
Abstract
Background
Previous studies showed that a probiotic-rich diet may improve the plasma lipid profile and lower the risk of cardiovascular disease (CVD). In a randomized controlled study we tested the possibility for regulation of plasma lipid profile using a kefir that contained the antioxidative probiotic strain, Lactobacillus fermentum ME-3 (DSM14241). The trial was performed in clinically healthy adults with borderline-high serum low-density lipoprotein-cholesterol (LDL-C) and/or high serum triglyceride (TG) levels based on guidelines from the European Cardiology Society and European Atherosclerosis Society.
Methods
One hundred sixty four participants meeting the inclusion criteria were included. Participants were randomised to receive 200 ml/day kefir, either with probiotic (PG; n = 71, 58 females, mean age 49.6 ± 6.5 y) or without probiotic (CG; n = 66, 58 females, mean age 49.9 ± 6.1 y). The probiotic contained 4 × 107 cfu/ml L. fermentum ME-3 (total 8 × 109 cfu/day). At 4 weeks (n = 71 PG and n = 66 CG) and at 8 weeks (n = 43 PG and n = 33 CG), we evaluated anthropologic, blood biochemical indices, and the faecal temporal persistence of the probiotic strain was assessed by real-time PCR.
Results
After 4 weeks, the lipid profiles were mostly similar between groups: only the values of oxidised LDL (ox-LDL) and TG were significantly reduced (P < 0.001 and P = 0.005, resp.). After 8 weeks, the PG group exhibited reductions in LDL-C (5 %, P = 0.001), ox-LDL (6 %, P < 0.0002), TG (17 %, P = 0.033). Next, the ratio of LDL-C to HDL-C was decreased only in the PG (P = 0.013) while in the CG it was significantly increased (P = 0.018). After completing the trial the changes in all above listed indices were significantly (P < 0.05) different between the PG and CG groups. In faeces, the prevalence of L. fermentum ME-3 increased after 4 and 8 weeks (both P < 0.001), but the counts, determined with real-time PCR, remained constant.
Conclusions
Eight weeks of consuming kefir with the antioxidative probiotic L. fermentum ME-3, reduced serum LDL-C, ox-LDL and TG values in clinically healthy volunteers with borderline-high lipid profile indices. Thus, L. fermentum ME-3 has potential to lower the risk of CVD that is tightly associated with maintenance of plasma lipid profile. This study was registered as current controlled trial http://www.controlled-trials.com/ISRCTN49744186 as ISRCTN49744186.
Similar content being viewed by others

Background
Cardiovascular disease (CVD) is the number one cause of death worldwide, and its incidence is rapidly increasing in countries with low- and mid-range economies [1, 2]. Abnormal function of the heart is related to dyslipidaemia, identified by the main marker, elevated levels of low-density lipoprotein-cholesterol (LDL-C), and the secondary marker, reduced levels of high-density cholesterol (HDL-C); these cholesterol disorders are summarized by the ratio of LDL: HDL. Numerous clinical trials have demonstrated that LDL-C can independently cause the development and progression of coronary heart disease [3]. In addition, elevated levels of triglycerides (TG) may contribute to dyslipidaemia. In a recent statement, the European Cardiology Society (ESC) and European Atherosclerosis Society (EAS) recommended that LDL-C should be used as a primary screening indicator for CVD risk estimations [4]. Thus, lowering the level of LDL-C remains the primary target of blood lipid-regulating strategies, including therapeutic dietary changes and pharmacological interventions [5].
Gut microbiota is an essential component of the complex host ecosystem involved in nutrition and health. A wide variety of host, microbiological, dietary, and environmental factors affect the metabolic relationships between gut mucosal epithelial cells and microbiota [6–9]. However, it remains unclear what role imbalanced intestinal microbiota might play in the pathophysiology of atherosclerosis, which underlies CVD. Cholesterol is obtained both through the diet and through highly-regulated biosynthesis. Serum cholesterol is predominantly eliminated from body in form of bile acids [10]. Among the normal microbiota of the gastrointestinal tract, some species and strains of lactic acid bacteria can assimilate cholesterol from consumed dietary products [11]. A meta-analysis of 13 probiotic studies indicated that a probiotic-rich diet improved the plasma lipid profile. Probiotic diets reduced the total cholesterol and LDL-C concentrations in the plasma of participants with high, borderline-high, and even normal cholesterol levels [12]. However, it is not clear how long the intervention should last to achieve a beneficial effect in healthy volunteers with borderline-high levels of LDL-C.
Lactobacillus fermentum ME-3 (DSM 14241) is a bacterial strain of human origin that was isolated from a healthy one-year-old child. L. fermentum ME-3 had both antimicrobial and antioxidant properties [13–16]. Various clinical studies have demonstrated several health benefits associated with consuming this strain [17–21].
In the present study, we aimed to evaluate the effects of the probiotic L. fermentum ME-3 on the plasma lipid profile in a randomised, double-blind, placebo-controlled, parallel-design, two-armed study (ISRCTN49744186). We included clinically healthy Estonian adults that had borderline-high levels of LDL-C and above optimum values of serum TG. We evaluated anthropologic, blood, biochemical, and faecal microbiological indices. Significant reductions in serum LDL-C and TG were the primary outcome measures for lowering the CVD risk due to borderline-high plasma lipid profiles.
