
Abstract
Pursuing dating relationships is important for many people’s well-being, because it helps them fulfill the need for stable social relationships. However, the neural underpinnings of decision-making processes during the pursuit of dating interactions are unclear. In the present study, we used a novel online speed dating paradigm where participants (undergraduate students, N = 25, aged 18–25 years, 52% female) received direct information about acceptance or rejection of their various speed dates. We recorded EEG measurements during speed dating feedback anticipation and feedback processing stages to examine the stimulus preceding negativity (SPN) and feedback-related brain activity (Reward Positivity, RewP, and theta oscillatory power). The results indicated that the SPN was larger when participants anticipated interest versus disinterest from their speed dates. A larger RewP was observed when participants received interest from their speed dates. Theta power was increased when participants received rejection from their speed dates. This theta response could be source-localized to brain areas that overlap with the physical pain matrix (anterior cingulate cortex, dorsolateral prefrontal cortex, and the supplementary motor area). This study demonstrates that decision-making processes—as evident in a speed date experiment—are characterized by distinct neurophysiological responses during anticipating an evaluation and processing thereof. Our results corroborate the involvement of the SPN in reward anticipation, RewP in reward processing and mid-frontal theta power in processing of negative social-evaluative feedback. These findings contribute to a better understanding of the neurocognitive mechanisms implicated in decision-making processes when pursuing dating relationships.
Similar content being viewed by others

The pursuit of dating relationships reflects a need for social affiliation, serves as a secondary reward that provides subjective pleasure, and fulfills biologically related needs (Ait Oumeziane et al., 2017). Neuroimaging evidence has shown that people in intimate relationships activate the dopamine reward system (i.e., the ventral tegmental area and caudate nucleus; Acevedo & Aron, 2014), which is associated with physical health and psychological well-being (Acevedo & Aron, 2014). Other studies have found that the breakup of an intimate relationship can cause social pain, which activates brain areas that overlap with experiencing physical pain (dorsal anterior cingulate cortex [dACC]; dorsolateral prefrontal cortex [dLPFC]; and anterior insula [AI]; Eisenberger, 2015; Seminowicz & Moayedi, 2017). Rejection by a loved one is commonly identified as a negative event that has profound meaning, often accompanied by negative emotions, anxiety, and depression, or even suicide or homicide (Fisher et al., 2010; Joel et al., 2019; van der Veen et al., 2019).
Although some studies have investigated the neural response to cues that either confirm an intimate relationship (Acevedo et al., 2012; Aron et al., 2005) or communicate romantic rejection (break-up; Fisher et al., 2010, Kross et al., 2011), less attention has been directed to the neural correlates of the pursuit of dating relationships—thus selecting potential dates and awaiting feedback regarding a match or mismatch. One of the challenges for this type of research is to simulate real-world situations that assess the pursuit of dating relationships in an ecologically valid way in a laboratory setting. Recently, van der Veen et al. (2019) developed an online dating task in which participants were presented with profiles of individuals of the other sex. Participants were instructed to decide whether these individuals were desirable or not. In a subsequent EEG session, participants were again shown photos of the same individuals, together with the participants’ evaluations of these individuals. The participants then received feedback from the speed date that could result in a match. That study found larger P3 responses when participants received positive versus negative evaluations from their speed date, which was interpreted to reflect the processing of the rewarding characteristics of romantic interest. Thus, the above-cited study examined the neural correlates of processing feedback from potential dates but did not examine the neural correlates of participants’ motivation regarding their decision-making during speed dating. In the current study, we used a novel speed dating paradigm that allowed us to examine (1) the speed dating decision-making stage, (2) the speed dating feedback anticipation stage, and (3) the speed dating feedback processing stage. Capitalizing on the high temporal precision of the EEG technique, we focused on event-related potentials that characterize anticipatory processes, as well as the processing of rewarding feedback as indexed with Reward Positivity. We also used time-frequency analyses to examine frequency-specific modulations in the EEG during the feedback processing stage.
Previous studies have used the stimulus preceding negativity (SPN) to study anticipatory motivation. The SPN is a slow negative potential that increases gradually before the feedback stimulus (Van der Molen et al., 2014). The SPN was considered to be an indicator of affective or motivational valence before giving action feedback (Böcker et al., 2001; Pornpattananangkul & Nusslock, 2015). A large body of studies revealed larger SPN amplitudes when anticipating reward versus nonreward (Donkers et al., 2005; Foti & Hajcak, 2012; Pornpattananangkul & Nusslock, 2015). Because the SPN reflects the anticipatory motivation before feedback (Pornpattananangkul & Nusslock, 2015), the SPN would be a feasible indicator to reveal differences in anticipatory motivation between feedback from the various speed dates in our current study.
During feedback processing, the event-related potential displays a prominent deflection in the ERP at around 250–350 ms that is sensitive to the valence of the feedback (such as rewarding feedback). For example, feedback signaling reward often is associated with a positive deflection, which has been termed reward positivity (RewP) (Foti et al., 2011). Previous studies have applied the RewP as a neural indicator for reward processing to monetary and social rewarding feedback (Ethridge et al., 2017). Several studies also have found that feedback signaling prediction errors (such as unexpected negative or positive feedback) result in a negative deflection in the ERP that co-occurs in the RewP time-window. This negative potential has been referred to as feedback-related negativity (FRN). According to reinforcement learning accounts, the FRN reflects the computation of negative reward prediction error (feedback is worse than expected) (Holroyd & Coles, 2002; Nieuwenhuis et al., 2004), whereas other accounts have found that the FRN is sensitive to unsigned prediction errors; that is, the FRN is increased for unexpected feedback regardless of its valence in both time-estimations tasks (Ferdinand et al., 2012), as well as social evaluative feedback processing (van der Molen et al., 2014, 2017, 2018).
There is ongoing debate on whether the feedback-related activity in the 250–350 ms time-window post-feedback reflects a single component (such as RewP or FRN) or whether it could reflect multiple components that are present dependent on the type of feedback presented (for example, reward or prediction errors during conflict monitoring; Holroyd et al., 2008, 2012; Proudfit 2015; Cavanagh et al., 2010; Cohen et al., 2011). Therefore, for simplicity, we refer to this component as the RewP but acknowledge the possibility that the ERP in this feedback processing time-window (250–350) is characterized by different aspects of processing of the feedback stimulus, and could therefore consist of multiple components.
Related to this issue of ERP component overlap, it has been recommended to examine ERPs in conjunction with time-frequency EEG activity, particularly when different components in the ERP hinder the appropriate quantification of these ERP components (Cohen et al., 2011). Studies using time-frequency decomposition of the EEG signal have revealed valuable information about the neural correlates of feedback processing in both cognitive and affective domains (Cavanagh et al., 2012; Yao et al., 2019). For example, enhanced frontal theta oscillatory activity has been observed when processing feedback that signals conflict, such as reward prediction errors (Janssen et al., 2016) and unexpected social rejection feedback (van der Molen et al., 2017; 2018, van der Veen et al., 2018). Notably, this enhancement in frontal theta power occurs in the same time-window as the RewP and therefore could provide important and complementary information that will help to elucidate the functional significance of feedback-related brain activity (for relevant discussions, see Cohen et al., 2011; Holroyd et al., 2012). Therefore, our study explores how EEG activity in the time domain (ERP) and time-frequency domain (theta power) is modulated by the processing of social feedback in a speed dating experiment.
