
Abstract
Fungi are a promising source for bioactive secondary metabolites against various agricultural pests. Soil samples were collected from the rhizosphere of various plants at El-Khatatba, Egypt, in May 2016. Sixty-two fungal isolates were locally isolated and screened against Tetranychus urticae Koch (Acari: Tetranychidae). Four fungal strains, showed potent control activities, were morphologically identified. Laboratory evaluation of the crude extracts of the selected strains, Aspergillus melleus, A. terreus, Emericella nidulans, and Chaetomium globosum, exhibited LC50 values of 10.27, 33.05, 14.68, and 22.40 mg/ml against females of T. urticae, respectively. Correspondent LC50 values that exhibited on eggs were 8.81, 23.17, 11.66, and 11.05 mg/ml. Consequently, secondary metabolites of A. melleus were separated by liquid chromatography/mass spectrometry. Compounds separated from the active fraction were identified as mellamide, ochratoxin C, nodulisporic acid, 7-Oxocurvularin, and 6-(4′-hydroxy-2′-methyl phenoxy)-(−)-(3R)-mellein. Obtained secondary metabolites are promising sources for biopesticides to be used in bio-rationale control of T. urticae.
Similar content being viewed by others

Background
Crop production is decreased annually by 18–26% because of arthropod pests (Culliney 2014). The two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae), can infest more than 200 plant species all over the world. Many reports showed that T. urticae can cause harsh damage to many agricultural crops such as vegetables, fruits, and ornamental plants (Fasulo and Denmark 2000). Management of T. urticae by using synthetic acaricides has been widely applied (Attia et al. 2013). Due to health and environmental hazards caused by chemical pesticides in addition to their effects on other non-target organisms such as predators, their use has been firmly regulated (Horikoshi et al. 2017). Moreover, the development of acaricidal resistance in T. urticae is highly increasing since they have an outstanding potential to develop such resistance. Therefore, the development of new acaricides with novel modes of action is increasingly needed (Marcic 2012).
Currently, many chemicals used in agriculture to control pests are originally derived from microbial metabolites (Horikoshi et al. 2017). Fungi and fungal metabolites exhibit a high toxicity to insects and mite pests; however, they show low toxicity to non-target organisms (Ragavendran and Natarajan 2015). Researchers thought that fungi use their products as well as mycotoxins as chemical defenses against different targets including insects and mites. For instance, mycotoxins such as aflatoxin B and trichothecenes exhibit toxicity to many insect pests (Srivastava et al. 2009). Consequently, microbial metabolites may be promising resources in a novel pesticide development (Horikoshi et al. 2017).
Beside predatory mites, plant extracts, and essential oils, microbial metabolites may be also promising tools in bio-rationale control of T. urticae (Attia et al. 2013). For example, thuringiensin produced by B. thuringiensis showed a potent acaricidal efficiency on T. urticae, T. cinnabarinus (Neal Jr et al. 1987), and Panonychus ulmi Koch (Vargas Mesina 1993). Pseudomonas fluorescens exhibited a strong efficacy on adults of Oligonychus coffeae Nietner under laboratory conditions (Roobakkumar et al. 2011). Also, an actinomycete-derived product, abamectin (avermectin) produced from Streptomyces avermitilis, demonstrated highly toxic effects on T. cinnabarinus (Wu and Liu 1997), T. urticae, Phyllocoptruta oleivora, Panonychus citri, and T. turkestani Ugarov and Nikolski (Putter et al. 1981).
The present study aimed to evaluate the control potential of ethyl acetate extract produced by different fungal strains against T. urticae under laboratory conditions. In addition, the active compound/s produced by the most effective strain will be separated, purified, and identified.
Materials and methods
All experiments were conducted at the laboratories of Plant Protection Department, Desert Research Centre, Egypt (2016–2018).
