
Abstract
Background
Foraging theory predicts that animals select patches that offer the highest net rate of energy gain. Hence, prey distribution patterns and spatiotemporal heterogeneity play important roles in determining animal feeding patch selection. For waterfowl foraging on buried aquatic plant tubers, the distribution and biomass of these plant organs vary with depth in the substrate. Since excavation costs also increase with depth, the energy intake of the animals foraging on these plants is highly sediment depth dependent.
Methods
Here, using observations of Swan Geese (Anser cygnoides) foraging on Vallisneria natans tubers, we test our hypothesis that geese feeding on tubers buried at intermediate sediment depth maximize their daily energy intake because of the interaction between tuber size and abundance with depth. To do this, we measured the distribution patterns of buried Vallisneria tubers under both undisturbed conditions and post-exploitation by geese (i.e. giving-up conditions). We investigated the relationship between tuber size and burial depth, and total tuber biomass within each sediment layer in undisturbed and exploited plots. Finally, we compared modelled Swan Goose daily energy intake feeding on Vallisneria tubers buried at different sediment layers (1–10, 11–20 and 21–30 cm below the surface).
Results
Dry weight of Vallisneria tubers linearly increased with burial depth, while average total dry weight density of tubers showed a unimodal relationship, peaking at intermediate levels. Not surprisingly, Swan Geese foraged most intensively on tubers buried at intermediate sediment depths, where they maximize their daily energy intake. Our results support our hypothesis that Swan Geese feeding on tubers at intermediate depths maximize their daily energy intake.
Conclusions
Our study is the first to quantify foraging strategies of Swan Geese during the wintering period, emphasizing the importance of plant traits on foraging selection of belowground foragers.
Similar content being viewed by others
Background
Animals need to satisfy their daily energy needs, but finding food is energetically costly, so consumers tend to select feeding patches which support the highest energetic intake rates (Murakami 1998), following the energy-maximizing hypothesis (Charnov 1976; Jeschke et al. 2002). Hence, prey or plant distribution patterns and their heterogeneity play an important role in determining densities of foraging predators and herbivores (the aggregative response, e.g., Rowcliffe et al. 1999). For example, herbivores feeding on highly digestible energy-rich tubers of plants aggregate in areas with the highest tuber densities, although this relationship is influenced by other factors affecting availability (Klaassen and Nolet 2007; Nolet and Gyimesi 2013).
Several waterfowl species feed on buried, over-wintering tubers of aquatic plants in underwater substrates, which may contribute to regulate the populations of these plants (Sponberg and Lodge 2005). The extraction costs associated with foraging on tubers increase with increasing tuber burial depth (Richman and Lovvorn 2009), so there is a selection pressure for tubers to be buried deeper in the substrate to avoid predation. However, for a given tuber size, the chance of germinating and sprouting to the surface successfully decreases with burial depth (Klaassen and Nolet 2007; Hamberg et al. 2017). For this reason, we would predict that plants balance the costs of such an avoidance strategy amongst deeply buried tubers by increasing their energy content to enhance the likelihood of successful germination and growth to the surface (Jokela et al. 2000). Hence, larger tubers are often found at deepest levels in the sediment, because costs of germination, shoot initiation and penetration to the surface increase with increasing burial depth (Santamaria and Rodriguez-Girones 2002).
Given this heterogeneity in size class distribution within the sediment, how the foraging strategies of tuber-feeding waterfowl respond to the belowground distribution patterns of over-wintering tubers is of great interest. Herbivores feeding on such belowground tubers may have a trade-off feeding on abundant, low quality tubers at shallow depths versus fewer but more profitable tubers deeper within the substrate, which cost more energy to extract. For example, despite a positive relationship between Fennel Pondweed (Potamogeton pectinatus) tuber size and burial depth, tuber density peaks at intermediate depths. Hence, among observed Bewick’s Swans (Cygnus columbianus) feeding on Fennel Pondweed tubers, studies showed that it was most profitable for swans to forage at intermediate depths (Santamaria and Rodriguez-Girones 2002; Hidding et al. 2009).