Taken together, the present study assessed the neurophysiological activity associated with pursuing dating relationships in anticipating and experiencing speed dating feedback. We introduced a novel “online dating” task in which participants saw and chose their liked and disliked speed dates and saw each speed date’s decision. This allowed us to capture brain activity during different stages of the speed dating process. We tested the following hypotheses: (1) The SPN would be larger when awaiting acceptance feedback than rejection feedback due to the high motivation to establish dating relationships (Aron et al., 2005); (2) there would be larger RewP amplitudes for processing match feedback, whereas the RewP would be smaller for rejection feedback; and (3) based on recent findings suggesting enhanced midfrontal theta power for processing unexpected social rejection feedback (Cristofori et al., 2013; van der Molen et al., 2017), rejection in our study would result in the largest increase in theta power relative to the other conditions. Exploratively, we performed source analyses to examine the neural underpinnings of the EEG components (SPN, RewP, theta power). Based on an intracranial and a recent EEG source-localization study of social exclusion (Cristofori et al., 2013) and unexpected rejection (Van der Molen et al., 2017), we expected that the rejection-induced theta power would be mainly associated with enhanced activity in neural regions associated with saliency detection with the ACC acting as a key neural source.
Method
Participants
Participants were recruited to participate in the experiment through posters, online ads, and school media. Twenty-six healthy participants aged 18–25 years were recruited from Shenzhen University in China. All of the participants reported being single and heterosexual. No participants had any current or past mental or psychiatric history. All participants were right-handed and had normal or corrected-to-normal vision. Data were excluded from one participant due to noisy EEG. Finally, data from 25 participants (mean age = 20.07, SD = 1.96, 13 women) were analyzed. Regarding the break-up status of the participants, 11 had never been in a romantic relationship and 14 had experienced a break-up within the previous few months (mean = 27.07 months; SD = 28.34 months; minimum = 6 months; maximum = 60 months).Footnote 1 The study protocol was approved by the ethics committee of the Faculty of Medicine at Shenzhen University, and all participants signed informed consent before inclusion. All participants received a reward of 80 Yuan after the experiment.
Stimuli and experimental procedure
We introduced a novel online speed dating task that combined the social-judgment paradigm (Somerville et al., 2006) and the speed dating paradigm (Cooper et al., 2013). Participants were informed that they were taking part in a multi-university speed dating study and were required to submit a digital photo of themselves with a neutral expression. Furthermore, on the day of the photo submission, a standardized photo of the participant (processed to the same size and background as the experimental material) was sent to the participant to ensure that he or she was satisfied with the final photo presented to their speed dates. Participants were allowed to replace the photos within a week of the first experiment if they were not satisfied with the current version. According to an earlier study (Gunther Moor et al., 2010), fictional participants from other universities were photographed with neutral faces. These fictional participants acted as speed dates in our experiment. The Self-Assessment Manikin (SAM; Bradley & Lang, 1994) was used to ensure that the photos used in the experiment had neutral expressions. Finally, we selected a total of 340 photos (170 males and 170 females) of potential speed dates. These photos were taken from different universities. All photos were cropped to a standard size (185 × 240 pixels) and replaced with a standard background color (R: 44, G: 44, B: 44).
Participants took part in three sessions: (1) a pre-task rating session, (2) the online speed dating session, and (3) the post-task rating session. During the pre-task rating session, participants were invited to the lab to provide their likeability rating of the speed dates. First, the participants provided their personal information, including name, gender, date of birth, height, weight, educational major and grade, phone number, and email address. The personal data were collected to increase the validity of involvement and were kept secure, with only research personnel having access to them. Identifiable personal information was not used in the analysis and was destroyed after experiment. Next, participants were shown photos of their speed dates (that is, individuals of the opposite sex). For each photograph, participants were instructed to rate how much they liked the speed date based on their first impression using a seven-point scale, ranging from “1, not at all”, to “7, very much” (Fig. 1a). After completing the pre-task rating session, participants were told that what they had just seen were the speed dates from other universities participating in this project. These speed dates also would complete ratings based on the first impressions of photos of the participant. In fact, the rating on the participants’ photos was not made by real speed dates but manipulated by the experimenter.

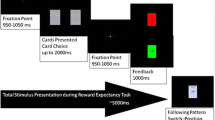
Experimental task. a Participants rated the likability of the speed dates before the task. b During the EEG experiment, participants were first required to judge whether they were interested in getting to know the speed date better, with a response window of 3,000 ms. Their choices were then highlighted and remained on the screen for 3,000 ms. Finally, the feedback from their speed date was presented for 2,000 ms, indicating whether they had been accepted or rejected by their speed date. c Participants rerated the likability of each speed date
Approximately 1 or 2 weeks after the first session, participants came back to the laboratory to complete the EEG task during the online speed dating session. Participants were shown photos of the speed dates and were instructed to make a judgment regarding the question, “Would you be interested in getting to know this person better?” Thereafter, the participants were shown feedback from their speed dates. This resulted in four different conditions: a Match condition (both the participant and the speed date answered “yes” to the question), a Rejection condition (the participant said “yes,” but the speed date said “no”), a Disinterest condition (both the participant and the speed date answered “no”), and an Unrequited condition (the participant said “no” and the speed date said “yes”). In addition, participants were told that only in case of a Match would they receive contact information of the speed dates.
Notably, during the EEG task, participants were free to choose who they wanted as a potential date and who they did not want. This is an important difference from previous studies (Cooper et al., 2013; van der Veen et al., 2019), in which participants were forced to choose at least 50 percent of the “yes” choices. We believe this manipulation would enhance the trustworthiness of experimental manipulations, and thus contribute to the ecological validity of the experiment.
A schematic of the EEG session is presented in Fig. 1b. Participants were shown photographs of the speed dates and asked to judge whether they were interested in getting to know that person better. Judgments were made by pressing one of two buttons (“F” and “J”), which corresponded with the “Y” (yes) and “N” (no) buttons on the computer screen. The positions of the “Y” and “N” buttons on the screen were counterbalanced between the participants. Participants were required to make a judgment within a 3,000-ms time window after the photo of the speed date appeared. If they did not respond within this time window, the trial ended and feedback from the speed date on that trial was not shown. Upon a button press, the button turned green for 3,000 ms to indicate the participants’ choice. Finally, feedback of the speed date was presented for 2,000 ms, with a “√” to indicate interest and “X” to indicate disinterest (210 × 210 pixels). In fact, the speed date’s feedback was randomly generated, so there was a 50% probability that the participant would receive feedback indicating interest from the speed date. Participants were only shown photos of speed dates of the opposite gender. For each participant, there were 10 practice trials (the same photos were used as practice trials for each gender) and 160 experimental trials.
Following the EEG session, participants completed a post-task likability rating in which they were again instructed to rate how much they liked each speed date (Fig. 1c). This allowed us to test whether participants’ pre-task rating was influenced by the experimental manipulation. Participants also indicated, on a seven-point Likert scale, how motivated they were to know about what their speed date thought of them (ranging from 1 “not at all” to 7 “very much”) and how pleasant they felt about the evaluation from the speed date (ranging from 1 “very unhappy” to 7 “very happy”). In addition, we administered the Rejection Sensitivity Questionnaire, Rosenberg Self-Esteem Scale and BDI-II Depression scale before the EEG task. Furthermore, the State-Trait Anxiety Inventory (STAI; Spielberger et al., 1983) and Positive and Negative Affect Schedule (PANAS; Watson et al., 1988) were used before and after the EEG session to measure state anxiety and mood changes, respectively. Moreover, participants were debriefed about the experiment after the last participant had finished the experiment and none of the participants reported suspicion about the experimental manipulation.
