
Abstract
Background
Bioactivities of plant-based compounds had been shown to be influenced by properties of the extracting solvents. This study investigated the influence of ethanolic extract of Xylopia aethiopica (Dunal) A. Rich on the haematological and biochemical parameters in apparently healthy Wistar Rats using solvent partitioning.
Methods
The procedures involved partitioning of ethanolic extract of X. aethiopica (Dunal) A. Rich into n-hexane and aqueous fractions, chosen based on their differences in polarities and dielectric constants. The samples were administered at doses of 100 mg/kg body weight and 200 mg/kg body weight orally once per day for a period of fourteen (14) days to apparently healthy male Wistar rats that had been randomly selected into seven (7) groups. Haematological parameters and plasma biochemical indices were determined using standard protocols after the rats were sacrificed on the 15th day.
Results
The results showed that the levels of haematocrit and related parameters in all the groups that received the extract when compared to the control increased significantly (p < 0.05). Additionally, no detrimental effect was observed on the levels of endogenous antioxidants, liver and kidney function indices in the rats compared to the control.
Conclusion
In summary, on the basis of these findings, this study suggests that the extracts may be capable to alleviate the pathologies that are associated with haematological disorders through improvement of endogenous antioxidant status. The study proposes further evaluation on the molecular mechanism of bioactive components of X. aethiopica (Dunal) A. Rich as agents that can prevents anaemia.
Similar content being viewed by others
Background
Oxidative stress had been established as one of the principal causative factors of haematological disorders which include destructions of blood cells leading to secondary complications such as anaemia, leukaemia and thrombocytopenia [1, 2]. Moreover, impairment in the functions of endogenous antioxidant systems are known to contribute to the elevation of free radicals and reactive oxygen species leading to oxidative stress. Major defence mechanisms against oxidative stress include enzymatic (catalase (CAT), glutathione peoroxidase (GPx), superoxide dismutase (SOD)) and non-enzymatic (vitamins C and E, phenolic compounds, reduced glutathione) systems. Superoxide dismutase 1 (SOD-1) deficiency and peroxiredoxin II (a member of the thioredoxin peroxidase family) have been separately been linked to development of anaemia in mice [3, 4]. However, studies on catalase deficient mice and GPx1- deficient mice had failed to establish the connections between these enzyme deficiencies and haematological parameters [5, 6]. Moreover, other studies have suggested that intake of antioxidants compounds might be a promising approach for the control of infection-mediated anaemia [7].
Plant based natural products and food containing antioxidant nutrients have been established to lower the incidence of oxidative stress-related pathologies [8]. These plant materials have been effective sources of traditional medicines against many ailments since ancient times. These medicinal plants contain various biologically active compounds which are believed to play essential roles in the amelioration of diseases that are associated with oxidative stress by reducing the concentrations free radicals in the system. Besides, many of these botanicals have been shown to possess haematopoietic ability partly due to their mineral compositions [9]. The fruit of Xylopia aethiopica (Dunal) A. Rich is a commonly used botanical in the treatment of inflammatory disorders and also as postpartum tonic due to its blood-boosting properties [10, 11]. The activities of these plant-based phytochemicals are greatly influenced by the physical and chemical properties of the extracting solvents as solute of similar polarity to the extracting solvent will suitably be dissolved in the solvent [12, 13]. The present study was therefore aimed at investigating the influence of extract partitioning based on polarities on haematological and biochemical parameters in healthy Wistar rats.
Methods
Plant material
The fruits of were gathered from the North central zone of Nigeria and air-dried at ambient temperature of 30 ± 1 °C. The dried sample was pulverised and extracted with ethanol. The ethanolic extract was allowed to dry and one-third of the ethanolic extract was further partitioned with n-hexane and aqueous fractions and kept in a freezer (-20 °C).
Animals
The rats were randomised into seven groups of six animals each, after acclimatisation in the experimental animal holding facility for 7 days. The rats administered with 5% DMSO v/v of distilled water were labelled as control in group 1. Rats in groups 2 to 7 consisted of two dosage groups (100 and 200 mg/kg body weight) administered the extracts (n-hexane partitioned fraction (HPE), aqueous partitioned fraction (APE), ethanolic crude extract (ECE),). The extracts were administered orally for 14 days. Fasted animals in all the groups were sacrificed 16 h after the last administration under diethylether anaesthesia. Blood samples were collected from each rat by cardiac puncture and dispensed into tubes containing ethylenediaminetetraacetic acid (EDTA) and heparin for haematological analysis and biochemical analysis, respectively. The blood samples in the tubes containing heparin were centrifuged at 650×g for 5 min to obtain the plasma for the subsequent biochemical analysis. The liver and the spleen were excised, washed twice with ice-cold sucrose solution (0.25 M) to remove residual blood. The liver and the spleen from each rat were homogenised separately at a tissue concentration of 0.25 g/ml in 0.25 M of ice-cold sucrose solution and centrifuged at 650×g for 5 min. The supernatant of the homogenates was separated and frozen thereafter for subsequent biochemical analyses.