The Swan Goose (Anser cygnoides) is a globally vulnerable species, which mainly breeds in Russia, Mongolia and China and overwinters now almost exclusively in China where it is largely restricted to the Yangtze flood plain (Kear 2005; Cao et al. 2008). During winter, when the water level drops below the substrate surface they excavate pits in the substrate from which they extract tubers (Zhang et al. 2011; Kim et al. 2013), although they have also been observed grazing on sedges, Carex and grasses (Fox et al. 2008). In this study, we test the hypothesis that Swan Geese feeding on belowground tubers of Vallisneria mostly extract tubers buried at intermediate sediment depth, because these layers offer the greatest net intake of energy.
Methods
Study area
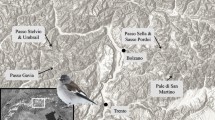
Shengjin Lake National Nature Reserve (30°15ʹ‒30°30ʹN; 116°55ʹ‒117°15ʹE), is situated south of the Yangtze River. It is a seasonally inundated freshwater lake of ca. 140 km2 and a shoreline of 156 km in winter (Fig. 1). During the wet season, water level increases to ca. 17 m a.s.l. and falls to lower than 10 m a.s.l. in the dry season. A sluice built in 1965 is used to regulate water flow between Shengjin Lake and the Yangtze River, allowing water retention in winter when river levels fall. Average annual precipitation is 1600 mm, with most rain during April to August. Average annual temperature is 16 °C and average January temperature is 4 °C.

Location of Shengjin Lake, China (inset bottom right) and positions of Site A and Site B at the lake as described in the text
Study sites
Data presented in these analyses were collected at Sites A and B at Shengjin Lake from mid-December 2008 to early February 2009 (Fig. 1). Site A (30°22′01.47″N; 117°04′10.99″E) was an open flat area covered by water until late November. After early winter water level recession, this site was used by Swan Geese during early December. From mid-December, this site was abandoned because of the combined effects of rapidly receding water levels and dry weather conditions, resulting in a larger area of unexploited habitat. Site B (30°22′01.47″N; 117°04′10.99″E) is a relatively enclosed bay ca. 80 m wide, heavily used by Swan Geese from mid-December 2008 to early February 2009. At both sites, the sediment type is clay and Vallisneria natans was the dominant aquatic plant over-wintering in the form of tubers buried in the muddy lake substrate.
Diet composition
In winter 2008/2009, Swan Goose faeces were collected at the two sites, separately stored in paper bags and oven-dried at 50 °C for 48 h using an oven (DHG-9053A, Shanghai). At least 20 faecal pellets from the same day and site were pooled and mixed thoroughly by hand. Vegetation samples were also collected at each site. Dicotyledonous plant epidermis patterns were derived by coating leaves with nail polish, the layer peeled off when dry and photographs of the impressions of cell patterns were taken under a microscope. Sub-samples of faeces were diluted with water in a petri dish, and drops taken from this suspension, placed on a glass slide, spread out evenly and covered with a cover slip. From each slide, 20 fields were identified using photomicrographs and area measured using a grid of 0.04 mm2 squares in the microscope eyepiece. However, no leaf epidermal fragments were found in the faeces, since all Swan Goose faeces contained fragments of Vallisneria tubers, readily identifiable by lack of cell structure and undifferentiated tissue, as well as compact dark flocculated fragments of tuber outer epidermis. A total of 100 fields were identified for each sample under the microscope (Owen 1975).
Determining Vallisneria tuber distribution
We observed the Swan Goose distribution every week at both Site A and Site B during the winter 2008/2009. The undisturbed distribution of buried Vallisneria tubers was determined in February 2009 at Site A. Seven 30 cm × 30 cm × 30 cm holes were excavated and tubers were extracted in situ within each of these quadrats within the substrate where they were unexploited by Swan Geese in these areas up to the sampling date. The quadrats were carefully excavated using a small shovel by removing mud layers to reveal each tuber. The vertical depth of each tuber from the substrate surface was measured to the nearest 1 cm from the surface level using a ruler. Tubers were put into separate paper bags. By contrast, Site B was subject to intensive foraging after the sampling date (Fig. 2). Hence, after prolonged use by feeding Swan Geese, tubers were again extracted at Site B following the same method from five 30 cm × 30 cm × 30 cm sample holes in February 2009. In the laboratory, all derived Vallisneria tubers were washed and remaining substrate was carefully removed using a toothbrush and dried at 50 °C for 48 h (Fig. 2). The dry weight of each tuber was determined to the nearest 0.001 g using an analytic balance (Sartorius, Germany).

