
Abstract
Background
Algal harvesting is a major cost which increases biofuel production cost. Algal biofuels are widely studied as third-generation biofuel. However, they are yet not viable because of its high production cost which is majorly contributed by energy-intensive biomass harvesting techniques. Biological harvesting method like fungal-assisted harvesting of microalgae is highly efficient but poses a challenge due to its slow kinetics and poorly understood mechanism.
Results
In this study, we investigate Aspergillus fumigatus–Chlorella pyrenoidosa attachment resulting in a harvesting efficiency of 90% within 4 h. To pinpoint the role of extracellular metabolite, several experiments were performed by eliminating the C. pyrenoidosa or A. fumigatus spent medium from the C. pyrenoidosa–A. fumigatus mixture. In the absence of A. fumigatus spent medium, the harvesting efficiency dropped to 20% compared to > 90% in the control, which was regained after addition of A. fumigatus spent medium. Different treatments of A. fumigatus spent medium showed drop in harvesting efficiency after periodate treatment (≤ 20%) and methanol–chloroform extraction (≤ 20%), indicating the role of sugar-like moiety. HR-LC–MS (high-resolution liquid chromatography–mass spectrometry) results confirmed the presence of N-acetyl-d-glucosamine (GlcNAc) and glucose in the spent medium. When GlcNAc was used as a replacement of A. fumigatus spent medium for harvesting studies, the harvesting process was significantly faster (p < 0.05) till 4 h compared to that with glucose. Further experiments indicated that metabolically active A. fumigatus produced GlcNAc from glucose. Concanavalin A staining and FTIR (Fourier transform infrared spectroscopy) analysis of A. fumigatus spent medium- as well as GlcNAc-incubated C. pyrenoidosa cells suggested the presence of GlcNAc on its cell surface indicated by dark red dots and GlcNAc-specific peaks, while no such characteristic dots or peaks were observed in normal C. pyrenoidosa cells. HR-TEM (High-resolution Transmission electron microscopy) showed the formation of serrated edges on the C. pyrenoidosa cell surface after treatment with A. fumigatus spent medium or GlcNAc, while Atomic force microscopy (AFM) showed an increase in roughness of the C. pyrenoidosa cells surface upon incubation with A. fumigatus spent medium.
Conclusions
Results strongly suggest that GlcNAc present in A. fumigatus spent medium induces surface changes in C. pyrenoidosa cells that mediate the attachment to A. fumigatus hyphae. Thus, this study provides a better understanding of the A. fumigatus-assisted C. pyrenoidosa harvesting process.
Similar content being viewed by others

Background
Microalgae showcase diverse applications for wastewater treatment and subsequent bioenergy generation. Large quantity of microalgal biomass is the foremost requirement in the biofuel route, for which several high-end/advanced photobioreactors have already been designed by several researchers [1,2,3]. However, the commercialization of algal biofuels still lags behind due to high cost of investment towards energy-intensive biomass harvesting techniques like centrifugation, membrane filtration and chemical based flocculation from photobioreactors [4,5,6]. Chemical processes are highly efficient, but the requirement of a lot of flocculant dosages renders the harvested biomass contaminated with undesirable chemicals [7, 8]. Biologically induced harvesting of algal cells is now being explored as a replacement for the conventional algal dewatering processes which contribute towards 3–15% of the algal biomass production cost [9, 10]. The harvested biomass can be processed for biofuel generation without any loss of quality [11, 12]. There have been quite a few studies on the algal–algal and algal–bacterial interactions for bio-harvesting of algal cells from bulk media [13,14,15,16,17,18,19,20]. A very recent approach is the use of filamentous fungi for harvesting algal cells [21,22,23]. In spite of having the potential for becoming a cost-effective process for algal harvesting, the lack of knowledge regarding the causative factors for the algal attachment to fungal pellets limits its application.
