
Abstract
Background
The main symptoms of schistosomiasis are granuloma and fibrosis, caused by Schistosoma eggs. Numerous types of cells and cytokines are involved in the progression of Schistosoma infection. As a class of innate immune cells, γδ T cells play critical roles in the early immune response. However, their role in modulating granuloma and fibrosis remains to be clarified.
Methods
Liver fibrosis in wild-type (WT) mice and T cell receptor (TCR) δ knockout (KO) mice infected with Schistosoma japonicum was examined via Masson’s trichrome staining of collagen deposition and quantitative reverse transcriptase-PCR (RT-PCR) of fibrosis-related genes. Granuloma was detected by hematoxylin-eosin (H&E) staining and quantified. Flow cytometry was used for immune cell profiling and for detecting cytokine secretion. The abundance of the related cytokines was measured using quantitative RT-PCR.
Results
The livers of S. japonicum-infected mice had significantly increased proportions of interleukin (IL)-17A producing γδ T cells and secreted IL-17A. Compared with the WT mice, TCR δ deficiency resulted in reduced pathological impairment and fibrosis in the liver and increased survival in infected mice. In addition, the profibrogenic effects of γδ T cells in infected mice were associated with enhanced CD11b+Gr-1+ cells, concurrent with increased expression of transforming growth factor (TGF)-β in the liver.
Conclusions
In this mouse model of Schistosoma infection, γδ T cells may promote liver fibrosis by recruiting CD11b+Gr-1+ cells. These findings shed new light on the pathogenesis of liver pathology in murine schistosomiasis.

Similar content being viewed by others


Background
Schistosomiasis is a neglected tropical disease caused by the parasites of the genus Schistosoma. Infection is prevalent in over 74 countries, and nearly 200 million people are at risk [1,2,3]. Following infection, liver fibrosis is the major pathological manifestation of the disease. This is mainly due to the deposition of Schistosoma eggs in the host liver. The excretory secretory antigens produced by the eggs induce strong immune responses and inflammation, and trigger granuloma formation around the eggs, which in turn leads to the activation of hepatic stellate cells (HSCs) and consequent liver fibrosis [4].
The mechanisms of fibrosis have been studied extensively in the murine model of schistosomiasis. One major mechanism is the switching of the immune pattern from a predominant T helper cell 1 (Th1) response to a Th2-dominant response, which promotes granuloma and fibrosis formation by secreting cytokines such as interleukin (IL)-4, IL-5 and IL-13 [5,6,7,8,9]. As a member of the IL-17 family, IL-17A is involved in various biological processes by binding its cognate receptor, IL-17RA [10,11,12,13]. Accumulating evidence demonstrates that IL-17A is related to a variety of inflammatory and autoimmune diseases such as inflammatory bowel diseases (IBD) and multiple sclerosis [14,15,16]. Multiple studies have demonstrated that Th17 cells, natural killer T cells and γδ T cells produce IL-17A [17,18,19]. Th17 promotes hepatic granuloma formation by secreting IL-17A [20,21,22]. Moreover, the absence of IL-17A signaling has been implicated in alleviating liver fibrosis in murine schistosomiasis [23].
Multiple cell types, including macrophages, T follicular helper cells (Tfh), regulatory T cells (Treg), and dendritic cells (DC) are activated and involved in granuloma formation and in the process of fibrosis [24,25,26,27,28]. γδ T cells are a subgroup of T cells that express a distinct T cell receptor (TCR) consisting of a γ and a δ chain, which allows rapid response to antigens without the requirement of major histocompatibility complex (MHC) presentation [29, 30]. γδ T cells play pivotal roles in autoimmunity, inflammatory diseases, and tumor development [31,32,33,34]. They also have important regulatory functions [35]. Of note, they are an important regulator in liver diseases, including liver infection, non-alcoholic fatty liver disease, autoimmune hepatitis, and liver fibrosis [36]. Although a correlation between γδ T cell and neutrophil recruitment and liver fibrosis has been implicated [37], the exact mechanism underlying the regulatory role of γδ T cells in liver fibrosis remains elusive. In the present study, we aimed to characterize the role of γδ T cells in the early formation of granuloma and fibrosis in the livers of TCR δ knockout (KO) mice infected with Schistosoma japonicum. We provide evidence that deleting TCR δ markedly ameliorates liver fibrosis and improves liver function. We explored the mechanism by which γδ T cells promote liver pathology in murine schistosomiasis. Our findings indicate that γδ T cells may promote liver fibrosis by recruiting CD11b+Gr-1+ cells, which thereby furthers understanding of fibrosis formation and provides a strategy for the basis of anti-schistosomiasis therapy.
Methods
Mice, parasite and infection
Eight-week-old male C57BL/6 mice were purchased from Shanghai Jiesijie Animal Feeding Co. Ltd. (Shanghai, China). TCR δ KO mice on C57BL/6 background were purchased from Jackson Laboratory (Bar Harbor, USA) and bred in our facility. For the comparison experiment, wild-type (WT) and KO mice were bred under specific pathogen-free conditions. In the infection experiments, 31 WT mice and 31 KO mice were each infected percutaneously via the shaved skin of the abdomen, with 20 ± 2 S. japonicum cercariae obtained from the National Institute of Parasitic Diseases, Chinese Center for Disease Control and Prevention (Shanghai, China). Fifteen WT mice and 15 TCR δ KO mice were randomly divided into three groups, and were sacrificed for pathological observation at 0, 4 and 6 weeks post-infection, respectively. Sixteen WT mice and 16 KO mice were used for the survival time observation, and the survival rates in each group were evaluated.