Main photograph shows a view of Site B at the study area at Shengjin Lake, China, showing the excavation pits created by Swan Geese (Anser cygnoides) (shown in detail below left) to extract the overwintering tubers of Vallisneria natans (shown cleaned of sediment below right)
Excavation pit measurements
After Swan Geese had exploited the two sites for feeding on buried tubers, both areas exhibited numerous tuber excavation pits created by the birds. The largest and smallest diameters and the greatest depths of the pits dug by Swan Geese were measured to the nearest centimetre using a ruler (site A: n = 50, site B: n = 28).
Estimated Swan Goose daily energy intake
We estimated daily energy intake of Swan Geese in each layer following the method described by Fox et al. (2011). Swan Goose intake rate was determined using focal sampling. In total, 28 focal samples of individuals were observed. Daily tuber intake was estimated based on the frequency of the observed goose found and swallowed the tuber. Time spent feeding was observed every 15 min using scan sampling based on the observation throughout daytime. Dropping rate was also recorded by focal sampling. Energy content for both tuber and dropping was analysed using a bomb calorimeter. In contrast to the earlier study, we further classified the tuber burial depth into three different layers (layer 1: 1‒10 cm, layer 2: 11‒20 cm and layer 3: 21‒30 cm) and separately calculated the mean tuber dry weight within each layer. Moreover, the observed intake rate (number of tubers consumed per minute) was the average value of three layers in the analysis of Fox et al. (2011). As tuber intake rate is closely linked to tuber density, we therefore adjusted the intake rate of each layer by multiplying the percent of tubers in each layer to determine the average intake rate in each layer. Finally, the daily energy intake was estimated by subtracting the energy content of daily output in droppings from that of daily food intake.
Statistical analysis
We calculated the average tuber dry weight density (g/m2) at each burial depth class based on the undisturbed data. To test if there was a relationship between tuber dry weight and tuber depth, a quadratic regression model was applied to ln-transformed tuber dry weight mass data from the unexploited area. A t-test was used to detect if there is a difference between undisturbed (original) and disturbed (giving-up) tuber total dry weight of each layer.
A t-test was also applied to test for differences in excavation pit greatest depth, largest and smallest diameters between Site A and Site B. The data were square root transformed to reach the assumption of normality. Statistical analyses were conducted in R 2.15.2 or SPSS 19.0.
Results
Diet analysis
Diet analysis showed that all 100 samples taken from December to January comprised 100% Vallisneria tuber fragments and those in February 98% (another 2% was grass), confirming that geese almost exclusively fed on Vallisneria tubers at these sites during these months.
Vallisneria tuber distribution
There was a positive relationship between tuber dry weight and tuber depth for undisturbed original tuber distributions down to 30 cm depth (dry weight (g) = 0.015 × depth (cm) + 0.039, p < 0.001, \(r_{{\;\;{\text{ad}}}}^{2} = \, 0.26\), n = 196; Fig. 3). A quadratic regression model captured the relationship between mean tuber dry weight density (g/m2) and tuber burial depth well, with highest tuber biomass densities at intermediate depths (\(r_{{\;\;{\text{adj}}}}^{2} = \, 0.31\); main term: b = 0.407, t = 3.292, p = 0.004; squared term: b = ‒0.014, t = ‒3.434, p < 0.03; Fig. 4).