When fungus is grown under submerged conditions with agitation, it forms a dense, compact hyphal structure termed as pellets [24]. When these pellet-forming filamentous fungi (PFF) were co-cultivated with algal cells, there is an orderly attachment of algal cells on the fungal pellets [22, 23, 25]. This phenomenon of algal–fungal attachment is of interest as it solves the problem of algal harvesting to a large extent. However, the studies with co-cultivation of fungi and algae have reported the interaction time to be between 24 and 72 h [22, 25,26,27,28,29,30,31]. In a more straight-forward approach, it has been observed that the attachment of algal cells (green algae or cyanobacteria) to fungal pellets could take place within 4–6 h using pre-cultivated fungal biomass [12, 32]. Such interaction of algae and PFF [12] is intriguing as it does not follow the simple kinetics of bio-harvesting [13, 33, 34]. Our recent study confirms that in addition to a specific set of physical conditions, the metabolically active fungus is mandatory for efficient attachment of the Chlorella pyrenoidosa cells to the Aspergillus fumigatus pellet [12]. This fact raises further queries on the biological context of C. pyrenoidosa–A. fumigatus attachment, which although seems far simpler than the complex algal–fungal associations in nature.
A complex symbiotic association between fungus and cyanobacteria exists in the form of lichens in natural systems [35]. In artificial systems, such as co-culturing of fungus (Aspergillus nidulans) with microalgae (Chlamydomonas), a mutualistic association for nutrient exchange has been reported as reflected by thinning of microalgal cell wall by enzymatic action of fungus. However, this mutualism may sometimes lead to antagonism, when the algal death occurs due to over secretions of these enzymes [36]. Relatively simple phytoplankton–parasitic fungal (Chytrids) interactions controlled by cell-to-cell contact also exist in nature, which are driven by chemotaxis [37]. In our recent studies, somewhat similar action of fungus was observed in algal–fungal pellets as fungus used algal cells as nutrient source by enzymatically degrading it [32]. However, what drives the interaction between C. pyrenoidosa and A. fumigatus at the molecular level is not clear. There are also no specific molecules reported for PFFs so far, although enough evidence for extracellular polymeric substances (EPS)-mediated aggregation exists [38,39,40,41].
The present study aims to find out the causative mechanism behind this interesting phenomenon. It also tries to answer questions like why a specific set of biological conditions is necessary for this process to occur. Experiments were designed to address several questions like is there any chemical signaling/mediating molecule which mediates the attachment of C. pyrenoidosa and A. fumigatus, what is the origin of this mediating molecule, i.e., C. pyrenoidosa or A. fumigatus, what is the type/nature of this mediating molecule, how does this molecule mediate the attachment, etc. However, this study raises further questions regarding the nature and type of receptors which may be responsible for the attachment process that needs to be investigated further.
Results
Role of extracellular metabolites in C. pyrenoidosa–A. fumigatus harvesting
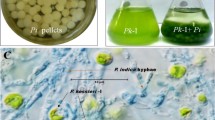
When A. fumigatus pellets and C. pyrenoidosa cells, suspended in respective spent medium (Fig. 1a), were mixed at 1:5 ratio, C. pyrenoidosa harvesting efficiency of 90% was observed after 4 h (Fig. 1b). This A. fumigatus–C. pyrenoidosa ratio and its harvesting efficiency were optimized in our earlier study [12] and has been referred to as control for all the experiments performed in the present study. To pinpoint the specific mediating molecule, i.e., whether the cell structure of C. pyrenoidosa, A. fumigatus or any of its extracellular metabolic by-product is mediating the attachment, several experiments were performed by eliminating the C. pyrenoidosa or A. fumigatus spent medium from the C. pyrenoidosa–A. fumigatus mixture. When unwashed C. pyrenoidosa cells were mixed with washed A. fumigatus pellets (Set II), the harvesting efficiency dropped to 20% compared to > 90% in the control (Set I). Interestingly, when washed A. fumigatus pellets were re-suspended in A. fumigatus spent medium (Set III), harvesting efficiency was similar to the control (Fig. 1b). However, when washed A. fumigatus pellets were resuspended in fresh PDB (Set IV), the harvesting efficiency was similar to that of washed A. fumigatus pellets, i.e., ≤ 20%. On the other hand, when washed C. pyrenoidosa cells were subjected to harvesting experiment with unwashed A. fumigatus pellets (Set V), > 90% harvesting efficiency was observed. C. pyrenoidosa culture without any A. fumigatus biomass showed ≤ 5% harvesting. Overall, significant reduction in harvesting efficiencies was seen (p < 0.05) when A. fumigatus spent medium was washed off. Therefore, impact of A. fumigatus spent medium on C. pyrenoidosa cells was explored further.