Haematological analysis
The haematological parameters including packed cell volume (PCV), red blood cell count (RBC), Haemoglobin concentration (Hb), mean cell volume (MCV), mean cell haemoglobin (MCH), Mean Cell Haemoglobin Concentration (MCHC), white blood cell (WBC), neutrophil (NEU), lymphocyte (LYM) and platelets (PLT) were determined using automated haematology (MAXCOM Veterinary Analyzer, MC-6200VET).
Endogenous antioxidant status
Peroxidase (POD) activity
The method described by Reddy et al. [14] was adopted for assaying the activity of peroxidase. Peroxidase converts H2O2 to H2O and O2 in the presence of a hydrogen donor, pyrogallol. The oxidation of pyrogallol to a coloured product (purpurgallin) can be followed spectrophotometrically at 430 nm. Exactly 0.1 ml of the sample was mixed with 2.9 ml of prepared working reagent (2.1 ml of distilled water, 0.32 ml of 100 mM potassium phosphate buffer (pH 6.0), 0.16 ml of 0.5% H2O2 and 0.16 ml of 5% pyrogallol). The mixture was read at 420 nm at a rate of 1 reading per minute for 3 min.
where: ΔA = change in absorbance, TV = total volume, SV = sample volume, ɛ = 12 (extinction coefficient of 1 mg/ml of purpurgullin at 430 nm)
Catalase (CAT) activity
Catalase activity was assayed according to the procedure reported by Sinha [15]. An unstable intermediate (perchromic acid) formed when chromic acetate, a reduction product of dichromate in acetic acid, is heated in the presence of H2O2 is monitored spectrophotometrically. Exactly 0.5 ml of sample was added to solution containing 0.01 M phosphate buffer (pH 7.0), 0.5 ml of 0.2 M H2O2 and 0.4 ml distilled water. The reaction is terminated by the addition of 2 ml of dichromoacetic acid mixture (5% potassium dichromate with glacial acetic acid (1:3 by volume)) and heated at 60 °C for 10 min. The absorbance was read at regular interval of 1 min at 620 nm against reagent blank.
The catalase activity will be obtained from the expression:
Where ΔA = change in absorbance, TV = total volume, SV = sample volume, ɛ = molar extinction = 40 M− 1 cm− 1
Superoxide dismutase (SOD) activity
The superoxide dismutase activity was determined by its ability to inhibit the auto-oxidation of epinephrine determined by the increase in absorbance at 480 nm as described by Sun and Zigma [16]. The reaction mixture (3 ml) contained 2.95 ml 0.05 M sodium carbonate buffer pH 10.2, 0.02 ml of liver homogenate and 0.03 ml of 2 mM epinephrine in 0.005 N HCl will be that will be used to initiate the reaction. The cuvette contained 2.95 ml buffer, 0.003 ml of substrate (epinephrine) and 0.02 ml of water. The absorbance was read at interval of 1 min for 3 min at 480 nm.
Where ΔA = change in absorbance, TV = total volume, SV = sample volume, ɛ = molar extinction = 4020 M− 1 cm− 1
Gamma-glutamyl transpeptidase (γ-GT) activity
γ –GT activity was estimated by the method of Szasz [17], where γ-GT enzyme reacts with L-γ - glutamyl-3-carboxy-p-nitroanilide (GLUPA) and glycyl-glycine to give L-γ - glutamyl-glycyl-glycine and 5-amino-2-nitrobenzoate. The sample (50 μl) was mixed with 0.5 ml of working reagent (126 mM glycyl-glycine and 3.3 mM GLUPA constituted in tris buffer (pH 8.0)) and the decrease in absorbance was read at 405 nm after 1 min at 1 min intervals for 3 min.
