
Abstract
Background
Basic knowledge of detailed spatiotemporal migration patterns is lacking for most migratory bird species. Using the smallest available geolocator, we aim to map autumn migration and wintering areas of north European wood warblers Phylloscopus sibilatrix and compare the spatiotemporal pattern with recoveries of individuals ringed across Europe.
Results
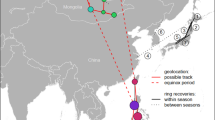
A tracked wood warbler migrated south-south-east to sub-Saharan Africa in Sudan and then west-south-west to winter in Côte d’Ivoire. The timing and route fits well within the distribution of ring recoveries although the westward movement after the Sahara crossing is not revealed by the ring recoveries, but only few recoveries south of Sahara exist.
Conclusions
The surprising westward movement south of the Sahara supplements the overall pattern revealed by ring recoveries and aids our understanding of the connectivity and site dependence in this generally declining species.
Similar content being viewed by others
Background
Our current understanding of the spatiotemporal migration systems in birds remains poor, especially for passerine species wintering in the tropics. This group is declining worldwide [1,2,3,4,5], and there is considerable interest in understanding the underlying causes, potentially to enhance conservation initiatives [6, 7].
Understanding causes of population declines are challenged by the lack of knowledge of migratory connectivity and spatiotemporal distribution across the annual cycle, including dependency on selected areas along the migration route. Despite increasing investment in tracking studies to reveal these patterns, individual migration routes remain elusive particularly for the smallest long-distance migratory birds.
The wood warbler Phylloscopus sibilatrix is one of the species where ring recoveries and observations so far provide only very limited information [8]. Ring recoveries indicate an initial southward migration direction with wintering areas found in the western and northern tropical forest belt of sub-Saharan Africa from Sierra Leone to western Kenya [9,10,11].
The wood warbler is strictly insectivorous and among the smallest Afro-Palearctic migrant species. It has a broad breeding range from across Europe and into Siberia [12]. Continent-wide declines by 35% have been reported [13]. In Denmark, wood warblers have declined by 2.5% per year from 1974 to 2016 [14].
The aim of this study is to map autumn migration and wintering areas of north European wood warblers Phylloscopus sibilatrix and compare the spatiotemporal pattern with recoveries of individuals ringed across Europe. We present a track retrieved from a geolocator recovered from a wood warbler, caught during the breeding season in Denmark, to identify its migration route, stopovers and wintering site.
Results
The tracked wood warbler departed from the Danish breeding site on 24 July 2014. From here, it migrated south (11 days) via a stopover in Italy (14 days) across the Sahara Desert (26 days) to a longer stopover (65 days) in Sudan. From here, it changed its migration direction to west-south-west with three shorter migration legs (each lasting 3–6 days) to stopovers in Chad (8 days), northern Nigeria (5 days) and Ghana (5 days) before arriving at its presumable main non-breeding area in Côte d’Ivoire (24 days before the battery power ended) (Fig. 1, Table 1). Arrival dates were 13 September in Sudan and 20 December in Côte d’Ivoire. Migration speed between stopovers varied considerably along the journey with the highest speed found on the migration stages westwards across tropical Africa (193-479 km/day) except the final stage before the main non-breeding arrival (67 km/day), while the Sahara crossing was relatively slow (120 km/day). The total distance covered was 8477 km in 52 migration days (mean = 163 km/day) and a total migration duration of 173 days (49 km/day including stopover days) (Fig. 1, Table 1).