Liver pathology and measurement of aspartate aminotransferase/alanine aminotransferase (AST/ALT) levels
Fresh liver samples were fixed in 10% neutral buffered formalin, then processed with routine paraffin-embedding procedures. The paraffin-embedded sections (5 μm) were dewaxed and stained with hematoxylin-eosin (H&E) or Masson’s trichrome for analyzing granuloma or fibrosis, respectively. Granulomas around single eggs were selected to assess the levels of granuloma and fibrosis. Each stained section was examined by optical microscopy under 100× magnification with identical settings. Thirty pictures were taken of granulomas around single eggs from three sections in each tissue. The areas featuring granuloma and fibrosis surrounding a single egg were estimated using ImageJ (NIH, Bethesda, MD, USA). Sera from individual infected mice were separated by centrifugation at 1000× g for 10 min. ALT and AST levels were detected by Adicon Clinical Laboratories, Inc. (Shanghai, China).
Isolation of liver non-parenchymal cells and spleen cells
Single-cell suspension of hepatic leukocytes were prepared using a conventional method according to a previous report with some modifications [17]. Briefly, infected or control mice were perfused with phosphate-buffered saline (PBS) via the portal vein. The excised liver was minced, then pressed through a 70-μm cell strainer (BD Biosciences, San Jose, CA, USA). The total hepatic cells were then resuspended in 40% Percoll solution (GE Healthcare, Chicago, IL, USA), and centrifuged for 15 min at 750× g. Then, the leukocytes were resuspended in erythrocyte-lysing buffer. Lastly, the cells were washed and resuspended in RPMI 1640 medium. For the spleen, all cells were washed and resuspended in RPMI 1640 medium immediately after they had been pressed through a 70-μm cell strainer (BD Biosciences).
Flow cytometry
To assess cytokine and biological marker expression on γδ T cells and CD11b+Gr-1+ cells, 2 × 106 cells per 100 μl were incubated with the following fluorescence-labeled antibodies for 30 min at 4 °C; fluorescein isothiocyanate (FITC) labeled anti-CD3; phycoerythrin (PE)-Cy7-labeled anti-γδ TCR; allophycocyanin (APC)-labeled anti-Vγ1; PE-labeled anti-Vγ2; Brilliant Violet 421-labeled anti-CD11b; APC-labeled anti-Gr-1; and PE-labeled anti-IL17RA (BioLegend, San Diego, CA, USA). After surface marker staining, the cells were permeabilized with Fix/Perm buffer (BD Biosciences) for 15 min, then incubated with the following fluorescence-labeled antibodies: Brilliant Violet 421-labeled anti-IL17A; Brilliant Violet 421-labeled anti-interferon (IFN)-γ; and PE-labeled anti-transforming growth factor (TGF)-β. The staining panels were as follows: γδ T cell, FITC-labeled anti-CD3, PE-Cy7-labeled anti-γδ TCR, APC-labeled anti-Vγ1, PE-labeled anti-Vγ2, Brilliant Violet 421-labeled anti-IL17A/Brilliant Violet 421-labeled anti-IFN-γ; CD11b+Gr-1+ cells, Brilliant Violet 421-labeled anti-CD11b, APC-labeled anti-Gr-1, PE-labeled anti-TGF-β/PE-labeled anti-IL17RA.
All flow cytometry data were acquired on an LSRFortessa X-20 cell analyzer (BD Biosciences) and analyzed with FlowJo software (Tree Star, Ashland, OR, USA).
Quantitative reverse transcriptase PCR (qRT-PCR)
The total RNA was extracted from liver tissue with TRIzol (Invitrogen, Carlsbad, CA, USA). For each sample, 2 μg of total RNA was reverse-transcribed using a complementary DNA (cDNA) reverse transcription kit (TaKaRa, Dalian, China). The comparative threshold cycle (2−ΔΔCq) was calculated to evaluate the relative mRNA expression, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) levels were used as a normalization control. The qRT-PCR primers used in this study are listed in Table 1.
Statistical analysis
All quantitative data were reported as the mean and standard error of the mean (SE). All samples were compared using an unpaired Student’s t-test or one-way analysis of variance (ANOVA). The survival rate was analysed by using the Kaplan–Meier method, the difference between survival curves was tested for statistical significance using the log-rank test. P < 0.05 was considered to indicate statistical significance. GraphPad Prism 6 (GraphPad Software, San Diego, CA, USA) was used for all statistical analysis and graphs.