a Schematic representations of experimental conditions in different experimental sets. b Harvesting efficiency of BG11 grown Chlorella pyrenoidosa and A. fumigatus pellets as per different experimental conditions (defined in 1a) after 4 h (n = 3). Significant drop in harvesting efficiency is seen in Set II and Set IV (p < 0.05)
Nature and characterization of the mediating molecule
To identify the nature of this extracellular molecule, different physical (heat) and chemical treatments (periodate, solvent extraction) of A. fumigatus spent medium were done before harvesting C. pyrenoidosa cells with untreated washed A. fumigatus pellets. The autoclaved A. fumigatus spent medium showed > 90% harvesting in 4 h. The aqueous phase of A. fumigatus spent medium after methanol: chloroform (1:1) extraction showed ≤ 20% harvesting efficiency. On the other hand, aqueous phase after hexane extraction exhibited > 90% harvesting efficiency. Periodate treatment of A. fumigatus spent medium leads to ≤ 20% harvesting efficiency. Harvesting efficiency of C. pyrenoidosa using A. fumigatus pellets with untreated spent was ≥ 90% (positive control). Harvesting efficiency of C. pyrenoidosa with washed A. fumigatus pellets (negative control) was ≤ 20% (Fig. 2).

Harvesting efficiency of C. pyrenoidosa cells by A. fumigatus pellets after different treatments of A. fumigatus spent medium. Significant drop in flocculation efficiency is seen after periodate and methanol–chloroform treatment (p < 0.05) which is similar to the negative control (washed A. fumigatus pellets) without any A. fumigatus spent medium
The composition of A. fumigatus spent medium was studied using HR-LC–MS. The flow-through obtained using Strata-X CW column showed no loss of harvesting efficiency. Hence, the HR-LC–MS studies of this flow-through liquid were done. The results indicated the presence of two sugar-like molecules in the spent medium, i.e., N-acetyl-d-glucosamine (GlcNAc) and glucose (Table 1). HR-LC–MS of the supernatant of washed A. fumigatus pellets (without spent medium) did not show the presence of GlcNAc or glucose.
Confirmation of the mediating molecule in C. pyrenoidosa–A. fumigatus interaction
Various treatments of A. fumigatus spent medium indicated sugar-like molecule to be responsible for harvesting. Hence, experiments were conducted to study the role of glucose and GlcNAc as a replacement of A. fumigatus spent medium during C. pyrenoidosa–A. fumigatus harvesting. Glucose, being the simplest form of sugar, was tested first. C. pyrenoidosa cells were incubated with 100-mM glucose and then harvesting experiments were carried out using washed A. fumigatus pellets devoid of any A. fumigatus spent medium. Results showed > 75% harvesting after 5 h, suggesting that glucose assisted in the harvesting process. Negative control of C. pyrenoidosa incubated without glucose did not show any harvesting with washed A. fumigatus pellets (Additional file 1: Figure S1).
The concentration of glucose in the supernatant at different stages of incubation and harvesting was analyzed using HPLC (Fig. 3). HPLC analysis showed the presence of glucose before and after incubation with C. pyrenoidosa cells. No other peak was observed in these spectra. However, after mixing with A. fumigatus pellets and harvesting for 5 h, no glucose was detected by HPLC, but 24 mM of GlcNAc was detected. This suggested that the conversion of glucose into GlcNAc took place during harvesting with A. fumigatus pellets. During HPLC, GlcNAc had a retention time of 8.7 min; while glucose had a retention time of 8.3 min. This was an interesting observation as after incubating C. pyrenoidosa with glucose, the end product after harvesting was not glucose but GlcNAc. HR-LC–MS analysis also confirmed the presence of GlcNAc after 5 h of harvesting the glucose-incubated C. pyrenoidosa cells (Fig. 3a–d). Therefore, the exact role of GlcNAc during harvesting was studied in further experiments. C. pyrenoidosa cells were incubated with 100-mM GlcNAc for 2 h and harvested with washed A. fumigatus pellets for 5 h. GlcNAc concentration was measured by HPLC during three stages of the experiments viz. initial, after 2-h incubation and after harvesting. The initial concentration of GlcNAc was 90.40 mM which decreased to 36 mM after incubation with C. pyrenoidosa. When the GlcNAc-incubated C. pyrenoidosa cells were subjected for harvesting with washed A. fumigatus pellets, harvesting efficiency was found to be > 85% and GlcNAc concentration was measured as 28 mM after 5 h. On the contrary, there was no GlcNAc detected in C. pyrenoidosa incubated without glucose/GlcNAc and subjected to harvesting with washed A. fumigatus pellets (Additional file 2: Table S1). No harvesting occurred in this case. However, when t test was performed between the harvesting efficiencies of C. pyrenoidosa cells incubated with glucose and GlcNAc, significant difference in harvesting efficiency was observed till 4 h. The difference was not significant after 4 h (Additional file 1: Figure S1).