Calculation: γ-GT(U/l) = ΔA/ min × 1190
Reduced glutathione (GSH)
The reduced glutathione content of the tissues and plasma as non-protein sulphyhydryls was estimated according to the method described by Sedlak and Lindsay [18]. Briefly 0.1 ml of 10% TCA was added to 1.0 ml of the sample to deproteinised the sample and centrifuged at 650×g for 5 min. Exactly 0.5 ml of supernatant was treated with 0.5 ml of Ellmans reagent (19.8 mg of 5.51-dithiobis (2-nitrobenzoic acid) (DTNB) in 100 ml 0f 0.1% sodium nitrate) and 3.0 ml of phosphate buffer (0.2 M, pH 8.0). The absorbance was read at 412 nm against the reagent blank.
where ΔA = change in absorbance, TV = total volume, SV = sample volume, ɛ = molar extinction =1.34 × 104 M− 1 cm− 1
Total thiol
Total thiol was estimated according to the methods described by Sedlak and Lindsay [18]. Exactly 0.5 ml of the sample was treated with 0.5 ml of Ellmans reagent (19.8 mg of 5.51-dithiobis (2-nitrobenzoic acid) (DTNB) in 100 ml 0f 0.1% sodium nitrate) and 3.0 ml of phosphate buffer (0.2 M, pH 8.0). The absorbance was read at 412 nm against the reagent blank.
where ΔA = change in absorbance, TV = total volume, SV = sample volume, ɛ = molar extinction = 1.34 × 104 M− 1 cm− 1.
Malondialdehyde
The index of lipid peroxidation (malondialdehyde) in the samples was determined according to the method described by Buege and Aust [19]. 1.0 ml of the sample was added to 2 ml of TCA-TBA-HCl reagent (0.37% thiobarbituric acid, 0.24 N HCl and 15% trichloro acetic acid in 1:1:1) and boiled for 15 min at 100 °C. The mixture was allowed to cool and centrifuged at 650×g for 10 min to remove. The absorbance of the supernatant was read against reagent blank at 532 nm.
Where, Abs = absorbance, TV = total volume, SV = sample volume, ɛ = molar extinction
Liver and kidney function makers
Total protein
The protein concentration in the serum and tissue homogenate was determined by the biuret method of Gornall et al. [20]. The principle is based on the formation of coloured complex when protein peptide interacts with cupric ions in an alkaline medium. Sample (or standard solution) (0.01 ml) was mixed with diluted (1:4 v/v distilled water) biuret reagent (100 mM sodium hydroxide, 16 mM sodium-potassium tartrate, 15 mM potassium iodide and 6 mM copper sulphate). The mixture was incubated at 28 °C for 30 min and the absorbance read at 550 nm against the reagent blank.
Aspartate aminotransferase (AST) activity
AST is measured by monitoring the concentration of oxaloacetate hydrazone formed with 2,4-dinitrophenylhydrazine as described by Reitman and Frankel [21]. Plasma (or homogenate) (0.1 ml) was mixed with 0.5 ml of reagent (100 mM L-aspartate and 2.0 mM α-oxoglutatrate in 100 mM phosphate buffer (pH 7.4)) and incubated for 30 min at 37 °C, after which 0.5 ml of 2.0 mM 2,4-dinitrophenylhydrazine was added and allowed to stand at 28 °C for 20 min. Precisely 5.0 ml of sodium hydroxide was added to the mixture and the absorbance was read at 546 nm after 5 min against reagent blank.
Alanine aminotransferase (ALT) activity
ALT was measured by monitoring the concentration of pyruvate hydrazone formed with 2,4-dinitrophenylhydrazine as described by Reitman and Frankel [21]. Exactly 0.1 ml of the plasma (or homogenate) was mixed with 0.5 ml of reacting solution which contained 200 mM L-alanine and 2.0 mM α-oxoglutatrate in 100 mM phosphate buffer (pH 7.4) and incubated for 30 min at 37 °C, after which 0.5 ml of 2 mM 2,4-dinitrophenylhydrazine was added and allowed to stand at 30 °C for 20 min. Accurately 5.0 ml of sodium hydroxide was added to the mixture and the absorbance was read at 546 nm after 5 min against reagent blank.
Alkaline phosphatase (ALP) activity
Determination of ALP was carried out according to the method described by Kind and King [22]. Precisely 0.05 ml of the plasma (or homogenate) was mixed with 3.0 ml of the substrate (10 mM p-nitophenylphosphate) reconstituted in 1 M diethanolamine buffer, pH 9.8 with 0.5 mM MgCl2. The absorbance was read at 1, 2, and 3 min at 405 nm.
Albumin
The principle is based on the binding of plasma albumin to 3,3′,5,5′-tetrabromo-mcresol sulphonephthalein (bromocresol green, BCG) according to Doumas et al. [23]. Precisely 3 μl of the sample (or standard) was mixed with 1 ml of diluted (13:87 v/v distilled water) BCG concentrate (75 mM succinate buffer, pH 4.2, 0.15 mM BCG). The mixture was left to incubate for 5 min at 27 °C and the absorbance of the sample (and of the standard) was measured against the reagent blank.