EEG recordings and processing
EEG data were recorded with 64 Ag/AgCl electrodes according to the 10–20 system (actiCAP, Brain Products, Germany; sampled at 1000 Hz). Electro-oculographic (EOG) signals were used to record ocular movements and eye blinks by using a surface electrode placed below the right eye. All impedances were kept below 10kΩ. EEG data were analyzed with BrainVision Analyzer 2.1 (Brain Products, Germany) and MATLAB (The MathWorks, Inc., Natick, MA).
EEG data were re-referenced offline to the bilateral mastoid electrodes and band-pass filtered between 0.1–30 Hz (48 db/oct) with a 50-Hz notch filter. We created 6,500-ms epochs that comprised 3,500 ms before speed date feedback onset and 3,000 ms after speed date feedback. Subsequently, independent component analysis (Lee, Girolami, & Sejnowski, 1999) was used to remove ocular artifacts. After additional baseline correction (see below for details on SPN and RewP), trials with voltage > ±80 μV were discarded. The number of artifact-free EEG epochs for further analyses is presented in Table 1. Although the number of trials differs between conditions (e.g., match vs. rejection feedback)—which is inherent to this social-decision making process—internal consistencies (as an index of reliability) of the ERPs per condition were excellent.Footnote 2
Event-related brain potential analyses
For SPN analyses, 3,500-ms artifact-free epochs were created comprising 3,300 ms pre-feedback and 200 ms post-feedback. In accordance with Van der Molen et al. (2014), we used the −2,400 to −2,000 ms pre-feedback interval for baseline correction. This interval ensured that no residual motor activity or decision-making processes were evident in the baseline correction period. By collapsing over the two conditions (i.e., interested and not interested in the speed date), we found a gradually increased SPN before the feedback onset. Thus, we calculated the mean amplitude of the 200 ms before the feedback onset at pooled frontocentral midline electrodes (Fz, FCz, Cz).Footnote 3
For RewP analysis, 1000 ms artifact-free epochs were created, including 200 ms before feedback onset and 800 ms post-feedback. The 200-ms pre-feedback time window was used for baseline correction. By collapsing over the four conditions, we found a pronounced RewP after the feedback during 255–355 ms. Thus, this time-window and pooled frontocentral midline electrodes (Fz, FCz, Cz) were used for assessment of the RewP.Footnote 4
Time-frequency power analyses
For each different type of feedback, artifact-free segments (−2,000 before and 2,000 ms after feedback onset) were transformed into the time-frequency domain using complex Morlet wavelets. For each segment, we obtained a complex time-frequency estimation with 30 logarithmically spaced steps, ranging from 1 to 30 Hz in the frequency domain. The Morlet parameter of the central frequency was set at 1 Hz and time resolution (in units of Full Width Half Maximum) was set to 5 s. The spectrogram was baseline-corrected using the subtraction approach at each frequency (Hu et al., 2014), in which −500 to −200 ms interval before the feedback onset was used as the baseline. By collapsing over the four conditions, we found a pronounced theta burst occurring 200–400 ms after the feedback at Fz, corresponding with findings from Van der Molen et al. (2017, 2018). Thus, we calculated the averaged theta power (4–8 Hz) in the 200–400-ms time-window following the feedback onset at Fz.
Source-localization analyses
Source-localization of theta power was performed on the single-trial level for each feedback condition using Brainstorm (Tadel et al., 2011), which is a free and documented software package available in Matlab (http://neuroimage.usc.edu/brainstorm). The default ICBM152 anatomy, distributed by the Montreal Neurological Institute (MNI), was used as a tessellated cortical mesh template surface due to the lack of individual MRI templates. The BrainProducts Easycap 64-channel layout was co-registered with the ICBM152 anatomy. The EEG forward model of volume currents was calculated by a symmetric boundary element model with OpenMEEG (Gramfort et al., 2010), with which the default layers and conductivities parameters and the adaptive integration method were applied. This forward model uses three realistic layers corresponding to the scalp (1,922 vertices, relative scalp conductivity = 1), the skull (1,922 vertices, relative skull conductivity = 0.0125), and the brain (1,922 vertices, relative brain conductivity = 1) (Ambrosini & Vallesi, 2016). The noise covariance matrix was based on the −500 to −200 ms baseline period before the feedback onset. Next, unconstrained cortical sources were calculated at the single trial level by using the depth-weighted minimum norm estimation (wMNE) approach (Baillet et al., 2001). This technique is robust to noisy EEG data and shows fair spatial resolution; it also provides reliable results for source-localization analysis of EEG data in the absence of individual MRI anatomies (Baillet et al., 2001). Finally, the source current strength (3 x5,005 vertices of the cortex surface) is obtained by multiplying the recorded EEG signal at each electrode on the time series by the wMNE inverse operator. Importantly, this linear transformation allows the time-frequency analysis to be calculated directly on the source space without changing the spectral characteristics of the underlying source (Ambrosini & Vallesi, 2016; Billeke et al., 2013). Z-score transformations were conducted to normalize the theta source results after averaging all the trials for each condition, using the 500 to −200 ms pre-feedback baseline as the reference interval. The Z-scores for source results were rectified in theta band (4–8 Hz) and averaged within the 200–400-ms post-feedback time windows for statistical analysis.
Statistical analysis
Statistical analyses were performed with IBM SPSS Statistics 21.0 (IBM, Armonk, NY). For behavioral data, we used t-tests to compare the reaction times (RTs) of two different choices (Participant Judgment: Yes or No), and the STAI and PANAS scores before and after the experiment (phase: pre and post). To investigate the change of likability ratings, we created the difference between post-task minus pre-task for the four feedback conditions (Match, Rejection, Unrequited, Disinterested). The difference scores as dependent variables were submitted to a 2 (Participant Judgment: Yes or No) × 2 (Speed date Feedback: Yes or No) ANOVA. The self-reported motivation ratings in two conditions (Participant Judgment: Yes or No) were compared by using paired t-test. Self-reported pleasantness ratings were submitted through a two-way ANOVA with Participant Judgment (Yes or No) by Speed date Feedback (Yes or No).
For EEG data, a paired t-test was used to compare the SPN of two judgments (Yes or No). Furthermore, RewP and Theta values were submitted separately into a 2 (Feedback Congruence: Congruent, Incongruent) by 2 (Feedback Valence: Positive/Negative) repeated measures ANOVA. Greenhouse-Geisser correction was used when sphericity was violated.
For theta source localization data, nonparametric cluster-based permutation testing was used to test for significant difference in source activity between each condition in the source space (Maris & Oostenveld, 2007) using Fieldtrip’s ft_sourcestatistics method (Oostenveld, Fries, Maris, & Schoffelen, 2011) as implemented in Brainstorm. The theta source data were averaged over frequency band (4–8 Hz) and time (200–400 ms), meaning that this test statistic only considered the spatial dimension. First, for every sample, a comparison of the two conditions was calculated based on the alpha level of 0.05 threshold. Samples that exceeded the critical t-values were then clustered and summed over t-values, which were based on spatial adjacency. Next, the cluster-level statistics were calculated. The Monte Carlo method was used for significance statistical testing with paired t-tests. The nonparametric cluster-level statistics was performed by calculating a p-value under 1000 random permutation distribution of the source data. The cluster-corrected alpha level of 0.05 was set for multiple comparisons.
