
Abstract
Septic syndromes represent a major although largely underrecognized healthcare problem worldwide, accounting for thousands of deaths every year. It is now agreed that sepsis deeply perturbs immune homeostasis by inducing an initial tremendous systemic inflammatory response which is accompanied by an anti-inflammatory process, acting as negative feedback. This compensatory inhibitory response secondly becomes deleterious as nearly all immune functions are compromised. These alterations might be directly responsible for worsening outcome, as they may play a major role in the decreased resistance to nosocomial infections in patients who survived initial resuscitation. Consequently, immunostimulatory therapies may now be assessed for the treatment of sepsis. This review focuses on immune dysfunctions described in septic patients and on their potential use as markers on a routine standardized basis for prediction of adverse outcome or of occurrence of secondary nosocomial infections. This constitutes a prerequisite to a staging system for individualized treatment for these hitherto deadly syndromes.
Similar content being viewed by others

The Old Ceremony
Septic syndromes represent a major, although largely underrecognized healthcare problem worldwide, accounting for thousands of deaths every year. Epidemiological studies estimated the number of cases of sepsis to be 750,000 in 1995 in the USA (resulting in more than 210,000 deaths) and nearly 60,000 in 2001 in France (1,2). Mortality remains high, ranging from 20% for sepsis to >50% for septic shock (1,3,4). More worrisome, a 75% increase in the incidence rate of severe sepsis has been observed over the past two decades, which may be partly explained by the improved care of the elderly and by associated comorbidities in patients [for example, cancer, diabetes (2)]. Therefore, as the population further ages, the incidence of sepsis is projected to increase significantly, leading, for example, to over a million cases of severe sepsis in 2020 in the USA alone (1). Accordingly, a recent study showed that the hospitalization rate for severe sepsis has almost doubled from 1993 to 2003 and that the population-based mortality rate has risen by two-thirds during the same period of time (4).
The provision of apparently adequate fluid resuscitation/goal-directed therapy, specific antibiotics, and low-dose steroids; management of blood glucose levels; aggressive operative intervention; selective use of activated protein C; and almost 20 years of anti-inflammatory clinical trials have had only a modest effect in decreasing mortality. The inability of these therapies to mitigate the devastating effects of this condition indicates that the initial hypotheses for sepsis pathophysiology have been misconstrued or inadequately addressed. Furthermore, treating septic patients as a group despite the extreme heterogeneity characterizing this population constitutes another explanation. In particular, the failure of anti-inflammatory strategies based on the postulate that death after sepsis is solely due to an overwhelming pro-inflammatory immune response is an illustrative example (5,6). Rather, we can now postulate that immune dysfunctions that are supposed to play a role in mortality may be different between a patient succumbing within the very first hour after sepsis and a patient surviving these first critical hours but dying later from a secondary nosocomial infection.
The New Skin
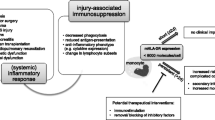
Several lines of evidence have established that death from septic shock may be due to the effect of distinct mechanisms over time. Early in septic shock, a massive release of inflammatory mediators (normally designated to trigger immune response against pathogens) occurs, which may be responsible for organ dysfunction and hypoperfusion (7,8). Concomitantly, the body develops compensatory mechanisms to prevent overwhelming inflammation and dampen an overzealous antiinfectious response (Figure 1). These negative feedback mechanisms, although having protective effects during the first hours, may paradoxically become deleterious as they persist over time. Indeed, considerable clinical and experimental evidence indicates that patients rapidly present with numerous compromised immune functions (7–9). Therefore, as our capacity to treat patients during the very first hour of shock has improved (early and aggressive initial supportive therapy), many patients now survive this critical step but eventually die later in a state of immunosuppression. Indeed, it has been observed that the majority of nonsurviving septic shock patients (>80%) die late after initial resuscitation and present with hallmarks of immunosuppression, whereas patients who survive are those who spontaneously recover immune functions without specific treatments (10). The necessity of an effective immune response to survive septic shock is further illustrated by a recent genomewide study of mRNA expression in patients (11). Among 14,500 genes, we observed that the 19 genes overexpressed in survivors were mainly involved in innate immune response [cytokine, chemokine receptors, effectors of the Toll-like receptor (TLR) pathways].

Simplified description of pro- and anti-inflammatory systemic immune responses over time after septic shock. The dashed lines represent pro- or anti-inflammatory responses; the bold line represents the result at the systemic level. The shift from a pro-inflammatory to an anti-inflammatory immune response predominant at the systemic level likely occurs before 24 h after diagnosis of shock.
These immune alterations are thought to play a major role in impaired clearance of microorganisms. They could explain patients’ difficulty to fight the primary bacterial infection and their decreased resistance to secondary nosocomial infections. As evidence, a proportion of nosocomial infections observed in septic patients are due to commensal organisms, which become pathogenic solely in fragile, immunocompromised hosts (12). Consequently, immunostimulatory therapies constitute an innovative strategy that deserves to be assessed for the treatment of sepsis. The goal is to improve bacterial killing at the primary site of infection and to prevent the development of nosocomial infections and the reactivation of dormant viruses (12). However, despite encouraging preliminary studies (interferon-γ, GM-CSF), we are still far from being able to initiate larger clinical trials. In particular, we need to be able to identify patients beforehand who would actually benefit from immunostimulating therapies. Indeed, in the absence of specific clinical signs of immune status in patients, it is necessary to determine the best biological tools for patients’ stratification according to their immune status (a missing step in most previous clinical trials). This would allow us to define the right action (that is, stimulating innate immunity and/or adaptive immunity, blocking apoptosis, restoring other altered functions) at the right time (early or delayed) in the right patient (individualized/tailored therapy). Also, this would allow us to conduct a close monitoring of patients’ immune and inflammatory responses to the drugs (drug efficacy or drug side effects-for example, reactivation of deleterious inflammatory response). Consequently, more knowledge of sepsis pathophysiology is needed not only if we are to develop better, more effective interventions, but also to identify for whom and when these interventions will be most efficacious.
Although the mechanistic and molecular bases for sepsis-induced immunosuppression are not exhaustively established, several features of the condition have been described, including enhanced leukocyte apoptosis, lymphocyte anergy, and deactivated monocyte functions (7–14). This review focuses on immune dysfunctions described in septic patients and on their potential use as markers on a routine standardized basis for prediction of adverse outcome or occurrence of secondary nosocomial infections.
Functional Testing
Because it directly measures ex vivo the capacity of a cell population to respond to an immune challenge, functional testing theoretically represents the best method to establish the diagnosis of immunoparalysis. Two hallmarks of sepsis are regularly reported: decreased monocyte capacity to release proinflammatory cytokines in response to lipopolysaccharide (LPS) challenge and lowered lymphocyte proliferation in response to antigens (tuberculin, tetanus toxin) or nonspecific (phytohemagglutinin, concanavalin A, anti-CD3, anti-CD28 antibodies) stimulations (15–17,21–28).
Lymphocyte anergy is illustrated by the loss of delayed-type hypersensitivity reaction to skin test recall antigens in patients. This has been described for a long time and is known to be associated with mortality and with the development of secondary infections (18–20). Indeed, a marked decrease in lymphocyte proliferation has been described in patients after severe injuries (sepsis, major surgery, severe burn or trauma) (15–17,21–28). This was associated with diminished production of cytokines such as interleukin (IL)-2 and γ-interferon (IFN-γ) by peripheral blood mononuclear cells (15,29,30). Manjuck et al. (16) observed that decreased proliferative response to tetanus toxin in severe septic patients was associated with diminished expression of monocyte HLA-DR and CD86 and of lymphocyte CD28 (all activator coreceptors during the process of antigen presentation), whereas expression of cytotoxic T lymphocyte antigen (CTLA)-4 (an inhibitory coreceptor) was increased on lymphocytes. Similarly, recent data in trauma correlated diminished lymphocyte proliferation with increased expression of inhibitory coreceptors on lymphocytes (28). Most important, it has been observed that the severity of this state of anergy correlated with poor outcome, increased occurrence of infectious complications, and subsequent multiorgan failure (16,21–28,31).
Usually performed with peripheral blood mononuclear cells (that is, lymphocytes and monocytes), the measurement of cell proliferation in vitro investigates both the capacity of lymphocytes to proliferate and of monocytes to present antigens (when performed with recall antigens) in a single test. Proliferation is usually assessed by [3H]thymidine uptake or, more recently, by the use of fluorescent probes [such as carboxyfluorescein diacetate succinimidyl ester (CFSE)]. However, as proliferation tests require long incubation time (two to three days for mitogens, five to seven days for recall antigens), they are not suitable for clinical decision-making and thus are not performed on a routine basis.
Concomitantly, many groups have investigated the capacity of patient cells (mainly antigen-presenting cells/monocytes) to release proinflammatory cytokines in response to LPS, other TLR agonists, or whole bacteria in vitro (see recent review in 32). These tests represent reliable methods to assess the phenomenon of endotoxin tolerance defined as a reduced responsiveness to a secondary LPS challenge following a first inflammatory response. Monocytes from septic patients usually present with a diminished capacity to release tumor necrosis factor (TNF)-α, IL-1α, IL-6, and IL-12, whereas the release of anti-inflammatory mediators (IL-1ra, IL-10) is not affected or even slightly increased (33–38). This shows that LPS can still activate monocytes but that the intracellular signaling has been turned to favor production of anti-inflammatory molecules, therefore supporting the concept of leukocyte reprogramming (39,40). Of note, cytokine measurements have usually been performed using homemade protocols, therefore leading to poor reproducibility. However, some results are now obtained with commercially available kits, allowing for better standardization (41,42). Still, large multicenter clinical trials are required to assess this test with regard to outcome and compare it with other markers of monocyte dysfunction such as reduced monocyte HLA-DR expression (mHLA-DR, see below).
In summary, although not suitable for routine monitoring and clinical decision-making, these functional tests remain essential to obtain further insights to the pathophysiology of sepsis-induced immunosuppression and assess the validity of surrogate markers, more easily measurable and standardizable.
Measurement of Circulating Mediators
Despite a large number of studies focusing on the measurement of plasmatic concentrations of various mediators, none of them has been consensually accepted. This might be explained by the complexity of the interactions between the numerous networks and amplification cascades involved in sepsis pathophysiology. Indeed, hundreds of mediators are likely to participate in sepsis pathophysiology, and their concentrations vary rapidly over time (43). Numerous correlations have been observed between the increased concentration of one particular circulating mediator and adverse outcome, but none of them is routinely used for patient staging and monitoring. This is especially true for pro-inflammatory cytokines. TNF-α, IL-6, and IL-8 have been extensively investigated during the last two decades without providing clear-cut results. An alternative strategy might be to stratify patients on the basis of decreased levels (instead of increased levels) of these proinflammatory mediators, which might subsequently be proposed as substitutive therapy. An example would be the study of circulating levels of GM-CSF. It has been documented that plasma concentrations of GM-CSF after sepsis were significantly depressed in nonsurvivors in comparison with survivors, whose levels were similar to healthy donors (44). This therefore provides a rationale for starting therapies aimed at restoring GM-CSF concentration.