Methods
Probiotic lactobacillus strain
The L. fermentum ME-3 strain was previously isolated from the gastrointestinal tract of healthy Estonian children [22]. The strain was deposited in the Deutsche Sammlung von Mikroorganismen und Zellkulturen (German Collection of Microorganisms and Cell Cultures GmbH) under the registration number, DSM 14241. The L. fermentum ME-3 strain used in this study was confirmed by molecular identification with polymerase chain reaction that employed an internally-transcribed spacer and 16S rRNA sequencing [23]. The strain’s safety was confirmed previously in an animal model [16] and in healthy Estonian adults (registered trials ISCRNT43435738 and ISCRNT53154826).
Activity for bile hydrolysation
Enzymatic deconjugation of bile acids by the bile salt hydrolase (BSH) activity of L. fermentum ME-3 was tested according to Lim et al. [24]. L. fermentum ME-3 strains were cultured overnight on de Man, Rogosa, Sharpe (MRS; Oxoid, UK) agar in microaerobic conditions at 37 °C. For control cultures, we used three reference strains (L. acidophilus 821–3, L. plantarum 299v, and L. plantarum BAA793). After incubation, cultures were standardised to McFarland 3.0. For testing BSH activity, sterilised paper disks were impregnated with the cultures and placed on MRS agar plates supplemented with 4 % (w/v) tauro-deoxycholic acid sodium salt (TDCA, Sigma, USA) and 0.37 g/l CaCl2. The plates were anaerobically incubated at 37 °C for 72 h, and the diameters of the precipitation zones around the disks were measured. BSH activity was then calculated by subtracting the disc diameter (DD, mm) from the inhibition zone diameter (IZD, mm), and dividing by two, as follows: BSH activity = (IZD-DD)/2. The activity of three consecutive cultures of each strain was expressed as median (CI 95 %) value.
Kefir preparation
The probiotic kefir containing L. fermentum ME-3 (trademark “Hellus”, 2005) was developed at Tere AS in Estonia. Briefly, kefir was prepared from pasteurised (92–95 °C for 5 min) cow’s milk with 2.5 % fat content; L. fermentum ME-3 was added as an adjunct starter. Commercial starter cultures — including kefir grains and mesophilic aromatic cultures of Lactococcus lactis subsp cremoris, Lactococcus lactis subsp lactis biovar diacetylactis, Leuconostoc sp, and Lactococcus lactis subsp lactis (Chr. Hansen, Denmark) — were added to the incubation tank. For production of “Hellus” kefir, freeze-dried L. fermentum ME-3 cultures were added to the kefir milk to a concentration of 1 × 106 colony-forming units (cfu)/ml. The kefir milk fermentation process lasted approximately 20–24 h, until achieving the required pH 4.5. After fermentation, the kefir was cooled to 8-10 °C in a heat exchanger.
The final kefir product possessed a homogenous consistency, creamy colour, shiny smooth surface, fresh and acidic taste, and mild yeast-like aroma. The average nutritional value of 100 g kefir was as follows: energy content 217 kJ/52 kcal, 3.1 g protein, 4.2 g carbohydrate, and 2.5 g fat. The content of L. fermentum ME-3 in the probiotic kefir was 4 × 107 cfu/ml. Similar kefir produced without the probiotic adjunct served as a control.
Subjects
This randomised, double-blind, placebo-controlled, parallel-design, two-armed study (registered trial ISRCTN49744186) was conducted in South Estonia, according to the guidelines laid down in the Declaration of Helsinki 1996–2000. The study was approved by the Ethics Committee of the University of Tartu (protocol number 210/T-3; 18.01.2011). All participants signed the written informed consent at enrolment and were given the option to withdraw from the study at any time.
Subject inclusion criteria were a desire to participate; age 35–65 years; blood lipid fractions: total cholesterol >5.2 mmol/l, LDL-C >3.4 mmol/l, and/or TG >1.7 mmol/l, in addition, subjects had to be willing to maintain a stable diet and physical activity level. Exclusion criteria included a history of gastrointestinal disease; food allergy or acute infection; use of any antimicrobial agent within the preceding 2 months; use of any regular concomitant medication within the preceding 2 weeks, including non-steroidal anti-inflammatory drugs and antioxidant products; pregnancy or breastfeeding; any serious organ or systemic disease; high blood pressure (>140/95 mmHg); eating disorder; extensive exercise; genetic hyperlipidaemia; drug or alcohol abuse; active weight loss >5 kg in the prior 3 months; participation in other studies within the last 30 days or during the study; and not wishing to participate. Also, subjects were excluded when they had used any concomitant treatment within the preceding 2 months that could influence evaluation of the efficacy and tolerability of the investigational study product, including lipid-lowering drugs (e.g., statins, bile acid sequestering agents, cholesterol absorption inhibitors, nicotinic acid) or dietary supplements (e.g., omega-3 fatty acids, calcium, oat fibre, niacin, green tea extract, plant sterols, soy protein, psyllium seed husk, or probiotics/prebiotics).
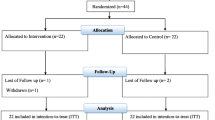
Participants were randomly assigned to receive either probiotic or placebo using a computer-generated randomisation list prepared by the independent statistician. The 164 eligible subjects were randomised to receive either probiotic treatment (PG; n = 83) or control kefir (CG; n = 81; Fig. 1).