Collection of soil samples
Soil samples were collected from EL-Khatatba, Egypt, in May 2016. Plants at different fields were examined by the aid of a × 10 magnification lens for spider mite infestation. Samples were collected from the rhizosphere of the spider mite-infested plants at a depth of 10–15 cm under the soil surface. Soil samples (500 g each) were placed in sterilized polyethylene bags and stored at 4 °C until use.
Isolation, purification, and identification of fungi by cultural and morphological methods
Isolation of soil fungi was done by a serial dilution method. Two types of culture media were used for isolation of fungi: potato dextrose agar (PDA) and Czapek’s Dox agar supplemented with yeast extract (5 g/l). All media were supplemented with chloramphenicol antibiotic (25 mg/l). Isolation of fungi was carried out from dilutions of 10−2 and 10−3, where 0.5 ml was taken from each dilution and placed into Petri dishes with a solid media, then spread, using a sterilized glass spreader and was left for 30 min before incubation at 28 °C for 4–7 days. Each morphological distinct, fungal colony was sub-cultured and purified, using standard techniques.
Primary screening of fungal isolates against T. urticae
Sixty-two fungal isolates were screened for their control activity by testing their culture filtrates against adult spider mite females. Four discs (1 cm) were cut from fresh culture of each fungal isolate and inoculated in 100 ml of potato dextrose broth. The broth cultures were incubated in shaking incubator at 28 °C, 150 rpm for 7 days. Fungal cultures were filtered, using sterilized filter papers and centrifuged. Finally, the filtrates were preserved in refrigerator at 4 °C until use within a week to avoid any contamination or alteration of metabolites. Bioassay of fungal filtrates on spider mites was carried out by leaf-dipping method. Three mulberry leaf discs (25 mm) were immersed for 5 s in each fungal filtrate and dried at room temperature. The treated leaf discs were placed on wetted cotton wool in Petri dishes. Each was lined with cotton lining to prevent mites from escaping. Ten females were transferred to each leaf disc, using a fine paintbrush and incubated at 25 °C. Control was immersed in potato dextrose broth and mortality rates were observed after 5 days.
Selection and identification of the most potent isolates
Morphological identification
Four isolates (KF23, KF45, KF40, and KF9) out of 62 were selected for further investigation, as they achieved > 50% mortality in T. urticae. Cultures were grown on Czapek’s yeast extract agar and incubated at 28 °C for 7–10 days. The identification of the most potent isolates was carried out on morphological basis, using culture characteristics, e.g., growth rate, color, and pigmentation, as well as microscopic features, e.g., conidiophores, conidia, production of sclerotia, and dimensions of the different microscopic fungal structures. The results were confirmed by the scientists of Mycological Centre, Assiut University, Egypt.
Effect of crude extract of the selected isolates on adult females of T. urticae
Twelve plugs of freshly prepared culture of each fungal isolate were inoculated in 300 ml potato dextrose broth media and incubated in shaking incubator at 28 °C, 150 rpm for 10 days. After incubation time, each fungal culture was filtered and subjected to extraction with ethyl acetate (1:1) triple. The crude extract was dried, using a rotary evaporator, and stored in a freezer until use. Four concentrations (5, 10, 20, 30 mg/ml) were prepared from the crude extract of each fungal isolate. Bioassay of each extract was undertaken using the leaf dipping method. Leaf discs of 25 mm were immersed in each of the four concentrations, and another disc group was dipped in ethanol and used as a check and left to dry at room temperature. Treated leaf discs were placed on wetted cotton in Petri dishes (9 cm) and lined with a cotton lining to prevent spider mites from escaping, and then 20 females were transferred and incubated at 25 °C. The mortality rates were recorded after 3 days. Each concentration was replicated three times.
Effect of crude extract of the selected isolates on eggs of T. urticae
The ovicidal activity of the previously prepared concentrations of ethyl acetate extract of selected isolates was tested as follows: 10 females were placed on mulberry leaf discs (25 mm), prepared in 9 cm Petri dish to obtain same-aged eggs to be used in the tests. Twenty-four hours later, females were removed from the discs, eggs were counted, and three leaf discs, carrying eggs, were dipped into each concentration. The control discs were dipped in 70% ethanol and were left to dry. Observations continued daily until hatching of eggs in the control group.