HPLC chromatograms of supernatant from glucose pre-incubated C. pyrenoidosa at different stages of incubation. a Initial glucose concentration (RT-8.3 min) of 94.36 mM. b Glucose concentration after 2 h of pre-incubation showing a reduced concentration up to 45 mM. b After harvesting, the peak shifted to RT-8.7 which corresponds to the peak of GlcNAc, depicting the formation of GlcNAc during the attachment process. d HR-LC–MS chromatogram of the supernatant after harvesting glucose pre-incubated C. pyrenoidosa and washed A. fumigatus pellets confirming the presence of GlcNAc (indicated by arrow)
Microscopic analysis and surface characterization of A. fumigatus spent medium-/GlcNAc-incubated C. pyrenoidosa cells during harvesting
HR-LC–MS and HPLC analyses suggested that washed A. fumigatus pellets could harvest C. pyrenoidosa cells in the presence of GlcNAc alone. This indicates that GlCNAc present in the A. fumigatus spent media is responsible for C. pyrenoidosa harvesting. To further investigate this observation, Con A staining of normal C. pyrenoidosa cells, A. fumigatus spent medium-incubated C. pyrenoidosa cells and GlcNAc-incubated C. pyrenoidosa cells was done. Results showed the absence of GlcNAc on the cell surface of normal cells. However, when the C. pyrenoidosa cells were incubated with A. fumigatus spent medium or GlcNAc, the presence of GlcNAc on its cell surface was indicated by dark red dots (Fig. 4a). Further, FTIR analysis of same samples was performed. The FTIR pattern of the A. fumigatus spent medium- and GlcNAc-incubated algae showed distinct peaks at 3430, 2922, 2131, 1657, 1564, 1380, 1319, 1161, 1079, and 894 cm−1. These peaks were found to be characteristic of GlcNAc when correlated with FTIR spectra of GlcNAc alone. No such peaks were observed in normal C. pyrenoidosa cells. The FTIR of C. pyrenoidosa alone showed the presence of phosphate (1048 cm−1), amide (1541 cm−1), carboxylic (1650 cm−1), alkenes (2919 cm−1) and hydroxyl (3300 cm−1) groups on its cell surface (Fig. 4b) [12].

a Representative brightfield and fluorescent micrographs of Concanavalin A stained (i and ii) normal C. pyrenoidosa, (iii and iv) C. pyrenoidosa cells incubated with A. fumigatus spent medium and (v and vi) C. pyrenoidosa cells incubated with GlcNAc. White arrows indicate the presence of GlcNAc on the C. pyrenoidosa cell surface giving bright red spots. b FTIR spectra of normal C. pyrenoidosa, A. fumigatus spent media-incubated C. pyrenoidosa, GlcNAc-incubated C. pyrenoidosa and GlcNAc powder alone. Arrows indicate the characteristic peaks of GlcNAc in spent medium-incubated as well as GlcNAc-incubated C. pyrenoidosa biomass
A distinct change in surface nature of C. pyrenoidosa cells incubated with A. fumigatus spent medium as well as GlcNAc for 2.5 h was observed through microscopic analysis (SEM and HR-TEM). The SEM image clearly showed more wrinkles on the C. pyrenoidosa cells after incubation as compared to the normal un-incubated C. pyrenoidosa cells (Fig. 5). The spent medium-incubated C. pyrenoidosa cells were elongated compared to the normal C. pyrenoidosa cells and appeared to be embedded in a matrix. Similar changes were observed when C. pyrenoidosa cells were incubated with GlcNAc. On the other hand, the SEM of the C. pyrenoidosa cells incubated with washed A. fumigatus pellets (which did not induce attachment) was found to be similar to the normal C. pyrenoidosa cells (Fig. 5).