Relationship between Vallisneria natans tuber dry weight and sediment depth at Shengjin Lake, China. The solid line shows the least squares fitted regression model for the relationship. The dotted lines show the 95% confidence intervals of predicted values

Relationship between average Vallisneria natans tuber dry weight density (g/m2) and tuber burial depth at Shengjin Lake, China. Predicted values (solid line) were calculated from back-transformed data on the basis of a quadratic regression on ln-transformed tuber dry weight density values
Comparing undisturbed (Site A) and post-feeding (Site B) total tuber dry weight in each layer indicated that the total dry weight in layer 2 was significantly less in Site B post exploitation by geese compared to site A where tubers were undisturbed (t = 3.014, df = 10, p = 0.013). There were no significant differences in total dry weight in layer 1 (t = 0.867, df = 10, p = 0.406) and layer 3 (t = ‒0.033, df = 10, p = 0.974; Fig. 5).

Comparing undisturbed and disturbed (giving-up) Vallisneria natans tuber dry weight in different sediment layers at Shengjin Lake, China. Error bars show 95% confident intervals
Excavation pit dimensions
Mean greatest depth, largest and smallest diameters of tuber pits excavated by Swan Geese were respectively 12.2 ± 0.62, 23.9 ± 1.68 and 18.0 ± 1.06 cm (mean ± 95% CI). There were no differences in depth (t = 1.759, df = 72.207, p = 0.083), long (t = ‒0.070, df = 76, p = 0.945) and short diameters (t = 1.059, df = 76, p = 0.293) of digging pits between site A and site B.
Daily energy intake
The average tuber dry weight of each layer from top to bottom was respectively 0.15 ± 0.02 g (mean ± 95% CI, n = 87), 0.26 ± 0.03 g (n = 97) and 0.35 ± 0.10 g (n = 17). The daily energy intake rate was highest in layer 2, with 2836 kJ per day, compared to 1372 kJ in layer 1 and 663 kJ in layer 3 (Table 1).
Discussion
We demonstrated a linear relationship between Vallisneria tuber dry weight and burial depth, which increased with increasing burial depth in a Yangtze wetland (Fig. 3). However, the tuber dry biomass showed a different pattern, with the highest total biomass levels found at intermediate sediment depth (Fig. 4). Interestingly, we found that Vallisneria tubers buried at intermediate depths were also most heavily harvested by Swan Geese (based on the comparison of undisturbed versus post-feeding total tuber dry weight in different sediment layers and measuring depths of extraction pits). Our energy intake estimations also suggested that Swan geese obtained the highest levels of energy intake by extracting Vallisneria tubers buried at intermediate sediment depth.
The increasing dry weight of Vallisneria tubers with sediment depth reflects results from a study of P. pectinatus tubers, which showed a similar pattern (Hidding et al. 2009). Tubers buried in deep sediment can decrease their predation risk, interpreted as a herbivore avoidance strategy (Santamaria and Rodriguez-Girones 2002). However, the possibility of successful sprouting decreases with increasing burial depth for a tuber of a given size for P. pectinatus (Santamaria and Rodriguez-Girones 2002) and Vallisneria americana (Rybicki and Carter 1986), which may explain the increasing size of tubers buried in deeper sediments. Our result also revealed that Vallisneria natans tubers deepest within the sediment had higher dry weights compared to tubers at shallower depths. Through such a strategy, tubers likely increase their fitness by decreasing their accessibility to Swan Geese, reducing the predation risk and increasing the chances of sprouting and reaching the surface in the following spring.
Our results also suggest that the total tuber dry weight peaked at intermediate depth (11‒20 cm). This result is consistent with the study on P. pectinatus in which the mean tuber density was larger at a depth of 10‒15 cm (Hidding et al. 2012). Although there were some larger tubers distributed in the deepest sediment, the tuber density was smaller, reducing profitability to Swan Geese in the face of mounting costs of extraction from the lowest levels. However, this result may indicate that plants have invested in deep burial of large tubers as an adaptation to herbivore predation. Such anti-herbivore strategy, achieved through phenotypic plasticity, is well known among plant species as a means to maximize relative fitness (Hanley et al. 2007; Ward et al. 2012). However, as our results are only based on 1 year of data, we are cautious about making such claims for V. natans, for which a long-term study is necessary to fully understand the plant’s tuber anti-herbivore strategy.