Confirmation of the most potent strain by DNA sequencing
The strain was cultured on PDA overlaid with cellophane at 27 °C for 3–5 days. Total genomic DNA (deoxyribonucleic acid) was directly extracted from fungal mycelia of the KF23 strain, using the Quick-DNA™ Fungal/Bacterial Microprep Kit (Zymo research #D6007). The internal transcribed spacers (ITS1 and ITS4) were amplified using the universal primer pair ITS1 (5′-CTTGGTCATTTAGAGGAAGTAA-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) (White et al. 1990) in a 50-μl reaction mixture comprising 5 μl of genomic DNA, 1 μl of each primer, 25 μl Maxima Hot Start PCR Master Mix (Thermo K1051), and 18 μl of sterile distilled water. The polymerase chain reaction (PCR) was carried out with the following parameters: an initial denaturation at 95 °C (10 min), 35 cycles of primer denaturation at 95 °C (30s), annealing at 57 °C (1 min), and extension at 72 °C (1 min 30s). A final elongation step was allowed at 72 °C for 10 min. The PCR products were purified using Gene JET™ PCR Purification Kit (Thermo K0701), according to the manufacturer’s instructions. Sequencing was done at GATC Company Germany by using ABI 3730xl DNA sequencer and using forward and reverse primers. The sequence obtained was subjected into Geneious pro 8.9 software and compared to ITS sequences available in Gene Bank by blast analysis. Finally, the phylogenetic tree was constructed to establish the taxonomic rank of the fungus.
Separation, purification, and identification of acaricidal secondary metabolites produced by A. melleus
Preparation of extracellular crude extract from A. melleus
Twenty-five Erlenmeyer flasks (250 ml), each containing 100 ml of PDB media supplemented with sucrose 3% (w/v), ammonium sulfate 0.75% (w/v), at initial pH value 4 and the flasks, were incubated at 30 °C for 6 days under static conditions. After the incubation period, the culture was collected and filtered by muslin cloth, then the clear filtrate was collected and subjected to extraction by Ethyl acetate triple (1:1), then the extraction solvent was dried, using vacuum under reduced pressure at 40 °C, and the crude extract was collected.
Separation of active compounds by thin-layer chromatography
Aluminum sheets (20×20 cm) with pre-coated silica gel 60 F254 (Merck, Germany) was used to separate active metabolites. The crude extract was applied in the form of bands at 1 cm from the bottom of the plate, using a capillary tube. Subsequently, the thin-layer chromatography (TLC) plate was then placed in a glass jar previously saturated with a developing solvent system of toluene to ethyl acetate to formic acid (6:3:1) (v/v). The glass jar was covered tightly. The developing system was left to rise on the TLC plate, until it reached the solvent front. The developed chromatogram was examined visually and observed under short (254 nm) and long (366 nm) wavelength ultraviolet light (UV). The retention factor (Rf) values of the separated spots were determined subsequently. For the detection of bioactive compounds, each band was scraped off the plate, eluted with methanol, and filtered, using a filter paper. The concentration of LC50 was prepared from each band and subjected to assay against T. urticae.