Results
The liver γδ T cells of infected mice had significantly increased IL-17A secretion
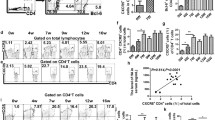
To profile the dynamic changes of the γδ T cells, we performed fluorescence-activated cell sorting and quantified the ratio of γδ T cells in the liver of S. japonicum-infected mice. Additional file 1: Figure S1 shows the gating strategy. The γδ T cells constituted approximately 2%, 3% and 2.8% of the total CD3+ T cells in the liver at 0, 4 and 6 weeks post-infection, respectively (Fig. 1a-c). During infection, the proportion of γδ T cells did not change greatly (F(2, 12) = 4.657, P = 0.0318) (Fig. 1d). However, the absolute number of γδ T cells was approximately 4.0 × 104, 7.5 × 104 and 10.0 × 104 at 0, 4 and 6 weeks post-infection, respectively, which was a significant increase (F(2, 12) = 100.7, P < 0.0001) (Fig. 1e). Notably, γδ T cell subtype analysis revealed mainly two subtypes: Vγ1 and Vγ2 (Fig. 1f-h). The percentage of Vγ1 cells peaked at 4 weeks post-infection (F(2, 12) = 49.46, P < 0.0001) (Fig. 1i), whereas that of the Vγ2 cells increased gradually post-infection (F(2, 12) = 11.98, P = 0.0014) (Fig. 1j). We then detected the cytokines secreted by the two subtypes. Additional file 2: Figure S2a-b shows the isotype controls. Vγ1 cells mainly secreted IFN-γ (Fig. 2a-c) rather than IL-17A (F(2, 12) = 7.481, P = 0.0078) (Fig. 2e-h), while Vγ2 cells secreted both IFN-γ (Fig. 2i-k) and IL-17A (Fig. 2m-o). IFN-γ from the Vγ1 cells increased at 4 weeks post-infection, and then decreased at 6 weeks post-infection (F(2, 12) = 44.56, P < 0.0001) (Fig. 2d), as did IFN-γ from the Vγ2 cells (F(2, 12) = 23.84, P < 0.0001) (Fig. 2l). However, IL-17A from the Vγ2 cells increased during infection (F(2, 12) = 31.37, P < 0.0001) (Fig. 2p). These results suggest that γδ T cells may participate in the liver pathological process in mice infected with S. japonicum.

Percentage of γδ T cells and their subsets in S. japonicum-infected mouse liver. a-c Flow cytometry analysis of percentage of γδ T cells in total CD3+ cells from liver leukocytes, and the representative dot plots at 0 (a), 4 (b) and 6 (c) weeks post-infection. d, e Statistical analysis of the percentage (d) and absolute number (e) of γδ T cells in CD3+ cells from liver leukocytes. f-h Flow cytometry analysis of Vγ1 and Vγ2 subset percentages among γδ T cells in total CD3+ T cells from liver leukocytes at 0 (f), 4 (g) and 6 (h) weeks post-infection, and the representative dot plots. i-j Statistical analysis of the percentage of Vγ1 (i) and Vγ2 (j) cells in total CD3+ T cells from liver leukocytes. The differences at 0, 4 and 6 weeks were analyzed using a one-way analysis of variance (ANOVA), n = 5 per group, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; ns, no significant difference

Vγ1 and Vγ2 cells secrete IFN-γ and IL-17A, respectively, in the liver. a-d Flow cytometric and statistical analyses of the percentage of IFN-γ from Vγ1 cells at 0 (a), 4 (b) and 6 (c) weeks post-infection. e-h Flow cytometric and statistical analyses of the percentage of IL-17A from Vγ1 cells at 0 (e), 4 (f) and 6 (g) weeks post-infection. i-l Flow cytometric and statistical analyses of the percentage of IFN-γ from Vγ2 cells at 0 (i), 4 (j) and 6 (k) weeks post-infection. m-p Flow cytometric and statistical analyses of the percentage of IL-17A from Vγ2 cells at 0 (m), 4 (n) and 6 (o) weeks post-infection. Liver leukocytes were stimulated with PMA (phorbol 12-myristate 13-acetate) +ionomycin for 4 h; intracellular IL-17A and IFN-γ staining were performed and representative dot plots are shown. The differences at 0, 4 and 6 weeks were analyzed using a one-way ANOVA, n = 5 per group, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; ns, no significant difference
Infected TCR δ KO mice had alleviated pathology and liver damage
To evaluate whether γδ T cells were involved in the pathogenesis of S. japonicum infection in mice, WT and TCR δ KO mice were infected with S. japonicum, and were sacrificed at 6 weeks post-infection. Hepatic granuloma size was determined using H&E staining (Fig. 3a, b). The average granuloma area in the KO mouse livers was much less than that of the WT mice (t(58) = 5.248, P < 0.0001) (Fig. 3c). Collagen deposition was detected using Masson’s trichrome staining (Fig. 3d, e), the KO mouse livers had significantly less severe fibrosis than that of the WT mice (t(58) = 6.481, P < 0.0001) (Fig. 3f). We also determined liver function (ALT/AST) at 0, 4 and 6 weeks after infection. The KO mice showed improved liver function in comparison with the WT mice at 4 and 6 weeks post-infection (t(8) = 2.690, P = 0.0275, t(8) = 2.982, P = 0.0175; and t(8) = 3.626, P = 0.0067, t(8) = 4.691, P = 0.0016, respectively) (Fig. 3g, h). Further, we monitored the survival rates of the mice. From week 11 onwards, the KO mice had significantly higher survival rates than the WT mice (P = 0.0301) (Additional file 3: Figure S3). Taken together, these data suggest that γδ T cell deficiency protects S. japonicum-infected mice from liver fibrosis and damage caused by Schistosoma eggs.

