
Abstract
Background
The long-lasting insecticidal nets (LLINs) and indoor residual spraying of insecticide (IRS) are major malaria vector control strategies in Mali. The success of control strategies depends on a better understanding of the status of malaria vectors with respect to the insecticides used. In this study we evaluate the level of resistance of Anopheles gambiae (sensu lato) to bendiocarb and the molecular mechanism that underlies it.
Methods
Larvae of An. gambiae (s.l.) were collected from breeding habitats encountered in the three study sites and bioassayed with bendiocarb. The ace-1 target site substitution G119S was genotyped using a TaqMan assay.
Results
The three species of the An. gambiae complex in Mali, i.e. An. arabiensis, An. coluzzii and An. gambiae (s.s.) were found in sympatry in the three surveyed localities with different frequencies. We observed a resistance and suspicious resistance of the three species to bendiocarb with a mortality rate ranging from 37% to 86%. The allelic frequency of the G119S mutation was higher in An. gambiae (s.s.) compared to the other two species; 42.86%, 25.61% and 16.67% respectively in Dangassa, Koula, and Karadié. The allelic frequency of G119S in An. coluzzii ranged from 4.5% to 8.33% and from 1.43% to 21.15% for An. arabiensis. After exposure to bendiocarb, the G119S mutation was found only in survivors. The survival of Anopheles gambiae (s.l) populations from the three surveyed localities was associated with the presence of the mutation.
Conclusions
The study highlights the implication of G119S mutation in bendiocarb resistance in An. gambiae (s.s.), An. arabiensis and An. coluzzii populations from the three surveyed localities.

Similar content being viewed by others

Background
Malaria vector control relies heavily on the use of long-lasting insecticidal nets (LLINs) and indoor residual spraying (IRS) in Mali. The wide deployment of these control tools is responsible for the current decline in malaria morbidity and mortality globally [1]. For example, LLINs alone are responsible for preventing approximately 68% of malaria deaths in Africa [2]. Four chemical classes of insecticide (organochlorines, pyrethroids, organophosphates, and carbamates) are recommended by the WHO for vector control [3, 4]. Pyrethroids are the only class of insecticide currently used in LLINs and unfortunately, a widespread resistance of the malaria vector is now being recorded with these insecticides. The proportion of malaria-endemic countries that monitored and subsequently reported pyrethroid resistance increased from 71% in 2010 to 81% in 2016 [5]. In addition, resistance to other classes of insecticides (carbamates and organophosphates) has been reported in several West African countries including Mali [6, 7]. Thus, the resistance of malaria vectors to all commonly used classes of insecticide in public health will compromise malaria control efforts in many countries if a good insecticide resistance plan is not put in place [1, 8, 9].
Mali experienced a nationwide distribution of LLINs and the implementation of IRS in selected districts with the support of the US Presidential Malaria Initiative (PMI) and other partners. Different classes of insecticides have been used in the course of this programme. Pyrethroids (lamda-cyhalothrin and deltamethrin) were first used between 2008 and 2010, then replaced with carbamates (bendiocarb) in 2011 and, recently, organophosphates (pirimiphos-methyl) was introduced in this programme. Resistance to carbamates and organophosphates has been well documented in many sub-Saharan countries [10,11,12,13,14]. It was first reported in Culex quinquefasciatus from Côte d’Ivoire [15]. Reduced susceptibility to organophosphates (Ops) and carbamates (CMs) was recently observed in An. gambiae populations in the centre and the north of Côte d’Ivoire [16] and Benin [17]. The main resistance mechanisms to CMs and OPs documented thus far are either metabolic or target-site mutations. Both resistance mechanisms are widespread in Anopheles gambiae (sensu lato) and Anopheles funestus, the main malaria vectors in Africa. Metabolic resistance to carbamates is often conferred by the upregulation of detoxification genes such as cytochrome P450s [18, 19]. Target-site resistance to CMs and OPs is conferred by a single point mutation causing acetylcholinesterase inhibition [20, 21]. The mutation encoded by the ace-1R gene induces a substitution from a glycine (GGC) codon to a serine (AGC) codon at position 119 (mutation G119S) in resistant mosquitoes [21]. An alternative mutation of the ace-1R gene (F290V) has been described in Culex pipiens; the latter is due to a substitution of phenylalanine (F) by valine (V) at position 290 [22].
In Mali, scarce data exist on carbamate and organophosphate resistance and the different mechanisms underlying this resistance. This study reports the presence of bendiocarb resistance in wild populations of An. arabiensis, An. coluzzii and An. gambiae (s.s.) in Mali and further investigates the allelic frequency of the G119S mutations in these wild populations of An. gambiae (s.l.).
