
Abstract
Background
Small crustaceans in the order Anostraca that lack a carapace are commonly referred to as fairy shrimps. Eubranchipus is an Anostracan genus distributed across the Holarctic region. E. uchidai has been recorded only in Japan and has been considered the only species of Eubranchipus in the region.
Results
We obtained fairy shrimps from three previously unsampled locations (Shiretoko and Chokai in Japan, and Khanka in Russia). Each specimen shares morphological similarities with E. uchidai, but can be discriminated by the second antenna of males and the genital morphology of females. Based on these specimens, we describe three new species of fairy shrimp: Eubranchipus asanumai n. sp. from the banks of the Shiretoko Five Lakes (Japan: Hokkaido), Eubranchipus hatanakai n. sp. from the base of Mt. Chokai (Japan: Yamagata) and Eubranchipus khankanus n. sp. from the banks of Lake Khanka (Russia: Primorsky). Phylogenetic analysis of mitochondrial DNA reveals significant differences from other known Eubranchipus species, including E. uchidai.
Conclusion
Both morphological and molecular analyses show that the three new species are similar to E. uchidai but constitute distinct species. Pronounced differences in female morphological features correspond to variability of the amplexus structure previously reported in the genus.
Similar content being viewed by others


Background
Crustaceans without a carapace in the order Anostraca (literally, “without a plate or shell”) are commonly referred to as “fairy shrimps” [1, 2]. Fairy shrimps occur in several types of bodies of water, including permanent and temporary salt lakes and alpine, tundra or rock pools [2, 3].
The Anostracan genus Eubranchipus Verill, 1870 is distributed across the Holarctic region [3,4,5,6]. Eighteen species are known so far [3, 6]. In Japan, Eubranchipus uchidai (Kikuchi, 1957) has been recorded from only two localities: Ishikari in Hokkaido and Shimokita in Aomori. This species was first described in Ishikari from male individuals [7] (Fig. 1) and later from female individuals [8]. Ooyagi [9] reported the species in Shimokita, more than 200 km across the sea to the south.

Geographical locations of fairy shrimp sample sites used in this study. Although Eubranchipus uchidai has also been reported from Shimotika [9], we were unable to obtain specimens from that site. A map was obtained from a personal freely available web site (http://shanghai-cool.jugem.jp/?eid=5) and was processed
We discovered three new localities (Shiretoko, Chokai and Khanka) of temporary snowmelt pools inhabited by fairy shrimps. Each locality is at a distance of more than 700 km from the other two (Fig. 1), and each yielded one specific type of fairy shrimp. These share similar male gonopods (Fig. 2): cylindrical, with a chitinized, subacute proximal, medial projection, with concave sides projecting a chitinized, acute, long spine laterally. This combination of characteristics is typical of the genus Eubranchipus [4, 5, 10]. In addition, they are similar to E. uchidai in their frons, second antenna apophyses, antennal appendage and distal antennomere.

Male gonopods. (a) Ishikari specimen, (b) Shiretoko specimen, (c) Chokai specimen, (d) Khanka specimen. Scale bars = 0.25 mm
The existing descriptive studies of E. uchidai noted above [7, 8] present a number of unresolved problems. Drawings of male individuals in both papers do not match, especially with regards to the second antennae, which typically have been used to identify Anostracans at the species level [11, 12]. Female descriptions lack genital morphology. There is thus not yet a clear morphological description for E. uchidai. We therefore review E. uchidai based on the materials from Ishikari (the type locality), provide a comparison with materials from the three new localities, and finally describe three new species.
While several molecular phylogenetic studies of fairy shrimp species have been carried out [13,14,15,16,17,18,19,20,21,22,23,24], there as yet exist none performed on Japanese specimens, and little is known about phylogenetic relationships among Eubranchipus species from Eurasia and North America and insular species in Japan. To clarify the systematic position of the newly found species relative to E. uchidai, we therefore also determined the nucleotide sequences of a part of the cytochrome oxidase subunit 1 (COI) and 16S ribosomal RNA (16S rRNA) in the mitochondrial genome, and derived phylogenetic relationships from these data.
Methods
Specimens
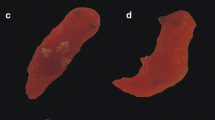
We collected fairy shrimps from temporary pools in Ishikari, Shiretoko (Fig. 3a and b), Chokai (Fig. 4a) and Khanka (Fig. 5a) by dip net and preserved them by immersion in ethyl alcohol. At the three new localities, photographs of live shrimp were taken beforehand (Fig. 3c, d, Fig. 4b, c, Fig.5b and c). Collection details are as follows.
-
1.
30 male, 29 female: Japan: Hokkaido: Ishikari: Ishikari coast: 21 May 2013: M. Hamasaki.
-
2.
4 male, 4 female: Japan: Hokkaido: Shari: Shiretoko Five Lakes: 29 May 2016: N. Takahashi, and 22 male, 22 female: Japan: Hokkaido: Shari: Shiretoko Five Lakes: 20 May 2017: N. Takahashi.
-
3.
20 male, 20 female: Japan: Yamagata: Yuza: base of Mt. Chokai: 15 April 2013: Y. Hatanaka.
-
4.
3 male, 2 female: Russia: Primorsky: Spassky: Lake Khanka: 29 April 2007: Y. Nagahata.

Shiretoko collection site and specimens. (a) Habitat 1, (b) Habitat 2, (c) Live male, (d) Live female

Chokai collection site and specimens. (a) Habitat, (b) Live male, (c) Live female