Decreased fibrosis and liver damage in S. japonicum-infected TCR δ KO mice. a, b H&E staining showing liver granuloma in WT (a) and TCR δ KO (b) mice at 6 weeks post-infection. c Statistical analysis of the granuloma area between the two groups. d, e Masson staining showing the liver fibrosis in WT (d) and KO (e) mice at 6 weeks post-infection. f Statistical analysis of the fibrosis area between the two groups. g, h Statistical analysis of serum ALT (g) and AST (h) in WT and KO mice at 0, 4 and 6 weeks post-infection. Differences were analyzed by using a Student’s t-test, n = 5 per group *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; ns, no significant difference
Reduced fibrosis is associated with decreased TGF-β and IL-17A in infected TCR δ KO mice
To study which factor is related to reduced fibrosis in S. japonicum-infected TCR δ KO mice, we extracted RNA from the liver tissue of the WT and KO mice at 6 weeks post-infection, and measured the mRNA transcripts of collagen, TGF-β, IL-17A and IL-13 using qRT-PCR. TCR δ deletion led to reduced expression of collagen I, TGF-β and IL-17A (t(4) = 3.296, P = 0.0301, t(4) = 3.218, P = 0.0324 and t(4) = 5.966, P = 0.004, respectively) (Fig. 4a-c), but not IL-13 (Fig. 4d). To rule out the difference between the number of Schistosoma adults and the Schistosoma egg burden, we examined the total number of adults (Fig. 4e) and egg burden (Fig. 4f) in each mouse at 6 weeks post-infection. There was no significant difference between the findings for the WT and KO mice. The involvement of IL-17A and TGF-β was implicated in the fibrotic effects in the S. japonicum infection [28, 38, 39]. Taken together, our results indicate the involvement of TGF-β in alleviating the fibrosis in TCR δ KO mice infected with S. japonicum.

The fibrogenic genes were downregulated in S. japonicum–infected TCR δ KO mice. a-d Real-time RT-PCR measurement of mRNA transcripts of collagen I (a), TGF-β (b), IL-17A (c), and IL-13 (d) in WT and KO mice at 6 weeks post-infection. e Adult worms in WT and KO mice at 6 weeks post-infection. f Hepatic egg burden in WT and KO mice at 6 weeks post-infection. Differences were analyzed by using Student’s t-test, n = 3 or n = 5 per group, *P < 0.05, **P < 0.01. Abbreviations: ns, no significant difference; EPG, egg per gram
The profibrotic effects of γδ T cells were associated with CD11b+Gr-1+ cells
In the WT mice, CD11b+Gr-1+ cells increased rapidly after infection (data not shown). To explore the relationship between γδ T cells and CD11b+Gr-1+ cells, we examined the percentage of CD11b+Gr-1+ cells in the liver and spleen from the WT and TCR δ KO mice at 0, 4 and 6 weeks post-infection. At 0 weeks post-infection, the WT and KO mouse livers had only about 1.4% and 1.3% CD11b+Gr-1+ cells, respectively (Fig. 5a, d), and there was no significant difference between the two groups (Fig. 5g). At 4 weeks post-infection, the percentage of CD11b+Gr-1+ cells in the WT and KO mouse livers increased to about 10.9% and 11.2%, respectively (Fig. 5b, e). At 6 weeks post-infection, the WT mouse livers had about 33% CD11b+Gr-1+ cells (Fig. 5c), while that in the KO mouse livers was about 20% (Fig. 5f), representing a significant reduction (t(8) = 7.028, P = 0.0001) (Fig. 5g). At 0 weeks post-infection, the WT and KO mouse spleens had only about 0.6% and 1.2% CD11b+Gr-1+ cells, respectively (Fig. 5h, k); there was no significant difference between the two groups (Fig. 5n). At 4 weeks post-infection, the percentage CD11b+Gr-1+ cells in the WT mouse spleens increased to about 3.5% (Fig. 5i), and that in the KO mice was about 1.6% (Fig. 5l); there was a significant difference between the two groups (t(8) = 7.212, P < 0.0001) (Fig. 5n). At 6 weeks post-infection, the percentage of CD11b+Gr-1+ cells in the WT mouse spleens continued to increase to around 15% (Fig. 5j). The percentage of CD11b+Gr-1+ cells in the KO mouse spleens decreased to around 7% (Fig. 5m), which was significantly lower than of wild-type mice (t(8) = 5.748, P = 0.0004) (Fig. 5n). These data suggest that the fibrotic effects of γδ T cells in S. japonicum infection may be related to CD11b+Gr-1+ cells.

The profibrogenic effects of γδ T cells in infected mice are associated with CD11b+Gr-1+ cells. a-c The percentage of CD11b+Gr-1+ cells in WT mouse liver at 0 (a), 4 (b) and 6 (c) weeks post-infection, and representative dot plots. d-f The percentage of CD11b+Gr-1+ cells in KO mouse liver leukocytes at 0 (d), 4 (e) and 6 (f) weeks post-infection, and representative dot plots. g Statistical analysis of the percentage of CD11b+Gr-1+ cells in WT and mouse liver leukocytes. h-j The percentage of CD11b+Gr-1+ cells in WT mouse spleen at 0 (h), 4 (i) and 6 (j) weeks post-infection, and representative dot plots. k-m The percentage of CD11b+Gr-1+ cells in KO mouse spleen at 0 (k), 4 (l) and 6 (m) weeks post-infection, and representative dot plots. n Statistical analysis of the percentage of CD11b+Gr-1+ cells in WT and KO mouse spleen. Differences were analyzed by using Student’s t-test, n = 5 per group, ***P < 0.001; ns, no significant difference
The possible mechanisms of γδ cell promotion of liver fibrosis
We interrogated the mechanism underlying the contribution of CD11b+Gr-1+ cells in mediating the fibrotic effects of γδ T cells. First, we examined the relationship between γδ cells and CD11b+Gr-1+ cells. Flow cytometric analysis showed that liver CD11b+Gr-1+ cells also expressed IL-17RA at 4 and 6 weeks post-infection (Fig. 6a, b). The same result was found for the spleen CD11b+Gr-1+ cells (Fig. 6c, d). As a receptor of IL-17A, IL-17RA may promote CD11b+Gr-1+ cell recruitment from the liver and spleen. In studying how CD11b+Gr-1+ cells promote fibrosis, we observed that the liver CD11b+Gr-1+ cells (WT and KO mice, about 35% and 20%, respectively) expressed TGF-β (Fig. 6e, f) (Additional file 2: Figure S2c shows the isotype). There was a significant difference between the two groups (t(8) = 3.974, P = 0.0041) (Fig. 6g). These results suggest that IL-17A secreted by γδ T cells may directly contribute to the recruitment of CD11b+Gr-1+ cells and that CD11b+Gr-1+ cells may be associated with liver fibrosis via the expression of TGF-β.