Migration of the tracked wood warbler from Denmark to Côte d’Ivoire [circles indicate stopover sites, error bars are standard deviations (SD)] compared to ringing and recovery sites (connected with grey lines) of birds ringed during breeding and autumn migration (n = 28). All recoveries south of Sahara of birds ringed in Europe are indicated as dark grey triangles (n = 5). Inset indicates timing and number of days used on migration (grey) and stopover days for each migration stage (colours in the stacked bar diagram match the colour of the circle for geographical reference). Shaded area in Africa indicates the main known wintering area of wood warbler [9]. Black dashed lines (latitudinal movement including equinox) and solid lines (longitudinal movement) connecting circles give a visual representation of the route of the tracked wood warbler (for actual raw position data over time, see Additional file 1: Figure S1). The geolocator battery died 13 January 2015
Ring recoveries generally indicate a south to south-west migration (Fig. 2). Migration through Europe occurs in the months of August and September generally later compared to the timing of the tracked individual (Fig. 3). Birds within ± 1° of the tracked individual’s breeding location and southern European stopover were observed in August (median: 10 August, IQR: 23 July–16 August, n = 35) and September (median: 14 September, IQR: 2 September–19 September, n = 21), respectively. The westernmost ring recoveries on the Iberian Peninsula and the widely spread sub-Saharan recoveries suggest that the Sahara crossing occurs on broad front at the species level. The westward movement after the Sahara crossing in the tracked individual is not evident in the ring recoveries where the westernmost sub-Saharan locations are also the earliest.

Polar plot showing migration direction of ring recoveries of birds ringed and recovered on autumn migration in the same year within Europe (n = 13, grey) and the tracked wood warbler (black) from the breeding area to the southern European stopover site. Birds recovered dead were excluded based on EURING codes: “circumstance and condition”

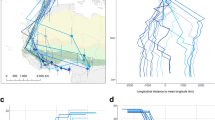
Latitude (left) and longitude (right) of stopover sites against time given with arrival and departure date for the wood warbler. Colours of stopovers correspond to colours in Fig. 1, while black lines indicate migration between sites. For comparison, we included latitude and longitude of all ringing data positions with ringing (n = 493, grey circles) and recovery date (n = 71, grey triangles) during breeding and autumn migration obtained from EURING (http://www.euring.org/edb). Dark grey triangles are sub-Saharan recoveries (n = 5). Grey lines are drawn between individuals ringed and recovered in the same year (n = 15). Birds recovered dead were excluded based on EURING codes: “circumstance and condition”
Discussion
The single tracked wood warbler migrated south-south-east to Sudan through Italy crossing the eastern part of the Sahara Desert. From Sudan, the bird moved slowly west to winter in Côte d’Ivoire. Through Europe, the spatiotemporal pattern was early, compared to that revealed by ring recoveries and the track revealed a surprising westward movement in sub-Saharan Africa during the season when only few recoveries exist. The main wintering site estimated in this study is within the overall non-breeding range of the species [9] and within the isoscape, but outside the predicted wintering distribution presented by Hobson et al. [10]. Migration speed was similar to some reports using ring recovery data comparing overall migration duration (e.g. 65 km/day in [15]).
The early migration through Europe compared to ringing data can perhaps be explained by a potentially more gradual migration pattern of wood warblers during autumn. There were some indications of shorter stopovers (< 5 days) within Europe in the longitudinal estimates of the tracked individual which could not be confirmed in latitudinal estimates due to the high variability caused by migration, equinox or shading in this part of the journey. Likewise, the main stopover in Europe may in fact have been two separate sites based solely on latitudinal estimates (Additional file 1: Figure S1).
The migration pattern had similarities to willow warbler [16]: the wood warbler stayed for 2 months immediately after crossing the Sahara Desert and before migration direction was changed towards west and migration restarted continuing for additionally 3373 km along areas with potentially suitable habitat [10]. The longer staging period after the Sahara crossing before reaching the final wintering site has also been found in other species, e.g. red-backed shrikes, great reed warblers and thrush nightingales [17,18,19], but not in redstarts [20] and some common nightingales stayed at the first site reached [21]. Even though the wood warbler is considered a habitat specialist, this study indicates that this individual may take advantage of the Sahel green-up during September–November [18] before continuing to the species-specific wintering areas in the continuous forest belt towards the south. The ring recoveries in sub-Saharan Africa did not support the westward movements. However, as there are only few sub-Saharan recoveries, the ring recoveries do not rule out that a significant proportion of birds could well be using the central route over Italy and dispersing westwards after the Sahara crossing.