Globulin
The globulin in the plasma was estimated as the difference between the total protein and the albumin expressed in g/l.
Uric acid
The analysis was carried out as described by Schulz [24]. Uric acid is converted by uricase to allantoin and hydrogen peroxide, which under the catalytic influence of peroxidase oxidises 3,5-dichloro-2-hydroxybenzdenesulfonic acid and 4-aminophenazone to form a red-violet N-(4-antipyryl)-3-chloro-5-sulfonate-p-benzo- quinoneimine. Precisely 20 μl of plasma (or standard) was mixed with 1 ml of the reagent prepared by reconstituting equal volume of enzyme reagent (0.25 mM 4- aminophenazone, 1000 U/l peroxidase and 200 U/l uricase) in 50 mM hepes buffer, pH 7.0that contained 4 mM 3,5-dichloro-2-hydroxybenzdenesulfonic acid and incubated for 5 min at 37 °C. The absorbance was measured against reagent blank at 520 nm within 30 min.
Urea
The principle is based on the formation of green indophenol from the reactions of ammonia generated by hydrolysis of urea with salicylate and hypochlorite. The colour intensity is proportional to the concentration of the urea in the sample Kaplan [25]. Exactly 10 μl of the plasma or standard was mixed with 1 ml of the working reagent (30,000 U/l urease reconstituted in 50 mM phosphate buffer that contained 2 mM EDTA, 400 mM sodium salycilate, 10 mM sodium nitroprusside) and incubated at 37 °C for 5 min after which 1 ml of a reagent solution containing 140 mM sodium hypochlorite and 150 mM sodium hydroxide was added and incubated at 37 °C for another 5 min. The absorbance was measured at 580 nm.
Creatinine
The principle is based on the reaction of picric acid in alkaline solution and creatinine and subsequent formation of coloured complex. The amount of complex formed is directly proportional to the concentration of creatinine in the sample as described by Bartles et al. [26]. Exactly 0.2 ml of plasma (or standard) was mixed with 2.0 ml of working reagent prepared by mixing equal volumes of 35 mM picric acid and 0.32 M sodium hydroxide. The absorbance was taken at 492 nm after 30 s and re-taken exactly 2 min later.
Statistical analysis
All data are reported as mean of six determinations ± standard error, unless otherwise stated, and analysed using one-way analysis of variance (ANOVA) with a Fisher’s Least Significant Difference post hoc test to determine significant differences (p < 0.05) between groups.
Results
Haematological parameters
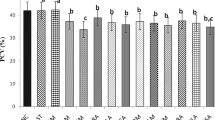
The effects of the extracts on the haematological parameters in healthy animals are presented in Table 1. The result showed increased haematocrit, RBC, and haemoglobin levels in all the groups that received the extracts, with the exception of APE200 mg/kg compared to the control. The MCHC level was significantly lowered in the test groups compared to the control. The level of WBC was significantly higher in the HPE100 mg/kg, HPE200 mg/kg, APE 100 mg/kg and ECE100 mg/kg; the mean percentage neutrophils were increased compared to the control in HPE200mg/kg, APE 200 mg/kg, ECE100mg/kg, and ECE200mg/kg groups while the opposite was true for percentage lymphocytes. The platelet levels were lowered in the groups that received ECE and APE at 200 mg/kg body weight compared to the control.
Endogenous antioxidant status
The effects of administration of the ethanolic extracts of the fruit of X. aethiopica (Dunal) A. Rich on the plasma GSH, lipid peroxidation, γ-GT and total thiol in healthy Wistar rats is shown in Table 2. There was no significant difference (p < 0.05) in the level of plasma GSH (μmol/g protein) across the groups compared to the control. The plasma MDA (μmol/l) level was lowered significantly in all the groups that received the extracts compared to the control. The plasma γ-GT (U/ g protein) level was also lowered only in HPE 100 mg/kg body weight, HPE 200 mg/kg body weight and ECE 100 mg/kg body weight than in the control. However, the total thiol (μmol/g protein) was found to be lowered in HPE 100 mg/kg body weight, HPE 200 mg/kg body weight, ECE 100 mg/kg body weight and APE 200 mg/kg body weight compared to the control group. The effects of the extracts on the antioxidant system in the liver of the rats are presented in Table 3. The liver CAT (U/g of protein) level was significantly higher only in ECE 100 mg/kg body weight and ECE 200 mg/kg body weight, similar trend was observed in the liver POD (U/g of protein) level except with the inclusion of HPE 200 mg/kg. However, HPE 200 mg/kg body weight was the only group that showed higher level of liver γ –GT (U/g of protein). The liver SOD (U/g of protein) was higher in HPE 100 mg/kg body weight and HPE 200 mg/kg body weight while the liver GSH (μmol/g protein) was higher in HPE 100 mg/kg body weight and APE 100 mg/kg body weight groups, compared to the control. The liver MDA (μmol/mg) was significantly (p < 0.05) lowered in the groups that received the n-hexane partitioned extract but elevated in the groups that received the ethanolic and the aqueous partitioned extracts while the liver total thiol (μmol/g of protein) level higher in all except in ECE 100 mg/kg body weight and APE 200 mg/kg body weight. The effect of the test samples on the spleen antioxidant system is shown in Table 4. There was no significant difference in the spleen SOD (U/g of protein), POD (U/g of protein), γ-GT (U/g of protein) levels in all the groups compared to the control. The spleen CAT (U/g of protein) level was higher only in ECE 200 mg/kg body weight; however, all the groups demonstrated higher levels of spleen GSH (μmol/g protein) except HPE200mg/kg body weight and APE 100 mg/kg body weight while only the groups ECE 100 mg/kg body weight, ECE 200 mg/kg body weight showed higher level of spleen MDA (μmol/mg of tissue). The spleen total thiol (μmol/g of protein) was higher in all except in HPE 200 mg/kg body weight and ECE 200 mg/kg body weight.