Flow chart of the study subjects. Legend: Randomised, double-blind, placebo-controlled, parallel-design, two-armed study (registered trial ISRCTN49744186). Participants were randomly assigned to receive either probiotic or placebo using a computer-generated randomisation list prepared by the independent statistician. The 164 eligible subjects were randomised to receive either probiotic treatment (PG; n = 83) or control kefir (CG; n = 81). The measurements were performed at 4 weeks with 137 participants (n = 71 PG and n = 66 CG) while 27 subjects withdrew. At 8 weeks 76 participants (n = 43 PG and n = 33 CG) were followed; while 51 subjects withdrew
The sample size was calculated for a two-armed intervention investigation that included 4 weeks of probiotic kefir consumption. We assumed a predicted change of 0.2 U in the LDL-C level between the probiotic and control groups during the treatment period. The planned sample size of 83 subjects provided sufficient power (0.90) to detect a significance level of P < 0.05 with the Student’s t-test.
Study protocol
Each group consumed 200 ml/day of kefir without (CG) or with (PG) the probiotic L. fermentum ME-3. The total amount of L. fermentum ME-3 consumed was 8 × 109 cfu/day. The trial lasted for 8 weeks. The weekly food diary was not performed but the appearance of gastrointestinal complaints was registered (Fig. 1). Measurements were carried out at the start of the study (baseline values) with 164 participants that met the inclusion criteria. We conducted measurements at 4 weeks with 137 participants (n = 71 PG and n = 66 CG) while 27 subjects withdrew (n = 12 PG and n = 15 CG) due to respiratory infections, intestinal complaints, and loss of contact. At 8 weeks 76 participants (n = 43 PG and n = 33 CG) were followed; while 51 subjects withdrew due to the infections, loss of contact and motivation (Fig. 1).
Clinical investigations
Subjects in both groups habitually consumed a Western-type diet, typically rich in potatoes, vegetables, meat, eggs, but also characterised with a high content of fibre (black bread) and several dairy products. The trial code was broken after 8 weeks. At each measurement time-point (baseline, 4 and 8 weeks), the subjects underwent a clinical examination. Plasma samples were collected after an overnight fast and abstinence from any medications, tobacco, alcohol, tea, and coffee. Each participant was evaluated for anthropometric indices. Body mass index (BMI) was calculated as the weight (kg) divided by squared height (m2) [25]. A sphygmomanometer was used to measure blood pressure, after subjects rested for 5 min in the sitting position. After the blood pressure measurement, blood samples were collected. Markers for determining cardiovascular health were selected based on suggestions by the NDA of the European Food Safety Authority [26]. At baseline, the two groups did not significantly differ in the selected anthropometric, clinical, and biochemical indices (Table 1). Similarly, at the end of the trial (8 weeks), the PG comprised 36 women and 7 men, with mean BMI 26.5 kg/m2, and mean age 50.4 y; the CG comprised 30 women and 3 men, with mean BMI 27.1 kg/m2, mean age 50.8 y. The two groups showed no significant differences. All volunteers were considered clinically healthy according to the tested baseline and reference values (Nordic Reference Interval Project; NORIP). Significant reductions in serum LDL-C and TG were considered the primary outcome measures for lowering the CVD risk score.
The obtained faecal samples were stored in a domestic refrigerator at 4 °C for no more than 2 h before transportation to the laboratory. There, the samples were frozen at −80 °C until analyses for determining the L. fermentum ME-3 prevalence and counts.
Measurement of oxidized LDL
Oxidized LDL (oxLDL) was measured with an ELISA kit (Cat. No. 10-1143-01, Mercodia AB, Uppsala, Sweden) [17]. This solid-phase, two-site enzyme immunoassay was based on the direct sandwich technique, in which two monoclonal antibodies were directed against separate antigenic determinants on the oxidized apolipoprotein B molecule [27]. During incubation, oxLDL in the sample reacted with the anti-oxLDL antibodies bound to the microtitration wells. After washing, a peroxidase-conjugated anti-human apolipoprotein B antibody recognised the oxLDL bound to the solid phase. After a second incubation and washing, the bound conjugate was detected in a reaction with 3,3′,5,5′-tetramethylbenzidine. The reaction was stopped by adding acid, and the colour intensity (U/l) was measured spectrophotometrically at 450 nm.
Metabolic indices (plasma glucose, total cholesterol, LDL-C, HDL-C, and TG) were analysed with standard laboratory methods and certified assays in the clinical laboratory of the Tartu University Clinics, Estonia. As references, we used indices for routine laboratory tests proposed by the Nordic Reference Interval Project (NORIP, http://www.tandfonline.com/doi/pdf/10.1080/00365510410006324#.Vhpmx27-VCE).
Molecular assessment of L. fermentum ME-3
Bacterial DNA was extracted from faecal samples with a QIAamp DNA stool mini kit (QIAgen, Hilden, Germany) with some modifications. Faeces (0.22 g) was suspended in 200 μl TE buffer (10 mM Tris, 10 mM EDTA pH 8, 20 mg/ml lysozyme, and 200 U/ml mutanolysin), and incubated for 1 h at 37 °C. To these samples, we added 0.3 g of 0.1-mm zirconia/silica beads and 1.4 ml ASL solution from the stool mini kit. The tubes were then agitated for 3 min at 5000 rpm in a mini-bead beater (Biospec Products Inc., USA). The protocol was completed as described by the manufacturer (QIAgen, Germany).