Lastly, Pearson correlation analyses were performed to assess the association between self-reported pleasantness ratings and the condition-specific RewP and theta power, as well as the association between self-reported motivation and condition-specific SPN. No significant associations were found (all ps > 0.23).
Results
Behavioral data
A paired t-test indicated that the RTs for “Yes” Judgments were significantly longer than “No” Judgments, t(1,24) = 5.425, p < 0.001, Cohen’s d = 1.09, indicating that it took longer for participants to show interest in their speed date (1,380.87 ± 65.34 ms) than to show disinterest (1,182.79 ± 50.51 ms; Fig. 2a). Furthermore, there was a significant main effect of Participant Judgment (F(1, 24) = 11.21; p = 0.003; ηp2 = 0.32), suggesting that the participants significantly increased the likeability for their speed dates if they deemed them as potential dates vs. non-potential dates.Footnote 5 In addition, participants reported a stronger motivation to meet their potential dates (5.32 ± 0.15) than non-potential dates did (4.12 ± 0.15; t (1, 24) = 5.77, p < 0.001, Cohen’s d = 1.15; Fig. 2b).

a RTs the participants’ judgment. b Self-reported motivation ratings. c Self-reported pleasantness ratings. Error bars represent standard errors. ***p < 0.001
As depicted in Fig. 2c, we found a significant main effect of speed date feedback in self-reported pleasantness ratings: F (1, 24) = 42.69; p < 0.001; ηp2 = 0.64. In addition, the interaction between judgment and speed date feedback was significant (F(1,24) = 33.65, p < 0.001, ηp2 = 0.58), indicating that participants reported more pleasantness after Match outcomes (5.72 ± 0.19) than Rejection outcomes (3.08 ± 0.16; p < 0.001). However, there was no significant difference between Unrequited outcomes (4.20 ± 0.20) and Disinterest outcomes (4.00 ± 0.21; p = 0.519). We also decomposed the interaction for Judgment. Results indicated that participants reported more pleasantness after Match outcomes (5.72 ± 0.19) than Unrequited outcomes (4.20 ± 0.20; p < 0.001). In addition, participants reported less pleasantness after Rejection outcomes (3.08 ± 0.16) than Disinterest outcomes (4.00 ± 0.21; p = 0.001). The scores on the STAI-S and PANAS scales did not differ between administration moments (i.e., before and after the speed dating task; all p values > 0.16).
We also measured levels of Rosenberg Self-Esteem, BDI-II Depression, and Rejection Sensitivity. Mean scores on the self-report measures are presented in Table 2. The personality trait scores were used as covariates in the behavioral analysis and EEG data; we did not find these personality trait scores to be related to participants’ behavioral responses (all ps > 0.075) and EEG results (all ps > 0.11). Therefore, these personality trait scores were not used for further analysis.
Stimulus preceding negativity (SPN)
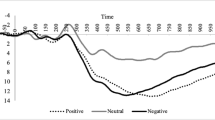
As depicted in Fig. 3, paired samples t-test indicated that the stimulus preceding negativity (SPN) was more negative when participants showed romantic interest (−3.98 ± 0.70 μV) relative to disinterest (−2.39 ± 0.56 μV) in their speed date; t(1,24) = −3.31, p = 0.003, Cohen’s d = −0.66).

a Feedback-locked grand-averaged ERP waveforms at pooled electrodes (Fz, FCz, Cz) for two participants’ judgments. b Average SPN amplitude for two participant’s judgments. c Scalp distribution represented by the average amplitude in a −200 to 0 ms time window. The blue shaded area indicates the baseline time window (−2,400 to 2,000 ms) and the gray shaded area indicates the quantified time window (−200 to 0 ms). In the EEG topographic map. *Electrodes used for calculation (from top to bottom, Fz, FCz, and Cz). Error bars represent standard errors. **p < 0.01
Reward positivity (RewP)
Grand-averaged ERPs at pooled electrodes (Fz, FCz, Cz) are depicted in Fig. 4a. We found a significant main effect of Feedback Valence (F(1,24) = 6.25, p = 0.020, ηp2 = 0.21) and a significant main effect of Feedback Congruency (F(1,24) = 28.29, p < 0.001, ηp2 = 0.54), which were included in a significant interaction between Feedback Valence x Feedback Congruency (F(1,24) =14.32, p = 0.001, ηp2 = 0.37)). Follow-up paired samples t-test revealed that the RewP to the speed date’s feedback (7.54 ± 1.44 μV) was significantly larger for Match outcomes relative to all other conditions (all ps < 0.001). The RewP was significantly larger for the Rejection condition (4.70 ± 1.25 μV) than the Disinterest condition (2.73 ± 0.91 μV; t(1,24) = 3.08, p = 0.005, Cohen’s d = 0.62), while the RewP between the Rejection (4.70 ± 1.25 μV) and Unrequited (4.04 ± 0.80 μV) conditions was not significant (t(1,24) = 0.96, p = 0.345, Cohen’s d = 0.19). In addition, the RewP was significantly larger for the Unrequited condition (4.04 ± 0.80 μV) than for the Disinterest condition (2.73 ± 0.91 μV; t(1,24) = 2.78, p = 0.010, Cohen’s d = 0.56).

a Feedback-locked grand-averaged ERP waveforms at pooled electrodes (Fz, FCz and Cz) in four conditions. b Average RewP amplitude per outcomes (c) Scalp distribution in four conditions for the average amplitude in a 255–355-ms time window indicated in A by the grey shaded area. Error bars represent standard errors. In the EEG topographic map, * represents the electrodes used for calculation (from top to bottom, Fz, FCz and Cz). ***p < 0.001, **p < 0.01, *p < 0.05
Theta power
As depicted in Fig. 5, we calculated the averaged theta power (4–8 Hz) in the 200–400-ms time-window following the feedback onset at Fz. The theta power yielded a significant main effect of Feedback Congruency (F(1,24) = 6.92, p = 0.015, ηp2 = 0.22) and a main effect of Feedback Valence (F(1,24) = 12.08, p = 0.002, ηp2 = 0.34), which were included in a significant interaction between Participant Judgment and Speed date Feedback (F(1,24) = 14.02, p = 0.001, ηp2 = 0.37). Follow-up paired samples t-test indicated that theta power was significantly higher in the Rejection condition than in all other conditions (all ps < 0.001). All other contrasts were not significant (all ps > 0.54). Exploratively, we examined feedback-related delta power (see supplemental material S2 for details) and observed a significant enhancement in delta power in both the Match (ps < 0.036) and Rejection (ps < 0.001) conditions relative to the Unrequited and Disinterest conditions.