Regarding pro-inflammatory cytokines, high mobility group box 1 (HMGB1) deserves to be specially investigated, as it represents a delayed/late mediator in sepsis (45). Its concentration continues to be elevated while other proinflammatory cytokines such as TNF-α or IL-6 have returned to normal (46,47). Moreover, HMGB1 may interact with other aspects of sepsis pathophysiology, including apoptosis, dysfunctional dendritic cells (DCs), maturation, or chemotaxis (48). It has also been shown that its persistent increase during sepsis is associated with unfavorable outcome (47,49). It would therefore be interesting to assess its concentration in a large cohort of septic patients.
On the other hand, in the particular context of immunosuppression, measuring persistently increased concentrations of anti-inflammatory mediators may be an indicator for the initiation of immunestimulating therapy. Lots of candidates have been proposed, but the most consistent data concerns IL-10. Gogos et al. (50) showed that IL-10 and IL-10/TNF-α ratios, among a panel of various cytokines (Il-1, IL-6, sTNF-RI and II), were the more powerful predictors of mortality in patients with severe sepsis both at admission and 48 h later. Van Dissel et al. (51) reported similar results: they observed that a high IL-10/TNF-α ratio was associated with unfavorable outcome in a group of 450 febrile patients. Accordingly, in critically ill septic patients at admission, higher values of IL-10 were measured in nonsurvivors and provided better prognostic information than IL-4 (52). An anti-inflammatory profile was also found to be associated with severity in children with meningococcal sepsis (53). We extended these results by illustrating that IL-10 levels remained higher in nonsurvivors until 15 days after the onset of septic shock in a group of 38 patients (54). In parallel, neither TNF-α nor transforming growth factor (TGF)-β values were able to differentiate survivors from nonsurvivors (54). In addition, we observed that IL-10 was the sole cytokine to correlate with mHLA-DR values. In particular, high IL-10 concentrations at the beginning of shock were negatively correlated with the nadir of mHLA-DR measurements during the 2 weeks of monitoring (54). Consequently, this initial value of IL-10 may reflect the severity of the forthcoming immunoparalysis. Similar results have been observed with IL-10 mRNA measurements in comparable cohorts of patients (49,55,56). Very recently, Leonidou et al. (57) confirmed these results. They observed that high IL-10 levels were associated with mortality after severe sepsis and, importantly, with stress-induced hyperglycemia, whereas TNF-α, IL-6, and TGF-β concentrations did not have any predictive value. Given the properties of IL-10 in suppressing the synthesis of numerous proinflammatory cytokines (59), its continued release may contribute to the immune dysfunctions observed after septic shock and thus may augment susceptibility to secondary microbial invasion (58–60).
Along with IL-10, TGF-β constitutes a potent immunosuppressive cytokine often described as being implicated in immunoparalysis (9) and as playing a role in monocyte deactivation during endotoxin tolerance (61). In numerous clinical studies, however, its plasma levels were found to be in normal range (or slightly above), and no difference has been observed between survivors and nonsurvivors (30,54,56,57,62). These results suggest that IL-10 might be more important than TGF-β in the pathophysiology of sepsis-induced immunoparalysis. This is in accordance with experimental data showing that blocking IL-10 is more efficient than blocking TGF-β in reversing endotoxin tolerance (59,61,62).
Many other mediators are known to be elevated during septic shock and to possess immunosuppressive properties; we can mention soluble HLA-G molecules, prostaglandin E2, α-melanocytestimulating hormone, adrenomedullin, and vaso-intestinal peptide (63–67). There is also evidence for a direct interaction between nervous, endocrine, and immune systems. Numerous mediators participating in this network and increased during sepsis behave as immunosuppressors: cortisol, norepinephrine, acetylcholine (66–70). However, they all need to be assessed in large clinical studies.
It should be noted that measuring concentrations of a single mediator (or a few mediators) for establishing patients’ immune status remains questionable because it provides only a partial view of the disorders. In many cases, determining what force dominates (pro- vs anti-inflammatory responses) on the basis of serum measurements will remain nearly impossible, because both responses are enhanced during septic shock. An alternative would be to use a panel of markers rather than a single parameter, allowing a better definition of individualized pro-/anti-inflammatory profiles (49,71–73).
Cellular Monitoring
Antigen-Presenting Cells: Dendritic Cells
Preliminary results in mice suggest the importance of dendritic cell (DC) dysfunction and apoptosis during sepsis. DCs are not only affected/killed by the systemic inflammatory response but may also contribute to the development of immune suppression during sepsis by impaired functions such as reduced IL-12 production or decreased capacity to activate T cells (74). Accordingly, in several murine studies, secondary infections and mortality have been reversed by administration of DCs purified from naive animals (75) or fms-like tyrosine kinase 3 lig-and (FLT3-L, a DC growth factor) (76,77).
In humans, Guisset et al. (78) have observed a reduced DC blood count in septic patients. Interestingly, this loss was more important in patients who died than in survivors. The number of circulating DCs was also inversely correlated with severity scores such as SAPS II (78). Ho et al. (79) monitored blood DCs in 56 patients before and after surgery. They observed that the number of DCs increased acutely in response to the stress of surgery but dropped below preoperative levels on days 2 to 3. In a postmortem study of spleen from 26 septic patients and 20 trauma patients, Hotchkiss et al. (80) observed that sepsis causes a dramatic reduction in the percentage area of spleen occupied by follicular DCs and that the number of MHC II+ interdigitating DCs was decreased in septic patients compared with trauma patients. Regarding DC functions, Faivre et al. (81) investigated the ability of monocytes to differentiate into functional DCs in vitro in patients undergoing surgery for peritonitis. They observed that, although the ability of patients’ deactivated monocytes to differentiate into mature DCs was faster than in healthy individuals, these DCs were unable to increase T-cell response. De et al. (82) observed that in trauma patients, monocytes with reduced IL-12 production and decreased ability to induce T-cell proliferation also had decreased capacity to differentiate into DCs in vitro. Overall, these studies suggest that, as observed in murine models of sepsis, human DCs may be not only decreased in blood after sepsis but also dysfunctional, which may therefore play a pivotal role in the development of sepsisinduced immunosuppression. Consequently, the monitoring and potential reversal of DC dysfunctions may represent an innovative therapeutic strategy for the treatment of sepsis.
Antigen-Presenting Cells: Monocytes
Monocytes from septic patients are characterized not only by decreased capacity to mount a pro-inflammatory reaction upon secondary bacterial challenge (as previously discussed), but also by impairment in antigen presentation, likely due to the lowered expression of major histocompatibility (MHC) class II molecules. Indeed, in septic patients, decreased cell-surface expression of HLA-DR molecules has regularly been observed on circulating monocytes. As opposed to circulating mediators, the major advantage of measuring a cellsurface marker such as mHLA-DR is that its level of expression is a result of the sum of the effects of multiple mediators, all of which are potentially regulated during septic shock. For example, mHLA-DR expression has been shown to be positively and negatively regulated by cytokines such as IFN-γ and IL-10, as well as by corticoids and catecholamines (83–86). There is general consensus that diminished mHLA-DR expression may be a reliable marker for immunosuppression in critically ill patients (47,87,88).
mHLA-DR Expression and Functional Parameters
Importantly, several studies have demonstrated the association of low mHLA-DR expression and impairment of various monocytic functions. Monocytes from septic patients with decreased mHLA-DR produced low amounts of TNF-α and IL-1 in response to bacterial challenges, including LPS and SEB or PMA stimulations (62). Similarly, lymphocytes from septic patients with low mHLA-DR have been shown to be unable to proliferate in response to tetanus toxin as a consequence of a failure in antigen presentation (16). Accordingly, Wolk et al. (89), in an experimental model of endotoxin tolerance, reproduced the decrease of mHLA-DR and showed that it is associated with impairment in T-cell proliferation and IFN-γ production in response to recall antigens. In a series of nine consecutive septic patients with immunosuppression, the administration of GM-CSF induced sustained mHLA-DR recovery accompanied by a restoration in ex vivo TNF-α production after LPS challenge (41). Recently, in surgical patients, Schneider et al. (90) observed that G-CSF-induced restoration of mHLA-DR was accompanied not only by increased lymphocyte proliferation and Th1 cytokine production (IL-2 and IFN-γ) in response to PHA but also by a better capacity to release inflammatory cytokines in a whole-blood model after LPS challenge. Finally and intriguingly, it has been proposed that the level of cell-surface MHC class II molecules directly modulates the degree of inflammatory response to LPS independently of CD14 and TLR4 equipment. In comparison with HLA-DR-positive cells, HLA-DR-negative cells secreted markedly reduced amounts of TNF-α and IL-8 (91). Overall, these data demonstrate that diminished mHLA-DR is a surrogate marker for monocyte dysfunction. Importantly, the restoration of normal mHLA-DR levels is associated with the recovery of monocyte functions.
mHLA-DR Expression and Clinical Study
Up to now, the decrease of mHLA-DR expression has mainly been assessed as a predictor of septic complications after trauma, surgery, or pancreatitis (28,92–98). In these studies, low levels of mHLA-DR (<40% of positive monocytes, normal >90%) were observed in patients who subsequently developed nosocomial infections. In contrast, in injured patients with uneventful recovery, mHLA-DR rapidly returned to normal values (in general, <1 week). In the work of Allen et al. (99), diminished mHLA-DR expression after cardiac surgery was found to be an independent predictor of septic complications after correction for usual clinical parameters in a multiple logistic regression analysis. Results in burn patients also indicate that low mHLA-DR expression is associated with secondary septic events (100,101). Similar results have been reported in septic shock. Patients who developed secondary infections exhibited persistent decrease of mHLA-DR expression (68). Our unpublished data obtained from >200 septic shock patients indicate that, seven days after the onset of shock, patients with mHLA-DR <40% will later develop nosocomial infections.