To establish a quantitative assay, we created plasmids that contained the PCR-amplified region of target bacteria with the pGEM-T vector system (Promega, Madison, WI). The PCR amplicon derived from L. fermentum ME-3 was inserted into the plasmid vector, and the recombinant vector was transformed into chemically competent E. coli. Plasmids were purified with a MaxiPrep kit (Qiagen), and then, serially diluted, and finally, measured in a spectrophotometric analysis (Quibit™, Invitrogen) [28]. The target DNA was quantified by comparing with serial, 10-fold dilutions (101 to 105 plasmid copies) of previously quantified plasmid standards. Comparisons between the plasmid standards and samples were run in triplicate.
Real-time PCR was performed with the ABI PRISM 7500 HT Sequence Detection System (Applied Biosystems, USA) equipped with optical grade 96-well plates. For detection of L. fermentum ME-3, the reaction mixture (25 μl) for the TaqMan assay contained 2× TaqMan Universal PCR Master Mix (PE Applied Biosystems, USA), 25 pmol primers (ME31 forward: 3′-CTTTTTACCGCCAAAAGCAG-5′; ME-3 reverse: 3′-AGCCCTTGTCGGTGATATTG-5′), 10 pmol of TaqMan probe (5′-FAM-TTGACACAGACTCGAGCAGT-Tamra-3′), and 200 ng of extracted DNA. The thermocycling program included an initial cycle of 95 °C for 10 min, followed by 45 cycles of 95 °C for 10 s and 60 °C for 1 min. Data was analysed with Sequence Detection Software, version 1.6.3 (Applied Biosystem, USA). L. fermentum ME-3 quantities were expressed as the log10 number of gene copies/g faeces.
Statistical analysis
Statistical analyses were performed with the R program, v. 3 · 0 · 2 (A Language and Environment, http://www.r-project.org) and GraphPad Prism, version 4 · 00 for Windows (GraphPad Software, San Diego, CA). All data were expressed as the mean and standard deviation (SD; Tables 1 and 2) or as the medians with two-sided confidence intervals for counts of BHS and L. fermentum ME-3. Sex and temporal colonisation with probiotic bacteria were compared between groups with Fisher’s exact test (Table 1, Fig. 3). Baseline and intervention data (BL 1–4 and BL 2–8 weeks) were compared with a paired t-test or with a Wilcoxon signed-rank test, according to the data distribution (Table 2). The change (mean ± SD) of individual biochemical indices from baseline of particular participants to 4 and 8 weeks in the probiotic and control groups was assessed with the paired t-test (Fig. 2). The extent of changes between probiotic and control groups (the PG and CG groups) was compared with the t-test. Differences were considered statistically significant when P was <0.05 (Table 2).

The reported changes, from the adjusted baseline 2 to 8 weeks, for individuals in the probiotic and control groups. Legend: Changes (mean ± SD) are shown for (LDL serum LDL-C, (TG) serum TG, and (oxLDL) serum oxLDL (paired t-test); the extent of changes was compared between probiotic and control groups (unpaired t-test)
Results
Bile salt hydrolysis activity
The bile salt hydrolysis activity (BSH) of L. fermentum ME-3 and L. plantarum BAA793 was equal: in both the median (CI 95 %) was 1.5 (1.5). Still it was somewhat higher than those of the other two control strains, L. acidophilus 821–3 and L. plantarum 299v with 1.0 (1.0) and 0.5 (048–1.48), respectively.
Changes in body mass index, blood pressure and lipid fractions
Consumption of probiotic kefir for 4 or 8 weeks did not influence the BMI nor blood pressure values. No BMI or blood pressure changes were observed in either the PG or CG (data not shown).
After 4 weeks, the lipid profiles were mostly similar between groups. Still, the TG level in the PG was reduced (P = 0.005), and this effect was sustained at 8 weeks of probiotic kefir consumption (17 % reduction, P = 0.033). The ox-LDL levels also decreased with probiotic intervention. In the PG, the ox-LDL decreased by 5 % at 4 weeks (P < 0.001) and by 6 % (P < 0.001) at 8 weeks, but no change was observed in the CG.
After 8 weeks of intervention, the PG group exhibited significant reduction in LDL-C (5 %, P = 0.001). Thus, after this period the differences between the PG and CG groups in the LDL-C, ox-LDL and TG indices (P = 0.037; 0.0002; 0.044; Fig. 2) could be well demonstrated.
Additionally we found that, in the CG, the total cholesterol level was increased from 6.0 mmol/l at baseline to 6.2 mmol/l after 8 weeks of kefir consumption (P = 0.031), which trend was not detected in the PG. To characterise additional clinically important changes in the lipid profile (also known as the atherogenic index), we calculated the ratio of LDL-C to HDL-C (Table 2). After 8 weeks, this ratio was decreased only in the PG (P = 0.013) while in the CG it was significantly increased (P = 0.018). As a result, the extent of changes between groups in total cholesterol and LDL-C/HDL-C ratio occurred significant (P values 0.014 and 0.004, respectively) which showed a beneficial effect from the probiotic kefir intervention absent with ordinary kefir.
Glycated haemoglobin (HbA1c)
Glycated haemoglobin levels were reduced after 4 and 8 weeks in both the PG and CG. However, no significant shifts were observed in the corresponding glucose values (Table 2).
Molecular detection of L. fermentum ME-3 with real-time PCR
The prevalence and counts of probiotic bacteria were evaluated in the collected faecal samples with real-time PCR and specific L. fermentum ME-3 primers. At baseline, L. fermentum ME-3 was found in samples from 10 individuals. The PG exhibited significant increases in the prevalence of L. fermentum ME-3. Only 8 % of subjects harboured L. fermentum ME-3 at baseline, but the prevalence rose to 62 % at 4 weeks and 56 % at 8 weeks (both P < 0.001) (Fig. 3). These results suggested that more than half the volunteers in the PG exhibited temporal colonization by L. fermentum ME-3.