Time-frequency power at Fz during the 200–400-ms post-feedback interval. a Time-frequency plots for the four conditions. b Average theta power per outcomes (c) Scalp distribution of theta power. Error bars represent standard errors. In the EEG topographic map, * represents the Fz electrode site. ***p < 0.001
Source localization for theta power
The neural sources underlying feedback-related theta activations are displayed in Fig. 6. For the rejection condition, the source maps reveal a distinct increase in theta power in comparison to the other feedback conditions. Nonparametric permutation testing was performed to test for significant condition differences in the theta activation patterns. These results are presented in Fig. 7. The contrasts between Rejection and the other conditions all revealed significant clusters, which suggests increased theta activation in the Rejection condition relative to the Match condition (cluster 1: size = 214, p = 0.024; cluster 2: size = 155, p = 0.045), the Disinterest condition (cluster 1: size = 181, p = 0.042; cluster 2: size = 175, p = 0.045), and the Unrequited condition (cluster: size = 174, p = 0.04). Although spatial precision of these condition differences cannot be inferred from these nonparametric permutation tests (Maris & Oostenveld, 2007; Sassenhagen & Draschkow, 2019), the source activity differences were observed within the 200–400-ms post-feedback interval over the prefrontal and cingulate cortices.

Theta oscillatory power source-localization maps during the 200–400-ms post-feedback window. Depicted are mid-sagittal slices (left and right) of theta power activation associated with the processing of romantic expression. The source activation maps are based on activation of at least 40 vertices (amplitude threshold of 50%)

Contrast maps of theta source activity for Rejection with the other conditions. The mid-sagittal slices (left and right) and axial views of the three contrasts are depicted. Only clusters of theta source activity that have passed cluster-based nonparametric permutation test and survived the correction are presented
We also performed source analysis on SPN and RewP, but no significant differences were found in source clusters between conditions (see Supplementary Material for details).
Discussion
The goal of this study was to examine the neural correlates associated with the acceptance and rejection related to online dating. Using a novel speed dating, while measuring brain activity with ERPs and EEG, we examined the anticipatory stage (waiting to receive the speed date’s feedback) and feedback processing stage (processing of the speed date’s feedback signaling either interest or disinterest). This set-up allowed us to disentangle the effects of reward (interest by the speed dates) versus nonreward (disinterest by the speed dates), as well as feedback conflict (for example, when feedback from the speed date was not in line with the judgments of the participant). Our behavioral data (measured during EEG experiment) showed greater emotional reactivity, shown by self-reported pleasantness, when participants received feedback from potential dates compared to non-potential dates. For electrophysiological data, participants showed a more negative SPN amplitude while awaiting social evaluation from potential dates than from non-potential dates, as expected. During the social evaluation feedback stage, Match outcomes (where both the participant and the speed date said “yes”) induced the largest RewP in all four conditions. Further, theta power was largest in response to Rejection outcomes (where the participant said “yes,” the speed date said “no”). This burst of theta power during rejection was source-localized to brain regions known to be relevant for processing physical and social pain (Cristofori et al., 2013; Eisenberger et al., 2003; Peyron et al., 2000), such as the ACC, the dlPFC, and supplementary motor cortices. To our knowledge, the present study is the first to explore patterns of neural activity during “pursuing dating relationships” and offers important insights into the role of SPN, RewP, and theta oscillations in dating relationship pursuit.
Our behavioral data revealed that participants “turned down” the majority of the speed date during the speed dating experiment and that decision times were significantly longer for showing interest in a speed date than rejecting a speed date. This effect could be interpreted to suggest that when individuals show interest in potential dates, they expose themselves to potential adverse effects (i.e., rejected by the speed date). Previous studies have shown that avoiding rejection is an important goal for most people (Baker & McNulty, 2013; Baumeister & Leary, 1995). When people try to pursue a new relationship, they consider whether exposing their intentions will make them experience the pain of rejection (Joel et al., 2019); in this case, the chance of being rejected by someone they like. In our study, the longer decision times for showing interest vs. disinterest could reflect minimization of the risk of adverse effects associated with the pursuit of dating relationships, and this risk is larger when showing interest vs. disinterest in a potential partner. As such, the decision times and trial numbers in our study provided an implicit index of the participants’ prudent behavior in choosing a potential dating partner, which confirmed our experimental manipulation.
Also, participants rated acceptance from potential dates as more pleasant than rejection. Because humans have a strong evolutionary motivation to have social interactions and relationships, social acceptance is highly rewarding and desired (Baumeister & Leary, 1995). Interestingly, we found no differences in emotional responses for nonpotential dates. That is, those speed dates for which the participant had no interest (Unrequited vs. Disinterest condition) showed similar self-pleasantness ratings, regardless of the feedback from nonpotential dates. In our study, the participants also reported a strong motivation to know the decision of potential dates, as shown by self-reported motivation. In fact, different social and motivational contexts affect how people feel about social feedback. People showed more pleasure and reward-related activity in response to social reward (for example, connection, cooperation, and conformity) for a close one rather than distant one (Hughes et al., 2018). Therefore, it is understandable that the participants had stronger motivation for potential dates, which may have increased the participants’ arousal in response to social feedback.
Our brain response and EEG results suggested clear differences in neural reactivity associated with feedback anticipation and processing thereof. During the speed date’s feedback anticipation stage, we observed a larger SPN when participants were hoping to receive interest. This result dovetails with prior findings on anticipating social evaluative feedback, where the SPN was found to be larger when individuals anticipated social acceptance feedback rather than rejection feedback (van der Molen et al., 2014). Our current SPN findings are also consistent with the participants’ self-report ratings, suggesting that participants have a strong motivation to know the potential partner’s social evaluation and feel that receiving feedback from a potential partner is more rewarding. Previous studies have found a larger SPN when people are more likely to receive reward than punishment (Donkers et al., 2005) or social acceptance versus rejection feedback (van der Molen et al., 2014). In our case, receiving interest from a potential dating partner meant the possibility of more subsequent communication, a strong social reward in a relationship (Cooper et al., 2013). This is consistent with previous studies that SPN reflects the anticipatory motivation before feedback (Pornpattananangkul & Nusslock, 2015). Thus, our results indicate that people have a high motivation to pursue interest from potential dates, which seems to be reflected in enhanced SPN amplitudes.
In contrast to previous social feedback anticipation studies, our current paradigm did not examine explicit expectancies from participants about the speed date’s feedback. That is, we examined what participants hoped for in terms of the feedback from the speed date, which is different from examining what feedback participants might expect to receive. For example, studies that have examined explicit expectancies about social evaluative feedback have found enhanced negativities in the typical RewP time-window (Dekkers et al., 2015; van der Molen et al., 2014, 2017; 2018), which increased significantly for unexpected social evaluative feedback. These studies used a social judgment paradigm in which participants were asked, “Do you think this person likes you?” Thus, explicit expectancies were measured that resulted in prediction errors (i.e., when feedback is not in line with participant’s expectancies). Instead, our study focused more on the participants’ own subjective preferences regarding the speed date by asking the question, “Would you be interested in getting to know this person better?” In this manner, motivational processes (hoping to receive positive answers from the romantically interesting speed date) seem to be more relevant in the current design than expectancies. However, similarly to van der Molen et al. (2014), we found enhanced SPN amplitudes while hoping to receive acceptance rather than rejection. This might suggest that the valence effects on the SPN are similar for explicit feedback expectancies and motivationally driven feedback anticipation.