Kidney and liver function makers
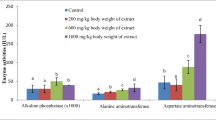
The results presented in Table 5 shows the effect of the oral administration of the ethanolic extracts and the partitioned extracts on kidney function. The plasma levels of urea, uric acid and creatinine can be used to assess the adverse effect of the extracts on the kidney function, and these were lowered in all the groups at p < 0.05 compared to the control. The results of liver function indices (ALT, AST and ALP) are shown in Table 6. It was found that the AST (U/g of protein) was lowered in the plasma of all the animals compared to the control. Similarly, the plasma ALT (U/g of protein) was also reduced in all the groups while the plasma ALP (U/g of protein) level only increased in HPE100 mg/kg body weight and HPE 200 mg/kg body weight, compared to the control. The level of AST in the liver was higher across the group compared to the control group except in ECE 200 mg/kg body weight and APE 200 mg/kg body weight; the liver ALT level increased across the groups except in APE 200 mg/kg body weight. Similarly, the liver ALP level increased across the groups compared to the control, except in ECE 200 mg/kg body weight.
Discussion
Haematological indices
The results of the effect of the extracts administered orally on the haematological indices in the experimental animals showed increased haematocrit, RBC, and haemoglobin levels in all the treated groups, with the exception of APE at 200 mg/kg body weight, compared to the control. The effect was more evident with the oral administration of HPE. These results are in conformity with earlier work done on the different extracts of the fruit of X. aethiopica (Dunal) A. Rich suggesting the effect of the extract on haemopoiesis in the experimental animals and its possibility in the maintenance of cellular membrane of RBCs and reversing anaemia in Wistar rats [10]. The MCHC level was significantly lowered in the groups that received the extracts compared to the control. The level of WBC was significantly higher in the groups that received the HPEs at both dosages, the ethanolic extract and the APE both at 100 mg/kg body weight; the mean percentage neutrophils were increased in the groups that received HPE 200 mg/kg body weight, ECE 100 mg/kg body weight, ECE 200 mg/kg body weight and APE 200 mg/kg body weight as compared to the control while the opposite was true for percentage lymphocytes. These findings are in agreement with a previous work which recognised X. aethiopica (Dunal) A. Rich as a booster of PCV, haemoglobin WBC and neutrophil in rat [27]. These conclusions confirm the conventional use of the extract as a tonic and immune booster. The increased number of the neutrophils may be considered a good response to boost the general resistance of the body against infection [28].. Moreover, the platelet levels were significantly lowered in the groups that received the APE and ECE at 200 mg/kg body weight. This suggests that the ethanolic and the hydrophilic aqueous partitioned extracts could possibly be effective in the management of cardiovascular events that are associated with clots and thrombus formation.