Swan Goose foraging strategy and the energy intake estimation
The diet analysis showed that Swan Geese intensively feed on Vallisneria tubers at Shengjin Lake although our former study showed that they also grazed on grasses and sedges (Fox et al. 2008). Swan Goose feeding behaviours can be affected by water level fluctuations (Zhang et al. 2011), influencing their diet composition. Field observation showed that Swan Geese spent more time on feeding when grazing on sedges comparing to excavating Vallisneria tubers (Fox et al. 2008). One potential explanation could be explained by differences in food digestibility, because the fibre content of Vallisneria tubers is much lower than that of graminoid species. Studies have shown that energy intake rate is often limited by the food processing rate or rate of nutrient absorption in the digestive tract (Kersten and Visser 1996; Wilmshurst et al. 1999; Oudman et al. 2014), the maximum rate of energy assimilation (Weiner 1992; Ydenberg 1998). In winter, birds need to maximize their probability of survival for breeding in the next year (Caraco 1979), hence increasing foraging efficiency and predator avoidance are paramount. As time limitation, predation and disturbance risk also increase with increasing feeding time (Chudzinska et al. 2015), and it is essential for Swan Geese to aim to maintain the highest feeding efficiency possible. Foraging on Vallisneria tubers may be more efficient in terms of energy intake and time spent feeding than grazing poorer quality graminoids. Such evidence was also found for Tundra Swans that fed on Fennel Pondweed tubers, because of the high-energy assimilation rate, which these provided (90%; Klaassen and Nolet 2007).
In line with our prediction, tubers buried at intermediate depth were most heavily harvested, based on the depth of extraction pits and comparing the differences between undisturbed and disturbed tuber dry weight between layers (Fig. 5). Optimal foraging theory predicts that animals should maximize their energy intake per unit time (Emlen 1966; Macarthur and Pianka 1966; Pyke 1984). Foraging on higher biomass patches (in this case intermediate sediment depths) can increase foraging success and increase energy intake rate (Bergman et al. 2001). To maximize fitness, animals behave as time minimizer or energy maximizer, although these may not be mutually exclusive (Langvatn and Hanley 1993). Anatidae species often use a strategy of energy maximization (Amano et al. 2004; Nolet et al. 2006; Wang et al. 2013). Digging for deeper buried tubers by Swan Geese is a time and energy-consuming process but geese benefit from the larger tuber sizes there. Our result showed that Swan Geese had a daily intake (2836 kJ) by digging for tubers at intermediate depth, where biomass density was double and fourfold compared to digging shallower or deeper layers respectively (Table 1). Daily energetic expenditure (DEE) of Swan Geese was calculated as 2200 kJ per day and the energy excreted from the production of droppings was estimated as 212 kJ per day (Fox et al. 2011). The net daily energy intake was then 2624 kJ for digging in intermediate deep sediments, as calculated from the difference between daily energy intake and energy excreted in droppings, which was higher than calculated DEE. Net energy intake acquired by digging in other layers was far less than DEE. Although we may overestimate daily energy intake as Swan Geese also fed in shallower sediments, the species can maximize their daily energy intake by foraging more at intermediate sediment depths. Migratory birds accumulate body mass in late winter in preparation for migration (Alerstam and Lindström 1990; Fox et al. 2003). By following this strategy, Swan Geese are potentially able to improve their energy balance and prepare for spring migration by active accumulation of energy stores.
Conclusions
In summary, our results are in agreement with our hypotheses that Swan Geese foraging on tubers buried at intermediate sediment depth maximize their daily energy intake. Although foraging in deeper sediments is energetically more costly, food biomass and density interact to play a critical role in determining the animals foraging selection, emphasizing the importance of plant traits on foraging selection of belowground foragers such as the Swan Goose.
References
Alerstam T, Lindström Å. Optimal bird migration: the relative importance of time, energy, and safety. In: Gwinner E, editor. Bird migration. Berlin: Springer; 1990. p. 331–51.
Amano T, Ushiyama K, Fujita G, Higuchi H. Alleviating grazing damage by white-fronted geese: an optimal foraging approach. J Appl Ecol. 2004;41:675–88.