Khanka collection site and specimens. (a) Habitat, (b) Live male, (c) Live female
Morphological characterization
We examined the morphology of the specimens with a stereo microscope, created manual drawings and carried out comparisons with the literature. At the genus level, we referred to [4, 5, 10, 25,26,27,28]; at the species level, we referred to [6,7,8, 26, 29,30,31,32,33,34,35,36,37]. We also used microscopic camera and created stacked images.
PCR and DNA sequencing for COI and 16S rRNA
Two individuals each from the three new location (Shiretoko, Chokai, and Khanka) and 54 E. uchidai individuals from Ishikari were used. Genomic DNA was extracted from muscle tissues using the standard protocol of the DNA Extractor FM Kit (Wako Pure Chemical Industries Ltd., Osaka, Japan).
The primers for COI were khCOI-F (5′-TTG ATC AGG TAT GGT AGG CAC-3′) and khCOI-R (5′-CGT GTT CAG ATT ACG ATC CG-3′). These were designed based on relatively well-conserved regions from a multiple alignment of COI sequences from species in other Anostraca species (e.g., Artemia, Eubranchipus and Streptocephalus because COI primers designed by some previous studies were ineffective with some specimens used in the present study. The primers for the 16S rRNA were kh16S-F (5′-GCC TGT TTA TCA AAA ACA TGG C-3′) and kh16S-R (5′-CGG TTT GAA CTC AGA TCA CG-3′). These primers were designed by modifying 16Sa (5′-CGC CTG TTT ATC AAA AAC AT-3′) and 16Sb (5′-CTC CGG TTT GAA CTC AGA TCA-3′) in [38]. Reactions for each set were performed in a 10.0 μl mixture containing approximately 10 ng genomic DNA, 0.5 μM of each primer, and 1× KAPA 2G Fast Multiplex PCR Kits HS (KAPA Biosystems, Wilmington, MA, USA). Thermal cycling was performed in a GeneAmp PCR system 9700 (Applied Biosystems, Waltham, MA, USA). A two-step PCR method was employed: 95 °C for 3 min, followed by 30 cycles of denaturation at 95 °C for 10 s, then annealing and extension at 60 °C for 2 min. The PCR products were confirmed by 1% agarose gel electrophoresis and purified using a High Pure PCR Product Purification Kit (Roche Diagnostics, Mannheim, Germany).
DNA sequencing was performed on the PCR products using the BigDye Terminator v1.1 Cycle Sequencing Kit and the ABI PRISM 310 Genetic Analyzer (Applied Biosystems, MA, USA). All sequencing was carried out at Yamagata University. Both strands of DNA were sequenced to confirm the sequence.
Although two specimens were sequenced for both COI and 16S from each of the three new locations, no nucleotide differences were found between the two sequences of any pair. We therefore assumed a single haplotype per location. In contrast, preliminary one-path sequencing for COI of E. uchidai indicated two types (44 for A type and 10 for B type as shown in Discussion) among 54 individuals. Thus two haplotypes are used for E. uchidai in the following phylogenetic analysis.
Phylogenetic analysis
COI nucleotide sequences for five Eubranchipus, six Chirocephalus, one Artemiopsis and one Branchinella species were retrieved from the DDBJ/EMBL/GenBank International Nucleotide Sequence Database, and were used in conjunction with our newly determined sequences for the phylogenetic analysis. Data for five Artemia species [24] were also included. Similarly, 16S rRNA nucleotide sequences were retrieved for three Eubranchipus, three Chirocephalus, one Artemiopsis, three Artemia and one Branchinella species.
Multiple alignments were performed using the MUSCLE program [39], implemented in MEGA7 software [40]. The maximum likelihood method [41] was used to construct phylogenetic trees of COI, 16S and COI + 16S data using the RAxML software [42] with the GTR + G model [43]. Bootstrap probabilities [44] were computed from 500 replicates. A Bayesian approach [45] implemented in the MrBayes package version 3.2 [46] with the GTR + G model, and a 10,000,000 generations series was used for phylogenetic inference of COI + 16S data. Phylogenetic networks were constructed using the Neighbor-Net method [47] with p-distance implemented in SplitsTree4 [48].
For a general overview of nucleotide differences among species, percentual pair-wise nucleotide difference per site for COI and 16S data was also estimated. Pair-wise p-distances in % were calculated using MEGA7.
Results
Eubranchipus uchidai (Kikuchi, 1957)
Here we review Eubranchipus uchidai (Fig. 6 and Fig. 7) based on the materials from the type locality at Ishikari. Morphological terminology predominantly follows [6, 25, 49].

Male Eubranchipus uchidai. (a) Left head anterior view, (b) Frons anterior view, (c) Posteromedial view of right antennal appendage with lateral view of left second antenna, (d) Right second antenna distal antennomere anteromedial view, (e) Right fifth leg ventral view, (f) Ventral view of head with posterior view of second antennas, (g) Left second antenna distal antennomere anterior view, (h) Left antennal appendage anterior view, (i) Left gonopod ventral view. Scale bars: A, C, F, H = 1 mm; B, D, E, G = 0.5 mm; I = 0.25 mm

Female Eubranchipus uchidai. (a) Left head anterior view, (b) Last three thoracic segments dorsal view, (c) Brood pouch ventral view, (d) Left side of brood pouch and abdomen lateral view. Scale bars: A, B = 0.5 mm; C, D = 1 mm
Male
Head (Fig. 6a and f) subspherical, with flattened ventral portion bearing a subtrapezoidal labrum. Frons (Fig. 6b) with a tiny, chitinized rostrum anteroinferiorly or absent. First antenna elongate, approximately the length of the second antenna proximal antennomere. Second antenna proximal antennomere subcylindrical, with a proximal, anteromedial conical apophysis. Apophysis chitinized, apically rounded and directed medially. Proximal antennomere with a posterobasal medial cone (Fig. 6f). Proximal antennomere bearing an anterior, proximal, lamellar antennal appendage (Fig. 6c and h) tapering from seven tenths of the length. Antennal appendage approximately the length of the second antenna and longer than its proximal antennomere. Antennal appendage with posterobasal medial-marginal spines directed posteriorly (Fig. 6c). Medial margin proximal fourth smooth, with remaining medial margin bearing short digitiform papillae continuously. Lateral margin proximal seventh smooth, with remaining lateral margin bearing long digitiform papillae continuously; papillae gradually increasing in size to distal fourth, reaching the length of the horizontal width of the adjacent appendage at the medial and then decreasing in size to apex. All digitiform papillae terminating in a short spine and studded shorter spines laterally. Second antenna proximal-distal joint thinly chitinized anteriorly, turning into an angulated sleeve during retraction of the distal antennomere. Second antenna distal antennomere (Fig. 6d and g) chitinized, curved as sickle, tapering apically. Distal antennomere medial margin center portion swollen medially, forming a concavity in both anterior and posterior sides (Fig. 6d). Anteromedial concavity continuing to the vertical distal end and posteromedial concavity joining to the distal end. Second antenna distal antennomere with a spiny, undeveloped, anterobasal bulge, and continuing with small spines along the ridge to the half of vertical distal. Distal antennomere medial margin with a subacute hook projection apically. Thoracic segments smooth, with eleven pairs of thoracopods on each side. Thoracopods serially homologous, increasing in size from the anterior pair to the fifth through eighth pairs, then decreasing in size posteriorly to the eleventh pair. Thoracopod V (Fig. 6e) with two preepipods, epipod, exopod, endopod and five endites. Preepipods and epipod lobiform, glabrous. Exopod oblong, margined with long plumose setae. Endopod broad, flabellate, margined with short, stout plumose setae. Endites I and II broad, rounded, and margined with numerous long plumose setae. Endites III, IV and V digitiform, margined with several stout plumose setae and several long plumose setae. Genital segments with a pair of gonopods ventrally. Gonopod (Fig. 6i) with a proximomedial, chitinized, subacute projection, approximately one third the length of the gonopod proximal rigid portion. Proximal rigid portion cylindrical, slightly constricted in medial, with concavity in superior side. Concavity of slightly shorter width than the proximal rigid portion. Concavity projects a distal eversible portion from lateral side. Distal eversible portion acute, spinulate, directed superolaterally, same length as proximal rigid portion, and studded with small denticles. Abdomen smooth, cylindrical, with six segments. Telson cylindrical. Cercopods typical for genus.
Female
Head (Fig. 7a) subspherical. First antenna elongate, approximately 1.5 times the length of the second antenna. Second antenna oblong, with a long spine apically and a short spine medially. Thorax smooth, but last three thoracic segments bearing a pair of dorsolateral lobes on each segment (Fig. 7b and d). Ninth lobe rounded, smaller than tenth and eleventh lobes. Tenth and eleventh linear lobes, same in size each other and project posteriorly. Thoracopods as in male. Brood pouch (Fig. 7c and d) subcylindrical, with tapered apex reach to the second abdominal segment. Apex half of the width of the abdomen. Abdomen (Fig. 7d) cylindrical, and with a pair of posterolateral spines on each segment from the last genital segment to the third abdominal segment. Posterolateral spines in third abdominal segment smaller. Telson and cercopods as in male.
Egg
Spherical, diameter approximately 420 μm, quite shallow engraving, densely covered with small scale-like projections 10–20 μm wide (Fig. 8a).