Methods
Study area
In Mali, the vector control strategy is based on LLINs, which was scaled up to universal coverage in 2014. In addition, from 2008 to 2016, Koulikoro district was one of the selected districts for IRS implementation in conjunction with LLINs. In the course of this IRS campaigns, three classes of insecticide (pyrethroids, carbamates and organophosphates) were used.
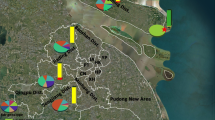
This study was conducted in the village of Dangassa (8° 12′ 38″ W, 12°8′ 46″ N) located in the health district of Ouéléssébougou, along the River Niger in the Sudan Savanna region, and the villages of Koula (7° 39′ 22″ W, 13° 7′ 28″ N) and Karadie (7° 36’34″ W, 13° 16′ 12″ N) in the health district of Koulikoro, in the Sudano-Sahelian region (Fig. 1). Malaria transmission mainly occurred during the rainy season (June to October) in both areas; however, the presence of the River Niger in the district of Ouéléssébougou makes it a year-round transmission area.

Map showing the study sites in administrative region of Koulikoro, Mali
The main economic activity in the each of the three localities is agriculture. The main crops are cotton, millet, peanuts and soybeans. Multiple classes of pesticides such as pyrethroids, carbamates and organophosphates are used to protect these crops against agricultural pests. Plasmodium falciparum is the predominant malaria parasite species found in all of the localities and is transmitted by species of the An. gambiae species complex. The prevalence of malaria infection varies from 40% to 50% in children under 5 years-old. Resistance to pyrethroids has been reported in all the three villages.
Mosquito collections
Larvae (L1 to L4) of An. gambiae (s.l.) were collected in different types of breeding habitats including puddles, brick pits, ponds, tires and animal footprints found in and around each village using the “dipping” technique as described by Service [23]. Collected larvae were transported to the insectary of ICER-Mali in Bamako, where they were maintained at 28 ± 2 °C and a relative humidity of 72 ± 5%. Larvae were separated into different instars in order to have adult mosquitoes emerging at the same time. Larvae were fed with Tetramin powder (Tetra, Herrenteich, Germany), a fish-food, until pupation stage and transferred into cages for adult production. Cotton wool pads soaked with 10% sugar solution were used for feeding emerging adult mosquitoes.
Insecticide susceptibility bioassays
Three to five days after emergence, adult mosquitoes were sexed before being subjected to the insecticide susceptibility test following the standard protocol of the World Health Organization [24, 25]. All tests were performed in the morning between 8:00 h and 12:00 h in a controlled air-conditioned room at 27 ± 1 °C and a relative humidity of 80%. Test papers impregnated with recommended diagnostic concentrations of 0.1% bendiocarb were used. Insecticide papers were obtained from the WHO reference centre at the Vector Control Research Unit, University Sains Malaysia. Mosquitoes were transferred from holding tubes to test tubes laminated with the insecticide-impregnated paper for one hour. After one-hour of exposure to the insecticide, mosquitoes were transferred again into holding tubes (no insecticide), kept in the insectary and fed with sugar solution. Mortalities were recorded at 24 h post-exposure to insecticide.
Alive mosquitoes were kept in 1.5 ml Eppendorf tubes containing RNA-later (Sigma-Aldrich, Steinheim, Germany) and the dead mosquitoes were kept in tubes with silica gel. All preserved mosquitoes were stored in a freezer at -20 °C for molecular identification and resistant gene screening. Data from molecular identification were used for subgrouping dead and alive mosquitoes into species of the An. gambiae complex, and determining susceptibility profiles of each species of this complex, i.e. An. arabiensis, An. coluzzii and An. gambiae (s.s.) to bendiocarb in each surveyed village.
Molecular identification of An. gambiae (s.l.)
For each village, dead and alive subgroups of An. gambiae (s.l.) exposed to bendiocarb were molecularly identified and separately sorted to record susceptibility profiles to bendiocarb of wild An. arabiensis, An. coluzzii and An. gambiae (s.s.). For this experiment, the genomic DNA of both alive and dead mosquitoes was extracted using the DNA extraction protocol described by Livak [26]. Specific DNA sequences were amplified using the technique of Santolamazza et al. [27] for identification of An. arabiensis, An. coluzzii and An. gambiae (s.s.).