On the other hand, low mHLA-DR has also been shown to be predictive of adverse outcome in different groups of critically ill patients (97,102,103). We recently observed that burn patients rapidly presented with decreased mHLA-DR expression in the first day after burn, and that this expression subsequently increased in patients who survived but remained low in nonsurvivors (101). Regarding severe sepsis and septic shock, the predictive value for outcome of decreased mHLA-DR remains somewhat conflicting. Although the decrease of mHLA-DR is generally well accepted, its correlation with mortality is not reported by every author. Perry et al. (104), Muller Kobold et al. (105), and Fumeaux and Pugin (106) did not observe any correlation, whereas Docke et al. (107), Saenz et al. (108), Hynninen et al. (52), Lekkou et al. (56), Monneret et al. (109), and Le Tulzo et al. (68) observed lower mHLA-DR in patients who died. However, those preliminary studies included a very limited number of patients and narrowed, in general, the monitoring at the first 48 h after onset of disease. Importantly, we observed that the difference in mHLA-DR values between survivors and nonsurvivors became significant only 48 h after the onset of shock (110). In 93 consecutive septic shock patients, we observed that, although mHLA-DR values were not significantly different between groups during the first 48 h, persisting low mHLA-DR values (<30% of positive monocytes at day 3 to 4 after ICU admission) identified a population of patients with a high mortality rate. Multivariate logistic regression analysis revealed that low mHLA-DR (<30%) at day 3 to 4 remained independently associated with mortality after adjustment for usual clinical confounders. Decreased mHLA-DR was even found to be a better predictor of mortality than SAPS II and SOFA scores (110). In parallel, we demonstrated that the loss of mHLA-DR expression was due to a global regulation at the transcriptional level of all MHC class II restricted genes and that the magnitude of this regulation affected clinical outcome (111).
Cutoff values, however, remain to be precisely defined regarding specific clinical context (they may be different depending on the type of injury) and elapsed time after the onset of sepsis/ injury. Therefore, multicenter clinical trials need to be designed with appropriate numbers of patients prospectively selected on the basis of well-defined criteria and ultimately classified on the basis of well-defined outcomes (87). A prerequisite for all centers performing mHLA-DR measurements should also be the standardization of the different protocols. For that purpose, a standardized test has recently been proposed. It is based on the use of calibrated beads allowing the conversion fluorescence intensity to number of antibodies fixed per cell, and is therefore indicative of the number of HLA-DR molecules per monocyte. Using this method, results become comparable between different laboratories (112). That said, the standardized use of an isotype control for determining the percentage of positive cells remains a robust and reproducible method when conducted appropriately. Ideally, in ensuing clinical studies, these two modes of expression of mHLA-DR (percentages versus number of HLA-DR molecules per cell) should be compared regarding their respective value as prognostic markers.
Other molecules have been demonstrated to be downregulated on the monocyte surface during sepsis: CD14, CD86, GM-CSF receptor, and CX3CR1 (113–118). Their decreased expression was correlated with mortality. Finally, another field of investigation concerns the respective roles of the numerous mechanisms that negatively regulate TLR-associated signaling pathways (for example, IRAK-M, SIGIRR, Tollip, SOCS, and MyD88s) in clinical samples (40). This might reveal both diagnostic markers and new therapeutic targets.
Studies of T-Lymphocyte Subsets
Because of their ability to interact not only with cells of the innate immune system but also with other cells of the adaptive response, T lymphocytes play a central role in the anti-infectious immune response, as both effectors and regulators of this response. This has been illustrated by the description of increased mortality, decreased bacterial clearance, and altered pro-inflammatory immune response after polymicrobial septic challenge in mice lacking both T and B cells (119,120). A growing body of evidence has now confirmed that the lymphocyte-mediated immune response may be dysfunctional after severe sepsis and may play a major role in the development of a state of immunoparalysis in patients.
Besides the functional defects that have been previously described, a marked decrease in the number of circulating lymphocytes has been observed in patients after sepsis and severe trauma (Table 1) (17,20,120–122). Most importantly, the extent of this lymphopenia has been correlated with the development of nosocomial infections in those patients (20,122).
During the phase of immunoparalysis, T lymphocytes have also been characterized by a shift toward Th2-type immune response. Several studies have observed that mononuclear cells from injured patients present with reduced levels of Th1 cytokines (including TNF-α, INF-γ, and IL-2) but increased levels of Th2 cytokines (mainly IL-4) and that the reversal of the Th2 response improves survival among patients with sepsis (7,123). This has been mainly demonstrated by the measurement of cytokine production by ELISA or flow cytometry after in vitro cell stimulation. With similar approaches, however, some authors instead observed global downregulation of Th1 and Th2 responses in patients after sepsis or severe trauma (24,25,29,30,124). We observed similar results based on the measurements of Th2 cell-surface marker (CRTH2) (125) or Th1/Th2 transcription factor mRNA by qRT-PCR (49). Interestingly, in those patients, mRNA expression of FOXP3 (specific transcription factor for regulatory T cells) was not affected, supporting the idea that regulatory functions are maintained during sepsis (or even increased) while effector responses are almost totally downregulated (126).
Regarding their phenotype, T lymphocytes have been characterized by the overexpression of inhibitory coreceptors during the phase of immunoparalysis. It has recently been demonstrated in trauma patients that anergic T cells had increased PD-1, CD47, and CTLA4 expression that would facilitate preferential triggering of negative signaling pathways during T-cell stimulation (28) and therefore lead to lymphocyte anergy. Moreover, this increase in corepressor receptors (in particular CTLA-4) appears to be associated with a decrease in the expression of co-activator receptors such as CD28 (16) or CD3 (127), which could also play a role in the development of immunoparalysis (128).
Finally, one likely major characteristic of T lymphocyte dysfunction after severe injury is the increase among patients’ circulating lymphocytes of a cell population with known regulatory properties. CD4+CD25+ regulatory T lymphocytes (Treg) have recently been reported to be a potent regulatory T-cell lineage, playing an essential role in the control of both adaptive and innate immune responses (129). An increase in the percentage of Treg has been described in septic shock patients (14). Importantly, this increase was observed immediately after the diagnosis of sepsis but persisted only in nonsurviving patients in association with augmented CTLA4 expression. These results were further extended by the observation that this relative increase was in fact due to a decrease of CD4+CD25− circulating T lymphocytes (Treg counterparts) and not so much to a change in the absolute Treg count in patients (126). This suggests a lower sensitivity of these cells to the apoptotic/ death mechanisms occurring after severe injury, as illustrated by the decreased cell number of every lymphocyte lineage but Treg in the circulating blood of septic patients (Table 1). Subsequently, a similar increase of Treg percentage has been observed in trauma patients (130) and in mice after polymicrobial septic challenge and stroke (131–134). In total, even if the actual role of these cells in the immune dysfunction occurring after severe injury remains to be demonstrated (inconclusive results of functional studies in murine models), based on their potent regulatory properties one could postulate a role for Treg in the development of lymphocyte anergy after severe injury. This hypothesis seems to be confirmed by our personal observations of a strong correlation between increased Treg/ effector ratio measured in whole blood after septic shock and decreased proliferative response of lymphocytes after mitogenic stimulation.
Regarding other populations of regulatory lymphocytes, a role for γδ T cells as well as natural killer (NK) T cells has been proposed in the development of immune dysfunction after severe injury mainly on the basis of animal models (135,136). These results have been in part confirmed by a clinical study demonstrating the decreased percentage of γδ T cells in septic shock patients (127). However, the preferential tissue location of those lymphocyte subpopulations and their low percentage in the circulating blood render their study in the human setting technically difficult and therefore their involvement in injury-induced immune dysfunction hard to demonstrate.
In total, there is compelling evidence that septic patients present with dysfunctions of their adaptive and innate immune responses, which might play a role in morbidity and mortality after severe injury.
Monitoring of Apoptosis
Increased apoptosis of immune effector cells has been shown to be a hallmark of sepsis, and a number of studies have suggested that this dysregulated process may contribute to the immune dysfunction and multiple organ failure observed during this deadly disease. Both the death-receptor and the mitochondrial pathways are likely to be involved in sepsis-induced apoptosis, as well as numerous inducers including steroids, cytokines (TNF-α, HMGB1), Fas ligand, heat shock proteins, oxygen radicals, nitric oxide, and some specific T lymphocytes (13,120,137,138).
Pioneering autopsy studies by Hotchkiss and colleagues (80,139,140) disclosed a profound, progressive, apoptosis-induced loss of cells of the adaptive immune system in the spleen, blood, and gut-associated lymphoid tissue of adults who had died of sepsis. Although no loss of CD8+ T cells or NK cells occurred, sepsis markedly decreased the level of B and CD4+ T cells. This loss was especially important because it occurred during the life-threatening infectious process, while clonal expansion of these cells might have been expected (7). Subsequent autopsy studies of pediatric and neonatal patients who died of sepsis have confirmed this profound apoptosis-induced depletion of CD4+ T cells and B cells (122,141). Accordingly, Le Tulzo et al. (142) observed a marked increase in apoptosis of circulating lymphocytes from septic shock patients compared with critically ill patients without sepsis and healthy volunteers. This induced a profound and persistent lymphopenia associated with poor outcome. Bilbault et al. (143) observed a severe downregulation of the expression of the anti-apoptotic gene BCL2 in circulating mononuclear cells from patients with severe sepsis. This was associated with reduced T-cell count and increased annexin V labeling. Most importantly, immediately after the onset of severe sepsis, this decrease was higher in nonsurvivors than survivors. A second study by this group confirmed these results by measuring the Bax/Bcl-xl and Bax/Bcl-2 ratios in septic shock patients (144).
Similarly, DCs appear to be apoptotic in spleens of septic patients (79). Regarding monocytes/macrophages, Williams et al. observed an increased apoptosis of circulating monocytes in septic patients (145). Adrie et al. found that septic patients exhibited an increased percentage of monocytes with a depolarized mitochondria (as a marker of apoptosis) compared with healthy individuals (146). Furthermore, among septic patients, this percentage was significantly higher in nonsurvivors than survivors. However, one limitation to appreciate the incidence of monocytes/macrophage apoptosis is that some of these changes may represent an increased role in clearance of apoptotic cells, which may make these cells look overtly more apoptotic as a result of handling a greater amount of apoptotic material (147). That said, the downmodulation of expression of CD14 (a cell-surface marker decreased during monocyte apoptosis) on monocytes from septic shock patients tends to confirm this increased apoptotic process, especially because a higher decrease was observed in patients who did not survive (114).
Paradoxically, neutrophil apoptosis is delayed in sepsis (148). Neutrophils are constitutively apoptotic in healthy individuals, and this program is normally activated within hours of maturation and release from marrow stores. As a result, the septic patient has an increased number of activated neutrophils, essential for pathogen eradication but also implicated in organ injury because of their excessive release of damaging proteases (148). The perplexing role of neutrophils in sepsis pathophysiology remains to be fully investigated.
Finally, increased apoptosis has also been observed in endothelial and epithelial cells. The hypothesis of endothelial cell apoptosis after sepsis is based on the observation that higher levels of “shed” endothelial cells were detected in the blood of septic shock patients (149). The M30 neoantigen, which is a product of cytokeratin 18 caspase cleavage and release, can be detected via ELISA during the process of epithelial cell apoptosis. Serum levels of M30 antigen are significantly increased in septic patients in comparison with noninfected trauma patients or healthy controls (150). Moreover, nonsurviving patients exhibit a significantly higher increase in M30 compared with surviving patients or healthy volunteers (150). Although it is strongly suspected that these cells are dysfunctional in septic patients, clear-cut documentation in the in vivo setting is difficult to obtain.