The possible mechanisms of γδ cell promotion of liver fibrosis. a, b CD11b+Gr-1+ cells in WT mouse liver expressed IL-17RA at 4 (a) and 6 (b) weeks post-infection. c, d CD11b+Gr-1+ cells in WT mouse spleen expressed IL-17RA at 4 (c) and 6 (d) weeks post-infection. e CD11b+Gr-1+ cells in WT mouse liver expressed TGF-β at 6 weeks post-infection. f CD11b+Gr-1+ cells in KO mouse liver expressed TGF-β at 6 weeks post-infection. g Statistical analysis of the percentage of TGF-β in CD11b+Gr-1+ cells from WT and KO mouse liver. Liver leukocytes were stimulated with PMA + ionomycin for 4 h; intracellular TGF-β staining was performed, and representative dot plots are shown. Student’s t-test was applied to two groups at six weeks, n = 5 per group, **P < 0.01
Discussion
Although the formation of granuloma and fibrosis limits the viability of Schistosoma eggs to a certain extent and helps improve the host survival rate, but the subsequent liver pathology is nonetheless very severe, and even leads to death. Therefore, more rational treatment of schistosomiasis requires more comprehensive understanding of granuloma and fibrosis formation. In the present study, we explored whether γδ T cells play a role in granuloma formation in the early stages of S. japonicum infection. Our results indicate that IL-17A-producing γδ T cells are involved in the processes of granuloma and fibrosis, although the abundance of the cells was relatively low. Moreover, the IL-17A-producing γδ T cells promoted the increase in CD11b+Gr-1+ cells. Further, we explored how γδ T cells promote CD11b+Gr-1+ cells. Our results suggest that γδ T cells may secrete IL-17A and mediate CD11b+Gr-1+ cell recruitment by engaging IL-17RA expressed on the CD11b+Gr-1+ cell surface. Next, we explored the possible mechanisms of fibrosis by CD11b+Gr-1+ cells. Our findings show that CD11b+Gr-1+ cells contribute to fibrogenesis by the high production of TGF-β.
γδ T cells are typically found at surface of barrier tissue such as the mucosa, lungs and skin. As innate T cells, γδ T cells are non-abundant; however, they play an important role in diverse diseases. For example, TCR-deficient mice are more susceptible to infection by bacteria and viruses, and have significantly increased incidence of tumors. γδ cells are well-documented as being involved in liver disease, such as non-alcoholic fatty liver disease, autoimmune hepatitis, liver fibrosis and cirrhosis and liver cancer [40,41,42,43]. Our results indicate that γδ T cells constitute approximately 3% of the total CD3+ T cells, and consist of two major subsets, which is consistent with previous reports [37, 44, 45]. In some disease models, Vγ1 cells, which are IFN-γ-producing γδ T cells, play an essential role in liver infection and liver cancer [46, 47]. In the present study, IFN-γ secretion by Vγ1 cells began to increase at 0 and 4 weeks post-infection, but then decreased at 6 weeks post-infection. Vγ2 cells, whose most striking feature is the secretion of IL-17A, are involved in immune responses and various diseases. They not only play protective roles against colitis and viruses, but also promote liver fibrosis [16, 48], especially, γδ T cells could exacerbate liver fibrosis by aiding the activation of HSCs in liver fibrosis induced by carbon tetrachloride injection [49]. Our observations show that during infection, the increase in Vγ2 cells was accompanied by an increase in their secreted IL-17A. These results suggest that IL-17A-producing γδ T cells (i.e. Vγ2 cells) may participate in liver pathological processes in mice infected with S. japonicum.
In the present study, we used TCR δ KO mice to confirm that γδ T cells are involved in liver fibrosis in S. japonicum-infected mice. Our results show that the KO mice had reduced granuloma area and attenuated liver fibrosis as compared with the WT mice. We selected granuloma caused by single eggs to ensure that the pathological improvement was due to the host immune response rather than the number of eggs, and this is consistent with previous reports [24, 50]. The improvement of liver function in the KO mice may have enhanced their survival rate as compared with the WT mice.
TGF-β and IL-13 are well-documented as two important cytokines of liver fibrosis in mice infected with S. mansoni and S. japonicum, and IL-13 is the key driving liver fibrosis [38, 39, 51, 52]. Here, TGF-β, and not IL-13, was decreased in the KO mouse livers at 6 weeks post-infection; IL-17A was also decreased in the KO mouse livers. These data suggest that the ameliorated liver fibrosis caused by the lower TGF-β was accompanied by decreased IL-17A expression.