Screening of ace-1 mutation in An. gambiae (s.l.) from the study sites
We determined the allelic frequency of ace-1 in each member of the An. gambiae complex, i.e. An. arabiensis, An. coluzzii and An. gambiae (s.s.) using the TaqMan SNP genotyping assay. The entire target marker ace-1R (G119S) was diluted in a total volume of 10 μl containing 2× qPCR Sensimix (Bioline, Cincinnati, USA), 80× primer/probe mix [Ace 1 Forward (5’-GGC CGT CAT GCT GTG GAT-3’); Ace1 Reverse (5’-GCG GTG CCG GAG TAG A-3’); ACE1-VIC (5′-TTC GGC GGC GGC T-3′); ACE1-6-FAM (5′-TTC GGC GGC AGC T-3′)], nuclease-free water and 1 μl template DNA. Probes were labeled with two specific fluorescent dyes, FAM and HEX. The reporter dye (FAM) was used to detect homozygous resistant genotypes (RR), while the quencher fluorescent dye (HEX) was used for the detection of homozygous susceptible genotypes (SS). Both FAM and HEX are specific for the detection of heterozygous resistant/susceptible genotypes (RS). Amplifications were performed in an Agilent MX3000 real-time qPCR machine (Agilent Technologies, Santa Clara, CA, USA) with cycling conditions of 95 °C for 10 min, followed by 40 cycles at 95 °C for 10 s and 60 °C for 45 s. FAM and HEX fluorescence was captured at the end of each cycle and genotypes were called from endpoint fluorescence using the MxPro software (Agilent).
Linking observed bendiocarb resistance to ace-1 mutation
To further analyze phenotypic and genotypic associations in observed resistance profiles, we conducted separately, on dead and alive mosquitoes, TaqMan SNP genotyping assays as described above [28]. We determined the allelic frequency of ace-1R (G119S) in dead and alive mosquitoes post-exposure to bendiocarb. These analyses were conducted at the Anopheles gambiae (s.l.) level but also disaggregated for each member (An. arabiensis, An. coluzzii and An. gambiae (s.s.)).
Data analysis
Following exposure of mosquito populations to bendiocarb, those with recorded mortalities between 98–100% were rated as susceptible. Populations showing mortality below 98% were rated as suspicious of resistance and populations showing mortality below 90% were rated as resistant [25]. Genotype distributions were recorded in an Excel datasheet (Microsoft Office 2017; Microsoft Corporation, Redmond, USA) and analysis performed using SPSS 25.0. Allelic frequencies were calculated using the following formula ƒ(R) = (2n.RR + n.RS)/2N, where n is the number of mosquitoes of a given genotype, RR represents the homozygote resistance allele, RS represents the heterozygote resistance allele, SS represents the susceptible allele, and N is the total number of mosquitoes tested. After PCR identification for each species per study site, the cumulative binomial exact test was used to test the significance of the resistance on the sample size where the recommended number was not reached. The Chi-square test or Fisher’s exact test where appropriate were used to test for significant associations between ace-1 genotypes and observed phenotypic resistance for the various exposed populations of An. gambiae (s.l.). The logistic regression model, with the alive status as the dependent variable, was used to measure the adjusted association between the independent variables (species and study site) and the dependent variable, and to compare different categories within each independent variable.
Results
Molecular identification within the An. gambiae complex in the three surveyed localities
Out of the 263 specimens of An. gambiae (s.l.) analyzed by PCR in the three investigated villages 110 (41.98%) were identified as An. gambiae (s.s.), 85 (32.3%) as An. arabiensis and 68 (25.9%) as An. coluzzii. Species composition of An. gambiae (s.l.) varied significantly between study sites (χ2 = 20.36, df = 4, P < 0.0001). Anopheles gambiae (s.s.) was predominant in Koula (48.8%, 41/84) and Karadié (47.2%, 42/89) (Fig. 2). Anopheles coluzzii was the most prevalent species in Dangassa (41.1%, 37/90) followed by Koula (22.6%, 19/84), while An. arabiensis was prevalent in Karadié (39.3%, 35/89) followed by Dangassa (28.9%, 26/90). In all other sites, An. gambiae (s.s.) was predominant. The three species were found in sympatry in all sites.

Frequency distribution of species of the Anopheles gambiae complex in wild populations exposed to diagnostic doses bendiocarb in all sites in 2016
Susceptibility to carbamates
Resistance of An. gambiae (s.l.) to bendiocarb was recorded in the three surveyed localities (Table 1, Fig. 3). In bivariate analysis, the rating of the populations as resistant was significant (P < 0.05) for all species with respect to localities within all strata (by locality and by species), except for An. arabiensis in Koula and Karadié (P = 0.21 and P = 0.27, respectively; testing the hypothesis of mortality < 90%) and An. coluzii in Karadié (P = 0.34), where the rating of the populations as suspicious resistant was significant (P < 0.05; testing the hypothesis of mortality < 97%). The highest mortality rate was observed within the latter three strata (83.33%, 85.71% and 83.33%, respectively, see Table 1). The resistance levels of An. gambiae (s.s.) were significantly high (P < 0.05) in all three localities. When this resistance was analyzed with respect to species of the An. gambiae complex, we recorded some variations in resistance patterns within the three species in the three localities (Fig. 3). All species populations were significantly rated as resistant in Dangassa (all P < 0.05). Highest mortalities were recorded for An. arabiensis in the localities of Koula and Karadié (83% and 86%, respectively).

