, & Hazeltine, 2013; Huffman & Pratt, 2017, Experiment 2, no-search condition) and task demands can determine which object-updating effects are observed (Memelink & Hommel, 2013; Schumacher & Hazeltine, 2016). Given that the memory task was a sub-task that occured on a minority of trials and the task relevant feature (the letter’s shape) is absent from the cue display, we believe that it is unlikely that the memory display would invoke the retrieval of the cue’s object file such that object-updating would be unncessary.
Given our suggestion regarding the role of sequential dependencies in cueing task, one related and noteworthy aspect of the current design is that in the color cue condition the color alternated between the cue and search target approximately one half of time. If the object-updating account of reduced cueing effects in the onset condition is correct, then why does it not reduce the cueing effect in the color condition where there are also non-spatial feature switches (see Folk & Remington, 2008; Irons, Folk, & Remington, 2012)? Previous studies have suggested that task demands can impact the type of features that are included within an object representation (Memelink & Hommel, 2013; Schumacher & Hazeltine, 2016) which is consistent with demonstrations that non-spatial feature repetition effects (benefits or costs) occur in cueing tasks when discrimination, but not detection, responses are required for the task (Klein et al., 2015).Footnote 4 In other words, stimuli are coded in terms of which features are necessary for completing the task. In the current task, the relevant feature for completing the task is color generally rather than any specific color. Therefore, it is possible the cues were coded for in terms of having a color or being white (or, possibly, as an onset) in the object file. This would lead to there being no cost for going from a red cue to a green target, because in both the relevant feature ‘color’ repeats which leads to the observed RT patterns.
In addition to providing a possible alternative explanation for the contingent capture effect, it is important to note that the sequential dependencies found in this study fit within a larger movement in cognitive science which attempts to understand cognition in terms of prediction coding (Clark, 2013; Friston & Kiebel, 2009). From this viewpoint, rather than the observed effects being accounted for by object files, they would be framed in prediction success or failure. For example, once a white onset cue is perceived, the best predictionFootnote 5 for what will appear next in that location is another white stimulus. When there is a violation of this assumption, there is a prediction error that must be resolved before the stimulus can be perceived, leading to an RT cost.
Notes
Recently, Gaspelin, Ruthruff, and Lien (2016) also hypothesized that attentional capture by ACS incompatible onset cues is masked by processing differences occurring after the cues capture attention.
While there are also inter-trial sequential dependency effects (e.g., Goller & Ansorge, 2015), here we are concerned primarily with such effects within a trial.
See Supplementary Material for Bayesian analyses of the RT and memory probe data for Experiments 1 and 2.
In a related observation, Folk and Anderson (2010) reported the possibility of a general color ACS which may also be accounted for in term object file updating with color as a feature.
When we say “best prediction” here, we mean the best prediction according to a lifetime of experiencing a world that has considerable stability rather than best prediction according to the experimental design in which repetition typically happens at chance levels.
References
Ansorge, U., Becker, S.I. (2012) Automatic priming of attentional control by relevant colors. Attention, Perception, & Psychophysics, 74(1), 83–104.
Born, S., Kerzel, D., & Pratt, J. (2015). Contingent capture effects in temporal order judgments. Journal of Experimental Psychology: Human Perception and Performance, 41(4), 995.
Carmel, T., & Lamy, D. (2014). The Same-Location Cost Is Unrelated to Attentional Settings: An Object-Updating Account. Journal of Experimental Psychology: Human Perception and Performance, 40, 1465–1478.
Clark, A. (2013). Whatever next? Predictive brains, situated agents, and the future of cognitive science. The Behavioral and Brain Sciences, 36, 181–204.
Clark, A. (2015) Radical Predictive Processing. The Southern Journal of Philosophy 53, 3–27.
Folk, C. L., & Anderson, B. A. (2010). Target-uncertainty effects in attentional capture: color-singleton set or multiple attentional control settings? Psychonomic Bulletin & Review, 17(3), 421–6.