Aside from these cellular measurements of apoptosis, the ability of septic or severely injured patients’ serum to induce apoptosis has been largely described on various cell culture populations (151–153). Accordingly, several groups observed increased serum levels of proapoptotic factors such as sFas or sFasL in septic and critically ill patients (17,154,155). Importantly, these markers exhibited direct correlation with the occurrence of multiple organ failure, and serum levels were higher in septic patients who died compared with survivors.
In conclusion, there is no doubt that sepsis induces extensive apoptosis of most immune cells. This may play a major role in sepsis-induced immune dysfunction and in patients’ altered capacity to clear microorganisms. This deleterious role of apoptosis has been largely demonstrated in numerous murine experiments in which its prevention improved the likelihood of survival. Several inhibitory strategies have been applied, which may point to novel therapeutic approaches in the clinic: caspase inhibitors, antiretroviral protease inhibitors, Fas targeting siRNA, Bcl-2 overexpression, CD95 inhibition (13,138,156,157). Most importantly, the degree of apoptosis of circulating lymphocytes correlates with sepsis severity and predicts fatal outcome, suggesting the importance of its measurement as a prognostic biomarker to help in stratifying patients. However, a major limitation may be the drawbacks inherent in this type of experiment, such as the necessity of rapid processing of the samples (especially regarding annexin V staining), hardly compatible with their use in ICUs (158). Furthermore, as methods used for studying apoptosis may often have a significant rate of false-positive results (especially the deoxyuridine triphosphate nick-end labeling assay), it is recommended to establish apoptosis on the basis of two or more methods of detection among DNA-hypoploidy, morphology, DNA laddering, annexin V staining, active caspase-3, or mitochondrial permeability measurements (13).
Toward a Genomic Approach
It is likely that determining a single parameter, as good as it may be, won’t be sufficient to characterize the complexity of septic patients’ immunological status, which rapidly changes over time. Therefore, the introduction of high-throughput technologies, such as oligonucleotide microarrays, represents an emerging opportunity to open up new dimensions in sepsis research. DNA microarrays allow genomewide assessment of changes in mRNA abundance, therefore usable to monitor changes in gene expression. Although the field that has reaped the most important benefit from the postgenomic era is cancer diagnostics, several recent microarray-based genomewide expression studies have contributed to the description of sepsis-induced host response (159,160). Initially, numerous studies examined the cellular response to different microbial pathogens or pathogen-associated molecular patterns (PAMPs) in vitro. Changes in gene expression after stimulation have been experimentally studied in human circulating leukocytes (161), DCs (162,163), and differentiated human macrophages (164) and in several animal models (165–167). Recent studies took advantage of this approach to assess whether it could successfully be applied to the differentiation between sterile systemic inflammation and sepsis in ICU patients (168–170).
Regarding outcome prediction and monitoring, several studies conducted in patients after surgery (171) or severe trauma (172–174) have shown promising results. We addressed this question in septic shock patients monitored after the early pro-inflammatory reaction, when the patient’s immune functions are known to be severely depressed (>48 h after the onset of shock) (11). Stratifying the results according to the outcome, we identified a set of 28 genes whose systemic transcriptional expression discriminated survivors with a sensitivity and specificity of 100% and 88%, respectively. Numerous genes in this list encoded for mediators of pathogen-recognition pathways, pro-inflammatory response, and cell-migration processes, supporting the hypothesis of the occurrence of better recovery of immune function in patients who survived. Among the genes upregulated in survivors, the chemokine receptor CX3CR1 showed the highest factor of change (≈8-fold increase in comparison with nonsurvivors). Additional investigations showed a persistent decrease of systemic CX3CR1 expression in monocytes from septic shock patients at both the mRNA and protein levels (118). Importantly, CX3CR1 downmodulation was significantly more pronounced in non-survivors. It might directly impair monocyte recruitment in tissue and contribute to patients’ inability to kill invading microorganisms (175). This illustrates the potential of a global transcriptional analysis to identify new molecules that may play a role in sepsis pathophysiology and may be used as prognostic markers. Similarly, Wong et al. (176) investigated the transcriptional response during the early phase of septic shock (<48 h) in children. Their data indicated that patients had decreased systemic expression of a large number of genes that either were dependent or played a direct role in zinc homeostasis. Among genes differently expressed between survivors and nonsurvivors, two isoforms of metallothionein-known to sequester zinc in the intracellular compartment-demonstrated increased expression in the nonsurvivors. Interestingly, these results have been recently extended by the analysis of additional data obtained 48 h after the first time point (177). This second study revealed time-dependent regulation of gene networks primarily related to immunity and inflammation and confirmed the persistent downregulation of genes related to zinc homeostasis.
Despite discrepancies in the very first transcriptome studies (due to differences in specific technological aspects), gene expression profiling is now recognized as providing meaningful data when conducted with rigorously validated and standardized methodologies. A recent study comparing data obtained from the latest, state-of-the-art platforms showed minimal technical variation and good correlation with qRT-PCR, the gold standard for mRNA measurement (178). Similarly, recent comparative cross-laboratory studies gave very promising data (179,180). Thus, because of patients’ heterogeneity and of the high complexity of sepsis pathophysiology, much will be gained from the next multicenter studies monitoring the dynamic behavior of gene expression over time (181,182).
The identification of polymorphic genome variations that may influence susceptibility to sepsis and/or alter its outcome represents another challenge (183). Although numerous studies have observed that single nucleotide polymorphisms (SNPs) in genes involved in sepsis response are associated with increased susceptibility to disease/adverse outcome, results were often inconsistent. This might be explained not only by methodological limitations, in terms of both study design and genotyping methods (184), but also by the complexity of sepsis pathophysiology. This also suggests that a panel of SNPs will probably be more efficient in characterizing the genetic background of the disease. The development of genomewide human SNP genotyping microarrays featuring thousands of genetic markers represents an emerging opportunity to address this question.
Future Therapeutic Strategies and Conclusion
Our understanding of the pathogenesis of sepsis has been oversimplified during the past decades and, as a result, many clinical trials addressed the pro-inflammatory side when there was no evidence that hyperinflammation was dominant in patients. Some issues remain before we gain a complete picture of events leading to immunosuppression: Are the major sepsis-induced inhibitory mechanisms all established? What is the physician-caused part of immunosuppression, because sedatives, catecholamines, insulinall immunosuppressive-are administered to the patient? Is the cellular energetic status crucial in maintaining immune functions? How important is the neuroendocrine-mediated part of immunosuppression? How preponderant is immune failure among other organ failures? Nonetheless, we can reasonably state that patients with sepsis present with features consistent with immunoparalysis. Consequently, stimulating the patient’s immune system may become a promising therapeutic strategy. As presented in this review, we delineate four types of therapeutic interventions: blocking soluble anti-inflammatory mediators, restoring antigen-presenting cell function, restoring T lymphocyte function, and blocking apoptosis (Table 2). In order not to repeat the mistakes from the past, we should keep in mind that targeting a single mediator/function among many immune dysfunctions may be inefficient.
Regarding an eventual blockade of IL-10 function, we presently lack enough consistent data in the human setting, and studies in animal models have provided contrasting results. AS101, with the capacity to inhibit IL-10, has been demonstrated to increase survival in septic mice (185), whereas anti-IL-10 antibodies did not improve bacterial clearance and mortality in murine models (186). That said, we may speculate that blocking a single mediator in a context where many inhibitory pathways are involved/activated, would remain inefficient (as was targeting a single pro-inflammatory mediator during the early cytokine cascade in septic shock). However, AS101 might still be interesting because it acts through different mechanisms (inhibition of IL-10, activation of macrophage functions, inhibition of IL-1β converting enzyme). Further investigations are required.
Several molecules (IFN-γ, G-CSF, GM-CSF) have been used to stimulate monocyte functions and gave interesting preliminary ex vivo results (increase in mHLA-DR expression, restoration of cytokine production). Several prospective randomized multicenter trials using IFN-γ have been conducted in trauma patients. However, despite interesting results regarding secondary end points in some subgroups of patients (decreased severity in nosocomial infections, decreased mortality in infected patients), they remained inconclusive regarding overall mortality or infection rates (187,188). Presneill et al. (189) presented preliminary GM-CSF data in 10 patients with sepsis-induced respiratory failure. They observed modest improvement in gas exchange, ARDS resolution, and alveolar leukocyte phagocytic functions, but it was not accompanied by enhanced survival. In a prospective, randomized, placebo-controlled trial, Rosenbloom et al. (190) investigated whether GM-CSF treatment can improve leukocyte functions and mortality in 40 septic patients. They observed a higher leukocyte count, increased mHLA-DR, and better cure/ improvement of infection in the treated group but no difference in mortality. Nevertheless, it should be noted that these trials were designed without patient stratification, whereas drug efficacy should be assessed only in patients with established impairment in monocyte function. To our knowledge, only two studies stratified septic and trauma patients with respect to mHLA-DR. Because of the small number of patients included, it is not yet possible to reach conclusions on the impact of IFN-γ treatment in septic patients, but promising results have shown a decrease in mortality and/or rate of nosocomial infection (107,191). A randomized, double-blind, placebo-controlled phase II trial using GM-CSF has just been completed in patients with severe sepsis and septic shock; results are still not available (https://doi.org/www.clinicaltrials.gov/ct2/show/NCT0 0252915). Given the central role of DCs in both innate and adaptive immune responses, increasing their number and restoring their function might constitute another potential treatment. In particular, the use of FLT3-L-known to activate and expand DCs-was shown to reverse immunoparalysis in a mouse model of endotoxin tolerance (77) and to increase resistance to P. aeruginosa opportunistic infections in burned mice (76). In human patients, phase II/III trials have been conducted in cancer patients, but not yet in sepsis. Interestingly, GM-CSF and FLT3-L have synergistic effects on the DC maturation process. Activation of TLRs may also allow upregulation of specific antimicrobial defenses; this approach is currently under investigation (192).
Augmenting T-cell function and fighting lymphopenia may represent another therapeutic strategy. For example, IL-7 is an essential cytokine for T-lymphocyte development, survival, expansion, and maturation in humans. Phase I clinical trials in cancer patients and HIV-infected patients have shown that T-cell expansion can be achieved at doses that are well tolerated (193,194). The use of ligands of co-activator receptors for effector T lymphocytes may also have beneficial effects. As an example, recent results by Scumpia et al. (128) have shown that anti-GITR agonistic antibodies were able to restore lymphocyte proliferation, prevent CD3 downmodulation, decrease bacteremia, and increase survival in a mouse model of sepsis. Intravenous use of immunoglobin therapy has also been proposed as an adjuvant treatment for sepsis, but its benefits remain unclear. The authors of recent meta-analyses recommend conducting larger clinical trials with patient stratification (195,196).