Separation and identification of the most active compound/s
Liquid column chromatography–mass spectrometry (LC-MS/MS) was used to separate active compounds in the highest active band. The band (Rf3) was subjected to identification on Agilent LC-MS/MS (Agilent Technologies, USA) that consists of C18 column model G1316A equipped with a diode array detector (model G1315D) and coupled to a 6420 Triple Quadrupole mass spectrometer equipped with an electrospray ionization (ESI) source operating in positive ionization mode. The mobile phase consisted of 5 mM ammonium formate containing 0.1% formic acid (solvent A) and acetonitrile (solvent B). A total of 5 μl of the sample was injected. The gradient elution program for LC analysis was applied as follows: 0–2 min, 40% B; 2–12 min, 40–85% B; 12–15 min, 85–90% B; 15–35 min, 90–95% B at a flow rate of 0.3 ml min−1 and the column temperature was set to 35 °C. The MS analysis used full-scan mode with the mass range set to 100–1200 m/z in positive and mode. The conditions of the ESI source were as follows: drying gas, high-purity nitrogen (N2), drying gas temperature was set to 350 °C, drying gas flow-rate was 11 L/min, the nebulizing gas (N2) was set at a pressure of 45 psi, a potential of 3500 V for the positive ionization mode was applied to the tip of the capillary, and the fragmentation voltage was 130 V. The acquisition and data analysis were controlled using Agilent LC-MS Software (Agilent, USA). The identification of the separated compounds was carried out by comparing their retention times and mass spectra provided by ESI-MS with the data obtained from previous studies that are carried out on the same A. melleus as well as those of authentic standards when available.
Data analysis
Control is corrected according to Abbott’s formula (Abbott 1925).
where T = dead mites in treatment and C = dead mites in control.
Data obtained from each dose-response bioassay were subjected to probit analysis (Finney 1971) to estimate LC50 values. For mortality and ovicidal activity experiments, the number of dead mite individuals and un-hatched eggs were counted and analyzed by one-way analysis of variance (ANOVA) (P = 0.05). Means were compared by Duncan’s test (Duncan 1955). Also, mortality percentages were calculated and corrected according to Abbott’s formula (Abbott 1925).
Results and discussion
Isolation and primary assay of fungi
Sixty-two fungal strains were isolated from the rhizosphere of different plants and tested for their control activity against the females of T. urticae. Thirty-four isolates showed insignificant differences in their efficiency, while 28 showed a considerable control activity against spider mite (Table 1). Among the effective isolates, KF23, KF40, KF45, and KF9, that showed the highest mortality rates, were selected for further experiments. Culture filtrates of KF23, KF9, KF40, and KF45 resulted in 53.69, 73.83, 50.34, and 67.11% mortality rate, respectively, 5 days after exposure. Similarly, the culture filtrates of B. bassiana and M. anisoplae highly reduced mite population (Yun et al. 2017). The obtained results were relatively higher than mortality rates of HtCRMB isolate of Hirsutella thompsonii Fisher that only exhibited 55.90% on T. urticae and citrus rust mite, Phyllocoptruta oleivora Ashmead, 9 days after application (Aghajanzadeh et al. 2006). Mehdi (2006) studied the effect of culture filtrates of some fungal isolates and found that the highest contact toxicities were 52.4, 52.4, 48.4, and 50.4%, exhibited by Alternaria alternata, A. terreus, Trichoderma viridae, and Eurotium eurotiorum, followed by A. pluriseptata, Stachybotrys atra, Trichoderma harzianum, T. koningii, and Ulocladium chartarum showed mortality rates of 42.5, 42.5, 46.5, 43.5, and 39.6%, respectively.
Morphological identification of the most effective isolates
Isolate KF23 grown on Czapek’s agar media forming colony covered with dense layer of sclerotia and conidiophores absent or produced sparingly in the center. Conidial structures are more abundant in older cultures. Sclerotia were yellow to brown and hard and slightly rounded. This isolate was confirmed as Aspergillus melleus Yukawa at the Mycological Centre, Faculty of Science, Assiut University, Egypt. Isolate KF40 was grown on Czapek’s medium forming a colony, colored dark orange yellow with yellow reverse. The colony attained a diameter of 31 mm after 7 days at 25 °C. Isolate KF40 was identified as A. terreus, according to Raper and Fennell (1965). Isolate KF9 was grown on potato dextrose agar medium producing flattened, black gray colony with white, lobed edges and dark yellow reverse. Growth was relatively slow and characterized by an abundance of olive green ascocarps with ascomal hairs. Spores were pale brown and lemon-shaped. These characters are matching well with Chaetomium globosum (Domsch et al. 1980). The isolate KF45 grew rapidly on Czapek’s agar forming green colony with purple shade. Cleistothecia are scattered on the colony, green-colored, and globose. Microscopic examination showed that ascospores were orange with two equatorial crests and in addition to the presence of globose Hülle cells. Accordingly, isolate KF45 was Emericella nidulans (Raper and Fennell 1965).