CD11b+Gr-1+ cells are involved in diverse disease such as IBD, tumor, and liver fibrosis [53,54,55]. CD11b+Gr-1+ cells and γδ T cells are reciprocally regulated [16, 56]. Our data show that CD11b+Gr-1+ cells in the WT mice increased rapidly during infection. However, CD11b+Gr-1+ cells in the KO mice were decreased in comparison. These data indicate that γδ T cells may promote liver fibrosis by recruiting CD11b+Gr-1+ cells. The notable characteristic of γδ T cells is the expression of IL-17A, and our data show that CD11b+Gr-1+ cells can express IL-17RA. IL-17RA or the IL-17RA/IL-17RC heterodimer is considered the receptor of IL-17A [57], so we speculate that γδ T cells promote CD11b+Gr-1+ cells via an IL-17A–IL-17RA pathway. Compared with WT mice, liver fibrosis is decreased in IL-17RA-deficient mice [14]. In addition, IL-17RA deficiency can prevent the development of tubulointerstitial fibrosis in the kidney [58]. In two different models of airway fibrosis, IL-17RA-deficient mice were protected against both airway inflammation and fibrosis [59]. Finally, our data imply that CD11b+Gr-1+ cells facilitate the development of liver fibrosis by expressing TGF-β. Similarly, CD11b+Gr-1+ cells can promote silica-induced lung fibrosis in mice [60]. Taken together, we speculate that IL-17A may recruit CD11b+Gr-1+ cells and promote liver fibrosis by expressing TGF-β. However, we should point out that more studies and evidence are needed to reveal and clarify the integrated mechanism involved.
Conclusions
We assessed the role of γδ T cells in liver fibrosis caused by acute murine schistosomiasis. We confirm that γδ T cells are involved in the development of liver fibrosis.
Availability of data and materials
The datasets supporting the conclusions of this article are included within the article and its additional files.
Abbreviations
- DC:
-
dendritic cell
- Tfh cells:
-
T follicular helper cells
- Treg:
-
regulatory T cells
- MHC:
-
major histocompatibility complex
- ALT:
-
aspartate aminotransferase
- AST:
-
aspartate aminotransferase, TGF-β: transforming growth factor β
- IFN-γ:
-
interferon-gamma
- IL-13:
-
interleukin-13
- IL-17A:
-
interleukin-17A
- COL 1A:
-
collagen 1A
References
Ross AG, Bartley PB, Sleigh AC, Olds GR, Li Y, Williams GM, et al. Schistosomiasis. N Engl J Med. 2002;346:1212–20.
Gryseels B, Polman K, Clerinx J. Kestens. Human schistosomiasis. Lancet. 2006;368:1106–18.
Gryseels B. Schistosomiasis. Infect Dis Clin North Am. 2012;26:383–97.
Pearce EJ, MacDonald AS. The immunobiology of schistosomiasis. Nat Rev Immunol. 2002;2:499–511.
Chiaramonte MG, Donaldson DD, Cheever AW, Wynn TA. An IL-13 inhibitor blocks the development of hepatic fibrosis during a T-helper type 2-dominated inflammatory response. J Clin Invest. 1999;104:777–85.
Fallon PG, Richardson EJ, McKenzie GJ, McKenzie AN. Schistosome infection of transgenic mice defines distinct and contrasting pathogenic roles for IL-4 and IL-13: IL-13 is a profibrotic agent. J Immunol. 2000;164:2585–91.
McKenzie GJ, Fallon PG, Emson CL, Grencis RK, McKenzie AN. Simultaneous disruption of interleukin (IL)-4 and IL-13 defines individual roles in T helper cell type 2-mediated responses. J Exp Med. 1999;189:1565–72.
Wynn TA. Fibrotic disease and the T(H)1/T(H)2 paradigm. Nat Rev Immunol. 2004;4:583–94.
Kamdem SD, Moyou-Somo R, Brombacher F, Nono JK. Host regulators of liver fibrosis during human schistosomiasis. Front Immunol. 2018;9:2781.
Kolls JK, Lindén A. Interleukin-17 family members and inflammation. Immunity. 2004;21:467–76.
Gaffen SL. Structure and signalling in the IL-17 receptor family. Nat Rev. Immunol. 2009;9:556–67.
Chang SH, Dong C. Signaling of interleukin-17 family cytokines in immunity and inflammation. Cell Signal. 2011;23:1069–75.
Lafdil F, Miller AM, Ki SH, Gao B. Th17 cells and their associated cytokines in liver diseases. Cell Mol Immunol. 2010;7:250–4.
Miossec P, Korn T, Kuchroo VK. Interleukin-17 and type 17 helper T cells. N Engl. J Med. 2009;361:888–98.
Wilke CM, Bishop K, Fox D, Zou W. Deciphering the role of Th17 cells in human disease. Trends Immunol. 2011;32:603–11.
Sun X, Cai Y, Fleming C, Tong Z, Wang Z, Ding C, et al. Innate γδ T17 cells play a protective role in DSS-induced colitis via recruitment of Gr-1+ CD11b+ myeloid suppressor cells. Oncoimmunology. 2017;6:e1313369.
Steinman L. A brief history of T(H)17, the first major revision in the T(H)1/T(H)2 hypothesis of T cell-mediated tissue damage. Nat Med. 2007;13:139–45.
Bendelac A, Savage PB, Teyton L. The biology of NKT cells. Annu Rev Immunol. 2007;25:297–336.
St Leger AJ, Desai JV, Drummond RA, Kugadas A, Almaghrabi F, Silver P, et al. An ocular commensal protects against corneal infection by driving an interleukin-17 response from mucosal γδ T cells. Immunity. 2017;47(148–58):e5.