Bergman CM, Fryxell JM, Gates CC, Fortin D. Ungulate foraging strategies: energy maximizing or time minimizing? J Anim Ecol. 2001;70:289–300.
Cao L, Barter M, Lei G. New Anatidae population estimates for eastern China: implications for current flyway estimates. Biol Conserv. 2008;141:2301–9.
Caraco T. Time budgeting and group-size—test of theory. Ecology. 1979;60:618–27.
Charnov EL. Optimal foraging, the marginal value theorem. Theor Popul Biol. 1976;9:129–36.
Chudzinska ME, van Beest FM, Madsen J, Nabe-Nielsen J. Using habitat selection theories to predict the spatiotemporal distribution of migratory birds during stopover—a case study of pink-footed geese Anser brachyrhynchus. Oikos. 2015;124:851–60.
Emlen JM. The role of time and energy in food preference. Am Nat. 1966;100:611–7.
Fox AD, Cao L, Zhang Y, Barter M, Zhao MJ, Meng FJ, Wang SL. Declines in the tuber-feeding waterbird guild at Shengjin Lake National Nature Reserve, China—a barometer of submerged macrophyte collapse. Aquat Conserv. 2011;21:82–91.
Fox AD, Glahder CM, Walsh AJ. Spring migration routes and timing of Greenland white-fronted geese—results from satellite telemetry. Oikos. 2003;103:415–25.
Fox AD, Hearn RD, Cao L, Cong PH, Wang X, Zhang Y, Dou ST, Shao XF, Barter M, Rees EC. Preliminary observations of diurnal feeding patterns of Swan Geese Anser cygnoides using two different habitats at Shengjin Lake, Anhui Province, China. Wildfowl. 2008;58:20–30.
Hamberg J, Findlay SEG, Limburg KE, Diemont SAW. Post-storm sediment burial and herbivory of Vallisneria americana in the Hudson River estuary: mechanisms of loss and implications for restoration. Restor Ecol. 2017;25:629–39.
Hanley ME, Lamont BB, Fairbanks MM, Rafferty CM. Plant structural traits and their role in anti-herbivore defence. Perspect Plant Ecol. 2007;8:157–78.
Hidding B, Klaassen M, de Boer T, de Vries PP, Nolet BA. Aquatic plant shows flexible avoidance by escape from tuber predation by swans. Basic Appl Ecol. 2012;13:50–8.
Hidding B, Nolet BA, van Eerden MR, Guillemain M, Klaassen M. Burial depth distribution of fennel pondweed tubers (Potamogeton pectinatus) in relation to foraging by Bewick’s swans. Aquat Bot. 2009;90:321–7.
Jeschke JM, Kopp M, Tollrian R. Predator functional responses: discriminating between handling and digesting prey. Ecol Monogr. 2002;72:95–112.
Jokela J, Schmid-Hempel P, Rigby MC. Dr. Pangloss restrained by the Red Queen—steps towards a unified defence theory. Oikos. 2000;89:267–74.
Kear J. Ducks, geese and swans. Oxford: Oxford University Press; 2005.
Kersten M, Visser W. The rate of food processing in the Oystercatcher: food intake and energy expenditure constrained by a digestive bottleneck. Funct Ecol. 1996;10:440–8.
Kim GY, Ji YK, Ganf GG, Lee CW, Joo GJ. Impact of over-wintering waterfowl on tuberous bulrush (Bolboschoenus planiculmis) in tidal flats. Aquat Bot. 2013;107(9):17–22.
Klaassen M, Nolet BA. The role of herbivorous water birds in aquatic systems through interactions with aquatic macrophytes, with special reference to the Bewick’s Swan—Fennel Pondweed system. Hydrobiologia. 2007;584:205–13.
Langvatn R, Hanley TA. Feeding-patch choice by Red Deer in relation to foraging efficiency—an experiment. Oecologia. 1993;95:164–70.
Macarthur RH, Pianka ER. On optimal use of a patchy environment. Am Nat. 1966;100:603–9.
Murakami M. Foraging habitat shift in the narcissus flycatcher, Ficedula narcissina, due to the response of herbivorous insects to the strengthening defenses of canopy trees. Ecol Res. 1998;13:73–82.