Eggs whole view. (a) E. uchidai, (b) E. asanumai n. sp. (c) E. hatanakai n. sp. (d) E. khankanus n. sp. Scale bars = 100 μm
Differential diagnosis
E. uchidai shares similar morphology with Eubranchipus claviger (Fisher, 1851) and Eubranchipus rostratus (Daday, 1910), although males of this species are identified by tiny rostrum (differs from E. rostratus); conical, apically rounded apophyses (differs from E. rostratus); antennal appendage posterior side with medial-marginal spines along less than one third of its length (differs from E. claviger); antennal appendage medial margin with digitiform papillae along more than three fifth of its length (differs from E. rostratus); and undeveloped anterobasal bulge of distal antennomere. Females can be identified by dorsolateral linear lobes of the last two thoracic segments, and number of posterolateral spines on abdomen.
Comments
Materials on male individuals from Ishikari matched Kikuchi’s original description except for one apophysis of the second antenna, as depicted in Fig. 2b of Moriya [8]. From this, we conclude that he drew the male left second antenna together with the unnecessary head and apophysis of the right second antenna in Fig. 2c. This combination was ambiguously indicated as “male left second antenna” in the caption, confusingly suggesting that the male second antenna possesses three apophyses.
Kikuchi [7] also drew short spines on the antennal appendage medial margin root in Fig. 2b (Kikuchi’s linear edge). However, this does not appear to be an accurate depiction, as spines project slightly medially along the medial margin of the antennal appendage posterobasally and are directed posteriorly. These posterobasal medial-marginal spines cannot be observed from the anterior side. We confirmed these spines for all specimens sampled in this study, including the three new species.
Description of three new species
Here we describe three new species of fairy shrimp: Eubranchipus asanumai n. sp. (Fig. 9), Eubranchipus hatanakai n. sp. (Fig. 10) and Eubranchipus khankanus n. sp. (Fig. 11). Both morphological and molecular analyses showed that the three new species are similar to E. uchidai but constitute distinct species.

Eubranchipus asanumai n. sp. (a) Male, left side of head anterior view, (b) Female, left side of head anterior view, (c) Female, last three thoracic segments dorsal view, (d) Male, right gonopod ventral view, (e) Female, left side of brood pouch and abdomen lateral view, (f) Male, left second antenna distal antennomere anterior view, (g) Male, left antennal appendage anterior view, (h) Female brood pouch ventral view. Scale bars A, E, G, H = 1 mm; B, C, F = 0.5 mm; D = 0.25 mm

Eubranchipus hatanakai n. sp. (a) Male, left side of head anterior view, (b) Female, left side of head anterior view, (c) Female, last three thoracic segments dorsal view, (d) Male, left gonopod ventral view, (e) Female, left side of brood pouch and abdomen lateral view, (f) Male, left second antenna distal antennomere anterior view, (g) Male, left antennal appendage anterior view, (h) Female brood pouch ventral view. Scale bars A, E, G, H = 1 mm; B, C, F = 0.5 mm; D = 0.25 mm