Effect of crude extract of the selected fungi on adults of T. urticae
Mortality rates exhibited by the ethyl acetate extract of each of the selected isolates are shown in Table 2. LC50 values on females exhibited by A. melleus, C. globosum, A. terreus, and E. nidulans were 10.27, 22.40, 33.05, and 14.68 mg/ml, respectively. It seems that there are no reports on the acaricidal activity of A. melleus or E. nidulans. The present results proved that the obtained isolates possessed a concentration-dependent strong acaricidal efficacy, since they caused high mortality rates in mite population at the concentration of 30 mg/ml where 91.67% and 81.67% were killed by A. melleus and E. nidulans. Also, C. globosum and A. terreus showed mortality rates of 76.67% and 56.67% at the highest concentration (30 mg/ml), respectively. Similarly, the crude extract of Hypocrella raciborskii Zimm at a concentration of 3% (w/v) showed residual toxicity of 80% in 3 days after application (Buttachon and Kijjoa 2013). Santamarina et al. (1987) found that the crude extract (10 mg/ml) of P. funiculosum caused 100% mortality in the population of Panonychus ulmi on the second day after treatment. Similar results were obtained by Jimenez et al. (1993) who tested P. funiculosum on adult females of T. urticae and recorded a mortality rate of 100% after 3 days.
Effect of crude extracts of the selected fungi on eggs of T. urticae
LC50 values exhibited on eggs were 8.81, 11.05, 23.17, and 11.66 mg/ml, when eggs of the spider mite were subjected to crude extracts of A. melleus, C. globosum, A. terreus, and E. nidulans, respectively (Table 3). Interestingly, the evaluated isolates showed a higher toxicity towards eggs than adults of T. urticae. Crude extracts (30 mg/ml) of A. melleus, E. nidulans, and C. globosum inhibited completely the hatchability of eggs. Similarly, fungal filtrates studied by Mehdi (2006) had more toxic action on eggs of T. urticae than adults. He also found that the filtrates of Alternaria alternate, A. pluriseptata, Aspergillus terreus, Trichoderma harzianum, T. koningii, T. viride, Eurotium eurotiorum, Ulocladium chartarum, and Stachybotrys atra inhibited hatchability by an average of 65.5, 58.3, 65.5, 60.6, 60.6, 54.1, 40, 52, 49.1, and 49.1%, respectively. Santamarina et al. (1987) reported that the crude extract (10 mg/ml) of P. funiculosum caused 52.17% reduction in eggs hatching of Panonychus ulmi.
Identification of A. melleus
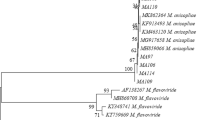
A. melleus showed the minimum LC50 value, so it was selected for further investigation. The identification of A. melleus was confirmed by DNA sequencing. The alignment of the ITS sequences of fungal isolate KF23 was done by the National Center for Biotechnology Information (NCBI) database. The phyllogenetic analysis showed that KF23 most resemble A. melleus strain CBS 112786 (Accession No. FJ491567) with 99% homology (Fig. 1).