During the speed date’s feedback processing stage, our results revealed the largest RewP amplitudes in the Match condition, which suggests an increased reward value when participants were presented with feedback signaling social acceptance by potential dates than nonpotential dates. This enhancement in the RewP after Match feedback might also relate to the participant’s individual preferences. A recent EEG study found that high-preference outcomes induce larger RewP amplitudes than medium reward outcomes (Peterburs et al., 2019). Although that study used chocolate as a reward rather than social feedback, individual preferences about social feedback are likely to have influenced the RewP amplitudes. For example, we found that RewP amplitudes were largest for feedback conditions in which participants indicated interest in the speed date (Match and Rejection). This finding is in line with a similar speed dating study by Van der Veen et al. (2019), who reported increased P3 amplitudes for Match feedback relative to the other feedback conditions. These authors interpreted this P3 enhancement after Match feedback to reflect the strong subjective motivational properties of this type of feedback. Future studies could further examine these potential subjective reward value effects on the RewP by examining participants’ judgments about the speed date on an interval scale, rather than binary judgments (Yes vs. No). Because subjective preferences show different levels, the interval scale can examine whether RewP is modulated by the linear variation of subjective preferences.
The current study also adds an important dimension to the existing literature on brain responses to feedback processing of pursuing dating relationships. For example, Van der Veen et al. (2019) only tracked brain responses during the presentation of feedback from the potential dates, and presented the participants’ judgment of the potential dates that was collected prior to the EEG session. Thus, participants passively watched their own judgments and the feedback of the potential dates. Moreover, participants in this study were forced to evaluate 50% of the speed date as “dateable.” Together, these manipulations might have resulted in “second thoughts” about participants’ judgments about the partners during the online speed dating session, as well as reduced task engagement in finding out the evaluation from the partners, and thereby confounding the reward processes that elicit the RewP. In the current paradigm, the willingness of participants and their speed dates was communicated in real time and participants were not forced to consider a fixed percentage of candidates as dateable. This might have contributed to the ecological validity of the paradigm, and particularly participants’ task engagement, a factor that is known to affect the amplitude of RewP (Bellebaum et al., 2010; Warren & Holroyd, 2012).
Previous studies have found that peer feedback incongruent with participants’ feedback expectancies resulted in larger feedback negativities (Dekkers et al., 2015; van der Molen et al., 2014, 2017, 2018). We did not observe this conflict effect in the time-domain EEG (that is, ERPs), but we did find it in the time-frequency domain of the EEG. Specifically, we observed that midfrontal theta power was exclusively stronger in the Rejection condition, where the participant’s interest in the potential partner was unanswered. This finding coincides with a series of EEG studies in which unexpected social rejection feedback resulted in a significant increase in mid-frontal theta power (van der Molen et al., 2017, 2018). This effect has been interpreted to reflect enhanced sensitivity of a social threat detection mechanism (Van der Molen et al., 2017, 2018). It has been postulated that humans have evolved a highly sensitive self-protection system that guards individuals from social disconnection (Eisenberger & Lieberman, 2004). Neuroimaging studies have revealed that the processing of social rejection cues is governed by brain regions that overlap with the physical pain matrix, such as the anterior insula (AI) and ACC, which are involved in cognitive-affective pathways that process information about the unpleasant value of a nociceptive stimulus (Eisenberger et al., 2003, 2007). Results from an intracranial EEG study by Cristofori et al. (2013) provided direct evidence that increased theta power during exclusion versus inclusion can be source-localized to brain areas overlapping with processing of physical pain, particularly the ACC and AI. Based on this overlap, those authors argued that enlarged theta power in response to social exclusion reflects a neural signature of social pain. Our source localization results show that theta-based source activity was significantly strongest in the Rejection condition. This activity was observed in the brain regions that cover the ACC and dlPFC—brain regions that have been shown to play a dominant role in the threat detection system (Eisenberger, 2015). However, source location results should be interpreted with caution because of their lack of precision compared to fMRI, for example. Due to the inverse problem, a limited number of electrodes and an infinite number of possible source locations result in relatively low spatial resolution (Asadzadeh et al., 2020). Our interpretation that the Rejection condition in this study elicited social pain is supported by the increased negative effect ratings from the participants self-reported pleasantness during rejection. However, this Rejection condition not only elicits intense social pain; it is also a condition that contains potential disconfirmation of an outcome that is hoped for and perhaps expected. Thus, implicit expectancies might have confounded the pure motivationally driven effects in this study.
Conclusions
This study has highlighted the different stages (speed dating decision making, awaiting, and processing of speed dating feedback) that characterize the pursuit of dating relationships. Specifically, our results confirm the notion that the stimulus preceding negativity is a neural correlate of reward anticipation and extend this view to anticipating a reward resulting from a potential dating relationship. We also suggest that the reward positivity reflects a neural response of the reward system related to receiving interest from potential dates, which is socially rewarding. Furthermore, our findings corroborate the idea that midfrontal theta oscillatory reactivity constitutes a neural signature of social pain resulting from social rejection by potential dates. Overall, our study highlights distinct sensitivity of anticipatory and feedback-related ERP components during the pursuit of dating relationships, and our results substantiate the important role of midfrontal theta oscillatory reactivity in regulating negative social effects.
Change history
01 October 2021
A Correction to this paper has been published: https://doi.org/10.3758/s13415-021-00949-y
Notes
Participants were divided into two groups according to whether they had been in a romantic relationship or not, which was used as a between-subject variable for subsequent analysis (behavioral data analysis and EEG data analysis). The main effects of the group were not significant (all ps > 0.151), and the interactions of any factor with the group were not significant (all ps > 0.086).
The split-half reliability was used to measure the internal consistency of the SPN and RewP at pooled electrodes (Fz, FCz, Fz). The odd and even trials are averaged and the correlation between the two is calculated (Threadgill et al., 2020), corrected using the Spearman-Brown prophecy formula (Nunnally et al., 1967). Spearman-Brown corrected split-half r of RewP for Match = 0.96, Rejection = 0.92, Unrequited = 0.89, and Disinterest = 0.92, and SPN for Yes judgment = 0.83 and No judgment = 0.84.
This positivity in the feedback-related ERP was already evident around the P2 component, so our RewP measure might have been subject to component overlap. However, PCA analysis revealed a distinct positive component with central dominance that yielded similar results as described for the RewP in this study (see Supplementary Material for details).
From here on we use the term potential dates to refer to speed dates that participants were romantically interested in, and nonpotential dates to refer to speed dates that participants were not romantically interested in.
References
Acevedo, B. P., & Aron, A. P. (2014). Romantic love, pair-bonding, and the dopaminergic reward system. Mechanisms of Social Connection: From Brain to Group, 55–70. https://doi.org/10.1037/14250-004
Acevedo, B. P., Aron, A., Fisher, H. E., & Brown, L. L. (2012). Neural correlates of long-term intense romantic love. Social Cognitive and Affective Neuroscience, 7(2), 145–159.
Ait Oumeziane, B., Schryer-Praga, J., & Foti, D. (2017). “Why don’t they ‘like’ me more?”: Comparing the time courses of social and monetary reward processing. Neuropsychologia, 107, 48–59. https://doi.org/10.1016/j.neuropsychologia.2017.11.001
Ambrosini, E., Vallesi, A. 2016. Asymmetry in prefrontal resting-state EEG spectral power underlies individual differences in phasic and sustained cognitive control. Neuroimage 124, 843–857. https://doi.org/10.1016/j.neuroimage.2015.09.035.
Aron, A., Fisher, H., Mashek, D. J., Strong, G., Li, H., & Brown, L. L. (2005). Reward, motivation, and emotion systems associated with early-stage intense romantic love. Journal of Neurophysiology, 94(1), 327–337.