Eubranchipus khankanus n. sp. (a) Male, left side of head anterior view, (b) Female, left side of head anterior view, (c) Female, last three thoracic segments dorsal view, (d) Male, left gonopod ventral view, (e) Female, left side of brood pouch and abdomen lateral view, (f) Male, right second antenna distal antennomere anterior view, (g) Male, left antennal appendage anterior view, (h) Female brood pouch ventral view. Scale bars A, E, G, H = 1 mm; B, C, F = 0.5 mm; D = 0.25 mm
Class Branchiopoda Latreille, 1817
Order Anostraca Sars, 1867
Family Chirocephalidae Daday, 1910
Genus Eubranchipus Verill, 1870
Eubranchipus asanumai n. sp. Takahashi
(LSID: urn:lsid:zoobank.org:act:FCB8F6EA-3E26-4BF7-8734-5ECA341A6B62)
Diagnosis - male
Head (Fig. 9a), frons, first antenna, second antenna proximal antennomere, apophysis and posterobasal medial cone as in E. uchidai. Second antenna proximal antennomere bearing an anterior, proximal, lamellar antennal appendage (Fig. 9g) tapering from the medial point onward. Antennal appendage approximately the length of the second antenna and longer than its proximal antennomere. Antennal appendage with posterobasal medial-marginal spines as in E. uchidai. Medial margin proximal fourth smooth, with remaining medial margin bearing short digitiform papillae and short spines sparsely. Lateral margin proximal eighth smooth, with remaining lateral margin bearing long digitiform papillae continuously; papillae gradually increasing in size to distal fifth, reaching 0.8 times the length of the horizontal width of the adjacent appendage at the medial, then decreasing in size to apex. All digitiform papillae ornamented with spines as in E. uchidai. Second antenna proximal-distal joint as in E. uchidai. Second antenna distal antennomere (Fig. 9f) chitinized, curved, tapering, with concavity, as in E. uchidai. Distal antennomere with a spiny, small anterobasal projection and continuing with spines along the ridge as in E. uchidai. Distal antennomere medial margin with an acute hook projection apically. Thorax, thoracopods, gonopod (Fig. 9d), abdomen, telson and cercopods as in E. uchidai.
Diagnosis - female
Head (Fig. 9b), first antenna, and second antenna as in E. uchidai. Thorax smooth, but last three thoracic segments bearing a pair of dorsolateral lobes on each segment (Fig. 9c and e). Ninth lobe rounded, smaller than tenth and eleventh lobes. Tenth and eleventh oval lobes projected posteriorly. Tenth lobe the largest. Thoracopods as in male. Brood pouch (Fig. 9e and h) subspherical with tapered apex reaching to the fourth abdominal segment. Apex a quarter the width of the abdomen. Abdomen (Fig. 9e) cylindrical, and with a pair of posterolateral spines on each segment from the last genital segment to the fourth abdominal segment. Posterolateral spines in fourth abdominal segment smaller. Telson and cercopods as in male.
Egg
Spherical, diameter approximately 380 μm, shallow engraving, densely covered with small scale-like projections 10–20 μm wide (Fig. 8b).
Authorship
N. Takahashi described E. asanumai n. sp.
Etymology
E. asanumai n. sp. is named after Takao Asanuma, a botanist of Shiretoko. N. Takahashi sincerely respects his steady, perseverant and pioneering work in areas with high brown bear density.
Type materials
Holotype: 1 male (15 mm), Japan: Hokkaido: Shari: Shiretoko Five Lakes, 20 May 2017, N: 44°07′53.5″, E: 145°04′56.5″, N. Takahashi, deposited Hokkaido University Museum type no. Invertebrate Collection of the Hokkaido University Museum (ICHUM) 5494. Paratypes: 2 males (15, 15 mm) and 3 females (16, 16, 17 mm), same collection data as holotype, deposited Hokkaido University Museum type nos. ICHUM 5495–5499; 2 males (17, 15 mm) and 2 females (12, 14 mm), Japan: Hokkaido: Shari: Shiretoko Five Lakes, 29 May 2016, N: 44°07′29.1″, E: 145°04′33.7″, N. Takahashi, deposited Lake Biwa Museum type nos. 1,430,008,288–8289.
Accession numbers in the DDBJ/EMBL/GenBank International Nucleotide Sequence Database are LC314404 for COI, and LC314405 for 16S rRNA, taken from an individual obtained from the same locality as the holotype. Here these sequences are considered as topogenetypes (proposed by [50]) of E. asanumai n. sp.
Type locality
Three hundred meters north of the outskirts of a trekking path (N: 44°07′53.5″, E: 145°04′56.5″, Fig. 3a) and beside a wooden walkway (N: 44°07′29.1″, E: 145°04′33.7″, Fig. 3b) in Shiretoko Five Lakes park area. The pool shown in Fig. 3a is in a forest of Quercus crispula Blume var. crispula, Betula ermanii Cham, and Abies sachalinensis (F. Schmidt) Mast. var. sachalinensis, and covered by a Sasa kurilensis (Rupr.) Makino & Shibata thicket more than two meters deep. The pool shown in Fig. 3b is in a grassland of low Sasa senanensis (Franch. & Sav.) Rehder.
Distribution and habitat
E. asanumai n. sp. is known only from the type locality within the Shiretoko National Park and World Heritage Area. Shiretoko Five Lakes and its surroundings consist of hummocks that are the remains of a debris avalanche about 4000 years ago [51] and provide a microrelief that supports snowmelt pools in spring. The species has not been observed co-occurring with other fairy shrimps.
Eubranchipus hatanakai n. sp. Takahashi & Hamasaki
(LSID: urn:lsid:zoobank.org:act:9D75832C-BC56-4131-A760-DB594B9C3D76)
Diagnosis - male
Head (Fig. 10a), frons, first antenna as in E. uchidai and E. asanumai n. sp. Second antenna proximal antennomere subcylindrical, with a proximal anteromedial, tubular apophysis. Apophysis chitinized, apically rounded, and directed anteriorly. Proximal antennomere with a posterobasal medial cone as in E.uchidai and E.asanumai n. sp. Proximal antennomere bearing an anterior, proximal, lamellar antennal appendage (Fig. 10g) tapering from seven tenths of the length. Antennal appendage shorter than the second antenna but longer than its proximal antennomere. Antennal appendage with posterobasal medial-marginal spines as in E. uchidai and E. asanumai n. sp. Medial margin proximal fifth smooth, with remaining medial margin bearing short digitiform papillae and short spines sparsely. Lateral margin proximal eighth smooth, with remaining lateral margin bearing long digitiform papillae continuously; papillae gradually increasing in size to distal fourth, reaching 1.4 times the length of the horizontal width of the adjacent appendage at the medial, then decreasing in size to apex. All digitiform papillae ornamented with spines as in E. uchidai and E. asanumai n. sp. Second antenna proximal-distal joint as in E. uchidai and E. asanumai n. sp. Second antenna distal antennomere (Fig. 10f) chitinized, curved and with concavity, as in E. uchidai and E. asanumai n. sp. but more elongating to the distal end than these two species. Second antenna distal antennomere with an anterobasal projection and continuing with spines, as in E. asanumai n. sp. Distal antennomere medial margin without a hook projection apically. Thorax, thoracopods, gonopod (Fig. 10d), abdomen, telson and cercopods as in E. uchidai and E. asanumai n. sp.
Diagnosis - female