Prevalence (%) of L. fermentum ME-3 observed among faecal samples obtained from subjects in the probiotic and control groups. Legend: The presence of L. fermentum ME-3 was determined with real-time PCR and L. fermentum ME-3-specific primers. The baseline values represent the numbers of BL1 both for PG and CG
The median counts did not change over time; at baseline, the median (CI 95 %) was 3.31 (2.20–4.95) log10 gene copies/g faeces, and at 4 and 8 weeks, the median counts were 4.83 (4.47–4.40) and 4.40 (4.11–4.67) log10 gene copies/g faeces, respectively.
Discussion
The present randomised, double-blind, placebo-controlled, parallel-design, two-armed study (ISRCTN 49744186) aimed to evaluate whether consumption of a functional dairy food product could cause reductions in borderline-high serum LDL-C and serum TG values in Estonian adults. The main finding of our study was the positive primary outcome: the significant reductions in LDL-C (5 %) and TG (17 %) levels in the volunteers that consumed probiotic kefir with L. fermentum ME-3 for 8 weeks. Present results show that daily consumption of kefir with antioxidative BSH-active L. fermentum ME-3 (DSM14241) over longer period of time is efficacious for regulating the lipid profile in borderline hyper-cholesterolaemic subjects. This finding supports the numerous clinical trials [3, 4] having demonstrated that LDL-C reduction is the main factor in prevention of the development of coronary heart disease. Earlier studies have demonstrated that some lactic acid bacteria, such as L. plantarum 299v and L. acidophilus L1, could lower LDL-C levels [29, 30], and that L. reuteri NCIMB 30242 after different periods of consumption (from 4 to 6 weeks) reduced both LDL-C and total cholesterol levels of adults [31]. In children with dyslipidemia a formulation of three strains of bifidobacteria caused up to 4 % reduction of TG and LDL-C values after 3 months consumption [32]. In contrast, other probiotic preparations, such as VSL#3 comprising different probiotic strains of Lactobacillus and Bifidobacterium species reduced the TG levels in critically ill patients, but did not influence the LDL-C [33]. In the present investigation, the gut microbial ecosystem was the target of the probiotic Lactobacillus sp., and the kefir served as a probiotic carrier. The antioxidative and anti-atherogenic effects of L. fermentum ME-3 were previously described in numerous in vitro experiments, animal experiments, and in human trials, with different dairy products, including goat milk, cheese, and yoghurt [14, 16–19].
The positive effects of lactobacilli on various markers of lipid metabolism in the human body are likely due to the sum of multiple different mechanisms [34]. Pereira et al. [35] demonstrated that short-chain fatty acid concentrations, specifically the molar proportion of propionate and/or bile salt deconjugation, comprised the major mechanism involved in the purported cholesterol-lowering properties of L. fermentum. Lee et al. (2010) revealed that several regulatory proteins mediated serum cholesterol reduction by a probiotic strain [36]. Lactic acid bacteria, including L. fermentum strains, could assimilate cholesterol, and thus, lower serum cholesterol levels [37]. However, the effect of Lactobacillus spp. strains on levels of serum cholesterol and cholesterol fractions is strain-specific, and depends on the origin, properties and formulation of a certain strain [38–40]. In the present study, the human origin strain, L. fermentum ME-3, exhibited considerable BHS activity. Moreover, we found that the reduction in LDL-C level was consistent with previous data that showed that L. fermentum ME-3 reduced the serum ox-LDL levels [17–19].
Another potential mechanism involved in the cholesterol-lowering properties of L. fermentum is related to the observation that L. fermentum ME-3 showed strong glycosyl-hydrolase activities. These activities included alpha-galactosidase, beta-galactosidase, alpha-glycosidase, beta-glycosidase, and beta-glycuronidase [41]. The concerted actions of these enzymes derived from L. fermentum caused the release of a number of bioactive compounds from various large complexes formed from foodstuffs; some of these bioactive compounds could affect the blood lipid profile [37]. For example, some aglycones derived from food-borne glycosides had positive effects on the blood lipid profile by increasing hepatic fatty acid oxidation and inhibiting cholesterol synthesis [42–44].
Additionally, it is widely known that the high postprandial response of serum TG is related to an elevated insulin response and an elevated hepatic synthesis of TG and cholesterol. We recently showed that consumption of L. fermentum ME-3 kefir after a standard breakfast led to a significantly reduced postprandial TG response [18]. This result may also contribute to the positive effects of L. fermentum ME-3 on the blood lipid profile, when it is consumed long-term, like that established in the present study. It was remarkable that L. fermentum ME-3 consumption reduced the TG more rapidly (4 weeks vs. 8 weeks) than it impacted the LDL-C levels.