Knowledge of links between breeding and non-breeding areas will help in the conservation of migratory songbirds. However, we also need to understand habitat requirements [22,23,24]. A number of recent studies have improved our knowledge of Phylloscopus warblers during breeding and non-breeding periods. Their preferred habitats in both seasons are broadleaved forests with a relatively open forest structure [16, 25,26,27]. A recent study showed no change in wood warbler abundance despite habitat conversion, indicating resilience of this species to loss of primary forest at the wintering grounds at least on the short term [28]. The combined knowledge of migration patterns (this study), breeding [26, 29] and non-breeding habitat requirements [28, 30] as well as other potential impacts at breeding sites [31] will form the basis for effective conservation strategies for declining migratory songbird species such as the wood warbler.
Because we could not track spring migration, it remains unknown whether the autumn route is traced back or a more direct route is chosen as indicated by more westerly recoveries in the Mediterranean in spring than autumn [32]. In addition, investigating the potential effect of improved vegetation conditions on timing of winter departure would be interesting as a general phenomenon.
While the tracked individual was an adult male, the ringing data included birds of both sexes and different age classes. Age and sex may influence the spatiotemporal migration patterns [33, 34]. Unfortunately, we were not able to account for age and sex in this study as most records are unknown.
The present study adds to our increasing understanding of the spatiotemporal distribution of more and more species of long-distance migratory songbirds across their annual cycle [35]. The next step will be to expand studies to determine the ecological requirements of migratory species throughout their annual cycle, e.g. during non-breeding and stopover for guiding future conservation initiatives. We can also study the link between breeding and non-breeding sites across different populations in space [36] and time [37] and build upon these new insights to start studying drivers of species-specific migration routes and schedules [18].
Conclusions
We present the spatiotemporal autumn migration of a wood warbler from breeding area to the main non-breeding area in sub-Saharan Africa. The migration of our tracked wood warbler fits well with the spatiotemporal distribution of ring recoveries, and we conclude that it is likely representative for wood warblers migrating from Denmark to sub-Saharan West Africa. The track also revealed a surprising westward movement in sub-Saharan Africa across a large part of the known non-breeding area, suggesting that the entire distribution of known ring recoveries can be covered by an individual during autumn migration. These findings provide the first insight into the migration strategy of this species and stopover locations with the potential for guiding future work connecting the entire annual circle for this declining songbird.
Methods
Using playback and mist nets, we trapped 17 male wood warblers in two different forests in Denmark; Tofte Skov (56°50′24″N, 10°14′24″E) and Gribskov (55°59′13″N, 12°18′30″E) during June 2014. Birds were ringed with a metal ring and fitted with archival light-level recorders (Intigeo–W30 from Migrate Technology Ltd, weight 0.3 g not including harness, expected recording time 7 months) using a leg-loop harness [38] made of 1-mm braided nylon string. Birds weighed 9.5–10.0 g, and thus, the logger with harness represented less than 3.3% of the body weight. On 12 June 2015, one individual was re-trapped in Tofte Skov. We did not recapture more tagged individuals although we performed forest surveys over a total of three weeks inspecting more than 200 singing males. The battery of the retrieved geolocator lasted until 13 January 2015 delivering data from the entire autumn migration and a large part of the wintering period.
We used the R package BAStag (version 0.1-3) to remove obvious false twilights (ten twilights removed, six sunsets and four sunrises over the months June–October) [39]. Data were log-transformed, and we used a threshold of 0.3 lux to identify twilight events. We used the R package GeoLight (version 2.0.0) to estimate coordinates from the light data [40]. Breeding area calibration resulted in a sun elevation angle of − 2.68. We removed data for 20 days on each side of autumn equinox (tolerance = 0.15) [41]. To estimate timing of stationary and movement periods, we visually inspected plots of longitude and latitude over time defining stopovers to be minimum 5 days (Additional file 1: Figure S1). During equinox, timing was estimated from longitudinal plots only. We used the R package geosphere version 1.5-7 [42] to estimate great circle distances between stopover sites. All analyses were done in R 3.5.1 [43].