Shainheit MG, Lasocki KW, Finger E, Larkin BM, Smith PM, Sharpe AH, et al. The pathogenic Th17 cell response to major schistosome egg antigen is sequentially dependent on IL-23 and IL-1beta. J Immunol. 2011;187:5328–35.
Wen X, He L, Chi Y, Zhou S, Hoellwarth J, Zhang C, et al. Dynamics of Th17 cells and their role in Schistosoma japonicum infection in C57BL/6 mice. PLoS Negl Trop Dis. 2011;5:e1399.
Chen D, Luo X, Xie H, Gao Z, Fang H, Huang J. Characteristics of IL-17 induction by Schistosoma japonicum infection in C57BL/6 mouse liver. Immunology. 2013;139:523–32.
Zhang Y, Huang D, Gao W, Yan J, Zhou W, Hou X, et al. Lack of IL-17 signaling decreases liver fibrosis in murine schistosomiasis japonica. Int Immunol. 2015;27:317–25.
Chen X, Yang X, Li Y, Zhu J, Zhou S, Xu Z, et al. Follicular helper T cells promote liver pathology in mice during Schistosoma japonicum infection. PLoS Pathog. 2014;10:e1004097.
Farwa A, He C, Xia L, Zhou H. Immune modulation of Th1, Th2, and T-reg transcriptional factors differing from cytokine levels in Schistosoma japonicum infection. Parasitol Res. 2018;117:115–26.
Romano A, Hou X, Sertorio M, Dessein H, Cabantous S, Oliveira P, et al. FOXP3+ regulatory T cells in hepatic fibrosis and splenomegaly caused by Schistosoma japonicum: the spleen may be a major source of Tregs in subjects with splenomegaly. PLoS Negl Trop Dis. 2016;10:e0004306.
Gong W, Huang F, Sun L, Yu A, Zhang X, Xu Y, et al. Toll-like receptor-2 regulates macrophage polarization induced by excretory-secretory antigens from Schistosoma japonicum eggs and promotes liver pathology in murine schistosomiasis. PLoS Negl Trop Dis. 2018;12:e0007000.
Xu L, Xue B, Zhou L, Qiu Z, Zhang X, Xu N, et al. NP 30 stimulates Th17 differentiation through DC in schistosomiasis japonicum. Parasite Immunol. 2018;40:e12528.
Hayday AC. γδ cells: a right time and a right place for a conserved third way of protection. Annu Rev Immunol. 2000;18:975–1026.
Chien Y, Meyer C, Bonneville M. γδ T cells: first line of defense and beyond. Annu Rev Immunol. 2014;32:121–55.
Paul S, Lal G. Role of gamma-delta (γδ) T cells in autoimmunity. J Leukoc Biol. 2015;97:259–71.
Silva-Santos B, Serre K, Norell H. γδ T cells in cancer. Nat Rev Immunol. 2015;15:683–91.
Papotto PH, Ribot JC, Silva-Santos B. IL-17+ γδ T cells as kick-starters of inflammation. Nat Immunol. 2017;18:604–11.
Akitsu A, Iwakura Y. Interleukin-17-producing γδ T (γδ17) cells in inflammatory diseases. Immunology. 2018;155:418–26.
Peters C, Kabelitz D, Wesch D. Regulatory functions of γδ T cells. Cell Mol Life Sci. 2018;75:2125–35.
Wang X, Tian Z. γδ T cells in liver diseases. Front Med. 2018;12:262–8.
Zheng L, Hu Y, Wang Y, Huang X, Xu Y, Shen Y, et al. Recruitment of neutrophils mediated by Vγ2 γδ T cells deteriorates liver fibrosis induced by Schistosoma japonicum infection in C57BL/6 mice. Infect Immun. 2017;85:e01020–116.
Chiaramonte MG, Schopf LR, Neben TY, Cheever AW, Donaldson DD, Wynn TA. IL-13 is a key regulatory cytokine for Th2 cell-mediated pulmonary granuloma formation and IgE responses induced by Schistosoma mansoni eggs. J Immunol. 1999;162:920–30.
Fichtner-Feigl S, Strober W, Kawakami K, Puri RK, Kitani A. IL-13 signaling through the IL-13alpha2 receptor is involved in induction of TGF-beta1 production and fibrosis. Nat Med. 2005;12:99–106.
Giles DA, Moreno-Fernandez M, Divanovic S. IL-17 axis driven inflammation in non-alcoholic fatty liver disease progression. Curr Drug Targets. 2015;16:1315–23.
Ferri S, Longhi MS, De Molo C, Lalanne C, Muratori P, Granito A, et al. A multifaceted imbalance of T cells with regulatory function characterizes type 1 autoimmune hepatitis. Hepatology. 2010;52:999–1007.
Pellicoro A, Ramachandran P, Iredale JP, Fallowfield JA. Liver fibrosis and repair: immune regulation of wound healing in a solid organ. Nat Rev Immunol. 2014;14:181–94.
Hoh A, Dewerth A, Vogt F, Wenz J, Baeuerle PA, Warmann SW, et al. The activity of γδ T cells against paediatric liver tumour cells and spheroids in cell culture. Liver Int. 2013;33:127–36.
Buus TB, Geisler C, Lauritsen JP. The major diversification of Vγ11+ and Vγ2+ thymocytes in mice occurs after commitment to the γδ T cell lineage. Eur J Immunol. 2016;46:2363–75.
Xiong N, Raulet DH. Development and selection of γδ T cells. Immunol Rev. 2007;215:15–31.