Scanning electron micrographs of (a) normal C. pyrenoidosa (b) C. pyrenoidosa incubated with washed A. fumigatus pellets (2.5 h) (c) C. pyrenoidosa incubated with A. fumigatus spent medium (2.5 h) (d) C. pyrenoidosa incubated with GlcNAc (2.5 h). The images shown above are the representative images for each treatment selected out of multiple frames. The figure shows the change in surface morphology of C. pyrenoidosa cells after incubating with A. fumigatus spent medium and GlcNAc while no such change in C. pyrenoidosa cells incubated with washed A. fumigatus pellets (2.5 h)
To study the ultrastructural changes, HR-TEM of the above samples was performed. The normal C. pyrenoidosa cells and cell incubated with washed A. fumigatus (without A. fumigatus spent medium/GlcNAc) showed intact and round cells having smooth cell wall and well-defined cell organelles (Fig. 6). TEM images of A. fumigatus spent medium- and GlcNAc-incubated C. pyrenoidosa cells showed identical morphological changes. Both the images showed the formation of small villi-like structures on the cell surface. However, the TEM images of C. pyrenoidosa cells incubated with washed A. fumigatus pellets did not show such morphological changes and were similar to that of normal C. pyrenoidosa cells. The changes in the C. pyrenoidosa surface after A. fumigatus spent medium/GlcNAc treatment or even after complete harvesting does not affect the integrity of the cell wall as depicted by SYTOX green staining. The red auto-fluorescence was observed in A. fumigatus spent medium-incubated C. pyrenoidosa cells indicating live cells. As SYTOX binds to nucleic acids, dead cells would have shown green fluorescent color (Additional file 3: Figure S2).

Transmission electron micrographs of (a) normal C. pyrenoidosa cells (b) C. pyrenoidosa incubated with washed A. fumigatus pellets (2.5 h) (c) C. pyrenoidosa incubated with A. fumigatus spent medium (2.5 h) (d) C. pyrenoidosa incubated with GlcNAc (2.5 h); CW denotes the cell wall of algal cells. The arrows show the formation of villi-like structures on the cell wall after incubation with A. fumigatus spent medium/GlcNAc. The images shown above are the representative images for each treatment selected out of multiple frames
To confirm the surface changes observed through SEM and HR-TEM, AFM analysis of normal C. pyrenoidosa cells and A. fumigatus spent medium-incubated C. pyrenoidosa cells was conducted. Increase in the roughness of the C. pyrenoidosa cells (RMS value of 91.0 nm) was seen when incubated with A. fumigatus spent medium as compared to the normal C. pyrenoidosa which showed RMS value of 76.5 nm (Fig. 7). The cell height also increased from 42.5 nm in normal C. pyrenoidosa to 53.8 nm in the A. fumigatus spent medium-incubated C. pyrenoidosa.

AFM analysis of (a) normal C. pyrenoidosa (RMS 76.5 nm) (b) spent medium treated C. pyrenoidosa (91.0 nm) showing change in the roughness of the cells after incubation with A. fumigatus spent medium
Discussion
Chemical harvesting, although quick, contaminates the biomass with unnecessary chemical contaminants [42]. Previous reports have suggested that biological harvesting of micro-algae improves the quality of the biomass for biofuel purpose. Wrede et al. [30] reported that there is a significant increase in lipid yield when microalgal cells were harvested with A. fumigatus. Increase in lipid content for algal–fungal pellets was also reported by Bhattacharya et al. [12]. In another study, Botryococcus braunii was harvested using A. fumigatus and the resultant biomass did not show any significant variation in the biomass composition. Apart from biodiesel aspect, harvesting of microalgae using filamentous fungi also increases its biogas potential. Prajapati et al. [32] reported enhanced bio-methane production from algal–fungal pellets due to the enzymatic degradation of microalgal cell wall by the fungi. All these studies show that harvesting algae with filamentous fungi is a favorable process for biofuel production. However, as the underlying mechanism of the process is not clearly understood, it is difficult to replicate the process at large scale. This study gives an insight into the communication mechanism between A. fumigatus and C. pyrenoidosa which could be exploited for fungal-assisted microalgal harvesting processes at large.