With the aim of comparing our single autumn migration track with the current knowledge, we have compiled all European ring recoveries of wood warblers in the EURING database (www.euring.org/edb). We focus on autumn migration of birds ringed or recaptured during breeding and autumn migration for comparison with the geolocator bird. In addition, we added the few other sub-Saharan recoveries as they may indicate approximate wintering areas.
The ringing data obtained from EURING consisted of 11,492 ringing and re-encounter events. One event lacked spatial information and was removed. For graphical representation, we included only re-encounter events further than 200 km from the ringing site. Furthermore, for comparability with the individual geolocator track, we included only data where the bird had been ringed and subsequently re-encountered during breeding (June) or autumn migration (01 July–31 December) and individuals with breeding status. In addition, we included five records of birds re-encountered in sub-Saharan Africa during autumn migration (latitude < 20°N). Included records consisted of birds of both sexes and first year as well as full-grown birds, although in most cases the sex was unknown.
To compare the timing of autumn migration between the ringing data and the tracked individual, we estimated the median date and interquartile range (IQR) of birds ringed or recovered within ± 1° north and south of the tracked individual’s breeding area and southern European stopover site.
References
Sanderson FJ, Donald PF, Pain DJ, Burfield IJ, van Bommel FPJ. Long-term population declines in Afro-Palearctic migrant birds. Biol Conserv. 2006;131:93–105.
Berthold P, Fiedler W, Schlenker R, Querner U. 25-year study of the population development of central European songbirds: a general decline, most evident in long-distance migrants. Naturwissenschaften. 1998;85:350–3.
Holmes RT, Sherry TW. Thirty-year bird population trends in an unfragmented temperate deciduous forest: importance of habitat change. Auk. 2001;118:589–609.
Robbins CS, Sauer JR, Greenberg RS, Droege S. Population declines in North American birds that migrate to the neotropics. Proc Natl Acad Sci USA. 1989;86:7658–62.
Kirby JS, Stattersfield AJ, Butchart SHM, Evans MI, Grimmett RFA, Jones VR, et al. Key conservation issues for migratory land- and waterbird species on the world’s major flyways. Bird Conserv Int. 2008;18:S49–73.
Vickery JA, Ewing SR, Smith KW, Pain DJ, Bairlein F, Škorpilová J, et al. The decline of Afro-Palaearctic migrants and an assessment of potential causes. Ibis. 2014;156:1–22.
Ockendon N, Johnston A, Baillie SR. Rainfall on wintering grounds affects population change in many species of Afro-Palaearctic migrants. J Ornithol. 2014;155:905–17.
Dowsett RJ, Backhurst GC, Oatley TB. Afrotropical ring recoveries of Palearctic migrants. 1. Passerines (Turdidae to Oeiolidae). Tauraco. 1988;1:29–63.
BirdLife International. BirdLife International and handbook of the birds of the World Bird—species distribution maps of the world; 2016.
Hobson KA, Van Wilgenburg SL, Wesołowski T, Maziarz M, Bijlsma RG, Grendelmeier A, et al. A multi-isotope (δ2H, δ13C, δ15N) approach to establishing migratory connectivity in Palearctic-Afrotropical Migrants: an example using wood warblers Phylloscopus sibilatrix. Acta Ornithol. 2014;49:57–69.
Urban EK, Fry CH, Keith S. The birds of Africa, vol. V. San Diego: Academic Press; 1997.
Cramp S. Handbook of the birds of Europe, the Middle East and North Africa: the birds of the Western Palearctic. Tyrant flycatchers to thrushes, vol. V. Oxford: Oxford University Press; 1988.
EBCC. European Bird Census Council. 2017. http://ebcc.birdlife.cz/trends-of-common-birds-in-europe-2017-update/. Accessed 5 Nov 2018.
Fenger M, Nyegaard T, Jørgensen MF. Overvågning af de almindelige fuglearter i Danmark 1975–2015. Årsrapport for Punkttællingsprogrammet. Dansk Ornitologisk Forening; 2016.
Hall-Karlsson S, Fransson T. Svensk ringmärkningsatlas. Vol. 3, Tättingar = Passerines. SOF Forläg; 2008.