Antioxidant status
In the apparently healthy rats, oral administration of the ethanolic extracts significantly lowered the plasma level of malondialdehyde (MDA) (Tables 2, 3 and 4). However, the ethanolic and the aqueous partitioned extracts significantly increased the hepatic MDA levels compared to the control. This is in conflict with the previous work by Folorunso et al. [29] who reported that oral administration of methanolic extract of X. aethiopica (Dunal) A. Rich at 400 mg/kg body weight could protect against hepatocellular damage. Moreover, the induced lipid peroxidation was not observed in the groups administered the n-hexane fraction, suggesting that the ‘pro-oxidant’ effect might be due to the presence of certain polar phytoconstituents of X. aethiopica (Dunal) A. Rich. Correspondingly, some dietary antioxidants such as vitamins, carotenoids and flavonoids have also been shown to act, sometimes, as pro-oxidants to initiate a mild degree of oxidative stress microenvironment under certain circumstances [30]; which may result in the up regulation of the endogenous antioxidant system in the cell resulting in the overall protection of the cell [31]. Though, the extent to which the phytochemicals can act as anti- and pro-oxidants in vivo is still poorly understood. Moreover, the protective effect against lipid peroxidation observed in the ECE and the HPE groups could be enhanced by the oil (fixed) in the samples. This is in agreement with the conclusion of Patil et al., [32] that the essential oil of Ageratum conyzoides possessed ability to prevent lipid peroxidation. Furthermore, Oladiji et al. [33] revealed that edible oil-based diets are not toxic but, paradoxically, they alleviate the pathogenesis of diseases linked with elevated concentrations of serum total cholesterol in rats. The plasma reduced glutathione (GSH) levels remained statistically unperturbed in the groups that received the extracts compared to the control. The ECE at 100 mg/kg body weight and the HPE at both dosages showed lowered levels of γ-GT in the plasma while the levels in the other groups are registered as not significant when compared to the control. The total thiol levels were significantly lowered except in the groups that received ethanolic extract. The observable reduction in the plasma total thiol levels may be due to low concentration of the extracellular protein (Table 6) and enzymes due to the possible protective ability of the extract in the cells.
Liver and kidney function markers
Damage to the liver is a widely used indicator of toxicity of medicinal plants in vivo. This issue of safety and toxicity has imposed great limitation to the use of medicinal plants and herbs. The aminotransferases (ALT and AST) and ALP are among serum marker enzymes of hepatic function, with their increase in the serum indicating hepatic damage [34]. The administration of the extract/fractions to the apparently healthy rats did not result in any toxicity or adverse effect. This is indicated by the reduction in the levels of the plasma aminotransferases and the phosphatase. The mean plasma protein was significantly higher in all the groups except in the APE 100 mg/kg group. Moreover plasma albumin level was higher in the HPEs and ECE 100 mg/kg. This suggests the apparent ability of the X. aethiopica (Dunal) A. Rich in the induction and stimulation of protein and albumin in the liver. This was also confirmed by the significant increase observed in the hepatic AST, ALT and ALP levels compared to the control. The effects of the extracts on the kidney function were assessed by evaluating the levels of uric acid, urea and creatinine in the plasma of the rats. Uric acid, urea and creatinine are derived from endogenous sources as products of metabolisms and usually cleared by the kidney following glomerulus filtration. They accumulate in the plasma in renal dysfunction; thus, they are biochemical indices in the assessment of nephrotoxicity [35]. There was no indication from the study that the extracts could initiate detrimental effects on the kidney functions as revealed by the plasma levels of creatinine, blood urea nitrogen, and uric acid when compared to the control animals.
Conclusion
The study suggests that the dried fruit of X. aethiopica (Dunal) A. Rich possess erythropoietin promoting activity. Thus, the extract and its fractions used in this study may hold promise in the prevention and/or treatment of anaemia. However, the biological activities vis-à-vis the haematological and biochemical indices observed in the apparently healthy rats were slightly influenced by the solvent partitioning of the ethanolic crude extract. The significant increase in total white blood cells and the differential white blood cell counts in the group treated with the aqueous partition extract indicated that the hydrophilic phytochemicals in the ethanolic extract may promote the immune-stimulatory activities. The significant decreases in platelets following oral administration of the ethanolic extract and aqueous partitioned extract demonstrate that the extracts may possibly be able to prevent thrombus formation and blood clotting, a property that can be harnessed in the events leading to cardiac arrest. Furthermore, X. aethiopica (Dunal) A. Rich stimulated endogenous, and may antioxidant system therefore lessen oxidative stress. This study therefore, scientifically confirms and supports the traditional use of the fruit of X. aethiopica (Dunal) A. Rich in enhancing haematological parameters and improving health. Hence, further studies are encouraged to establish their clinical relevance in the management of haematological disorders.
Abbreviations
- APE:
-
Aqueous partitioned fraction of ethanolic extract of X. aethiopica (Dunal) A. Rich
- ECE:
-
Ethanolic crude extract X. aethiopica (Dunal) A. Rich
- HPE:
-
N-hexane partitioned fraction of ethanolic extract of X. aethiopica (Dunal) A. Rich
References
Bracci R Normal and abnormal effects of free radicals in blood cells. Pediatric Res 1999; 45:771.