Mortality of wild populations of species of the Anopheles gambiae complex exposed to diagnostic doses of bendiocarb in all sites in 2016
The logistic regression models testing adjusted association between bendiocarb resistance (low mortality rate) with the species and with the localities were globally both significant (P = 0.036 and P = 0.006, respectively). There was a significant difference between the mortality rate of An. arabiensis compared to An. gambiae (s.s.) (OR = 0.439, P = 0.011). However, the adjusted difference of the mortality rate between localities were not significant (all P > 0.05) (Table 2).
Distribution of ace-1 mutation in wild populations of sampled Anopheles species
Two hundred and sixty mosquitoes from Koula (n = 84), Karadié (n = 89 ) and Dangassa (n = 90) were assayed for molecular form and analyzed for their ace-1 genotype (Table 3). All specimens carrying the gene were heterozygotes. Generated data for the allelic frequency of the ace-1R mutation revealed relatively high frequency of this mutation in wild populations of An. gambiae (s.l.). from Dangassa compared to Koula and Karadié. At Dangassa, the mutation was dominant in An. gambiae (s.s.) (42.9%) followed by An. arabiensis (21.2%) and An. coluzzii (4.1%) with significant differences between species (χ2 = 10.3, df = 2, P = 0.006). At Koula, the mutation remained with high frequency in An. gambiae (s.s.) (25.6%) followed by An. coluzzii and An. arabiensis (χ2 = 18.33, df = 2, P < 0.0001); a similar trend was recorded in Karadié (χ2 = 11.53, df = 2, P = 0.003) (Fig. 4). We observed a relatively high frequency of ace-1 mutation in An. arabiensis from Dangassa (21.2%) compared to An. arabiensis from Koula and Karadié, which had low levels of mutation (2.1% and 1.4%, respectively) (χ2 = 21.12, df = 2, P < 0.0001; Fig. 4). For the An. coluzzii the difference between the three localities was not significant (χ2 = 1.05, df = 2, P = 0.59). In contrast, the distribution of ace-1 in An. gambiae (s.s.) was significantly different across the localities with the high presence in Dangassa (42.86%) (χ2 = 6.31, df = 2, P = 0.043).

Allelic frequency of the ace-1 mutation in wild populations of species of the Anopheles gambiae complex from the surveyed localities
Association between ace-1 mutation frequencies and recorded bendiocarb resistance profiles in the three surveyed localities
The analysis of the presence of the resistant allele of ace-1 in alive and dead individuals following exposure to bendiocarb showed consistent associations between mosquito survivors after bendiocarb exposure and the presence of the mutation (Table 4); indeed, none of the dead mosquitoes carried the mutation. The mutation was never found in the population of individuals dying post-exposure to bendiocarb. The survival of Anopheles gambiae (s.l.). populations from the three surveyed localities was associated with the presence of the mutation. However, the fact that some survivors did not carry the mutation highlights the presence of other bendiocarb resistance mechanisms in these localities.
Association between ace-1 mutation frequencies and recorded bendiocarb resistance profiles in members of the An. gambiae complex in the three surveyed localities
When data of allelic frequencies of G119S mutation were disaggregated at the species level (Table 4), we recorded an association of survival after bendiocarb exposition and the presence of ace-1 mutations in An. gambiae (s.s.). Allelic frequencies of 40.4%, 46.7% and 42.9% were recorded in surviving An. gambiae (s.s.) populations at Koula, Karadié and Dangassa, respectively. Similarly, most bendiocarb surviving An. arabiensis and An. coluzzii also carried the G119S mutation, but not at high levels as in An. gambiae (s.s.) with a few exceptions (Table 4).