Lerche-Jørgensen M, Willemoes M, Tøttrup AP, Snell KRS, Thorup K. No apparent gain from continuing migration for more than 3000 kilometres: willow warblers breeding in Denmark winter across the entire northern Savannah as revealed by geolocators. Mov Ecol. 2017;5:1–7.
Tøttrup AP, Klaassen RHG, Kristensen MW, Strandberg R, Vardanis Y, Lindström Å, et al. Drought in Africa caused delayed arrival of European songbirds. Science. 2012;338:1307.
Thorup K, Tøttrup AP, Willemoes M, Klaassen RHG, Strandberg R, Vega ML, et al. Resource tracking within and across continents in long-distance bird migrants. Sci Adv. 2017;3:e1601360.
Koleček J, Hahn S, Emmenegger T, Procházka P. Intra-tropical movements as a beneficial strategy for Palearctic migratory birds. R Soc Open Sci. 2018;5:171675.
Kristensen MW, Tøttrup AP, Thorup K. Migration of the common redstart (Phoenicurus phoenicurus): a Eurasian songbird wintering in highly seasonal conditions in the West African Sahel. Auk. 2013;130:258–64.
Hahn S, Emmenegger T, Lisovski S, Amrhein V, Zehtindjiev P, Liechti F. Variable detours in long-distance migration across ecological barriers and their relation to habitat availability at ground. Ecol Evol. 2014;4:4150–60.
Willemoes M, Tøttrup AP, Lerche-Jørgensen M, Jacobsen EM, Reeve AH, Thorup K. Spatial behaviour and density of three species of long-distance migrants wintering in a disturbed and non-disturbed woodland in northern Ghana. Bird Conserv Int. 2017;28:59–72.
Zwarts L, Bijlsma RG, van der Kamp J, Sikkema M, Wymenga E. Moreau’s paradox reversed, or why insectivorous birds reach high densities in Savanna trees. Ardea. 2015;103:123–44.
Morel GGJ, Morel M-YM. Habitat use by Palaearctic migrant passerine birds in West Africa. Ibis. 1992;134:83–8.
Stostad HN, Menéndez R. Woodland structure, rather than tree identity, determines the breeding habitat of Willow Warblers Phylloscopus trochilus in the northwest of England. Bird Study. 2014;61:246–54.
Mallord JW, Charman EC, Cristinacce A, Orsman CJ. Habitat associations of Wood Warblers Phylloscopus sibilatrix breeding in Welsh oakwoods. Bird Study. 2012;59:403–15.
Pasinelli G, Grendelmeier A, Gerber M, Arlettaz R. Rodent-avoidance, topography and forest structure shape territory selection of a forest bird. BMC Ecol. 2016;16:1–18.
Mallord JW, Orsman CJ, Roberts JT, Boafo K, Skeen RQ, Sheehan DK, et al. Apparent resilience of a declining Afro-Palaearctic migrant to forest loss on the wintering grounds. Int J Avian Sci. 2018;160:805–15.
Huber N, Kéry M, Pasinelli G. Occupancy dynamics of the Wood Warbler Phylloscopus sibilatrix assessed with habitat and remote sensing data. Ibis. 2017;159:623–37.
Mallord JW, Orsman CJ, Roberts JT, Skeen R, Sheehan DK, Vickery JA. Habitat use and tree selection of a declining Afro-Palaearctic migrant at sub-Saharan staging and wintering sites. Bird Study. 2016;63:459–69.
Szymkowiak J, Thomson RL, Kuczyński L. Wood warblers copy settlement decisions of poor quality conspecifics: support for the tradeoff between the benefit of social information use and competition avoidance. Oikos. 2016;125:1561–9.
Bønløkke J, Madsen JJ, Thorup K, Petersen KT, Bjerrum M, Rahbek C. The Danish Bird Migration Atlas. Humlebæk: Rhodos; 2006.