Phylogenetic tree of the ITS region sequences of the fungal isolate (Aspergillus melleus KF23) aligned with closely related sequences accessed from Gene Bank. Branches show bootstrap values (%), random seed (226,649), no. of replicates = 100; scale bar indicates nucleotides substitutions per site
Identification of active metabolites of A. melleus
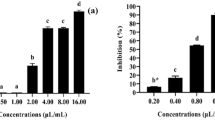
Partial separation of active compounds in A. melleus crude extract by TLC resulted in five major bands. Four of them were visualized at long UV wavelength at 366 nm and one was seen under short UV wavelength at 254 nm. A band colored florescent blue at a Rf1 value of 0.97, florescent green band at Rf2 value of 0.72, yellow band at Rf3 value 0.55, blue band at Rf5 of 0.1, and brown band at Rf4 0.43. Each of the five bands was eluted by methanol and subjected to toxicity bioassay against females of T. urticae. The mortality rates are as shown in Fig. 2. The Rf3 band exhibited the highest mortality percent of 57.05%, followed by Rf1 (28.52%), Rf2 (10.07%), Rf5 (11.74%), and Rf4 (8.39%). Therefore, it was selected for further purification and identification by LC-MS/MS.

Lethal activity of TLC fractions of ethyl acetate extract of A. melleus on T. urticae. Error bars are corresponding to standard error. Different letters (superscript a and b) indicate a significant difference according to Duncan’s multiple range test
Liquid chromatography/mass spectrometry
The metabolites in the active fraction separated by TLC, which had Rf3 of 0.55, were analyzed by LC-MS/MS and identified by comparing to the same authentic compounds. From LC/MS data, 13 compounds were detected (Fig. 3). Only five compounds were identified, while the others are needed further studies for identification.

Chromatogram shows the peaks separated by LC from the band (Rf3)
The first identified one appeared on liquid chromatography profile at a retention time (Rt) of 4.854 min had a molecular mass of 306 resulting from ESI-MS m/z = 306 [M+]; it might be 7-oxocurvularin (Fig. 4) and it had the molecular formula C16H18O6. The second identified compound appeared at Rt 10.399 min and a molecular mass of 679 deduced from m/z of 679 [M+] might be a nodulisporic acid with fragment ion at m/z = 702 [M + Na] deduced from m/z 701 [M − 1] (Fig. 5). The molecular formula of the nodulisporic acid is C43H53NO6. The third peak detected at Rt 15.298 min and a molecular weight of 381 deduced from m/z 382 [M + 1] might be mellamide (Fig. 6) which had the molecular formula of C23H31N3O2. The fourth identified peak, Rt (18.025 min) with molecular weight of 431, resulted from m/z = 431 and fragment ion of m/z = 358 (Fig. 7). This peak might be ochratoxin C with a molecular formula of C22H22NO6Cl. The last identified one appeared at Rt 21.243 min with a molecular mass of 301 deduced from m/z = 300 [M − 1] and fragment ion at m/z = 258 [M + 2], 149 [M − 1], and 121[M − 3] (Fig. 8). This peak might be 6-(4′-hydroxy-2′-methyl phenoxy)-(−)-(3R)-mellein (C19H24O3). Further investigations are required to characterize the other eight compounds. It was reported that A. melleus produces a wide range of polyketides (Garson et al. 1984) and naphthoquinone pigments such as xanthomegnin, viomellein, and viopurpurin (Durley et al. 1975). Similarly, Ondeyka et al. (2003) reported an insecticidal activity of mellamide, xanthomegnin, viomellein, and ochratoxin A isolated from A. melleus against Aedes aegypti. Also, A. melleus produces aspyrone that exhibited nematicidal activity against Pratylenchus penetrans by 80% at a concentration of 300 mg/l (Kimura et al. 1996).

Mass chart shows the fragment ions of 7-oxocurvularin

Mass chart shows the fragment ions of nodulisporic acid

Mass chart shows the fragment ions of mellamide

Mass chart shows the fragment ions of ochratoxin C

Mass chart shows the fragment ions of 6-(4′-hydroxy-2′-methyl phenoxy)-(−)- (3R)-mellein
Conclusion
A. melleus, E. nidulans, C. globosum, and A. terreus showed promising acaricidal activities against the two-spotted spider mite, T. urticae, females and eggs. Further investigations are still needed to identify their bioactive secondary metabolites and to evaluate their efficacy under greenhouse and field trials.