Discussion
Currently, there is growing concern that the widespread resistance of the malaria vector to pyrethroid insecticides reduces the effectiveness of LLINs and IRS and compromises the current decline in malaria morbidity and mortality globally. An insecticide resistance management strategy may help to moderate these effects. The first step of insecticide resistance management is the determination and monitoring of the resistance mechanisms prevailing in the area. In Mali, the target site mutation, specifically the knockdown resistance (kdr) allele has been well documented [7, 29, 30] but data on the other resistance mechanisms are scarce. In this study, we assessed the status of An. gambiae (s.l.) to CMs and determined the frequency of the ace-1 mutation in the health districts of Koulikoro (with LLINs plus IRS), and Ouéléssébougou (with only LLINs).
Molecular identification within the An. gambiae complex in the three surveyed localities
Our results revealed the sympatric presence of An. gambiae (s.s.), An. arabiensis and An. coluzzii in the three sites with different proportions as reported by a number of studies in Mali [7, 31, 32] . Anopheles gambiae (s.s.) was the major species in Koula and Karadié and An. coluzzii in Dangassa. This is certainly related to the types of breeding habitats found in each of the two areas. In Koula and Karadié, larval habitats are rainfall dependent and sunny, such as brick pits, tire prints and puddles, which are preferred by An. gambiae (s.s.). In Dangassa, the flooded plain that separates the village from the River Niger constitutes the favorable environment for the development of An. coluzzii [33, 34].
Susceptibility to carbamates
All three species of the An. gambiae complex (An. gambiae (s.s.), An. arabiensis and An. coluzzii) found in the study sites were resistant to bendiocarb in Dangassa, while An. gambiae (s.s.) was rated as resistant in all the three localities at the significance lavel of P < 0.05. There was variation in the level of resistance by species and by area. In Dangassa, cotton and vegetable cropping is very common and therefore the use of agricultural pesticides in addition to the wide deployment of the LLINs may have triggered a selection pressure on mosquitoes, as reported in many studies across West Africa [35,36,37,38,39]. The resistance observed in Koula and Karadié can be attributed to the use of bendiocarb in the IRS campaigns from 2011 to 2013 in these localities. Indeed, previous data from sentinel sites of the National Malaria Control Programme across the country showed low resistance of species of the An. gambiae complex to bendiocarb [6, 7]; resistance of species of the complex to the carbamate class of insecticide has been reported elsewhere in Africa in many studies [10, 13, 40, 41].
Distribution of ace-1R mutation in wild populations of sampled Anopheles species
Our study showed the presence of the G119S mutation in the three species and at all study sites. The allelic frequencies of the G119S mutation were higher in An. gambiae (s.s.) than in An. coluzzii in all study sites. Similar observations were reported in Burkina Faso [42] and in Ghana [12]. However, higher allelic frequencies of the G119S mutation was reported in An. coluzzii compared to An. gambiae (s.s.) in Côte d’Ivoire [11]. In Mali, the only study on the G119S mutation was carried out by Cissé et al. [7] who reported a low frequency of the mutation in these two species. We also noticed a higher frequency of the mutation G116S in An. arabiensis in Dangassa, in the Sudan Savanna region, compared to Karadié and Koula, in the Sahel region. Similar observations were made in Burkina Faso [43].
Association between ace-1R mutation frequency and recorded bendiocarb resistance profiles in the three surveyed localities
Our study showed that all resistant specimens carried the resistant allele (R) in its heterozygous form. The absence of a homozygous specimen is probably due to the high mortality of resistant homozygotes caused by the G119S mutation. Indeed, many studies [21, 44, 45] reported that homozygous resistant individuals are most likely to die during pupation than susceptible individuals. Therefore, in the area where the ace-1 resistant allele is present, the resistant mosquitoes will mainly be of the heterozygous (ace-1 RS) status. Djogbénou et al. [46] reported that the presence of the ace-1 RS affects the body size of adult mosquitoes, with resistant individuals being smaller than the susceptible ones. Our results showed that the G119S mutation was present in alive specimens of all study sites. This observation is in accordance with other studies [47, 48] supporting the association of the G119S mutation with the survival of An. gambiae (s.l.) populations. Our results also showed that the ace-1 gene frequency was associated with An. arabiensis resistance to bendiocarb in Dangassa, An. coluzzii in Koula and Dangassa, and An. gambiae (s.s.) in all surveyed localities.
Conclusions
This study documented the resistance levels to bendiocarb of all three species of the An. gambiae complex in Mali. We have also shown the link between the molecular mechanism and the observed phenotypic resistance and implicated the G119S mutation in bendiocarb resistance in An. gambiae (s.l.) populations. Periodic updates of data on resistance of major vectors are required for a rational planning of insecticide deployment and insecticide resistance mitigation.
Availability of data and materials
Data supporting the conclusions of this article are included within the article. The datasets analyzed during the present study are available from the corresponding author upon reasonable request.