In the present study, we tested healthy volunteers with borderline-high levels of lipid fractions at baseline. This design enabled us to document the efficacy of L. fermentum ME-3 consumption. Other authors have also highlighted a need to select groups with borderline values of biomarkers from the general population [12]. The present study recruitment design was quite similar to that used in testing efficacy in individuals with diseases, where the proved diagnosis served as the basis for consecutive patient selection. In our study, all randomly selected individuals of both intervention groups were healthy and agreed to maintain a standard Western-type diet rich in potatoes, vegetables, meat, eggs, one also characterized by a high content of grain fibre and dairy products. A limitation of this study is that there is no evaluation of the dietary intake in terms of its possible implications. Variations in diet over weeks may be responsible for some of the changes observed in lipid profile between probiotic and ordinary kefir. However, none of the routine clinical indices such as body mass index or blood pressure changed either in the probiotic or control groups. Thus the work presented here suggests the special regulation of the host lipid metabolism with a probiotic strain Lactobacillus fermentum ME-3. The main limitation of this study was the high number of drop-outs. Remarkably, only two persons in the PG and two in the CG complained of gastrointestinal symptoms, the other drop-out persons were affected mainly with an outbreak of respiratory infections or discontinued the intervention for loss of motivation to follow the prescriptions during the long study. However, we observed significant changes in the primary outcome measure LDL-C.
Previous studies have shown particularly good faecal recovery of L. fermentum ME-3 after consumption of probiotic yoghurt and cheese [17, 45]. The probiotic kefir tolerates the technology involved in preparation, and it remains viable throughout its shelf life [18]. However, the probiotic strain was detected in only 62 % of faecal samples after 4 weeks and 56 % after 8 weeks. These results may be due to the relatively low dose consumed (8 × 109 cfu/day). Saxelin et al. (1995) reported that a dose above 1010 cfu/day was necessary for even brief persistence of a probiotic strain [46]. Nevertheless, the L. fermentum ME-3 counts in positive faecal samples were quite high (up to 106 copy numbers/g) and the beneficial shifts in lipid profiles confirmed the presence of L. fermentum ME-3 in the gut, seemingly in the higher parts, where metabolites are absorbed. A recent clinical trial tested 3 weeks of consuming probiotic cheese with 2 × 1010 cfu/day of L. plantarum Tensia. Similarly, faecal recovery was detected in only 64 % of patients on a hypocaloric diet, although compliance was high, due to strict control by a hospital nurse [47]. Moreover, the faecal counts of L. plantarum Tensia did not exceed 103gene copies/g, which was substantially lower than with L. fermentum ME-3. Our results suggested that the intervention should last at least 8 weeks, based on the expected primary outcome, despite the equivalent prevalence of L. fermentum ME-3 temporal colonization after 4 and 8 weeks. Interestingly, the prevalence of L. fermentum as species was reported to be 21 % in Estonian adults [48]. In the present study, we found that 9 % of subjects at baseline were persistently colonized with L. fermentum ME-3 strain.
In Estonia, L. fermentum ME-3 has been marketed for the past 10 years in dairy products. This may have facilitated its symbiosis with the hosts. In future investigations, it would be interesting to assess how a sustainable colonization of the probiotic strain is influenced by baseline data of different indigenous lactobacilli to regulate the health indices of the general population.
It was recently shown that total cholesterol does not rank very high in either its long-term impact or as a lipid marker, in terms of its clinical validity, response to changes in therapy, and long-term signal-to-noise ratio [2] Thus, our findings that the probiotic L. fermentum ME-3 had a beneficial effect on LDL-C, ox-LDL, the LDL-C to HDL-C ratio, and TG levels indicated that the blood lipid anti-atherogenic profile had improved, which translated as a reduced risk of CVD.
Conclusion
Eight weeks of consuming kefir with the antioxidative probiotic L. fermentum ME-3, reduced serum LDL-C, ox-LDL and TG values in clinically healthy volunteers with borderline-high lipid profile indices. Thus, L. fermentum ME-3 has the potential to lower the risk of CVD that is tightly associated with maintenance of plasma lipid profile.
References
Mendis S, Puska P, Norrving B. Global atlas on cardiovascular disease prevention and control. World Health Organization, Geneva, Switzerland; 2011 (Available at: http://whqlibdoc.who.int/publications/2011/9789241564373_eng.pdf).
Glasziou PP, Irwig L, Kirby AC, Tonkin AM, Simes RJ. Which lipid measurement should we monitor? An analysis of the LIPID study. BMJ Open. 2014;4:e003512.
Saenger A. Cardiovascular risk assessment beyond LDL cholesterol: non-HDL cholesterol, LDL particle number and apolipoprotein B. Mayo Clinic Communiqué 2011;36(6):1–9.
Reiner Z, Catapano AL, De Backer G, Graham I, Taskinen M, Wiklund O, et al. ESC/EAS Guidelines for the management of dyslipidaemias: The Task Force for the management of dyslipidaemias of the European Society of Cardiology (ESC) and the European Atherosclerosis Society (EAS). Eur Heart J. 2011;32:1769-818.
National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III). Third Report of the National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III) final report. Circulation 2002;106:3143–421.
Morelli L. Postnatal development of intestinal microflora as influenced by infant nutrition. J Nutr. 2008;138:1791S–5S.
Canny GO, McCormick BA. Bacteria in the intestine, helpful residents or enemies from within? Infect Immun. 2008;76:3360–73.
Cani PD, Delzenne NM. The Role of the Gut Microbiota in Energy Metabolism and Metabolic Disease. Curr Pharm Des. 2009;15:1546–58.
Sommer F, Baeckhed F. The gut microbiota - masters of host development and physiology. Nat Rev Microbiol. 2013;11:227–38.
Ishibashi S, Schwarz M, Frykman PK, Herz J, Russell DW. Disruption of cholesterol 7α-hydroxylase gene in mice. I. Postnatal lethality reversed by bile acid and vitamin supplementation. J Biol Chem. 1996;271:18017–23.