Availability of data and materials
The data and material of this manuscript are available on reasonable request.
Abbreviations
- ANOVA:
-
Analysis of variance
- DNA:
-
Deoxyribonucleic acid
- ESI:
-
Electrospray ionization
- ITS:
-
Internal transcribed spacer
- LC50 :
-
The median lethal concentration
- LC-MS/MS:
-
Liquid chromatography–mass spectrometry
- NCBI:
-
National Center for Biotechnology Information
- PCR:
-
Polymerase chain reaction
- PDA:
-
Potato dextrose agar
- Rf:
-
Retention factor
- rpm:
-
Revolutions per minute
- Rt:
-
Retention time
- TLC:
-
Thin layer chromatography
- UV:
-
Ultraviolet
References
Abbott WS (1925) A method of computing the effectiveness of an insecticide. J Econ Entomol 18(2):265–267
Aghajanzadeh S, Mallik B, Chandrashekar SC (2006) Toxicity of culture filtrate of Hirsutella thompsonii Fisher against citrus rust mite, Phyllocoptruta oleivora Ashmead (Acari: Eriophyidae) and two spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae). Int J Agric Biol 8(2):276–279
Attia S, Grissa KL, Lognay G, Bitume E, Hance T, Mailleux AC (2013) A review of the major biological approaches to control the worldwide pest Tetranychus urticae (Acari: Tetranychidae) with special reference to natural pesticides. J Pest Sci 86(3):361–386
Buttachon S, Kijjoa A (2013) Acaricidal activity of Hypocrella raciborskii Zimm. (Hypocreales: Clavicipitaceae) crude extract and some pure compounds on Tetranychus urticae Koch (Acari: Tetranychidae). Afr J Microbiol Res 7(7):577–585
Culliney TW (2014) Crop losses to arthropods. In: Pimentel D, Peshin R (eds) Pesticide problems. Integrated pest management, vol 3. Springer, Dordrecht, pp 201–225
Domsch KH, Gams W, Anderson TH (1980) Compendium of soil fungi. Academic, London
Duncan DB (1955) Multiple range and multiple F-tests. Biometrics 11(1):1–42
Durley RC, MacMillan J, Simpson TJ, Glen AT, Turner WB (1975) Fungal products. Part XIII. Xanthomegnin, viomellin, rubrosulphin, and viopurpurin, pigments from Aspergillus sulphureus and Aspergillus melleus. J Chem Soc Perkin Trans 1(2):163–169
Fasulo TR, Denmark HA (2000) Two spotted spider mite, Tetranychus urticae Koch (Arachnida: Acari: Tetranychidae). University of Florida Cooperative Extension Service, Institute of Food and Agricultural Sciences, EDIS http://entnemdept.ufl.edu/creatures/orn/twospotted_mite.htm
Finney DJ (1971) Probit Analysis, 3rd edn. Cambridge University Press, London
Garson MJ, Staunton J, Jones PG (1984) New polyketide metabolites from Aspergillus melleus: structural and stereochemical studies. J Chem Soc Perkin Trans 1:1021–1026
Horikoshi R, Goto K, Mitomi M, Oyama K, Sunazuka T, Ōmura S (2017) Identification of pyripyropene A as a promising insecticidal compound in a microbial metabolite screening. J Antibiot 70(3):272
Jimenez M, Atienza J, Hernandez E (1993) Bioproduction of an extract from Penicillium funiculosum Thom with activity against Ceratitis capitata and Tetranychus urticae. Appl Microbiol Biotechnol 39(4–5):615–616
Kimura Y, Nakahara S, Fujioka S (1996) Aspyrone, a nematicidal compound isolated from the fungus, Aspergillus melleus. Biosci Biotechnol Biochem 60(8):1375–1376
Marcic D (2012) Acaricides in modern management of plant-feeding mites. J Pest Sci 85(4):395–408
Mehdi HMR (2006) The effect of some fungi in biocontrol in two spotted spider mite Tetranychus urticae (Koch.) (Tetranychidae: Acari). J Basrah Res (Sciences) 32(2B):12–26