Abbreviations
- NMCP:
-
National Malaria Control Programmes
- WHO:
-
World Health Organization
- LLINs:
-
long-lasting insecticidal nets
- IRS:
-
indoor residual spraying
- ace-1 R :
-
acetylcholinesterase-1 resistance
- SNP:
-
single-nucleotide polymorphism
- R:
-
resistant
- S:
-
susceptible
References
WHO. Plan mondial pour la gestion de la résistance aux insecticides chez les vecteurs du paludisme Résumé dʼorientation (mai 2012). Geneva: World Health Organization; 2012. www.who.int/malaria/publications/atoz/gpirm/fr/. Accessed 22 Apr 2019.
Bhatt S, Weiss DJ, Cameron E, Bisanzio D, Mappin B, Dalrymple U, et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature. 2015;526:207–11.
WHO. Global insecticide use for vector-borne disease control. A 10-year assesment (2000–2009). 5th edn. Geneva; World Health Organization; 2011.
Van den Berg H, Zaim M, Yadav RS, Soares A, Ameneshewa B, Mnzava A, et al. Global trends in the use of insecticides to control vector-borne diseases. Environ Health Perspect. 2012;120:577–82.
WHO: World malaria report 2017. Geneva; World Health Organization; 2017.
Keita M, Traore S, Sogoba N, Dicko AM, Coulibaly B, Sacko A, et al. Susceptibility status of Anopheles gambiae sensu lato to insecticides commonly used for malaria control in Mali. Bull Soc Pathol Exot. 2016;109:39–45.
Cisse MB, Keita C, Dicko A, Dengela D, Coleman J, Lucas B, et al. Characterizing the insecticide resistance of Anopheles gambiae in Mali. Malar J. 2015;14:327.
Alemayehu E, Asale A, Eba K, Getahun K, Tushune K, Bryon A, et al. Mapping insecticide resistance and characterization of resistance mechanisms in Anopheles arabiensis (Diptera: Culicidae) in Ethiopia. Parasit Vectors. 2017;10:407.
Namountougou M, Simard F, Baldet T, Diabate A, Ouedraogo JB, Martin T, et al. Multiple insecticide resistance in Anopheles gambiae s.l. populations from Burkina Faso, West Africa. PLoS ONE. 2012;7:e48412.
Oduola AO, Idowu ET, Oyebola MK, Adeogun AO, Olojede JB, Otubanjo OA, et al. Evidence of carbamate resistance in urban populations of Anopheles gambiae s.s. mosquitoes resistant to DDT and deltamethrin insecticides in Lagos, south-western Nigeria. Parasit Vectors. 2012;5:116.
Ahoua Alou LP, Koffi AA, Adja MA, Tia E, Kouassi PK, Kone M, et al. Distribution of ace-1R and resistance to carbamates and organophosphates in Anopheles gambiae s.s. populations from Côte d’Ivoire. Malar J. 2010;9:167.
Essandoh J, Yawson AE, Weetman D. Acetylcholinesterase (Ace-1) target site mutation 119S is strongly diagnostic of carbamate and organophosphate resistance in Anopheles gambiae s.s. and Anopheles coluzzii across southern Ghana. Malar J. 2013;12:404.
Antonio-Nkondjio C, Poupardin R, Tene BF, Kopya E, Costantini C, Awono-Ambene P, et al. Investigation of mechanisms of bendiocarb resistance in Anopheles gambiae populations from the city of Yaounde, Cameroon. Malar J. 2016;15:424.
Camara S, Koffi AA, Ahoua Alou LP, Koffi K, Kabran JK, Kone A, et al. Mapping insecticide resistance in Anopheles gambiae (s.l.) from Côte d’Ivoire. Parasit Vectors. 2018;11:19.
Chandre F, Darriet F, Doannio JM, Riviere F, Pasteur N, Guillet P. Distribution of organophosphate and carbamate resistance in Culex pipiens quinquefasciatus (Diptera: Culicidae) in West Africa. J Med Entomol. 1997;34:664–71.
N’Guessan R, Darriet F, Guillet P, Carnevale P, Traore-Lamizana M, Corbel V, et al. Resistance to carbosulfan in Anopheles gambiae from Ivory Coast, based on reduced sensitivity of acetylcholinesterase. Med Vet Entomol. 2003;17:19–25.
Corbel V, N’Guessan R, Brengues C, Chandre F, Djogbenou L, Martin T, et al. Multiple insecticide resistance mechanisms in Anopheles gambiae and Culex quinquefasciatus from Benin, West Africa. Acta Trop. 2007;101:207–16.