Gilliland SE, Nelson CR, Maxwell C. Assimilation of cholesterol by Lactobacillus acidophilus. Appl Environ Microbiol. 1985;49:377–81.
Guo Z, Liu XM, Zhang QX, Shen Z, Tian FW, Zhang H, et al. Influence of consumption of probiotics on the plasma lipid profile: a meta-analysis of randomised controlled trials. Nutr Metab Cardiovasc Dis. 2011;21:844–50.
Kullisaar T, Zilmer M, Mikelsaar M, Vihalemm T, Annuk H, Kairane C, et al. Two antioxidative lactobacilli strains as promising probiotics. Int J Food Microbiol. 2002;72:215–224.
Annuk H, Shchepetova J, Kullisaar T, Songisepp E, Zilmer M, Mikelsaar M. Characterization of intestinal lactobacilli as putative probiotic candidates. J Appl Microbiol. 2003;94:403–12.
Hutt P, Shchepetova J, Loivukene K, Kullisaar T, Mikelsaar M. Antagonistic activity of probiotic lactobacilli and bifidobacteria against entero- and uropathogens. J Appl Microbiol. 2006;100:1324–32.
Truusalu K, Naaber P, Kullisaar T, Tamm H, Mikelsaar R-H, Zilmer K, et al. The influence of antibacterial and antioxidative probiotic lactobacilli on gut mucosa in a mouse model of Salmonella infection. Micr Ecol Health Dis. 2004;16:180–187.
Kullisaar T, Songisepp E, Mikelsaar M, Zilmer K, Vihalemm T, Zilmer M. Antioxidative probiotic fermented goats’ milk decreases oxidative stress-mediated atherogenicity in human subjects. Br J Nutr. 2003;90:449–56.
Kullisaar T, Shepetova J, Zilmer K, Songisepp E, Rehema A, Mikelsaar M, et al. An antioxidant probiotic reduces postprandial lipemia and oxidative stress. Centr Eur J Biol. 2011; 6: 32–40.
Songisepp E, Kals J, Kullisaar T, Mändar R, Hütt P, Zilmer M, et al. Evaluation of the functional efficacy of an antioxidative probiotic in healthy volunteers. Nutr J. 2005;4;22.
Hutt P, Andreson H, Kullisaar T, Vihalemm T, Unt, E, Kals J, et al. Effects of a synbiotic product on blood antioxidative activity in subjects colonized with Helicobacter pylori. Lett Appl Microbiol. 2009;48:797–800.
Kaur S, Kullisaar T, Mikelsaar M, Eisen M, Rehema A, Vihalemm T, et al. Successful management of mild atopic dermatitis in adults with probiotics and emollients. Cent Eur J Med, 2008;3:215–220.
Mikelsaar M, Annuk H, Stsepetova J, Mändar R, Sepp E, Björksten B. Intestinal lactobacilli of Estonian and Swedish children. Microb Ecol Health Dis. 2002;14:75–80.
Mikelsaar M, Zilmer M. Lactobacillus fermentum ME-3 - an antimicrobial and antioxidative probiotic. Microb Ecol Health Dis. 2009;21:1–27.
Lim HJ, Kim SY, Lee WK. Isolation of cholesterol-lowering lactic acid bacteria from human intestine for probiotic use. J Vet Sci. 2004;5:391–5.
WHO. Physical status: the use and interpretation of anthropometry. Report of a WHO Expert Committee. World Health Organ Tech Rep Ser. 1995;854:1–452.
EFSA. Guidance on the scientific requirements for health claims related to antioxidants, oxidative damage and cardiovascular health. EFSA J. 2011;9:2474. doi:10.2903/j.efsa.2011.2474.
Holvoet P, Vanhaecke J, Janssens S, Van de Werf F, Collen D. Oxidized LDL and malondialdehyde-modified LDL in patients with acute coronary syndromes and stable coronary artery disease. Circulation. 1998;98:1487–94.
Bartosch S, Fite A, Macfarlane GT, McMurdo ME. Characterization of bacterial communities in feces from healthy elderly volunteers and hospitalized elderly patients by using real-time PCR and effects of antibiotic treatment on the fecal microbiota. Appl Environ Microbiol. 2004;70:3575–81.
Naruszewicz M, Johansson ML, Zapolska-Downar D, Bukowska H. Effect of Lactobacillus plantarum 299v on cardiovascular disease risk factors in smokers. Am J Clin Nutr. 2002;76:1249–55.
Anderson JW, Gilliland SE. Effect of fermented milk (yogurt) containing Lactobacillus acidophilus L1 on serum cholesterol in hypercholesterolemic humans. J Am Coll Nutr. 1999;18:43–50.
Jones ML, Martoni CJ, Prakash S. Cholesterol lowering and inhibition of sterol absorption by Lactobacillus reuteri NCIMB 30242: a randomized controlled trial. Eur J Clin Nutr. 2012;66:1234–41.
Guardamagna O, Amaretti A, Puddu PE, Raimondi S, Abello F, Cagliero P, et al. Bifidobacteria supplementation: effects on plasma lipid profiles in dyslipidemic children. Nutrition, 2014;30,831–836. doi:10.1016/j.nut.2014.01.014.
Sanaie S, Ebrahimi-Mameghani M, Mahmoodpoor A, Shadvar K, Ej GS. Effect of a Probiotic Preparation (VSL#3) on CardiovascularRisk Parameters in Critically-Ill Patients. J Cardiovasc Thorac Res. 2013;5:67–70.