Asadzadeh, S., Rezaii, T. Y., Beheshti, S., Delpak, A., & Meshgini, S. (2020). A systematic review of EEG source localization techniques and their applications on diagnosis of brain abnormalities. Journal of Neuroscience Methods, 339, 108740.
Baillet, S., Riera, J.J., Marin, G., Mangin, J.F., Aubert, J., Garnero, L. (2001). Evaluation of inverse methods and head models for EEG source localization using a human skull phantom. Physics in Medicine and Biology 46, 77–96. https://doi.org/10.1088/00319155/46/1/306.
Baker, L. R., & McNulty, J. K. (2013). When low self-esteem encourages behaviors that risk rejection to increase interdependence: The role of relational self-construal. Journal of Personality and Social Psychology, 104, 995–1018.
Baumeister, R. F., & Leary, M. R. (1995). The need to belong: desire for interpersonal attachments as a fundamental human motivation. Psychological Bulletin, 117(3), 497.
Bellebaum, C., Kobza, S., Thiele, S., & Daum, I. (2010). It was not MY fault: event-related brain potentials in active and observational learning from feedback. Cerebral Cortex, 20(12), 2874–2883.
Billeke, P., Zamorano, F., Cosmelli, D., Aboitiz, F., 2013. Oscillatory brain activity correlates with risk perception and predicts social decisions. Cerebral Cortex 23, 2872–2883. https://doi.org/10.1093/cercor/bhs269.
Böcker, K. B. E., Baas, J. M. P., Kenemans, J. L., & Verbaten, M. N. (2001). Stimulus-preceding negativity induced by fear: a manifestation of affective anticipation. International Journal of Psychophysiology, 43(1), 77–90.
Bradley, M. M., & Lang, P. J. (1994). Measuring emotion: the self-assessment manikin and the semantic differential. Journal of Behavior Therapy and Experimental Psychiatry, 25(1), 49–59.
Cavanagh, J.F., Frank, M.J., Klein, T.J., Allen, J.J.B. (2010). Frontal theta links prediction errors to behavioral adaptation in reinforcement learning. NeuroImage 49 (4), 3198–3209. https://doi.org/10.1016/j.neuroimage.2009.11.080.
Cavanagh, J. F., Zambrano‐Vazquez, L., & Allen, J. J. (2012). Theta lingua franca: A common mid‐frontal substrate for action monitoring processes. Psychophysiology, 49(2), 220–238. https://doi.org/10.1111/j.1469-8986.2011.01293.x.
Cohen, M. X., Wilmes, K., & Vijver, I. V. (2011). Cortical electrophysiological network dynamics of feedback learning. Trends in Cognitive Sciences, 15(12), 558–566. https://doi.org/10.1016/j.tics.2011.10.004
Cooper, J. C., Dunne, S., Furey, T., & O’Doherty, J. P. (2013). The role of the posterior temporal and medial prefrontal cortices in mediating learning from romantic interest and rejection. Cerebral Cortex, 24(9), 2502–2511.
Cristofori, I., Moretti, L., Harquel, S., Posada, A., Deiana, G., Isnard, J., ... & Sirigu, A. (2013). Theta signal as the neural signature of social exclusion. Cerebral Cortex, 23(10), 2437–2447.
Dekkers, L.M.S., van der Molen, M.J.W., Moor, B.G., van der Veen, F.M., van der Molen, M.W. (2015). Cardiac and electro-cortical concomitants of social feedback processing in women. Soc. Cogn. Affect. Neurosci. 10, 1506–1514. https://doi.org/10.1093/scan/nsv039.
Donkers, F. C., Nieuwenhuis, S., & Van Boxtel, G. J. (2005). Mediofrontal negativities in the absence of responding. Cognitive Brain Research, 25(3), 777–787.
Eisenberger, N. I. (2015). Social pain and the brain: controversies, questions, and where to go from here. Annual Review of Psychology, 66, 601–629.
Eisenberger, N.I., Lieberman, M.D., 2004. Why rejection hurts: a common neural alarm system for physical and social pain. Trends in Cognitive Sciences, 8, 294–300. https://doi.org/10.1016/j.tics.2004.05.010.
Eisenberger, N. I., Lieberman, M. D., & Williams, K. D. (2003). Does rejection hurt? An fMRI study of social exclusion. Science, 302(5643), 290–292.
Eisenberger, N. I., Gable, S. L., & Lieberman, M. D. (2007). Functional magnetic resonance imaging responses relate to differences in real-world social experience. Emotion, 7(4), 745.
Ethridge, P., Kujawa, A., Dirks, M. A., Arfer, K. B., Kessel, E. M., Klein, D. N., & Weinberg, A. (2017). Neural responses to social and monetary reward in early adolescence and emerging adulthood. Psychophysiology, 54(12), 1786–1799. https://doi.org/10.1111/psyp.12957
Ferdinand, N. K., Mecklinger, A., Kray, J., & Gehring, W. J. (2012). The processing of unexpected positive response outcomes in the mediofrontal cortex. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 32(35), 12087–12092. https://doi.org/10.1523/JNEUROSCI.1410-12.2012
Fisher, H. E., Brown, L. L., Aron, A., Strong, G., & Mashek, D. (2010). Reward, addiction, and emotion regulation systems associated with rejection in love. Journal of Neurophysiology, 104(1), 51–60.
Foti, D., & Hajcak, G. (2012). Genetic variation in dopamine moderates neural response during reward anticipation and delivery: Evidence from event-related potentials. Psychophysiology, 49(5), 617–626.
Foti, D., Weinberg, A., Dien, J., & Hajcak, G. (2011). Event-related potential activity in the basal ganglia differentiates rewards from nonrewards: response to commentary. Human Brain Mapping, 32(12), 2267–2269. https://doi.org/10.1002/hbm.21357
Gramfort, A., Papadopoulo, T., Olivi, E., Clerc, M., 2010. OpenMEEG: opensource software for quasistatic bioelectromagnetics. Biomed Eng Online, 9, 45. https://doi.org/10.1186/1475-925X-9-45.
Gunther Moor, B., Crone, E. A., & van der Molen, M. W. (2010). The heartbrake of social rejection: heart rate deceleration in response to unexpected peer rejection. Psychological Science, 21(9), 1326–1333.
Holroyd, C. B., & Coles, M. G. (2002). The neural basis of human error processing: reinforcement learning, dopamine, and the error-related negativity. Psychological Review, 109(4), 679.
Holroyd, C. B., Pakzad-Vaezi, K. L., & Krigolson, O. E. (2008). The feedback correct-related positivity: Sensitivity of the event-related brain potential to unexpected positive feedback. Psychophysiology, 45(5), 688–697.
Holroyd, C. B., HajiHosseini, A., & Baker, T. E. (2012). ERPs and EEG oscillations, best friends forever: comment on Cohen et al. Trends in Cognitive Sciences, 16(4), 192–193. https://doi.org/10.1016/j.tics.2012.02.008
Hu, L., Xiao, P., Zhang, Z. G., Mouraux, A., & Iannetti, G. D. (2014). Single-trial time–frequency analysis of electrocortical signals: Baseline correction and beyond. Neuroimage, 84, 876–887.
Hughes, B. L., Leong, J. K., Shiv, B., & Zaki, J. (2018). Wanting to like: Motivation influences behavioral and neural responses to social feedback. bioRxiv, 300657. https://doi.org/10.1101/300657.