Mazzulla S, Schella A, Gabriele D, Baldino N, Sesti S, Perrotta E, Costabile A, de Cindio B. Oxidation of human red blood cells by a free radical initiator: effects on rheological properties. Clin Hemorh Microcirculation. 2015;60:375–88.
Lee TH, Kim SU, Yu SL, Kim SH, Park S, Moon HB, Dho SH, Kwon KS, Kwon HJ. Han YH. Et al. (2003) Peroxiredoxin II is essential for sustaining life span of erythrocytes in mice. Blood. 2003;101:5033–8.
Starzyński RR, Canonne-Hergaux F, Willemetz A, Gralak MA, Woliński J, Styś A, Olszak J, Lipiński P. Haemolytic anaemia and alterations in hepatic iron metabolism in aged mice lacking cu, Zn-superoxide dismutase. Biochem J. 2009;420(3):383–90.
Ho YS, Xiong Y, Ma W, Spector A, Ho DS. Mice lacking catalase develop normally but show differential sensitivity to oxidant tissue injury. J Biol Chem. 2004;279(31):32804–12.
Johnson RM, Goyette G, Ravindranath Y, Ho Y. Red cells from glutathione peroxidase-1-deficient mice have nearly normal defenses against exogenous peroxides. Blood. 2000;96:1985–8.
Iuchi Y. Anemia Caused by Oxidative Stress. In Anemia, Dr. Donald Silverberg (Ed.), 2012, InTech, Available from: http://www.intechopen.com/books/anemia/anemia-caused-by-oxidative-stress.
Stanković N, Mihajilov-Krstev T, Zlatković B, Stankov-Jovanović V, Mitić V, Jović J, Čomić L, Kocić B, Bernstein N. Antibacterial and antioxidant activity of traditional medicinal plants from the Balkan Peninsula. NJAS - Wageningen J Life Sci. 2016;78:21–8.
Ferri-Lagneau KF, Moshal KS, Grimes M, Zahora B, Lv L, Sang S, Leung T. Ginger stimulates hematopoiesis via bmp pathway in zebrafish. PLoS One. 2012;7(6):e39327.
Abaidoo CS, Woode E, Alhassan A. An evaluation of the effect of ethanolic fruit extracts of Xylopia aethiopica on haematological and biochemical parameters in male rats. Der Pharmacia Sinica. 2011;2(2):39–45.
Oso BJ, Oyewo EB, Oladiji AT. Ethanolic, n-hexane and aqueous partitioned extracts of Xylopia aethiopica fruit modulated inflammatory responses in turpentine oil induced acute inflammation in male Wistar rats. Int J Res Health Sci. 2017;5(2):1–10.
Oso BJ, Oyeleke O, Soetan O. Influence of different solvent polarities on the phenolics, flavonoids and antioxidant properties of the fruit of Xylopia aethiopica (Dunal) a. Rich Trends Phytochem Res. 2018;2(2):97–102.
Oso BJ, Boligon AA, Oladiji AT. Metabolomic profiling of ethanolic extracts of the fruit of Xylopia aethiopica (Dunal) a. Rich using gas chromatography and high-performance liquid chromatography techniques. J Pharmacogn Phytochem. 2018;7(1):2083–90.
Reddy KP, Subhani SM, Khan PA, Kumar KB. Effect of light and benzyl adenine and dark-treated gravinig rice (Oryza sativa) leaves- changes in peroxidises activity. Plant Cell Physiol. 1995;26:987–94.
Sinha AK. Colorimetric assay of catalase. Anal Biochem. 1972;47:389–94.
Sun M, Zigma S. An improved spectrophotometer assay of superoxide dismutase based on epinephrine autoxidation. AnalBiochem. 1978;90:81–9.
Szasz G. A kinetic photometric method for serum γ-glutamyl transpeptidase. Clin Chem. 1969;15:124–36.
Sedlak J, Lindsay RH. Estimation of total, protein-bound, and non-protein sulfhydryl groups in tissues with Ellman’s reagent. Anal Biochem. 1958;25(1):192–205.
Buege JA, Aust SD. Microsomal lipid peroxidation. Meth Enzymol. 1978;52:302–10.
Gornall AC, Bardwawill CJ, David MM. Determination of serum protein by means of the biuret reaction. J Biol Chem. 1949;177:751–6.
Reitman S, Frankel S. Colorimetric method for estimation of AST and ALT. Am J Clin Path. 1957;28:56–63.
Kind PR, King EG. Colourimetric determination of alkaline phosphatase. J Clin Pathol. 1954;7:322–6.