Schmaljohann H, Meier C, Arlt D, Bairlein F, van Oosten H, Morbey YE, et al. Proximate causes of avian protandry differ between subspecies with contrasting migration challenges. Behav Ecol. 2016;27:321–31.
Smith RJ, Moore FR. Arrival timing and seasonal reproductive performance in a long-distance migratory landbird. Behav Ecol Sociobiol. 2005;57:231–9.
McKinnon EA, Love OP. Ten years tracking the migrations of small landbirds: lessons learned in the golden age of bio-logging. Auk. 2018;135:834–56.
Webster MS, Marra PP, Haig SM, Bensch S, Holmes RT. Links between worlds: unraveling migratory connectivity. Trends Ecol Evol. 2002;17:76–83.
Bauer S, Lisovski S, Hahn S. Timing is crucial for consequences of migratory connectivity. Oikos. 2016;125:605–12.
Naef-Daenzer B. An allometric function to fit leg-loop harnesses to terrestrial birds. J Avian Biol. 2007;38:404–7.
Wotherspoon S, Sumner M, Lisovski S. BAStag: basic data processing for British Antarctic Survey archival tags. 2016. https://github.com/SWotherspoon/BAStag/. Accessed 5 Nov 2018.
Lisovski S, Hahn S. GeoLight—processing and analysing light-based geolocator data in R. Methods Ecol Evol. 2012;3:1055–9.
Lisovski S, Hewson CM, Klaassen RHG, Korner-Nievergelt F, Kristensen MW, Hahn S. Geolocation by light: accuracy and precision affected by environmental factors. Methods Ecol Evol. 2012;3:603–12.
Hijmans RJ. Geosphere: spherical trigonometry. 2016. https://cran.r-project.org/web/packages/geosphere/index.html/. Accessed 5 Nov 2018.
R Core Team. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing; 2018. http://www.r-project.org/. Accessed 5 Nov 2018.
Tøttrup AP, Pedersen L, Thorup K. Data from: Autumn migration and wintering site of a wood warbler Phylloscopus sibilatrix breeding in Denmark identified using geolocation. Movebank Data Repository; 2018. https://doi.org/10.5441/001/1.5jp8mq2v
Fair J, Paul E, Jones J. Guidelines to the use of wild birds in research. Washington: Ornithological Council; 2010.
Authors’ contributions
APT, LP, KT carried out field work. APT, LP, KT analysed the data. APT, LP, KT drafted the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We thank Troels Eske Ortvad for fieldwork assistance and two anonymous reviewers for suggestions improving the manuscript.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The tracking data used in this study are available on Movebank (movebank.org, study name: “Autumn migration of wood warbler”) and are published in the Movebank Data Repository [44]. Ringing data are from the EURING database (http://www.euring.org/edb).
Ethics approval and consent to participate
This study was carried out in strict accordance with Guidelines to the use of wild birds in research of the Ornithological Council [45]. The Copenhagen Bird Ringing Centre with permission from the Danish Nature Agency approved capturing and tagging of wood warblers.
Funding
APT thanks Aage V. Jensen Naturfond for support. KT thanks The Danish Council for Independent Research for support to the MATCH Project (1323-00048B). KT, LP, APT thank the Danish National Research Foundation for support to the Center for Macroecology, Evolution and Climate.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Additional file
Additional file 1: Figure S1.
Observed changes in estimated longitude (left) and latitude (right) over time from deployment of the geolocator on a wood warbler in June 2014 until the battery died 13 January 2015. Colours denote stopover sites and migratory segments following Fig. 1 in the main paper (black: active migration, pink: breeding site, orange: southern Europe stopover, light blue: Sudan stopover, green: Chad stopover, yellow: Nigeria stopover, dark blue: Ghana stopover, red: Côte d’Ivoire presumed wintering area). Dashed vertical line indicate equinox.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Tøttrup, A.P., Pedersen, L. & Thorup, K. Autumn migration and wintering site of a wood warbler Phylloscopus sibilatrix breeding in Denmark identified using geolocation. Anim Biotelemetry 6, 15 (2018). https://doi.org/10.1186/s40317-018-0159-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40317-018-0159-x