Head (Fig. 10b), first antenna, second antenna as in E. uchidai and E. asanumai n. sp. Thorax smooth, but last three thoracic segments bearing a pair of dorsolateral lobes on each segment (Fig. 9c and e). Ninth lobe rounded, smaller than tenth and eleventh lobes. Tenth oval lobe and eleventh lanceolate lobe projected posteriorly. Eleventh lobe cover half of the genital segments. Thoracopods as in male. Brood pouch (Fig. 9e and h) subcylindrical with tapered apex reaching to the third abdominal segment. Apex a quarter the width of the abdomen. Abdomen (Fig. 10e) cylindrical, and with a pair of posterolateral spines on each segment from the last genital segment to the second abdominal segment. Telson and cercopods as in male.
Egg
Spherical, diameter approximately 350 μm, bearing burrs at 10 μm intervals (Fig. 8c).
Authorship
N. Takahashi and M. Hamasaki described E. hatanakai n. sp.
Etymology
E. hatanakai n. sp. is named after Yushi Hatanaka, who discovered this species.
Type materials
Holotype: 1 male (22 mm), Japan: Yamagata: the base of Mount Chokai (Chokai-zan), 15 April 2013, N: 39°05′24.06″, E: 139°53′52.3″, Y. Hatanaka, deposited Hokkaido University Museum type no. ICHUM 5500. Paratype: 1 female (14 mm), same collection data as holotype, deposited Hokkaido University Museum type no. ICHUM 5501.
Accession numbers in the DDBJ/EMBL/GenBank International Nucleotide Sequence Database are LC314402 for COI, and LC314403 for 16S rRNA, taken from an individual obtained from the same locality as the holotype. Here these sequences are considered as topogenetypes (proposed by [50]) of E. hatanakai n. sp.
Type locality
A temporary snowmelt pool in a reedy marsh in secondary forest of Quercus spp. and Zelkova serrata (Thunb.) Makino, covered with Sasa spp., at the base of Mount Chokai (Fig. 4a).
Distribution and habitat
E. hatanakai n. sp. is known only from the type locality, from temporary pools created by snowmelt in forest groves on the south side of the base of Mount Chokai. This area is a lava-plateau formed by volcanic activity. The species has not been observed co-occurring with other fairy shrimps.
Eubranchipus khankanus n. sp. Takahashi & Moriya
(LSID: urn:lsid:zoobank.org:act:9BB1B0ED-176B-468F-A692-6A745889F93F)
Diagnosis - male
Head (Fig. 11a), frons, first antenna as in E. uchidai, E. asanumai n. sp. and E. hatanakai n. sp. Second antenna proximal antennomere subcylindrical, with a proximal anteromedial, conical apophysis. Apophysis apically rounded and directed medially as in E. uchidai and E. asanumai n. sp. but less chitinization. Proximal antennomere with a posterobasal medial cone as in E. uchidai, E. asanumai n. sp. and E. hatanakai n. sp. Proximal antennomere bearing an anterior, proximal, lamellar antennal appendage (Fig. 11g) tapering from seven tenths of the length. Antennal appendage approximately the length of the second antenna and longer than its proximal antennomere. Antennal appendage with posterobasal medial-marginal spines as in E. uchidai, E. asanumai n. sp. and E. hatanakai n. sp. Medial margin proximal seventh smooth, with remaining medial margin bearing short digitiform papillae and short spines sparsely. Lateral margin proximal seventh smooth, with remaining lateral margin bearing long digitiform papillae continuously; papillae gradually increasing in size to distal second, reaching 0.8 times the length of the horizontal width of the adjacent appendage at the medial, then decreasing in size to apex. All digitiform papillae ornamented with spines as in E. uchidai, E. asanumai n. sp. and E. hatanakai n. sp. Second antenna proximal-distal joint as in E. uchidai, E. asanumai n. sp. and E. hatanakai n. sp. Second antenna distal antennomere (Fig. 11f) chitinized, curved, tapering and with concavity, as in E. uchidai and E. asanumai n. sp. Distal antennomere with an anterobasal projection and continuing with spines, as in E. asanumai n. sp. and E. hatanakai n. sp. Distal antennomere medial margin with an obtuse projection apically. Thorax, thoracopods, gonopod (Fig. 11d), abdomen, telson and cercopods as in E. uchidai, E. asanumai n. sp. and E. hatanakai n. sp.
Diagnosis - female
Head (Fig. 11b), first antenna, second antenna as in E. uchidai, E. asanumai n. sp. and E. hatanakai n. sp. Thorax smooth, but last three thoracic segments bearing a pair of dorsolateral lobes on each segment (Fig. 10c and e). Ninth lobe rounded, smaller than tenth and eleventh lobes. Tenth and eleventh linear lobes as in E. uchidai, projected posteriorly slightly. Thoracopods as in male. Brood pouch (Fig. 10e and h) subcylindrical with tapered apex reaching to the third abdominal segment. Apex one tenth the width of the abdomen. Abdomen (Fig. 11e) cylindrical, and with a pair of posterolateral spines on each segment from the last genital segment to the second abdominal segment. Telson and cercopods as in male.
Egg
Spherical, diameter approximately 400 μm, shallow engraving and densely covered with small scale-like projections 10–20 μm wide as in E. asanumai n. sp. (Fig. 8d).
Authorship
N. Takahashi and H. Moriya described E. khankanus n. sp.
Etymology
E. khankanus n. sp. is named after Lake Khanka in Primorsky, Russia, the type locality of this new species.
Type materials
Holotype: 1 male (18 mm), Russia: Primorsky: Spassky: South bank of Lake Khanka, 29 April 2007, N: 44°36′19.2″, E: 132°42′02.2″, Y. Nagahata, deposited Hokkaido University Museum type no. ICHUM 5502. Paratypes: 1 female (16 mm), same collection data as holotype, deposited Hokkaido University Museum type no. ICHUM 5503; 2 males (16, 17 mm) and 1 female (20 mm), same collection data as holotype, deposited Lake Biwa Museum type nos. 1,430,008,290–8291.
Accession numbers in the DDBJ/EMBL/GenBank International Nucleotide Sequence Database are LC314406 for COI, and LC314407 for 16S rRNA, taken from an individual obtained from the same locality as the holotype. Here these sequences are considered as topogenetypes (proposed by [50]) of E. khankanus n. sp.
Type locality
Small puddle in completely reclaimed hilly area with open forest of Salix spp. (Fig. 5a). This snowmelt puddle was shallow and contained some aquatic Hemipterans, Coleopterans and Lepidurus sp.
Distribution and habitat
E. khankanus n. sp. is known only from the type locality. Marshes are spreading around this area. The species has not been observed co-occurring with other fairy shrimps.
Nucleotide sequences and phylogenetic trees
We obtained a 488 bp sequence for COI. One haplotype each was obtained from E. hatanakai n. sp., E. asanumai n. sp. and E. khankanus n. sp. Two haplotypes were obtained from E. uchidai, which we designated “haplotype A” and “haplotype B”. There were 12 nucleotide differences (ten transitions and two transversions) in the 488 bp between the two haplotypes, all of which were synonymous. Nucleotide difference between the two types was 2.5% (≈ 12/488 × 100). We could not, however, find any morphological differences between the two haplotypes.
A phylogenetic tree (Fig. 12) for Anostraca species was constructed using the COI sequences (Additional file 1: Table S1). Some studies [52,53,54] have found that the Chirocephalidae and Thamnocephalidae are more closely related to each other than they are to the Artemiidae, so we used the Artemia species as outgroups. In the Chirocephalidae cluster, Chirocephalus and Eubranchipus species from the North America were indicated as sister genera. Surprisingly, the Eubranchipus species from northern Japan and the Russian Far East formed a distinct cluster separate from the other Eubranchipus species (Fig. 12). E. hatanakai n. sp., E. asanumai n. sp. and E. khankanus n. sp. are unambiguously distinct from E. uchidai.