Neal JW Jr, Lindquist RK, Gott KM, Casey ML (1987) Activity of the thermostable p-exotoxin of Bacillus thuringiensis Berliner on Tetranychus urticae and T. cinnabarinus. J Agric Entomol 4(1):33–40
Ondeyka JG, Dombrowski AW, Polishook JP, Felcetto T, Shoop WL, Guan Z, Singh SB (2003) Isolation and insecticidal activity of mellamide from Aspergillus melleus. J Ind Microbiol Biotechnol 30(4):220–224
Putter I, Mac Connell JG, Preiser FA, Haidri AA, Ristich SS, Dybas RA (1981) Avermectins: novel insecticides, acaricides and nematicides from a soil microorganism. Experientia 37(9):963–964
Ragavendran C, Natarajan D (2015) Insecticidal potency of Aspergillus terreus against larvae and pupae of three mosquito species Anopheles stephensi, Culex quinquefasciatus, and Aedes aegypti. Environ Sci Pollut Res 22(21):17224–17237
Raper KB, Fennell DI (1965) The genus Aspergillus. The Williams &Wilkins company, Baltimore (686 pages)
Roobakkumar A, Babu A, Kumar DV, Sarkar S (2011) Pseudomonas fluorescens as an efficient entomopathogen against Oligonychus coffeae Nietner (Acari: Tetranychidae) infesting tea. J Entomol Nematol 3(5):73–77
Santamarina MP, Jimenez M, Sanchis V, Garcia F, Hernandez E (1987) A strain of Penicillium funiculosum Thorn with activity against Panonychus ulmi Koch (Acar.: Tetranychidae). J Appl Entomol 103(1–5):471–476
Srivastava CN, Maurya P, Sharma P, Mohan L (2009) Prospective role of insecticides of fungal origin. Entomol Res 39(6):341–355
Vargas Mesina RR (1993) Thuringiensin toxicity to Tetranychus urticae Koch and Panonychus ulmi (Koch) (Tetranychidae) and effects on cuticle development of immature stages of T. urticae. Ph.D. Dissertation, Lincoln University, New Zealand
White TJ, Bruns T, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA gene for phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols: a guide to methods and applications. Academic, San Diego, pp 315–322
Wu Y, Liu X (1997) Toxicity and biological effect of abamectin on Tetranychus cinnabarinus in lab. Acta Agric Boreali Sin 12(1):108–111
Yun HG, Kim DJ, Lee JH, Ma JI, Gwak WS, Woo SD (2017) Comparative evaluation of conidia, blastospores and culture filtrates from entomopathogenic fungi against Tetranychus urticae. Int J Ind Entomol 35(1):58–62
Acknowledgements
Thanks are due to Prof. Mohamed Abdelaziz Balah, Plant Protection Department, Desert Research Center, for the interpretation of LC-MS/MS data and identification of the separated compounds.
Funding
There are no funding sources for this manuscript.
Author information
Authors and Affiliations
Contributions
The conception and design of the study were done by all authors; the last author (GAH) performed the experimental part under the supervision of MEO, AAAE, and MAN. The analysis and interpretation of the data were done by the first author (MEO) and AAAE. Both MAN and GAH prepared the manuscript draft; then the manuscript was revised for important intellectual content by MEO and AAAE. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This article does not contain any studies with human participants or animals.
Consent for publication
The manuscript has not been published in completely or in part elsewhere.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Osman, M.E., Abo Elnasr, A.A., Nawar, M.A. et al. Myco-metabolites as biological control agents against the two-spotted spider mite, Tetranychus urticae Koch (Acari: Tetranychidae). Egypt J Biol Pest Control 29, 64 (2019). https://doi.org/10.1186/s41938-019-0166-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41938-019-0166-0