Finally, strategies designed at blocking apoptosis, including caspase inhibitors, overexpression of Bcl-2, and inhibition of Fas/FasL signaling, have demonstrated survival improvement in animal models of sepsis as well (13,138,156,157). That said, no therapeutic strategy is sufficiently developed for clinical use. An alternative might be provided by HIV protease inhibitors, whose activity is partly mediate through anti-apoptotic effects. Administration of ritonavir improved survival in a murine model of sepsis, even when given after the onset of the disease (197). As these protease inhibitors are well toler ated in patients, we may expect enticing possibilities in sepsis. A phase I trial is currently underway, investigating in healthy volunteers the effects of these drugs in boosting the immune system (https://doi.org/www.clinicaltrials.gov/ct2/show/NCT00346619). Of note, drugs aimed at blocking apoptosis may be used as adjunctive agents in association with molecules targeting monocytes or leukocytes.
In summary, both arms of immunity, innate and adaptive, are severely dysfunctional in septic patients. This very likely contributes to the development of nosocomial infections and patients’ inability to clear primary infections. Consequently, therapies capable of restoring immune function represent a new worthwhile strategy. Although we cannot predict that these therapies will be efficacious, they surely deserve to be fully investigated, considering the high mortality that has characterized septic syndromes for 25 years. In order not to repeat the mistakes from the past, an absolute prerequisite for clinical trials is to systematically assess patients’ immune functions to be able to define individualized immunotherapy.
References
Angus DC, Wax RS (2001) Epidemiology of sepsis: an update. Crit. Care. Med. 29:S109–16.
Brun-Buisson C, Meshaka P, Pinton P, Vallet B (2004) EPISEPSIS: a reappraisal of the epidemiology and outcome of severe sepsis in French intensive care units. Intensive. Care. Med. 30:580–8.
Brun-Buisson C, Roudot-Thoraval F, Girou E, Grenier-Sennelier C, Durand-Zaleski I (2003) The costs of septic syndromes in the intensive care unit and influence of hospital-acquired sepsis. Intensive. Care. Med. 29:1464–71.
Dombrovskiy VY, Martin AA, Sunderram J, Paz HL (2007) Rapid increase in hospitalization and mortality rates for severe sepsis in the United States: a trend analysis from 1993 to 2003. Crit. Care. Med. 35:1244–50.
Zeni F, Freeman B, Natanson C (1997) Anti-inflammatory therapies to treat sepsis and septic shock: a reassessment. Crit. Care. Med. 25:1095–100.
Remick DG (2007) Pathophysiology of sepsis. Am. J. Pathol. 170:1435–44.
Hotchkiss RS, Karl IE (2003) The pathophysiology and treatment of sepsis. N. Engl. J. Med. 348:138–150.
Annane D, Bellissant E, Cavaillon JM (2005) Septic shock. Lancet 365:63–78.
Munford RS, Pugin J (2001) Normal responses to injury prevent systemic inflammation and can be immunosuppressive. Am. J. Respir. Crit. Care. Med. 163:316–321.
Monneret G, Lepape A, Voirin N, Bohe J, Venet F, Debard AL, Thizy H, Bienvenu J, Gueyffier F, Vanhems P (2006) Persisting low monocyte human leukocyte antigen-DR expression predicts mortality in septic shock. Intensive. Care. Med. 32:1175–83.
Pachot A, Lepape A, Vey S, Bienvenu J, Mougin B, Monneret G (2006) Systemic transcriptional analysis in survivor and non-survivor septic shock patients: a preliminary study. Immunol. Lett. 106:63–71.
Pugin J (2007) Immunostimulation is a rational therapeutic strategy in sepsis. Novartis. Found. Symp. 280:21–7.
Wesche DE, Lomas-Neira JL, Perl M, Chung CS, Ayala A (2005) Leukocyte apoptosis and its significance in sepsis and shock. J. Leukoc. Biol. 78:325–37.
Monneret G, Debard AL, Venet F, Bohe J, Hequet O, Bienvenu J, Lepape A (2003) Marked elevation of human circulating CD4+CD25+ regulatory T cells in sepsis-induced immunoparalysis. Crit. Care. Med. 31:2068–71.
Lederer JA, Rodrick ML, Mannick JA (1999) The effects of injury on the adaptive immune response. Shock 11:153–9.
Manjuck J, et al. (2000) Decreased response to recall antigens is associated with depressed costimulatory receptor expression in septic critically ill patients. J. Lab. Clin. Med. 135:153–60.
Roth G, Moser B, Krenn C, Brunner M, Haisjackl M, Almer G, Gerlitz S, Wolner E, Boltz-Nitulescu G, Ankersmit HJ (2003) Susceptibility to programmed cell death in T-lymphocytes from septic patients: a mechanism for lymphopenia and Th2 predominance. Biochem. Biophys. Res. Commun. 308:840–6.
Meakins JL, Pietsch JB, Bubenick O, Kelly R, Rode H, Gordon J, MacLean LD (1977) Delayed hypersensitivity: indicator of acquired failure of host defenses in sepsis and trauma. Ann. Surg. 186:241–50.
Rode HN, Christou NV, Bubenik O, Superina R, Gordon J, Meakins JL, MacLean LD (1982) Lymphocyte function in anergic patients. Clin. Exp. Immunol. 47:155–61.
Christou NV, Meakins JL, Gordon J, Yee J, Hassan-Zahraee M, Nohr CW, Shizgal HM, MacLean LD (1995) The delayed hypersensitivity response and host resistance in surgical patients: 20 years later. Ann. Surg. 222:534–46.
O’Mahony JB, Palder SB, Wood JJ, McIrvine A, Rodrick ML, Demling RH, Mannick JA (1984) Depression of cellular immunity after multiple trauma in the absence of sepsis. J. Trauma 24:869–75.
Faist E, Kupper TS, Baker CC, Chaudry IH, Dwyer J, Baue AE (1986) Depression of cellular immunity after major injury: its association with posttraumatic complications and its reversal with immunomodulation. Arch. Surg. 121:1000–5.
Hensler T, Hecker H, Heeg K, Heidecke CD, Bartels H, Barthlen W, Wagner H, Siewert JR, Holzmann B (1997) Distinct mechanisms of immunosuppression as a consequence of major surgery. Infect. Immun. 65:2283–91.
Puyana JC, Pellegrini JD, De AK, Kodys K, Silva WE, Miller CL (1998) Both T-helper-1- and T-helper-2-type lymphokines are depressed in posttrauma anergy. J. Trauma 44:1037–45.
De AK, Kodys KM, Pellegrini J, Yeh B, Furse RK, Bankey P, Miller-Graziano CL (2000) Induction of global anergy rather than inhibitory Th2 lymphokines mediates posttrauma T cell immunodepression. Clin. Immunol. 96:52–66.
Pellegrini JD, De AK, Kodys K, Puyana JC, Furse RK, Miller-Graziano C (2000) Relationships between T lymphocyte apoptosis and anergy following trauma. J. Surg. Res. 88:200–6.
Laudanski K, et al. (2006) Cell-specific expression and pathway analyses reveal alterations in trauma-related human T cell and monocyte pathways. Proc. Natl. Acad. Sci. U. S. A. 103:15564–9.
Bandyopadhyay G, De A, Laudanski K, Li F, Lentz C, Bankey P, Miller-Graziano C (2007) Negative signaling contributes to T-cell anergy in trauma patients. Crit. Care. Med. 35:794–801.
Murphy T, Paterson H, Rogers S, Mannick JA, Lederer JA (2003) Use of intracellular cytokine staining and bacterial superantigen to document suppression of the adaptive immune system in injured patients. Ann. Surg. 238:401–10.
Heidecke CD, Hensler T, Weighardt H, Zantl N, Wagner H, Siewert JR, Holzmann B (1999) Selective defects of T lymphocyte function in patients with lethal intraabdominal infection. Am. J. Surg. 178:288–92.
Ochoa JB, Makarenkova V (2005) T lymphocytes. Crit. Care. Med. 33:S510–3.
Cavaillon JM, Adib-Conquy M (2007) Determining the degree of immunodysregulation in sepsis. Contrib. Nephrol. 156:101–11.
Munoz C, Carlet J, Fitting C, Misset B, Bleriot JP, Cavaillon JM (1991) Dysregulation of in vitro cytokine production by monocytes during sepsis. J. Clin. Invest. 88:1747–54.
van Deuren M, van der Ven-Jongekrijg J, Demacker PN, Bartelink AK, van Dalen R, Sauerwein RW, Gallati H, Vannice JL, van der Meer JW (1994) Differential expression of proinflammatory cytokines and their inhibitors during the course of meningococcal infections. J. Infect. Dis. 169:157–61.
Munoz C, Misset B, Fitting C, Bleriot JP, Carlet J, Cavaillon JM (1991) Dissociation between plasma and monocyte-associated cytokines during sepsis. Eur. J. Immunol. 21:2177–84.
Rigato O, Salomao R (2003) Impaired production of interferon-gamma and tumor necrosis factor-alpha but not of interleukin 10 in whole blood of patients with sepsis. Shock 19:113–6.
Stanilova SA, Karakolev ZT, Dimov GS, Dobreva ZG, Miteva LD, Slavov ES, Stefanov CS, Stanilov NS (2005) High interleukin 12 and low interleukin 10 production after in vitro stimulation detected in sepsis survivors. Intensive Care Med. 31:401–7.
Cavaillon JM, Adrie C, Fitting C, Adib-Conquy M (2003) Endotoxin tolerance: is there a clinical relevance? J. Endotoxin. Res. 9:101–7.
Zhang X, Morrison DC (1993) Lipopolysaccharide structure-function relationship in activation versus reprogramming of mouse peritoneal macrophages. J. Leukoc. Biol. 54:444–50.
Cavaillon JM, Adib-Conquy M (2006) Bench-to-bedside review: endotoxin tolerance as a model of leukocyte reprogramming in sepsis. Crit. Care 10:233.
Nierhaus A, Montag B, Timmler N, Frings DP, Gutensohn K, Jung R, Schneider CG, Pothmann W, Brassel AK, Schulte Am Esch J (2003) Reversal of immunoparalysis by recombinant human granulocyte-macrophage colony-stimulating factor in patients with severe sepsis. Intensive Care Med. 29:646–51.
Ploder M, Pelinka L, Schmuckenschlager C, Wessner B, Ankersmit HJ, Fuerst W, Redl H, Roth E, Spittler A (2006) Lipopolysaccharide-induced tumor necrosis factor alpha production and not monocyte human leukocyte antigen-DR expression is correlated with survival in septic trauma patients. Shock 25:129–34.
Marshall JC (2003) Such stuff as dreams are made on: mediator-directed therapy in sepsis. Nat. Rev. Drug Discov 2:391–405.
Perry SE, Mostafa SM, Wenstone R, McLaughlin PJ (2002) Low plasma granulocyte-macrophage colony stimulating factor is an indicator of poor prognosis in sepsis. Intensive Care Med. 28:981–4.
Wang H, et al. (1999) HMG-1 as a late mediator of endotoxin lethality in mice. Science 285:248–51.