Drissi F, Merhej V, Angelakis E, Kaoutari A, Carriere F, Henrissat B, et al. Comparative genomics analysis of Lactobacillus species associated with weight gain or weight protection. Nutr Diabetes. 2014;4:e109.
Pereira DI, McCartney AL, Gibson GR. An in vitro study of the probiotic potential of a bile-salt-hydrolyzingLactobacillus fermentum strain, and determination of its cholesterol-lowering properties. Appl Environ Microbiol. 2003;69:4743–52.
Lee J, Kim Y, Yun HS, Kim JG, Oh S, Kim SH. Genetic and Proteomic Analysis of Factors Affecting Serum Cholesterol Reduction by Lactobacillus acidophilus A4. Appl Environ Microbiol. 2010;76:4829–35.
Tulumoglu S, Kaya HI, Simsek O. Probiotic characteristics of Lactobacillus fermentum strains isolated from tulum cheese. Anaerobe. 2014;30:120–5.
Tanaka H, Doesburg K, Iwasaki T, Mierau I. Screening of lactic acid bacteria for bile salt hydrolase activity. J Dairy Sci. 1999;82:2530–5.
Ataie-Jafari A, Larijani B, Alavi Majd H, Tahbaz F. Cholesterol-lowering effect of probiotic yogurt in comparison with ordinary yogurt in mildly to moderately hypercholesterolemic subjects. Ann Nutr Metab. 2009;54:22–7.
Jones ML, Martoni CJ, Parent M, Prakash S. Cholesterol-lowering efficacy of a microencapsulated bile salt hydrolase-active Lactobacillus reuteri NCIMB 30242 yoghurt formulation in hypercholesterolaemic adults. Br J Nutr. 2012;107:1505–13.
Uskova MA. Investigation of probiotic lactic acid bacteria as biologically active components of food. Moscow: PhD dissertation; 2010.
Zang M, Xu S, Maitland-Toolan KA, Zuccollo A, Hou X, Bingbing J, et al. Polyphenols stimulate AMP-activated protein kinase, lower lipids, and inhibit accelerated atherosclerosis in diabetic LDL receptor-deficient mice. Diabetes. 2006;55:2180–91.
Mulvihill EE, Allister EM, Sutherland BG, DiMattia AS, Khami M, Koppesd JB, et al. Naringenin prevents dyslipidemia, apolipoprotein B overproduction, and hyperinsulinemia in LDL receptor-null mice with diet-induced insulin resistance. Diabetes. 2009;58:2198–210.
Chen G, Wang H, Zhang X, Yang S-T. Nutraceuticals and Functional Foods in the Management of Hyperlipidemia. Crit Rev Food Sci Nutr. 2014;54:1180–201.
Songisepp E, Kullisaar T, Hutt P, Elias P, Brilene T, Zilmer M, et al. A new probiotic cheese with antioxidative and antimicrobial activity. J Dairy Sci 2004; 87:2017–23.
Saxelin M, Pessi T, Salminen S. Fecal recovery following oral administration of Lactobacillus strain GG (ATCC 53103) in gelatine capsules to healthy volunteers. Int J Food Microbiol. 1995;25:199–203.
Sharafedtinov KK, Plotnikova OA, Alexeeva RI, Sentova TB, Songisepp E, Stsepetova J, et al. Hypocaloric diet supplemented with probiotic cheese improves body mass index and blood pressure indices of obese hypertensive patients - a randomized double-blind placebo-controlled pilot study. Nutr J. 2013;12:138.
Štšepetova J, Sepp E, Kolk H, Lõivukene K, Songisepp E, Mikelsaar M. Diversity and metabolic impact of intestinal Lactobacillus species in healthy adults and the elderly. Br J Nutr. 2011;105:1235–44.
Acknowledgements
This study was financially supported by the European Union through the European Regional Development Fund (SFOS reg. no. 3.2.0701.11-0023), the Estonian Ministry of Education and Research (target No. SF0180132s08 and financing of scientific collections KOGU-HUMB), and the University of Tartu (grant no.SARMBARENG). We appreciate the technical assistance from Laboratory for Clinical and Physiological Research of the Bio-Competence Centre of Healthy Dairy Products, the assistance by PhD student, Merle Rätsep, and the help with statistics of Karolin Toompere.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
University of Tartu, Estonia holds the US patent (US7,244 424 B2 July 17,2007) on the antioxidative probiotic Lactobacillus fermentum ME-3 while 3/6 authors of the manuscript MM, TK and MZ are among the authors of the patent. For the study the authors of the paper have got financial support only from EU regional project and Estonian Science foundations. Thus, the authors of the manuscript declare no direct competing financial and non-financial interests.
Authors’ contributions
MM, ES, and MZ participated in the design and coordination of the study; JS applied the molecular methods; PH investigated the patients and collected the clinical data; TK, KZ, and PH performed the clinical laboratory tests and microbiological assays; MM, JS, PH, ES, TK, and MZ participated in writing the manuscript. All authors read and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Mikelsaar, M., Sepp, E., Štšepetova, J. et al. Regulation of plasma lipid profile by lactobacillus fermentum (probiotic strain ME-3 DSM14241) in a randomised controlled trial of clinically healthy adults. BMC Nutr 1, 27 (2015). https://doi.org/10.1186/s40795-015-0020-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40795-015-0020-z