Nolet BA, Gyimesi A. Underuse of stopover site by migratory swans. J Ornithol. 2013;154:695–703.
Nolet BA, Gyimesi A, Klaassen RHG. Prediction of bird-day carrying capacity on a staging site: a test of depletion models. J Anim Ecol. 2006;75:1285–92.
Oudman T, Onrust J, de Fouw J, Spaans B, Piersma T, van Gils JA. Digestive capacity and toxicity cause mixed diets in Red Knots that maximize energy intake rate. Am Nat. 2014;183:650–9.
Owen M. An assessment of fecal analysis technique in waterfowl feeding studies. J Wildl Manag. 1975;39:271–9.
Pyke GH. Optimal foraging theory—a critical review. Annu Rev Ecol Syst. 1984;15:523–75.
Richman SE, Lovvorn JR. Predator size, prey size and threshold food densities of diving ducks: does a common prey base support fewer large animals? J Anim Ecol. 2009;78:1033–42.
Rowcliffe JM, Sutherland WJ, Watkinson AR. The functional and aggregative responses of a herbivore: underlying mechanisms and the spatial implications for plant depletion. J Anim Ecol. 1999;68:853–68.
Rybicki NB, Carter V. Effect of sediment depth and sediment type on the survival of Vallisneria Americana Michx grown from tubers. Aquat Bot. 1986;24:233–40.
Santamaria L, Rodriguez-Girones MA. Hiding from swans: optimal burial depth of sago pondweed tubers foraged by Bewick’s swans. J Ecol. 2002;90:303–15.
Sponberg AF, Lodge DM. Seasonal belowground herbivory and a density refuge from waterfowl herbivory for Vallisneria americana. Ecology. 2005;86:2127–34.
Wang X, Fox AD, Cong PH, Cao L. Food constraints explain the restricted distribution of wintering Lesser White-fronted Geese Anser erythropus in China. Ibis. 2013;155:576–92.
Ward D, Shrestha MK, Golan-Goldhirsh A. Evolution and ecology meet molecular genetics: adaptive phenotypic plasticity in two isolated Negev desert populations of Acacia raddiana at either end of a rainfall gradient. Ann Bot. 2012;109:247–55.
Weiner J. Physiological limits to sustainable energy budgets in birds and mammals—ecological implications. Trends Ecol Evol. 1992;7:384–8.
Wilmshurst JF, Fryxell JM, Colucci PE. What constrains daily intake in Thomson’s gazelles? Ecology. 1999;80:2338–47.
Ydenberg RC. Behavioral decisions about foraging and predator avoidance. In: Dukas R, editor. Cognitive ecology: the evolutionary ecology of information processing and decision making. Chicago: University of Chicago Press; 1998. p. 343–78.
Zhang Y, Cao L, Barter M, Fox AD, Zhao MJ, Meng FJ, Shi HQ, Jiang Y, Zhu WZ. Changing distribution and abundance of Swan Goose Anser cygnoides in the Yangtze River floodplain: the likely loss of a very important wintering site. Bird Conserv Int. 2011;21:36–48.
Authors’ contributions
YZ, ADF, and LC conceived the idea. YC and YZ conducted the field work. YC, YZ and WFD analysed the data. YZ and ADF wrote the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We thank the staff of the Shengjin Lake National Nature Reserve for facilitating the studies.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The data used and analyzed in this study are available from the corresponding authors on reasonable request.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Not applicable.
Funding
This study was supported by the Natural Science Foundation of Jiangsu Province (Grant No. BK20170922), the Key Strategic Program of the Chinese Academy of Sciences, Water Ecological Security Assessment and Great Strategy Research of Middle and Lower Yangtze River (Grant No. ZDRW-ZS-2017-3).
Author information
Authors and Affiliations
Corresponding authors
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Chen, Y., Zhang, Y., Cao, L. et al. Wintering Swan Geese maximize energy intake through substrate foraging depth when feeding on buried Vallisneria natans tubers. Avian Res 10, 6 (2019). https://doi.org/10.1186/s40657-019-0145-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40657-019-0145-x