Sunden-Cullberg J, et al. (2005) Persistent elevation of high mobility group box-1 protein (HMGB1) in patients with severe sepsis and septic shock. Crit. Care Med. 33:564–73.
Angus DC, Yang L, Kong L, Kellum JA, Delude RL, Tracey KJ, Weissfeld L (2007) Circulating high-mobility group box 1 (HMGB1) concentrations are elevated in both uncomplicated pneumonia and pneumonia with severe sepsis. Crit. Care Med. 35:1061–7.
Bianchi ME (2007) DAMPs, PAMPs and alarmins: all we need to know about danger. J. Leukoc. Biol. 81:1–5.
Pachot A, et al. (2005) Longitudinal study of cytokine and immune transcription factor mRNA expression in septic shock. Clin. Immunol. 114:61–9.
Gogos CA, et al. (2000) Pro- versus anti-inflammatory cytokine profile in patients with severe sepsis: a marker for prognosis and future therapeutic options. J. Infect. Dis. 181:176–80.
van Dissel JT, et al. (1998) Anti-inflammatory cytokine profile and mortality in febrile patients. Lancet 351:950–3.
Hynninen M, et al. (2003) Predictive value of monocyte histocompatibility leukocyte antigen-DR expression and plasma interleukin-4 and -10 levels in critically ill patients with sepsis. Shock 20:1–4.
Carrol ED, Thomson AP, Jones AP, Jeffers G, Hart CA (2005) A predominantly anti-inflammatory cytokine profile is associated with disease severity in meningococcal sepsis. Intensive Care Med. 31:1415–9.
Monneret G, et al. (2004) The anti-inflammatory response dominates after septic shock: association of low monocyte HLA-DR expression and high interleukin-10 concentration. Immunol. Lett. 95:193–8.
Abe R, et al. (2007) Up-regulation of interleukin-10 mRNA expression in peripheral leukocytes predicts poor outcome and diminished human leukocyte antigen-DR expression on monocytes in septic patients. J. Surg. Res. Aug 20 [Epub ahead of print].
Lekkou A, et al. (2004) Cytokine production and monocyte HLA-DR expression as predictors of outcome for patients with community-acquired severe infections. Clin. Diagn. Lab. Immunol. 11: 161–7.
Leonidou L, Mouzaki A, Michalaki M, Delastic AL, Kyriazopoulou V, Bassaris HP, Gogos CA (2007) Cytokine production and hospital mortality in patients with sepsis-induced stress hyperglycemia. J. Infect. 55:340–6.
Oberholzer A, Oberholzer C, Moldawer LL (2002) Interleukin-10: a complex role in the pathogenesis of sepsis syndromes and its potential as an anti-inflammatory drug. Crit. Care. Med. 30:S58–63.
Sfeir T, et al. (2001) Role of interleukin-10 in monocyte hyporesponsiveness associated with septic shock. Crit. Care. Med. 29:129–33.
Muehlstedt SG, Lyte M, Rodriguez JL (2002) Increased IL-10 production and HLA-DR suppression in the lungs of injured patients precede the development of nosocomial pneumonia. Shock 17:443–50.
Randow F, et al. (1995) Mechanism of endotoxin desensitization: involvement of interleukin 10 and transforming growth factor beta. J. Exp. Med. 181:1887–92.
Astiz M, et al. (1996) Monocyte response to bacterial toxins, expression of cell surface receptors, and release of anti-inflammatory cytokines during sepsis. J. Lab. Clin. Med. 128:594–600.
Monneret G, Voirin N, Krawice-Radanne I, Bohe J, Lepape A, Rouas-Freiss N, Carosella ED (2007) Soluble human leukocyte antigen-G5 in septic shock: marked and persisting elevation as a predictor of survival. Crit. Care. Med. 35:1942–7.
Martinez C, et al. (2002) Anti-inflammatory role in septic shock of pituitary adenylate cyclase-activating polypeptide receptor. Proc. Natl. Acad. Sci. U. S. A. 99:1053–8.
Yaqub S, Solhaug V, Vang T, Solberg R, Aasen A, Tasken K, Wang JE (2003) Ahuman whole blood model of LPS-mediated suppression of T cell activation. Med. Sci. Monit. 9:BR120–6.
Catania A, Cutuli M, Garofalo L, Airaghi L, Valenza F, Lipton JM, Gattinoni L (2000) Plasma concentrations and anti-L-cytokine effects of alpha-melanocyte stimulating hormone in septic patients. Crit. Care. Med. 28:1403–7.
Christ-Crain M, Morgenthaler NG, Struck J, Harbarth S, Bergmann A, Muller B (2005) Midregional pro-adrenomedullin as a prognostic marker in sepsis: an observational study. Crit. Care 9:R816–24.
Le Tulzo Y, et al. (2004) Monocyte human leukocyte antigen-DR transcriptional downregulation by cortisol during septic shock. Am. J. Respir. Crit. Care. Med. 169:1144–51.
Elenkov IJ, Wilder RL, Chrousos GP, Vizi ES (2000) The sympathetic nerve: an integrative interface between two supersystems: the brain and the immune system. Pharmacol. Rev. 52:595–638.
Tracey KJ (2007) Physiology and immunology of the cholinergic antiinflammatory pathway. J. Clin. Invest. 117:289–96.
Osuchowski MF, Welch K, Yang H, Siddiqui J, Remick DG (2007) Chronic sepsis mortality characterized by an individualized inflammatory response. J. Immunol. 179:623–30.
Osuchowski MF, Welch K, Siddiqui J, Remick DG (2006) Circulating cytokine/inhibitor profiles reshape the understanding of the SIRS/CARS continuum in sepsis and predict mortality. J. Immunol. 177:1967–74.
Oberholzer A, Souza SM, Tschoeke SK, Oberholzer C, Abouhamze A, Pribble JP, Moldawer LL (2005) Plasma cytokine measurements augment prognostic scores as indicators of outcome in patients with severe sepsis. Shock 23:488–93.
Huang X, Venet F, Chung CS, Lomas-Neira J, Ayala A (2007) Changes in dendritic cell function in the immune response to sepsis: cell- and tissue-based therapy. Expert. Opin. Biol. Ther. 7:929–38.
Benjamim CF, Lundy SK, Lukacs NW, Hogaboam CM, Kunkel SL (2005) Reversal of long-term sepsis-induced immunosuppression by dendritic cells. Blood 105:3588–95.
Toliver-Kinsky TE, Cui W, Murphey ED, Lin C, Sherwood ER (2005) Enhancement of dendritic cell production by fms-like tyrosine kinase-3 lig-and increases the resistance of mice to a burn wound infection. J. Immunol. 174:404–10.
Wysocka M, Montaner LJ, Karp CL (2005) Flt3 ligand treatment reverses endotoxin tolerance-related immunoparalysis. J. Immunol. 174:7398–402.
Guisset O, Dilhuydy MS, Thiebaut R, Lefevre J, Camou F, Sarrat A, Gabinski C, Moreau JF, Blanco P (2007) Decrease in circulating dendritic cells predicts fatal outcome in septic shock. Intensive Care Med. 33:148–52.
Ho CS, Lopez JA, Vuckovic S, Pyke CM, Hockey RL, Hart DN (2001) Surgical and physical stress increases circulating blood dendritic cell counts independently of monocyte counts. Blood 98:140–5.
Hotchkiss RS, Tinsley KW, Swanson PE, Grayson MH, Osborne DF, Wagner TH, Cobb JP, Coopersmith C, Karl IE (2002) Depletion of dendritic cells, but not macrophages, in patients with sepsis. J. Immunol. 168:2493–500.
Faivre V, Lukaszewicz AC, Alves A, Charron D, Payen D, Haziot A (2007) Accelerated in vitro differentiation of blood monocytes into dendritic cells in human sepsis. Clin. Exp. Immunol. 147:426–39.
De AK, Laudanski K, Miller-Graziano CL (2003) Failure of monocytes of trauma patients to convert to immature dendritic cells is related to preferential macrophage-colony-stimulating factor-driven macrophage differentiation. J. Immunol. 170:6355–62.
Koppelman B, Neefjes JJ, de Vries JE, de Waal Malefyt R (1997) Interleukin-10 down-regulates MHC class II alphabeta peptide complexes at the plasma membrane of monocytes by affecting arrival and recycling. Immunity 7:861–71.
Krakauer T, Oppenheim JJ (1993) IL-1 and tumor necrosis factor-alpha each up-regulate both the expression of IFN-gamma receptors and enhance IFN-gamma-induced HLA-DR expression on human monocytes and a human monocytic cell line (THP-1). J. Immunol. 150:1205–11.
Schwiebert LM, Schleimer RP, Radka SF, Ono SJ (1995) Modulation of MHC class II expression in human cells by dexamethasone. Cell. Immunol. 165:12–93.
Basta PV, Moore TL, Yokota S, Ting JP (1989) A beta-adrenergic agonist modulates DR alpha gene transcription via enhanced cAMP levels in a glioblastoma multiforme line. J. Immunol. 142: 2895–901.
Fumeaux T, Pugin J (2006) Is the measurement of monocytes HLA-DR expression useful in patients with sepsis? Intensive Care Med. 32:1106–8.
Tschoeke SK, Moldawer LL (2005) Human leukocyte antigen expression in sepsis: what have we learned? Crit. Care Med. 33:236–7.
Wolk K, et al. (2000) Impaired antigen presentation by human monocytes during endotoxin tolerance. Blood 96:218–23.
Schneider C, et al. (2004) Perioperative recombinant human granulocyte colony-stimulating factor (filgrastim) treatment prevents immunoinflammatory dysfunction associated with major surgery. Ann. Surg. 239:75–81.
Piani A, et al. (2000) Expression of MHC class II molecules contributes to lipopolysaccharide responsiveness. Eur. J. Immunol. 30:3140–6.
Hershman MJ, et al. (1990) Monocyte HLA-DR antigen expression characterizes clinical outcome in the trauma patient. Br. J. Surg. 77:204–7.
Wakefield CH, Carey PD, Foulds S, Monson JR, Guillou PJ (1993) Changes in major histocompatibility complex class II expression in monocytes and T cells of patients developing infection after surgery. Br. J. Surg. 80:205–9.
Giannoudis PV, et al. (2000) Immediate IL-10 expression following major orthopaedic trauma: relationship to anti-inflammatory response and subsequent development of sepsis. Intensive Care Med. 26:1076–81.
Ditschkowski M, et al. (1999) HLA-DR expression and soluble HLA-DR levels in septic patients after trauma. Ann. Surg. 229:246–54.
Satoh A, et al. (2002) Human leukocyte antigen-DR expression on peripheral monocytes as a predictive marker of sepsis during acute pancreatitis. Pancreas 25:245–50.
Tschaikowsky K, et al. (2002) Coincidence of pro-and anti-inflammatory responses in the early phase of severe sepsis: longitudinal study of mononuclear histocompatibility leukocyte anti-gen-DR expression, procalcitonin, C-reactive protein, and changes in T-cell subsets in septic and postoperative patients. Crit. Care. Med. 30:1015–23.