Edi CV, Djogbenou L, Jenkins AM, Regna K, Muskavitch MA, Poupardin R, et al. CYP6 P450 enzymes and ACE-1 duplication produce extreme and multiple insecticide resistance in the malaria mosquito Anopheles gambiae. PLoS Genet. 2014;10:e1004236.
Ibrahim SS, Ndula M, Riveron JM, Irving H, Wondji CS. The P450 CYP6Z1 confers carbamate/pyrethroid cross-resistance in a major African malaria vector beside a novel carbamate-insensitive N485I acetylcholinesterase-1 mutation. Mol Ecol. 2016;25:3436–52.
Weill M, Lutfalla G, Mogensen K, Chandre F, Berthomieu A, Berticat C, et al. Comparative genomics: insecticide resistance in mosquito vectors. Nature. 2003;423:136–7.
Weill M, Malcolm C, Chandre F, Mogensen K, Berthomieu A, Marquine M, et al. The unique mutation in ace-1 giving high insecticide resistance is easily detectable in mosquito vectors. Insect Mol Biol. 2004;13:1–7.
Alout H, Berthomieu A, Hadjivassilis A, Weill M. A new amino-acid substitution in acetylcholinesterase 1 confers insecticide resistance to Culex pipiens mosquitoes from Cyprus. Insect Biochem Mol Biol. 2007;37:41–7.
Service MW. Mosquito ecology: field sampling methods. London: Chapman and Hall; 1993.
Gillies MT, Coetzee M. A supplement to the Anophelinae of Africa South of the Sahara (Afrotropical Region). Johannesburg: The South African Institute for Medical Research; 1987.
WHO. Test procedures for insecticide resistance monitoring in malaria vector mosquitoes (Second edition). Geneva: World Health Organization; 2016.
Livak KJ. Organization and mapping of a sequence on the Drosophila melanogaster X and Y chromosomes that is transcribed during spermatogenesis. Genetics. 1984;107:611–34.
Santolamazza F, Mancini E, Simard F, Qi Y, Tu Z, della Torre A. Insertion polymorphisms of SINE200 retrotransposons within speciation islands of Anopheles gambiae molecular forms. Malar J. 2008;7:163.
Bass C, Nikou D, Vontas J, Williamson MS, Field LM. Development of high-throughput real-time PCR assays for the identification of insensitive acetylcholinesterase (ace-1R) in Anopheles gambiae. Pest Biochem Phys. 2010;96:80–5.
Fanello C, Petrarca V, della Torre A, Santolamazza F, Dolo G, Coulibaly M, et al. The pyrethroid knock-down resistance gene in the Anopheles gambiae complex in Mali and further indication of incipient speciation within An. gambiae s.s. Insect Mol Biol. 2003;12:241–5.
Tripet F, Wright J, Cornel A, Fofana A, McAbee R, Meneses C, et al. Longitudinal survey of knockdown resistance to pyrethroid (kdr) in Mali, West Africa, and evidence of its emergence in the Bamako form of Anopheles gambiae s.s.. Am J Trop Med Hyg. 2007;76:81–7.
Adamou A, Dao A, Timbine S, Kassogue Y, Yaro AS, Diallo M, et al. The contribution of aestivating mosquitoes to the persistence of Anopheles gambiae in the Sahel. Malar J. 2011;10:151.
Keita M, Baber I, Sogoba N, Maiga HM, Diallo M, Doumbia S, et al. Vectorial transmission of malaria in a village along the Niger River and its fishing hamlet (Kenieroba and Fourda, Mali). Bull Soc Pathol Exot. 2014;107:356–68.
Sogoba N, Vounatsou P, Bagayoko MM, Doumbia S, Dolo G, Gosoniu L, et al. The spatial distribution of Anopheles gambiae sensu stricto and An. arabiensis (Diptera: Culicidae) in Mali. Geospat Health. 2007;1:213–22.
Dolo G, Briet OJ, Dao A, Traore SF, Bouare M, Sogoba N, et al. Malaria transmission in relation to rice cultivation in the irrigated Sahel of Mali. Acta Trop. 2004;89:147–59.
Chouaibou MS, Fodjo BK, Fokou G, Allassane OF, Koudou BG, David JP, et al. Influence of the agrochemicals used for rice and vegetable cultivation on insecticide resistance in malaria vectors in southern Côte d’Ivoire. Malar J. 2016;15:426.
Hien AS, Soma DD, Hema O, Bayili B, Namountougou M, Gnankine O, et al. Evidence that agricultural use of pesticides selects pyrethroid resistance within Anopheles gambiae sl populations from cotton growing areas in Burkina Faso, West Africa. PLoS ONE. 2017;12:0173098.