Folk, C. L., Remington, R. W., & Johnston, J. C. (1992). Involuntary covert orienting is contingent on attentional control settings. Journal of Experimental Psychology: Human Perception and Performance Enhanced, 18, 1030–1044.
Folk, C. L., & Remington, R. W. (2008). Bottom-up priming of top-down attentional control settings. Visual Cognition, 16(2-3), 215-231.
Friston, K., & Kiebel, S. (2009). Predictive coding under the free-energy principle. Philosophical Transactions of the Royal Society of London, 364, 1211–1221.
Gaspelin, N., Leonard, C. J., & Luck, S. J. (2015). Direct Evidence for Active Suppression of Salient-but-Irrelevant Sensory Inputs. Psychological Science , 26, 1740–1750.
Gaspelin, N., Ruthruff, E., & Lien, M. C. (2016). The problem of latent attentional capture: Easy visual search conceals capture by task-irrelevant abrupt onsets. Journal of Experimental Psychology: Human Perception and Performance, 42(8), 1104.
Goller, F., & Ansorge, U. (2015). There is more to trial history than priming in attentional capture experiments. Attention, Perception, & Psychophysics, 77, 1574-1584.
Halvorson, K. M., Wagschal, T. T., & Hazeltine, E. (2013). Conceptualization of task boundaries preserves implicit sequence learning under dual-task conditions. Psychonomic Bulletin & Review, 20, 1005-1010.
Hommel, B. (1998). Event files: Evidence for automatic integration of stimulus-response episodes. Visual Cognition, 5, 183-216.
Huffman, G., & Pratt, J. (2017). The action effect: Support for the biased competition hypothesis. Attention, Perception, & Psychophysics, 1-12.
Huffman, G., Al-Aidroos, N., & Pratt, J. (2017). Salience drives non-spatial feature repetition effects in cueing tasks. Attention, Perception, & Psychophysics, 79, 212-222.
Irons, J. L., Folk, C. L., & Remington, R. W. (2012). All set! Evidence of simultaneous attentional control settings for multiple target colors. Journal of Experimental Psychology: Human Perception and Performance, 38(3), 758.
Kahneman, D., Treisman, A, & Gibbs, B. J. (1992). The reviewing of object files: object-specific integration of information. Cognitive Psychology, 24, 175–219.
Kim, M. S., & Cave, K. R. (1995). Spatial attention in visual search for features and feature conjunctions. Psychological Science, 6(6), 376-380.
Klein, R. M., Wang, Y., Dukewich, K. R., He, S., & Hu, K. (2015). On the costs and benefits of repeating a nonspatial feature in an exogenous spatial cuing paradigm. Attention, Perception, & Psychophysics, 77, 2293-2304.
Lien, M. C., Ruthruff, E., Goodin, Z., & Remington, R. W. (2008). Contingent attentional capture by top-down control settings: converging evidence from event-related potentials. Journal of Experimental Psychology: Human Perception and Performance, 34(3), 509.
Memelink, J., & Hommel, B. (2013). Intentional weighting: a basic principle in cognitive control. Psychological Research, 77, 249–59.
Schumacher, E. H., & Hazeltine, E. (2016). Hierarchical Task Representation: Task Files and Response Selection. Current Directions in Psychological Science, 25, 449–454. 5085
Sperling, G. (1960). The information available in brief visual presentations. Psychological monographs: General and applied, 74(11), 1.
Theeuwes, J. (1992). Perceptual selectivity for color and form. Perception & Psychophysics, 51, 599–606.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
ESM 1
(DOCX 12 kb)
Rights and permissions
About this article

Cite this article
Huffman, G., Antinucci, V.M. & Pratt, J. The illusion of control: Sequential dependencies underlie contingent attentional capture. Psychon Bull Rev 25, 2238–2244 (2018). https://doi.org/10.3758/s13423-017-1422-5
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13423-017-1422-5