Walsh DS, et al. (2005) Characterization of circulating monocytes expressing HLA-DR or CD71 and related soluble factors for 2 weeks after severe, non-thermal injury. J. Surg. Res. 129:221–30.
Allen ML, et al. (2002) Early postoperative monocyte deactivation predicts systemic inflammation and prolonged stay in pediatric cardiac intensive care. Crit. Care. Med. 30:1140–5.
Sachse C, Prigge M, Cramer G, Pallua N, Henkel E (1999) Association between reduced human leukocyte antigen (HLA)-DR expression on blood monocytes and increased plasma level of interleukin-10 in patients with severe burns. Clin. Chem. Lab. Med. 37:193–8.
Venet F, Tissot S, Debard AL, Faudot C, Crampe C, Pachot A, Ayala A, Monneret G (2007) Decreased monocyte human leukocyte antigen-DR expression after severe burn injury: correlation with severity and secondary septic shock. Crit. Care. Med. 35:1910–1917.
Haveman JW, van den Berg AP, van den Berk JM, Mesander G, Slooff MJ, de Leij LH, The TH (1999) Low HLA-DR expression on peripheral blood monocytes predicts bacterial sepsis after liver transplantation: relation with prednisolone intake. Transpl. Infect. Dis. 1:146–52.
Haveman JW, van den Berg AP, Verhoeven EL, Nijsten MW, van den Dungen JJ, The HT, Zwaveling JH (2006) HLA-DR expression on monocytes and systemic inflammation in patients with ruptured abdominal aortic aneurysms. Crit. Care. 10:R119.
Perry SE, et al. (2003) Is low monocyte HLA-DR expression helpful to predict outcome in severe sepsis? Intensive Care Med. 29:1245–52.
Muller Kobold AC, et al. (2000) Leukocyte activation in sepsis; correlations with disease state and mortality. Intensive Care Med. 26:883–92.
Fumeaux T, Pugin J (2002) Role of interleukin-10 in the intracellular sequestration of human leukocyte antigen-DR in monocytes during septic shock. Am. J. Respir. Crit. Care Med. 166:1475–82.
Docke WD, et al. (1997) Monocyte deactivation in septic patients: restoration by IFN-gamma treatment. Nat. Med. 3:678–81.
Saenz JJ, et al. (2002) Early prognosis in severe sepsis via analyzing the monocyte immunophenotype. Intensive Care Med. 27:970–7.
Monneret G, et al. (2002) Analytical requirements for measuring monocytic HLA-DR by flow cytometry: application to the monitoring of patients with septic shock. Clin. Chem. 48:1589–92.
Monneret G, et al. (2006) Persisting low monocyte human leukocyte antigen-DR expression predicts mortality in septic shock. Intensive Care Med. 32:1175–83.
Pachot A, Monneret G, Brion A, Venet F, Bohe J, Bienvenu J, Mougin B, Lepape A (2005) Messenger RNA expression of major histocompatibility complex class II genes in whole blood from septic shock patients. Crit. Care Med. 33:31–8.
Docke WD, et al. (2005) Monitoring temporary immunodepression by flow cytometric measurement of monocytic HLA-DR expression: a multicenter study. Clin. Chem. 51:2341–7.
Heinzelmann M, Mercer-Jones M, Cheadle WG, Polk HC Jr (1996) CD14 expression in injured patients correlates with outcome. Ann. Surg. 224:91–6.
Venet F, Pachot A, Debard AL, Bohe J, Bienvenu J, Lepape A, Powell WS, Monneret G (2006) Human CD4+CD25+ regulatory T lymphocytes inhibit lipopolysaccharide-induced monocyte survival through a Fas/Fas ligand-dependent mechanism. J. Immunol. 177:6540–7.
Aalto H, Takala A, Kautiainen H, Siitonen S, Repo H (2007) Monocyte CD14 and soluble CD14 in predicting mortality of patients with severe community acquired infection. Scand. J. Infect. Dis. 39:596–603.
Wolk K, Hoflich C, Zuckermann-Becker H, Docke WD, Volk HD, Sabat R (2007) Reduced monocyte CD86 expression in postinflammatory immunodeficiency. Crit. Care Med. 35:458–67.
Pangault C, Le Tulzo Y, Tattevin P, Guilloux V, Bescher N, Drenou B (2006) Down-modulation of granulocyte macrophage-colony stimulating factor receptor on monocytes during human septic shock. Crit. Care Med. 34:1193–201.
Pachot A, Faudot C, Cazalis MA, Mougin B, Lepape A, Monneret G (2007) Decreased expression of CX3CR1 in septic shock: transcriptional regulation by LPS, cortisol and soluble fractalkine. Inflammation Res. 56:S131.
Shelley O, Murphy T, Paterson H, Mannick JA, Lederer JA (2003) Interaction between the innate and adaptive immune systems is required to survive sepsis and control inflammation after injury. Shock 20:123–9.
Hotchkiss RS, Osmon SB, Chang KC, Wagner TH, Coopersmith CM, Karl IE (2005) Accelerated lymphocyte death in sepsis occurs by both the death receptor and mitochondrial pathways. J. Immunol. 174:5110–8.
Menges T, Engel L, Welters I, Wagner RM, Little S, Ruwoldt R, Wollbrueck M, Hempelmann G (1999) Changes in blood lymphocyte populations after multiple trauma: association with posttraumatic complications. Crit. Care Med. 27:733–40.
Felmet KA, Hall MW, Clark RS, Jaffe R, Carcillo JA (2005) Prolonged lymphopenia, lymphoid depletion, and hypoprolactinemia in children with nosocomial sepsis and multiple organ failure. J. Immunol. 174:3765–72.
Spolarics Z, Siddigi M, Siegel JH, Garcia ZC, Stein DS, Denny T, Deitch EA (2003) Depressed interleukin-12-producing activity by monocytes correlates with adverse clinical course and a shift toward Th2-type lymphocyte pattern in severely injured male trauma patients. Crit. Care Med. 31: 1722–9.
Wick M, Kollig E, Muhr G, Koller M (2000) The potential pattern of circulating lymphocytes TH1/TH2 is not altered after multiple injuries. Arch. Surg. 135:1309–14.
Venet F, Lepape A, Debard AL, Bienvenu J, Bohe J, Monneret G (2004) The Th2 response as monitored by CRTH2 or CCR3 expression is severely decreased during septic shock. Clin. Immunol. 113:278–84.
Venet F, Pachot A, Debard AL, Bohe J, Bienvenu J, Lepape A, Monneret G (2004) Increased percentage of CD4+CD25+ regulatory T cells during septic shock is due to the decrease of CD4+CD25− lymphocytes. Crit. Care Med. 32:2329–31.
Venet F, Bohe J, Debard AL, Bienvenu J, Lepape A, Monneret G (2005) Both percentage of gammadelta T lymphocytes and CD3 expression are reduced during septic shock. Crit. Care. Med. 33:2836–40.
Scumpia PO, et al. (2007) Treatment with GITR agonistic antibody corrects adaptive immune dysfunction in sepsis. Blood 110:3673–81.
Sakaguchi S (2004) Naturally arising CD4+ regulatory T cells for immunologic self-tolerance and negative control of immune responses. Annu. Rev. Immunol. 22:531–62.
MacConmara MP, et al. (2006) Increased CD4+ CD25+ T regulatory cell activity in trauma patients depresses protective Th1 immunity. Ann. Surg. 244:514–23.
Scumpia PO, et al. (2006) Increased natural CD4+CD25+ regulatory T cells and their suppressor activity do not contribute to mortality in murine polymicrobial sepsis. J. Immunol. 177: 7943–9.
Wisnoski N, Chung CS, Chen Y, Huang X, Ayala A (2006) The contribution of CD4+CD25+ T-regulatory-cells to immune suppression in sepsis. Shock 27:251–7.
Offner H, Subramanian S, Parker SM, Wang C, Afentoulis ME, Lewis A, Vandenbark AA, Hurn PD (2006) Splenic atrophy in experimental stroke is accompanied by increased regulatory T cells and circulating macrophages. J. Immunol. 176: 6523–31.
Chen X, Bäumel M, Männel DN, Howard OM, Oppenheim JJ (2007) Interaction of TNF with TNF receptor type 2 promotes expansion and function of mouse CD4+CD25+ T regulatory cells. J. Immunol. 179:154–61.
Schneider DF, Glenn CH, Faunce DE (2007) Innate lymphocyte subsets and their immunoregulatory roles in burn injury and sepsis. J. Burn Care Res. 28:365–79.
Matsushima A, et al. (2004) Early activation of gammadelta T lymphocytes in patients with severe systemic inflammatory response syndrome. Shock 22:11–5.
Chang KC, Unsinger J, Davis CG, Schwulst SJ, Muenzer JT, Strasser A, Hotchkiss RS (2007) Multiple triggers of cell death in sepsis: death receptor and mitochondrial-mediated apoptosis. FASEB J. 21:708–19.
Hotchkiss RS, Nicholson DW (2006) Apoptosis and caspases regulate death and inflammation in sepsis. Nat. Rev. Immunol. 6:813–22.
Hotchkiss RS, et al. (1999) Apoptotic cell death in patients with sepsis, shock, and multiple organ dysfunction. Crit. Care Med. 27:1230–51.
Hotchkiss RS, et al. (2001) Sepsis-induced apoptosis causes progressive profound depletion of B and CD4+ T lymphocytes in humans. J. Immunol. 166:6952–63.
Toti P, De Felice C, Occhini R, Schuerfeld K, Stumpo M, Epistolato MC, Vatti R, Buonocore G (2004) Spleen depletion in neonatal sepsis and chorioamnionitis. Am. J. Clin. Pathol. 122:765–71.
Le Tulzo Y, et al. (2002) Early circulating lymphocyte apoptosis in human septic shock is associated with poor outcome. Shock 18:487–94.
Bilbault P, et al. (2004) Transient Bcl-2 gene down-expression in circulating mononuclear cells of severe sepsis patients who died despite appropriate intensive care. Intensive. Care Med. 30:408–15.
Bilbault P, Lavaux T, Launoy A, Gaub MP, Meyer N, Oudet P, Pottecher T, Jaeger A, Schneider F (2007) Influence of drotrecogin alpha (activated) infusion on the variation of Bax/Bcl-2 and Bax/Bcl-xl ratios in circulating mononuclear cells: a cohort study in septic shock patients. Crit. Care Med. 35:69–75.
Williams TE, Ayala A, Chaudry IH (1997) Inducible macrophage apoptosis following sepsis is mediated by cysteine protease activation and nitric oxide release. J. Surg. Res. 70:113–8.
Adrie C, et al. (2001) Mitochondrial membrane potential and apoptosis peripheral blood monocytes in severe human sepsis. Am. J. Respir. Crit. Care Med. 164:389–95.