Nucleotide differences among the four species (excluding the comparison between A and B haplotypes of E. uchidai) were 9.4–15.6% (Additional file 2: Table S2). These values are comparable to values among Chirocephalus species (7.2–19.7%) and among Artemia species (6.4–20.1%). In addition, all nucleotide substitutions among the four species were synonymous, indicating that the COI sequences determined in this study are subject to functional constraints and do not constitute pseudogenes such as nuclear mitochondrial (numt) DNA segments.
We also obtained a 489 bp sequence for 16S rRNA. As above, one haplotype was obtained each from E. hatanakai n. sp., E. asanumai n. sp. and E. khankanus n. sp., and comparison of the two haplotypes from E. uchidai showed seven nucleotide differences (six transitions and one transversion).
A phylogenetic tree (Fig. 13) was constructed using these sequences (Additional file 3: Table S3). In this case, the collected Eubranchipus species from northern Japan and the Russian Far East formed a cluster with the North American and European Eubranchipus species, but were nevertheless distinct from them, as in the COI tree.

Phylogenetic tree of fairy shrimp species based on 16S rRNA nucleotide sequence data. The tree was constructed using the maximum likelihood method [41] and the GTR + G model [43]. Scale bar: 0.05 nucleotide substitutions per site. Bootstrap values (percentages of 500 replicates) are shown at the nodes
Nucleotide differences among the four species (excluding the comparison between A and B haplotypes of E. uchidai) were 2.6–4.3% (Additional file 4: Table S4). These values are somewhat low but comparable to values among Chirocephalus species (6.4–9.2%) and among Artemia species (3.8–10.0%).
Phylogenetic trees reconstructed from both COI and 16S data showed lower bootstrap values for E. uchidai, E. khankanus n. sp., E. hatanakai n. sp. and E. asanumai n. sp., excluding 98% of A and B haplotypes of E. uchidai in the COI tree. When we reconstructed phylogenetic networks with E. uchidai, E. khankanus n. sp., E. hatanakai n. sp. and E. asanumai n. sp. using COI and 16S data, large reticulations were observed in both phylogenetic networks (Additional file 5: Figure S1). These reticulations indicate incompatible phylogenetic information, as may be caused by parallel and/or backward substitutions. In general, if there is no incompatible phylogenetic information in the data, a phylogenetic network automatically becomes an unrooted phylogenetic tree. Thus, those parallel and/or backward substitutions result in lower bootstrap values.
We reconstructed a phylogenetic tree based on the concatenated COI and 16S data (Fig. 14). Maximum likelihood and Bayesian methods both yielded identical topologies. The phylogenetic tree is also showed that E. hatanakai n. sp., E. asanumai n. sp. and E. khankanus n. sp. are unambiguously distinct from E. uchidai, and the Eubranchipus species from northern Japan and the Russian Far East formed a distinct cluster separate from the Eubranchipus species from North America.

Phylogenetic tree of fairy shrimp species based on COI and 16S rRNA nucleotide sequence data. The tree was constructed using the maximum likelihood method [41] and the GTR + G model [43]. Scale bar: 0.1 nucleotide substitutions per site. Bootstrap (percentages of 500 replicates)/Bayesian credibility values are shown at the nodes
Discussion
Comparison with Holarctic congeners
Far eastern species are readily separated from North American eleven species based on obvious morphological characteristics. Males are homologous in terms of a two-segmented second antenna bearing a lamellar antennal appendage. But the distal antennomere develops a ramus and lacks a complicated concavity, and the length of papillae on the antennal appendage does not exceed the width of the appendage. There are no North American species with a rostrum on the frons (note that the rostrum is not a labral appendage like as in Eubranchipus bundyi Forbes, 1876), and females do not have abdominal spines. Brood pouches are similar, basically subcylindrical with a short tapering apex.
The European species Eubranchipus grubii (Dybowski, 1860) shares more homological features with Far Eastern species. The male distal antennomere lacks a ramus and has a curved concavity. The frons (between the second antennae) possesses a distinct rostrum half the length of the proximal antennomere. The antennal appendage bears long papillae. The female brood pouch is similar but thoracic lobe is lateral.
To this extent, a morphological comparison of males agrees with the mitochondrial 16S sequence results in establishing a difference of Far Eastern species versus North American and European species. However, considering the morphological differences between European and North American species, the low divergence of these species is suggestive and may indicate mismatched morphological and molecular groupings. In females, the presence of abdominal spines corresponds with the separation from Far Eastern species suggested by the molecular grouping.
The inland Eurasian species Eubranchipus birostratus (Fisher, 1851), Eubranchipus hankoi (Dudich, 1927) and Eubranchipus vladimiri (Vekhoff and Vekhova, 1992) are similar in many respects. Males’ distal antennomeres are sickle-like, have complicated concavities, and feature a large, spiny anterobasal projection. The frons projects a small rostrum which is however bifurcated. Remarkably, the antennal appendage has no digitiform papillae. The female brood pouch and thoracic dorsolateral lobe are similar, but they possess a broad abdomen without spines.
The arctic Eurasian species E. claviger and E. rostratus are most similar to the Far Eastern species examined in the present study. Males’ proximal antennomeres bear posterobasal medial cones. The distal antennomere has a small, spiny anterobasal projection and complicated concavity. The antennal appendage develops digitiform papillae. The frons possesses a small, simply-shape rostrum. Females are similar to those of inland Eurasian species but possess a cylindrical abdomen with spines.
Systematic placement of males and identifiable females in far eastern species
The differential morphologies of the four Eubranchipus species are summarized in Table 1 and Fig. 15. Anostracan males have second antenna that are modified into large claspers to amplex the female during copulation, which have been used as an identifying characteristic at the species level [11, 12]. The male of E. hatanakai n. sp. is readily identified by its longer distal, hookless antennomere and a tubular apophysis on the proximal antennomere. The other two new species and E. uchidai males are morphologically more similar with a distal antennomere hook and conical apophyses, but can still be distinguished from each other. This morphological relationship corresponds with mitochondrial COI and COI + 16S results regarding earlier divergence of E. hatanakai n. sp. and the closely related status of E. uchidai with E. asanumai n. sp. and E. khankanus n. sp. Of the species analyzed in the present study, the female E. asanumai n. sp. is readly identified by its subspherical brood pouch and the largest tenth thoracic dorsolateral lobes. However, differences were also discernible among the other three analyzed females, as in the males of those species. These included the shape of the brood pouch, the shape of thoracic dorsolateral lobes, and different numbers of posterolateral spines on abdomen. However, these combinations of female characteristics allow less systematic clarification than in males, and correspondence with the molecular analyses carried out in the present study is not readily established.