Doumas BT, Watson WA, Bigsg HG. Albumin standards and measurement of serum-albumin with bromocresol green. Clin Chim Acta. 1971;31:87–92.
Schulz A. Uric acid. In: Kaplan LA. Clinical Chemistry. The C.V. Mosby Co. St. Louis. Toronto, Princeton; 1984. p. 1261–6.
Kaplan A. Urea nitrogen and urinary ammonia. In: Meites S, editor. Standard method of clinical chemistry. New York: Academic Press Inc.; 1965. p. 245–56.
Bartles H, Bohmer M, Heirli C. Colorimetric kinetic method for creatinine determination in serum and urine. Clin Chem Acta. 1972;37:193.
Taiwo IA, Bola O, Francis-Garuba PN. Haematological properties of aqueous extracts of Phyllantus amarus (Schum and Thonn.) and Xylopia aethiopica (Dunal) a. Rich in albino rats. Ethno-Med. 2009;3(2):99–103.
Oso BJ, Oyewo EB, Oladiji AT. Phytochemical composition, antioxidant capabilities and immunomodulatory in vitro indices of Xylopia aethiopica fruit extracts. Adv Pharm J. 2018;3(1):29–37.
Folorunso OS, Ogungbe BF, Ikanone CEO. Methanolic extract of Xylopia aethiopica ameliorates acetaminophen-induced liver damage in male Wistar rats. Annl Rev Res Biol. 2013;3(4):1074–90.
Halliwell B. Dietary polyphenols: good, bad or indifferent for your health? Cardiovasc Res. 2007;73(2):341–7.
Moini H, Packer L, Saris NE. Antioxidant and prooxidant activities of alpha-lipoic acid and dihydrolipoic acid. Toxicol Appl Pharmacol. 2002;182(1):84–90.
Patil RP, Nimbalkar MS, Jadhav UU, Dawkar VV, Govindwar SP. Antiaflatoxigenic and antioxidant activity of an essential oil from Ageratum conyzoides L. J Sci Food Agr. 2010;90:608–14.
Oladiji AT, Jacob TO, Yakubu MT. Antianaemic potentials of aqueous extract of Sorghum bicolor (L.) Moench stem bark in rats. J Ethnopharmacol. 2007;111:651–6.
Shaarawy SM, Tohamy AA, Elgendy SM, Elmageed ZYA, Bahnasy A, Mohamed MS, Kandil E, Matrougui K. Protective effects of garlic and silymarin on NDEA-induced rat’s hepatotoxicity. Int J Biol Sci. 2009;5(6):549–57.
Ali BH, Ben TH, Basheer AA. Sex related differences in the susceptibility of rat to gentamicin nephrotoxicity: influence of gonadectomy and hormonal replacement therapy. Ind J Pharmacol. 2001;33:369–73.
Ethical approval and consent to participate
All experiments were performed in accordance with ethical laws and with the approval of the University of Ilorin Ethics Committee (UERC) with UERC Approval Number UERC/ASN/2015/047.
Funding
Not applicable
Availability of data and materials
The dataset supporting the conclusions of this article is included as tables in the uploaded additional files.
Author information
Authors and Affiliations
Contributions
BJO conceived, designed and performed the experiments under the supervision of ATO. BJO wrote the manuscript draft with the assistance of EBO. Revision of the manuscript was done by all the authors. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Authors’ information
Dr. B.J. Oso is a lecturer in the Department of Biochemistry, Faculty of Science, Kings University Odeomu, Nigeria. He obtained his B.Sc., M.Sc. and Ph.D in Biochemistry from the University of Ado Ekiti (now, Ekiti State University, Ado Ekiti), Nigeria, University of Ibadan, Nigeria and University of Ilorin, Nigeria, respectively.
Dr. E.B. Oyewo is a lecturer in the Department of Biochemistry, Federal University of Agriculture, Abeokuta, Nigeria. He obtained his B.Sc., M.Sc. and Ph.D. in Biochemistry from the University of Ilorin, Nigeria.
Prof. A.T. Oladiji obtained her B.Sc., M.Sc. and Ph.D. in Biochemistry from the University of Ilorin, Nigeria. She is a Professor of Biochemistry at the Faculty of Life Sciences, University of University of Ilorin., Nigeria.
Consent for publication
Not applicable
Competing interests
Not applicable
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Oso, B.J., Oyewo, E.B. & Oladiji, A.T. Influence of ethanolic extracts of dried fruit of Xylopia aethiopica (Dunal) A. Rich on haematological and biochemical parameters in healthy Wistar rats. Clin Phytosci 5, 9 (2019). https://doi.org/10.1186/s40816-019-0104-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40816-019-0104-4