Agrati C, D’Offizi G, Narciso P, Abrignani S, Ippolito G, Colizzi V, et al. Vδ1T lymphocytes expressing a Th1 phenotype are the major γδ T cell subset infiltrating the liver of HCV-infected persons. Mol Med. 2001;7:11–9.
Gao Y, Yang W, Pan M, Scully E, Girardi M, Augenlicht LH, et al. γδ T cells provide an early source of interferon γ in tumor immunity. J Exp Med. 2003;198:433–42.
Wang X, Ma K, Chen M, Ko KH, Zheng BJ, Lu L. IL-17A promotes pulmonary B-1a cell differentiation via induction of Blimp-1 expression during influenza virus infection. PLoS Pathog. 2016;12:e1005367.
Seo W, Eun HS, Kim SY, Yi HS, Lee YS, Park SH, et al. Exosome-mediated activation of toll-like receptor 3 in stellate cells stimulates interleukin-17 production by γδ T cells in liver fibrosis. Hepatology. 2016;64:616–31.
He X, Xie J, Wang Y, Fan X, Su Q, Sun Y, et al. Down-regulation of microRNA-203-3p initiates type 2 pathology during schistosome infection via elevation of interleukin-33. PLoS Pathog. 2018;14:e1006957.
Gressner AM, Weiskirchen R. Modern pathogenetic concepts of liver fibrosis suggest stellate cells and TGF-beta as major players and therapeutic targets. J Cell Mol Med. 2006;10:76–99.
Kresina TF, He Q, Degli Esposti S, Zern MA. Gene expression of transforming growth factor beta 1 and extracellular matrix proteins in murine Schistosoma mansoni infection. Gastroenterology. 1994;107:773–80.
Haile LA, von Wasielewski R, Gamrekelashvili J, Krüger C, Bachmann O, Westendorf AM, et al. Myeloid-derived suppressor cells in inflammatory bowel disease: a new immunoregulatory pathway. Gastroenterology. 2008;135:871–81.
Kumar V, Patel S, Tcyganov E, Gabrilovich DI. The nature of myeloid-derived suppressor cells in the tumor microenvironment. Trends Immunol. 2016;37:208–20.
Weston CJ, Zimmermann HW, Adams DH. The role of myeloid-derived cells in the progression of liver disease. Front Immunol. 2019;10:893.
Rutkowski MR, Stephen TL, Svoronos N, Allegrezza MJ, Tesone AJ, Perales-Puchalt A, et al. Microbially driven TLR5-dependent signaling governs distal malignant progression through tumor-promoting inflammation. Cancer Cell. 2015;27:27–40.
Krstic J, Obradovic H, Kukolj T, Slavko M, Ivana OD, Diana B, et al. An overview of interleukin-17A and interleukin-17 receptor a structure, interaction and signaling. Protein Pept Lett. 2015;22:570–8.
Ramani K, Tan RJ, Zhou D, Coleman BM, Jawale CV, Liu Y, et al. IL-17 receptor signaling negatively regulates the development of tubulointerstitial fibrosis in the kidney. Mediators Inflamm. 2018;2018:5103672.
Yanagisawa H, Hashimoto M, Minagawa S, Takasaka N, Ma R, Moermans C, et al. Role of IL-17A in murine models of COPD airway disease. Am J Physiol Lung Cell Mol Physiol. 2017;312:122–30.
Lebrun A, Lo Re S, Chantry M, Izquierdo Carerra X, Uwambayinema F, Ricci D, et al. CCR2 monocytic myeloid-derived suppressor cells (M-MDSCs) inhibit collagen degradation and promote lung fibrosis by producing transforming growth factor-β1. J Pathol. 2017;243:320–30.
Acknowledgements
We are grateful to Mr Yuxin Xu at the National Institute of Parasitic Diseases, Chinese Center for Disease Control and Prevention, for assisting with the mouse infection experiments, and to Dr Haipeng Liu at Shanghai Pulmonary Hospital, Tongji University, for helping with the draft of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (Nos. 8177225 and 81971969 to JC). The funders had no role in the study design, data collection and analysis, the decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
JC and LS conceived and designed the experiment. LS, WG, YS, LL, XZ, TL, TTC and YH performed the experiments. JC contributed reagents and materials. LS and JC analyzed the data. LS wrote the draft and JC revised the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This study was carried out in strict accordance with the recommendations of the Guide for the Care and Use of Laboratory Animals of the Ministry of Science and Technology, China. The protocol was approved by the Laboratory Animals Welfare and Ethics Committee of the National Institute of Parasitic Diseases, Chinese Center for Diseases Control and Prevention (Permit Number: NIPD-2017001). All the animal carcasses were disposed of by the Shanghai Animal Harmless Treatment Center. A special effort was made to reduce the number of mice used in the study, to provide them with the most comfortable conditions possible, and to minimize animal suffering where possible.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1: Figure S1.
Gating strategy for γδ T cells.
Additional file 2: Figure S2.
Isotype controls for intracellular staining. a, b Isotype controls (BV421-labeled IgG2a) for IFN-γ (a) and IL-17A (b). c Isotype controls (PE-labeled IgG2a) for TGF-β.
Additional file 3: Figure S3.
Survival curve of WT and TCR δ KO mice. The survival of two groups was monitored weekly for up to 16 weeks post-infection (n = 15 per group for one experiment).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Sun, L., Gong, W., Shen, Y. et al. IL-17A-producing γδ T cells promote liver pathology in acute murine schistosomiasis. Parasites Vectors 13, 334 (2020). https://doi.org/10.1186/s13071-020-04200-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-020-04200-4