Combination of the key morphologies (male second antenna apexes) and phylogenic tree shown in Fig. 14
Female Anostracans generally offer few identifying characteristics useful for identification at the species level, and most species have primarily been described on the basis of male characteristics. Fewer females are found in proposed identification keys [6, 12, 49, 55]. Species separation based on females is possible for a part of the genus Eubranchipus based on the morphology of the amplexial groove, as shown by Rogers [11]. Following this author, the anostracan amplexus can be classified into three types. Species of the genus Eubranchipus possess a “lock and key” morphology, the lock being the female amplexial groove (the region on the female thorax between the base of the brood pouch and the last pair of legs) and the key being the male second antenna. Rogers [11] found that all female Eubranchipus in his collection possessed a well-developed amplexial groove and could be readily separated at the species level based on its morphology.
Conclusion
We reviewed one endemic species and described three new species of Anostracan based on both male and female characteristics as well as molecular data. Six decades after H. Kikuchi first described E. uchidai, morphological variations among females of Far Eastern species of the genus Eubranchipus were shown to complement his findings.
Abbreviations
- 16S rRNA:
-
16S ribosomal RNA
- COI:
-
cytochrome oxidase subunit 1
- DDBJ:
-
DNA Data Bank of Japan
- EMBL:
-
European Molecular Biology Laboratory
- GTR:
-
General time reversible
- ICHUM:
-
Invertebrate Collection of the Hokkaido University Museum
References
Eriksen CH, Belk D. Fairy shrimps of California's puddles, pools, and playas. California: Mad River Press. 1999. p. 196.
Rogers DC. Branchiopoda (Anostraca, Notostraca, Laevicaudata, Spinicaudata, Cyclestherida). In: Likens GF, editor. Encyclopedia of inland waters. 2. Oxford: Academic Press; 2009. p. 242–9.
Rogers DC. A conceptual model for Anostracan biogeography. J Crustac Biol. 2015;35(5):686–99.
Belk D. Uncovering the Laurasian roots of Eubranchipus. Hydrobiologia. 1995;298:241–3.
Rogers DC. The development of the male second antenna in Polyartemiella hazeni (Murdoch, 1884) with a morphological definition of the Chirocephalidae (Crustacea: Anostraca). Zootaxa. 2003;251(1):12.
Rogers DC. Two new cryptic Anostracan (Branchiopoda: Streptocephalidae, Chirocephalidae) species. J Crustac Biol. 2014;34(6):862–74.
Kikuchi H. Occurrence of a new fairy shrimp, Chirocephalopsis uchidai sp. nov., from Hokkaido, Japan (Chirocephalidae Anostraca). J Fac Sci Hokkaido Univ Ser 6 Zool. 1957;13:59–62.
Moriya H. Notes on a fairy shrimp, Eubranchipus uchidai (Kikuchi) (Anostraca), from Japan. Hydrobiologia. 1985;120:97–101.
Ooyagi A. First records of a fairy shrimp, Eubranchipus uchidai Kikuchi (Anostraca), in Honshu, Japan, with some notes on its ecology. J Nat Hist Aomori. 1996;1:25–9. (in Japanese)
Brendonck L, Belk D. On potentials and relevance of the use of copulatory structures in Anostracan taxonomy. Hydrobiologia. 1997;359:83–92.
Rogers DC. The amplexial morphology of selected Anostraca. Hydrobiologia. 2002;486:1–18.
Rogers DC. Female-based characters for anostracan (Crustacea: Branchiopoda) identification: a key for species of California and Oregon, USA. Hydrobiologia. 2002;486:125–32.
Sanoamuang LO, Murugan G, Weekers PH, Dumont HJ. Streptocephalus sirindhornae, new species of freshwater fairy shrimp (Anostraca) from Thailand. J Crustac Biol. 2000;20(3):559–65.
Ketmaier V, Zarattini P, De Matthaeis E, Cobolli M, Mura G. Intra- and inter-specific relationships in the six Italian species of the fairy shrimp genus Chirocephalus: combining allozyme and mtDNA data. J Zool Syst Evol Res. 2003;41(4):276–85.
Stenderup JT, Olesen J, Glenner H. Molecular phylogeny of the Branchiopoda (Crustacea)–multiple approaches suggest a ‘diplostracan’ancestry of the Notostraca. Mol Phylogenet Evol. 2006;41(1):182–194.
MuNOz J, GOMez A, Green AJ, Figuerola J, Amat F, Rico C. Phylogeography and local endemism of the native Mediterranean brine shrimp Artemia salina (Branchiopoda: Anostraca). Mol Ecol. 2008;17(13):3160–77.
Tizol-Correa R, Maeda-Martínez AM, Weekers PH, Torrentera L, Murugan G. Biodiversity of the brine shrimp Artemia from tropical salterns in southern México and Cuba. Curr Sci. 2009:81–7.
McCafferty SS, Warren N, Wilbur C, Shumway S. Genetic divergence among Massachusetts populations of the vernal pool fairy shrimp Eubranchipus vernalis (Crustacea: Anostraca). Northeast Nat. 2010;17(2):285–304.
Jeffery NW, Elías-Gutiérrez M, Adamowicz SJ. Species diversity and phylogeographical affinities of the Branchiopoda (Crustacea) of Churchill, Manitoba, Canada. PLoS One. 2011;6(5):e18364.
Ketmaier V, Marrone F, Alfonso G, Paulus K, Wiemann A, Tiedemann R, Mura G. Mitochondrial DNA regionalism and historical demography in the extant populations of Chirocephalus kerkyrensis (Branchiopoda: Anostraca). PLoS One. 2012;7(2):e30082.
Reniers J, Vanschoenwinkel B, Rabet N, Brendonck L. Mitochondrial gene trees support persistence of cold tolerant fairy shrimp throughout the Pleistocene glaciations in both southern and more northerly refugia. Hydrobiologia. 2013;714(1):155–67.
Zhang H, Luo Q, Sun J, Liu F, Wu G, Yu J, Wang W. Mitochondrial genome sequences of Artemia tibetiana and Artemia urmiana: assessing molecular changes for high plateau adaptation. Sci China Life Sci. 2013;56(5):440.
Eimanifar A, Van Stappen G, Marden B, Wink M. Artemia biodiversity in Asia with the focus on the phylogeography of the introduced American species Artemia franciscana Kellogg, 1906. Mol Phylogenet Evol. 2014;79:392–403.
Eimanifar A, Van Stappen G, Wink M. Geographical distribution and evolutionary divergence times of Asian populations of the brine shrimp Artemia (Crustacea, Anostraca). Zool J Linnean Soc. 2015;174:447–58.
Linder F. Contributions to the morphology and taxonomy of the Branchiopoda Anostraca. Zoologiska Bidrag från Uppsala. 1941;20:101–303.
Brtek J. Einige Notizen zur Taxonomie der Familie Chirocephalidae Daday 1910. Annotnes Zool Bot (Bratislava). 1966;33:1–65. (in German)
Belk D, Brtek J. Checklist of the Anostraca. Hydrobiologia. 1995;298:315–53.
Rogers DC. Anostraca catalogus (Crustacea: Branchiopoda). Raffles Bull Zool. 2013;61(2):525–46.
Fischer S. Branchiopoden und Entomostraceen. In: v. Middendorff A Th, editor. Reise in den Äussersten Norden und Osten Sibiriens 2 [Zoologie](1) [Wirbellose Thiere: Annulaten, Echinodermen, Insecten, Krebse, Mollusken, Parasiten]. St. Petersburg: 1851;149–62. (in German)