Swan R, Chung CS, Albina J, Cioffi W, Perl M, Ayala A (2007) Polymicrobial sepsis enhances clearance of apoptotic immune cells by splenic macrophages. Surgery 142:253–61.
Hoesel LM, et al. (2005) Harmful and protective roles of neutrophils in sepsis. Shock 24:40–7.
Mutunga M, Fulton B, Bullock R, Batchelor A, Gascoigne A, Gillespie JI, Baudouin SV (2001) Circulating endothelial cells in patients with septic shock. Am. J. Respir. Crit. Care Med. 163:195–200.
Roth GA, et al. (2004) Elevated serum levels of epithelial cell apoptosis-specific cytokeratin 18 neoepitope m30 in critically ill patients. Shock 22: 218–20.
Kumar A, Kumar A, Michael P, Brabant D, Parissenti AM, Ramana CV, Xu X, Parrillo JE (2005) Human serum from patients with septic shock activates transcription factors STAT1, IRF1, and NF-kappaB and induces apoptosis in human cardiac myocytes. J. Biol. Chem. 280:42619–26.
Minambres E, Lopez-Escribano H, Ballesteros MA, Pena M, Lopez-Hoyos M (2005) Apoptosis of Jurkat cells induced by serum of patients with acute severe brain injury. Intensive Care Med. 31:791–8.
Ballesteros MA, Lopez-Hoyos M, Munoz P, Marin MJ, Minambres E (2007) Apoptosis of neuronal cells induced by serum of patients with acute brain injury: a new in vitro prognostic model. Intensive Care Med. 33:58–65.
De Freitas I, Fernandez-Somoza M, Essenfeld-Sekler E, Cardier JE (2004) Serum levels of the apoptosis-associated molecules, tumor necrosis factor-alpha/tumor necrosis factor type-I receptor and Fas/FasL, in sepsis. Chest 125:2238–46.
Papathanassoglou ED, Moynihan JA, McDermott MP, Ackerman MH (2001) Expression of Fas (CD95) and Fas ligand on peripheral blood mononuclear cells in critical illness and association with multiorgan dysfunction severity and survival. Crit. Care Med. 29:709–18.
Hotchkiss RS, Coopersmith CM, Karl IE (2005) Prevention of lymphocyte apoptosis: a potential treatment of sepsis? Clin. Infect. Dis. 41:S465–9.
Wesche-Soldato DE, Swan RZ, Chung CS, Ayala A (2007) The apoptotic pathway as a therapeutic target in sepsis. Curr. Drug Targets 8:493–500.
Greineder CF, Nelson PW, Dressel AL, Erba HP, Younger JG (2007) In vitro and in silico analysis of annexin V binding to lymphocytes as a biomarker in emergency department sepsis studies. Acad. Emerg. Med. 14:763–71.
Cobb JP, O’Keefe GE (2004) Injury research in the genomic era. Lancet 363:2076–83.
Feezor RJ, Cheng A, Paddock HN, Baker HV, Moldawer LL (2005) Functional genomics and gene expression profiling in sepsis: beyond class prediction. Clin. Infect. Dis. 41(Suppl 7):S427–35.
Feezor RJ, Oberholzer C, Baker HV, Novick D, Rubinstein M, Moldawer LL et al. (2003) Molecular characterization of the acute inflammatory response to infections with gram-negative versus gram-positive bacteria. Infect. Immun. 71:5803–13.
Huang Q, et al. (2001) The plasticity of dendritic cell responses to pathogens and their components. Science 294:870–5.
Zhao B, Bowden RA, Stavchansky SA, Bowman PD (2001) Human endothelial cell response to gram-negative lipopolysaccharide assessed with cDNA microarrays. Am. J. Physiol. Cell. Physiol. 281:C1587–95.
Nau GJ, Richmond JF, Schlesinger A, Jennings EG, Lander ES, Young RA (2002) Human macrophage activation programs induced by bacterial pathogens. Proc. Natl. Acad. Sci. U. S. A. 99:1503–8.
Yu SL, et al. (2004) Differential gene expression in gram-negative and gram-positive sepsis. Am. J. Respir. Crit. Care Med. 169:1135–43.
Chinnaiyan AM, et al. (2001) Molecular signatures of sepsis: multiorgan gene expression profiles of systemic inflammation. Am. J. Pathol. 159:1199–1209.
Chung TP, et al. (2006) Molecular diagnostics in sepsis: from bedside to bench. J. Am. Coll. Surg. 203:585–98.
Prucha M, et al. (2004) Expression profiling: toward an application in sepsis diagnostics. Shock 22:29–33.
Johnson SB, Lissauer M, Bochicchio GV, Moore R, Cross AS, Scalea TM (2007) Gene expression profiles differentiate between sterile SIRS and early sepsis. Ann. Surg. 245:611–21.
Ramilo O, et al. (2007). Gene expression patterns in blood leukocytes discriminate patients with acute infections. Blood 109:2066–77.
Feezor RJ, et al. (2004) Genomic and proteomic determinants of outcome in patients undergoing thoracoabdominal aortic aneurysm repair. J. Immunol. 172:7103–9.
Biberthaler P, et al. (2003) Initial RNAexpression in human monocytes after multiple injury: a screening pilot study on potentially trauma-sensitive factors by using the microarray-technique. Eur. J. Med. Res. 8:473–84.
Schneider EM, et al. (2006) MAPkinase gene expression, as determined by microarray analysis, distinguishes uncomplicated from complicated reconstitution after major surgical trauma. Ann. N. Y. Acad. Sci. 1090:429–44.
Bogner V, et al. (2007) Gene expression profiles are influenced by ISS, MOF, and clinical outcome in multiple injured patients: a genome-wide comparative analysis. Langenbecks Arch. Surg. 392:255–65.
Auffray C, Fogg D, Garfa M, Elain G, Join-Lambert O, Kayal S, Sarnacki S, Cumano A, Lauvau G, Geissmann F (2007) Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science 317:666–70.
Wong HR, et al. (2007) Genome-level expression profiles in pediatric septic shock indicate a role for altered zinc homeostasis in poor outcome. Physiol. Genomics 30:146–55.
Shanley TP, et al. (2007) Genome-level longitudinal expression of signaling pathways and gene networks in pediatric septic shock. Mol. Med. 13:495–508.
de Reynies A, et al. (2006) Comparison of the latest commercial short and long oligonucleotide microarray technologies. BMC Genomics 7:51.
Bammler T, et al. (2005) Standardizing global gene expression analysis between laboratories and across platforms. Nat. Methods 2:351–6.
Irizarry RA, et al. (2005) Multiple-laboratory comparison of microarray platforms. Nat. Methods 2:345–50.
Storey JD, Xiao W, Leek JT, Tompkins RG, Davis RW (2005) Significance analysis of time course microarray experiments. Proc. Natl. Acad. Sci. U. S. A. 102:12837–42.
Calvano SE, et al. (2005) Anetwork-based analysis of systemic inflammation in humans. Nature 437:1032–7.
Dahmer MK, Randolph A, Vitali S, Quasney MW (2005) Genetic polymorphisms in sepsis. Pediatr. Crit. Care Med. 6:S61–73.
Papathanassoglou ED, Giannakopoulou MD, Bozas E (2006) Genomic variations and susceptibility to sepsis. AACN Adv. Crit. Care 17:394–422.
Kalechman Y, Gafter U, Gal R, Rushkin G, Yan D, Albeck M, Sredni B (2002) Anti-IL-10 therapeutic strategy using the immunomodulator AS101 in protecting mice from sepsis-induced death: dependence on timing of immunomodulating intervention. J. Immunol. 169:384–92.
Murphey ED, Sherwood ER (2006) Bacterial clearance and mortality are not improved by a combination of IL-10 neutralization and IFN-gamma administration in a murine model of post-CLP immunosuppression. Shock 26:417–24.
Polk HC Jr, Cheadle WG, Livingston DH, Rodriguez JL, Starko KM, Izu AE, Jaffe HS, Sonnenfeld G (1992) Arandomized prospective clinical trial to determine the efficacy of interferongamma in severely injured patients. Am. J. Surg. 163:191–6.
Dries DJ, et al. (1994) Effect of interferon gamma on infection-related death in patients with severe injuries: a randomized, double-blind, placebo-controlled trial. Arch. Surg. 129:1031–41.
Presneill JJ, Harris T, Stewart AG, Cade JF, Wilson JW (2002) A randomized phase II trial of granulocyte-macrophage colony-stimulating factor therapy in severe sepsis with respiratory dysfunction. Am. J. Respir. Crit. Care Med. 166:138–43.
Rosenbloom AJ, Linden PK, Dorrance A, Penkosky N, Cohen-Melamed MH, Pinsky MR (2005) Effect of granulocyte-monocyte colony-stimulating factor therapy on leukocyte function and clearance of serious infection in nonneutropenic patients. Chest 127:2139–50.
Nakos G, et al. (2002) Immunoparalysis in patients with severe trauma and the effect of inhaled interferon-γ. Crit. Care Med. 30:1488–94.
Leaver SK, Finney SJ, Burke-Gaffney A, Evans TW (2007) Sepsis since the discovery of Toll-like receptors: disease concepts and therapeutic opportunities. Crit. Care Med. 35:1404–10.
Alpdogan O, van den Brink MR (2005) IL-7 and IL-15: therapeutic cytokines for immunodeficiency. Trends Immunol. 26:56–64.
Snyder KM, Mackall CL, Fry TJ (2006) IL-7 in allogeneic transplant: clinical promise and potential pitfalls. Leuk. Lymphoma 47:1222–8.
Turgeon AF, Hutton B, Fergusson DA, McIntyre L, Tinmouth AA, Cameron DW, Hebert PC (2007) Meta-analysis: intravenous immunoglobulin in critically ill adult patients with sepsis. Ann. Intern. Med. 146:193–203.
Norrby-Teglund A, Haque KN, Hammarstrom L (2006) Intravenous polyclonal IgM-enriched immunoglobulin therapy in sepsis: a review of clinical efficacy in relation to microbiological etiology and severity of sepsis. J. Intern. Med. 260:509–16.
Weaver JG, Rouse MS, Steckelberg JM, Badley AD (2004) Improved survival in experimental sepsis with an orally administered inhibitor of apoptosis. FASEB J. 18:1185–91.
Acknowledgments
This review does not represent an exhaustive listing of studies aimed at monitoring immunological functions in the critically ill patient. We sincerely apologize for works not cited in this manuscript. G.M. thanks all the Immunology Laboratory staff at Hôpital E. Herriot for constant support over the years. None of the authors has any potential financial conflict of interest related to this manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Monneret, G., Venet, F., Pachot, A. et al. Monitoring Immune Dysfunctions in the Septic Patient: A New Skin for the Old Ceremony. Mol Med 14, 64–78 (2008). https://doi.org/10.2119/2007-00102.Monneret
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2119/2007-00102.Monneret