Janssen, D. J., Poljac, E., & Bekkering, H. (2016). Binary sensitivity of theta activity for gain and loss when monitoring parametric prediction errors. Social Cognitive and Affective Neuroscience, 11(8), 1280–1289. https://doi.org/10.1093/scan/nsw033
Joel, S., Plaks, J. E., & MacDonald, G. (2019). Nothing ventured, nothing gained: People anticipate more regret from missed romantic opportunities than from rejection. Journal of Social and Personal Relationships, 36(1), 305–336.
Kappenman, E. S., & Luck, S. J. (2016). Best Practices for Event-Related Potential Research in Clinical Populations. Biological Psychiatry. Cognitive Neuroscience and Neuroimaging, 1(2), 110–115. https://doi.org/10.1016/j.bpsc.2015.11.007
Kross, E., Berman, M. G., Mischel, W., Smith, E. E., & Wager, T. D. (2011). Social rejection shares somatosensory representations with physical pain. Proceedings of the National Academy of Sciences, 108(15), 6270–6275.
Lee, T. W., Girolami, M., & Sejnowski, T. J. (1999). Independent component analysis using an extended infomax algorithm for mixed subgaussian and supergaussian sources. Neural Computation 11(2), 417-441. https://doi.org/10.1162/089976699300016719.
Maris, E., Oostenveld, R., 2007. Nonparametric statistical testing of EEG- and MEGdata. Journal of Neuroscientific Methods 164, 177–190. https://doi.org/10.1016/j.jneumeth.2007.03.024.
Nieuwenhuis, S., Yeung, N., Holroyd, C. B., Schurger, A., & Cohen, J. D. (2004). Sensitivity of electrophysiological activity from medial frontal cortex to utilitarian and performance feedback. Cerebral Cortex, 14(7), 741–747. https://doi.org/10.1093/cercor/bhh034
Nunnally, J. C., Bernstein, I. H., & Berge, J. M. T. (1967). Psychometric theory. McGraw-Hill.
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011). FieldTrip: open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Computational Intelligence and Neuroscience, 2011. https://doi.org/10.1155/2011/156869.
Peterburs, J., Sannemann, L., & Bellebaum, C. (2019). Subjective preferences differentially modulate the processing of rewards gained by own vs. observed choices. Neuropsychologia, 132, 107139. https://doi.org/10.1016/j.neuropsychologia.2019.107139
Peyron, R., Laurent, B., & Garcia-Larrea, L. (2000). Functional imaging of brain responses to pain. A review and meta-analysis (2000). Neurophysiologie Clinique/Clinical Neurophysiology, 30(5), 263–288. https://doi.org/10.1016/s0987-7053(00)00227-6.
Pornpattananangkul, N., & Nusslock, R. (2015). Motivated to win: Relationship between anticipatory and outcome reward-related neural activity. Brain and Cognition, 100, 21–40.
Proudfit, G.H. (2015). The reward positivity: from basic research on reward to a biomarker for depression. Psychophysiology 52(4), 449–459. https://doi.org/10.1111/psyp.1237.
Sassenhagen, J., & Draschkow, D. (2019). Cluster-based permutation tests of MEG/EEG data do not establish significance of effect latency or location. Psychophysiology, 56(6), e13335. https://doi.org/10.1111/psyp.13335
Seminowicz, D. A. , & Moayedi, M. . (2017). The dorsolateral prefrontal cortex in acute and chronic pain. Journal of Pain, 18(9):1027-1035.
Somerville, L. H., Heatherton, T. F., & Kelley, W. M. (2006). Anterior cingulate cortex responds differentially to expectancy violation and social rejection. Nature Neuroscience, 9(8), 1007.
Spielberger, C. D., Gorsuch, R. L., Lushene, R., Vagg, P. R., Jacobs, G. A. (1983). Manual for the state-trait anxiety inventory. Consulting Psychologist Press
Tadel, F., Baillet, S., Mosher, J.C., Pantazis, D., Leahy, R.M. (2011). Brainstorm: a userfriendly application for MEG/EEG analysis. Computational Intelligence and Neuroscience. 2011, 879716. https://doi.org/10.1155/2011/879716.
Threadgill, A. H., Ryan, J., Jordan, C., & Hajcak, G. (2020). The reward positivity: Comparing visual and auditory feedback. Biological Psychology, 154, 107907. https://doi.org/10.1016/j.biopsycho.2020.107907
Van der Molen, M. J.W., Poppelaars, E. S., Van Hartingsveldt, C. T., Harrewijn, A., Gunther Moor, B., & Westenberg, P. M. (2014). Fear of negative evaluation modulates electrocortical and behavioral responses when anticipating social evaluative feedback. Frontiers in Human Neuroscience, 7, 936. https://doi.org/10.1016/j.biopsycho.2018.02.016
Van der Molen, M. J. W., Dekkers, L. M., Westenberg, P. M., van der Veen, F. M., & Van der Molen, M. W. (2017). Why don’t you like me? Midfrontal theta power in response to unexpected peer rejection feedback. NeuroImage, 146, 474–483.
van der Molen, M. J.W., Harrewijn, A., & Westenberg, P. M. (2018). Will they like me? Neural and behavioral responses to social-evaluative peer feedback in socially and non-socially anxious females. Biological Psychology, 135, 18–28. https://doi.org/10.1016/j.biopsycho.2018.02.016.
van der Veen, F. M., Burdzina, A., & Langeslag, S. J. E. (2019). Don’t you want me, baby? Cardiac and Electrocortical Concomitants of Romantic Interest and Rejection: Running Title: Romantic Interest and Rejection. Biological Psychology, 146, 107707. https://doi.org/10.1016/j.biopsycho.2019.05.007.
Warren, C. M., & Holroyd, C. B. (2012). The Impact of Deliberative Strategy Dissociates ERP Components Related to Conflict Processing vs. Reinforcement Learning. Frontiers in Neuroscience, 6, 43. https://doi.org/10.3389/fnins.2012.00043
Watson, D., Clark, L.A., Tellegen, A. (1988). Development and validation of brief measures of positive and negative affect: the PANAS scales. Journal of Personality and Social Psychology, 54(6), 1063.
Yao, M., Lei, Y., Li, P., Ye, Q., Liu, Y., Li, X., & Peng, W. (2019). Shared sensitivity to physical pain and social evaluation. The Journal of Pain, 21(5-6), 677-688.
Acknowledgments
This study was supported by the National Natural Science Foundation of China (31671150), Shenzhen-Hong Kong Institute of Brain Science-Shenzhen Fundamental Research Institutions (2019SHIBS0003), and Shenzhen Basic Research Scheme (JCYJ20150729104249783).
Data and Code Availability Statement
The data and code that support the findings of this study are available from the corresponding author [Hong Li, E-mail: lihongwrm@vip.sina.com] upon reasonable request.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
"The original online version of this article was revised:" plus the same explanatory text of the problem as in the erratum/correction article.
Supplementary Information
ESM 1
(DOCX 1.24 mb)
Rights and permissions
About this article

Cite this article
Zhang, X., van der Molen, M.J.W., Otieno, S.C.S.A. et al. Neural correlates of acceptance and rejection in online speed dating: An electroencephalography study. Cogn Affect Behav Neurosci 22, 145–159 (2022). https://doi.org/10.3758/s13415-021-00939-0
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13415-021-00939-0