Yadouleton AW, Asidi A, Djouaka RF, Braima J, Agossou CD, Akogbeto MC. Development of vegetable farming: a cause of the emergence of insecticide resistance in populations of Anopheles gambiae in urban areas of Benin. Malar J. 2009;8:103.
Akogbeto MC, Djouaka RF, Kinde-Gazard DA. Screening of pesticide residues in soil and water samples from agricultural settings. Malar J. 2006;5:22.
Diabate A, Baldet T, Chandre F, Akoobeto M, Guiguemde TR, Darriet F, et al. The role of agricultural use of insecticides in resistance to pyrethroids in Anopheles gambiae s.l. in Burkina Faso. Am J Trop Med Hyg. 2002;67:617–22.
Aikpon R, Agossa F, Osse R, Oussou O, Aizoun N, Oke-Agbo F, et al. Bendiocarb resistance in Anopheles gambiae s.l. populations from Atacora department in Benin, West Africa: a threat for malaria vector control. Parasit Vectors. 2013;6:192.
Rakotondranaivo T, Randriamanarivo SF, Tanjona MR, Vigan-Womas I, Randrianarivelojosia M, Ndiath MO. Evidence of insecticide resistance to pyrethroids and bendiocarb in Anopheles funestus from Tsararano, Marovoay District, Madagascar. Biomed Res Int. 2018;2018:5806179.
Dabire KR, Diabate A, Namontougou M, Djogbenou L, Kengne P, Simard F, et al. Distribution of insensitive acetylcholinesterase (ace-1R) in Anopheles gambiae s.l. populations from Burkina Faso (West Africa). Trop Med Int Health. 2009;14:396–403.
Dabire RK, Namountougou M, Diabate A, Soma DD, Bado J, Toe HK, et al. Distribution and frequency of kdr mutations within Anopheles gambiae s.l. populations and first report of the ace1 G119S mutation in Anopheles arabiensis from Burkina Faso (West Africa). PLoS ONE. 2014;9:e101484.
Djogbenou L, Chandre F, Berthomieu A, Dabire R, Koffi A, Alout H, et al. Evidence of introgression of the ace-1R mutation and of the ace-1 duplication in West African Anopheles gambiae s.s. PLoS ONE. 2008;3:2172.
Asidi AN, N’Guessan R, Koffi AA, Curtis CF, Hougard JM, Chandre F, et al. Experimental hut evaluation of bednets treated with an organophosphate (chlorpyrifos-methyl) or a pyrethroid (lambda-cyhalothrin) alone and in combination against insecticide-resistant Anopheles gambiae and Culex quinquefasciatus mosquitoes. Malar J. 2005;4:25.
Djogbenou L, Noel V, Agnew P. Costs of insensitive acetylcholinesterase insecticide resistance for the malaria vector Anopheles gambiae homozygous for the G119S mutation. Malar J. 2010;9:12.
Edi CV, Koudou BG, Jones CM, Weetman D, Ranson H. Multiple-insecticide resistance in Anopheles gambiae mosquitoes. Southern Côte d’Ivoire. Emerg Infect Dis. 2012;18:1508–11.
Weetman D, Mitchell SN, Wilding CS, Birks DP, Yawson AE, Essandoh J, et al. Contemporary evolution of resistance at the major insecticide target site gene Ace-1 by mutation and copy number variation in the malaria mosquito Anopheles gambiae. Mol Ecol. 2015;24:2656–72.
Acknowledgements
The study received support from the National Institutes of Allergy and Infectious Diseases (NIAID) through the West African International Center of Excellence for Malaria Research (ICEMR): NIAID U19 AI 089696 and U19 AI 129387. We thank the research team and the population of all study sites for their collaboration. We also acknowledge the technical assistance and collaboration of Boubacar Coulibaly, Romaric Akoton, Yaya Ibrahim Coulibaly and Moussa Diallo.
Funding
This work was funded by the TDR/WHO under grant No. B20388.
Author information
Authors and Affiliations
Contributions
MK and NS designed the study and monitored its implementation. MK and OT analyzed and interpreted the results. MK, BT and FK conducted field sampling and carried out bioassay tests. MK and FZ contributed in laboratory analysis of sampled mosquitoes to detect the target site mutation. MK wrote the manuscript which was critically revised by OT, NS, SD and RD. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Keïta, M., Kané, F., Thiero, O. et al. Acetylcholinesterase (ace-1R) target site mutation G119S and resistance to carbamates in Anopheles gambiae (sensu lato) populations from Mali. Parasites Vectors 13, 283 (2020). https://doi.org/10.1186/s13071-020-04150-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-020-04150-x