Dybowski B. Beitrag zur Phyllopoden-Fauna der Umgegend Berlins, nebst kurzen Bemerkungen über Cancer paludosus O. F M Archiv fur Naturgeschichte. 1860;26:195–204. (in German)
Sars GO. The Phyllopoda of the Jana-expedition. Ann Mus Zool Acad Sci St Petersbourg. 1897;2:461–93.
Daday de Dees E. Monographie Systématique des Phyllopodes Anostracés. Ann Sc Nat Zool Sér 9 1910;11:91–489. (in French)
Dudich E. Neue Krebstiere in der Fauna Ungarns. Arch Balatonicum. 1927;1:344–8. (in German)
Belk D. Key to the Anostracan (fairy shrimps) of North America. Southwest Nat. 1975;20:91–103.
Vekhoff NV, Vekhova TP. A new species of Anostraca (Crustacea) from the water body in Central Russia. Vestnik Zool. 1992;2:63–6. (in Russian)
Brtek J, Mura G. Revised key to the families and genera of the Anostraca with notes on their geographical distribution. Crustaceana. 2000;73(9):1037–88.
Rogers DC, Jensen JB, Floyd TM. Eubranchipus stegosus n. sp. (Crustacea: Anostraca), a bizarre new species of fairy shrimp from the southeastern United States. Hydrobiologia. 2004;528:201–7.
Richter S, Olesen J, Wheeler WC. Phylogeny of Branchiopoda (Crustacea) based on a combined analysis of morphological data and six molecular loci. Cladistics. 2007;23:301–36.
Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32:1792–7.
Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–4.
Felsenstein J. Evolutionary trees from DNA sequences: a maximum likelihood approach. J Mol Evol. 1981;17:368–76.
Stamatakis A. RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics. 2014;30:1312–3.
Tavaré S. Some probabilistic and statistical problems in the analysis of DNA sequences. Lect Math Life Sci. 1986;17:57–86.
Felsenstein J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution. 1985;39:783–91.
Yang Z, Rannala, B. Bayesian phylogenetic inference using DNA sequences: a Markov chain Monte Carlo method. Mol Biol Evol. 1997;14:717–724.
Ronquist F, Huelsenbeck JP. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics. 2003;19:1572–4.
Bryant D, Moulton V. Neighbor-net: an agglomerative method for the construction of phylogenetic networks. Mol Biol Evol. 2004;21:255–65.
Huson DH, Bryant D. Application of phylogenetic networks in evolutionary studies. Mol Biol Evol. 2006;23:254–267.
Timms BV. A revised identification guide to the fairy shrimps (Crustacea: Anostraca: Anostracina) of Australia. Museum Victoria Science Reports. 2015;19:1–44.
Chakrabarty PG. A concept to help integrate molecular phylogenetics and taxonomy. Zootaxa. 2010;2632:67–8.
Goto Y, Gouchi N, Matsuda I. Radiocarbon dating of the Minamidake debris-avalanche deposit, Shiretoko-iozan volcano, eastern Hokkaido. Japan Bull Volcanol Soc Japan. 2011;56(4–5):161–7.
Remigio EA, Hebert PD. Affinities among anostracan (Crustacea: Branchiopoda) families inferred from phylogenetic analyses of multiple gene sequences. Mol Phylogenet Evol. 2000;17:117–128.
Weekers PH, Murugan G, Vanfleteren JR, Belk D, Dumont HJ. Phylogenetic analysis of anostracans (Branchiopoda: Anostraca) inferred from nuclear 18S ribosomal DNA (18S rDNA) sequences. Mol Phylogenet Evol. 2002;25:535–544.
deWaard JR, Sacherova V, Cristescu ME, Remigio EA, Crease TJ, Hebert PD. Probing the relationships of the branchiopod crustaceans. Mol Phylogenet Evol. 2006;39:491–502.
Timms BV. An identification guide to the brine shrimps (Crustacea: Anostraca: Artemiina) of Australia. Museum Victoria Science Reports. 2012;16:1–36.
Acknowledgments
We are grateful for the aid of Seiki Igarashi of the Hokkaido Research Organization and of members of the Rausu Municipal Museum. We thank two anonymous reviewers and the editor for their valuable comments.
Availability of data and materials
Holotype and paratypes Eubranchipus asanumai n. sp. were deposited in Hokkaido University Museum, Sapporo, Hokkaido, Japan (type nos. ICHUM 5494–5499), and additional paratypes were deposited in Lake Biwa Museum, Kusatsu, Shiga, Japan (type nos. 1,430,008,288–8289). Holotype and paratype Eubranchipus hatanakai n. sp. were deposited in Hokkaido University Museum (type nos. ICHUM 5500, 5501). Holotype and paratype Eubranchipus khankanus n. sp. were deposited in Hokkaido University Museum (type nos. ICHUM 5502, 5503), and additional paratypes were deposited in Lake Biwa Museum (type nos. 1,430,008,290–8291). All DNA sequences were deposited into the DDBJ/EMBL/GenBank International Nucleotide Sequence Database (Additional file 1: Table S1 and Additional file 3: Table S3).
Author information
Authors and Affiliations
Contributions
NT, TK and KU conceived and designed the study. NT, MH, HM, KI, YH, YN, YAT and KU collected fairy shrimp specimens. NT, MH, HM, YN and KU performed the experiments. NT, TK, MH, HM, YN and KU analyzed the data. NT, TK, MH and KU wrote the paper. NT created the drawings and microscopic images. NT, TK, MH, YH, YN and KU created the figures. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
N. Takahashi researched and collected the fairy shrimps in special protection zone of Shiretoko national park, with the permission nos. 1,604,271 and 1,704,061 of the Kushiro environmental office, Ministry of the environment. Y. Nagahata and Y. A. Tshistjakov obtained approval for collection of specimens from the Russian Science Academy. No approvals were required from specimens of other locations in this study. Ethical approvals are not required at Yamagata University for research conducted on invertebrates such as shrimps used in the study.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
Table S1. COI nucleotide sequence data used in the study. (XLS 33 kb)
Additional file 2:
Table S2. Pair-wise nucleotide difference per site in % for COI data. (XLS 36 kb)
Additional file 3:
Table S3. 16S rRNA nucleotide sequence data used in the study. (XLS 31 kb)
Additional file 4:
Table S4. Pair-wise nucleotide difference per site in % for 16S rRNA data. (XLS 35 kb)
Additional file 5:
Figure S1. Phylogenetic networks of Far East Eubranchipus species based on COI and 16S data. (PDF 7 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Takahashi, N., Kitano, T., Hatanaka, Y. et al. Three new species of the fairy shrimp Eubranchipus Verill, 1870 (Branchiopoda: Anostraca) from northern Japan and far Eastern Russia. BMC Zool 3, 5 (2018). https://doi.org/10.1186/s40850-018-0029-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40850-018-0029-2