Chemical communication between microbial cells is an inherent property which dictates the attachment and aggregation of cells [41, 43]. Such communications mediated by secretion of extracellular chemical signaling molecules could impose inter-beneficial partnerships for both the species [44]. In natural systems, the algae and fungus exhibit a host–parasite relationship as seen in chytrid–phytoplankton association [37]. In artificial systems, the co-culturing of Chlamydomonas reinhardtii and A. nidulans strains has been reported to show obligate mutualism in terms of nitrogen and carbon exchange within the species. The fungus converts glucose into CO2, which is being used by algae for its growth. On the other hand, the nitrogen fixing Chlamydomonas sp. reduces nitrite to ammonia, which fungus can use as a nitrogen source [36, 45]. During this nutrient exchange, the cell wall of Chlamydomonas sp. attached to fungus was reported to show thinning due to the action of fungal remodeling enzymes [45].
The present study can be closely related to the above-mentioned cases as it also reports extracellular secretions from A. fumigatus, mediating cell wall changes in the C. pyrenoidosa cells. These secretions might help A. fumigatus to provide signal to the C. pyrenoidosa cell and bring it into close vicinity for nutritional benefits or enzymatic degradation [32]. Similar phenomenon of microalgal–fungal attachment has been reported by our group with wide range of microalgal species encompassing green algae (Chlorella sp.), blue–green algae (Chroococcus sp.) as well as mixed consortia [12, 32].
Fungi produce various exopolysaccharides during its growth [38], which have been known to mediate cell–cell adhesion. The present study has identified an extracellular causative factor produced by the fungus A. fumigatus which is responsible for the C. pyrenoidosa–A. fumigatus attachment. Experiments done with washed and unwashed A. fumigatus pellets have shown the presence of a mediating molecule in the A. fumigatus spent medium that is crucial for the attachment process. Washed A. fumigatus pellets could not induce attachment since they were devoid of any A. fumigatus spent medium. Fresh PDB also did not have the mediating molecule, and its harvesting efficiency harvesting was similar to the washed A. fumigatus pellets. This observation suggested that the causative factor is the extracellular metabolite produced by actively growing fungi. The result was also in agreement with our recent observations that relatively old (72-h grown) or autoclaved A. fumigatus pellets failed to harvest the C. pyrenoidosa cells due to their low metabolic activity and damaged hyphae [12]. A. fumigatus spent medium is a cocktail of extracellular metabolites including EPS which is a complex mixture of polysaccharides, proteins, nucleic acids and amyloids [40]. EPS production by Aspergillus sp. is reported for bio-flocculation [46]. However, to pin-point the causative factor, it is necessary to know the nature of the mediating molecule.
The autoclaving of the A. fumigatus spent medium did not affect the harvesting efficiency of the fungus. This indicated that the mediating factor was not proteinaceous in nature as autoclaving would denature the protein. Chloroform and methanol mixture was able to extract the polar organic components from the A. fumigatus spent medium. Loss of activity after this treatment indicated that the mediating factor was a polar organic compound. This was further confirmed using hexane as a solvent for extraction. Treatment of the A. fumigatus spent medium with hexane did not show any loss in harvesting activity. As hexane is able to extract only non-polar organic compounds, it further confirmed that the molecule is a polar organic compound. Further insights into the nature of the molecule were obtained by treating the A. fumigatus spent medium with sodium per-iodate. Per-iodate targets any saccharide/polysaccharide leading to its oxidation. Loss of harvesting activity after per-iodate treatment of the A. fumigatus spent medium strongly suggested that the mediating factor was a sugar-like molecule. The HR-LC–MS analysis of the A. fumigatus spent medium also showed the presence of two sugars, glucose and GlcNAc which might be the triggering factors for the attachment process. Previously, the bioflocculant produced by Achromobacter xylosoxidans was found to be composed mainly of carbohydrate hetero polymer [47]. In another study, characterization of a bioflocculant produced by Aspergillus flavus showed that it contained 69.7% sugar of which 1.8% was amino sugar-like GlcNAc [46]. In a recent study, it has been found that an amino sugar galactosaminogalactan mediates attachment of A. fumigatus to epithelial cells [48].
In the present study, we were able to pinpoint the particular molecule that could replicate the C. pyrenoidosa–A. fumigatus attachment process in the absence of A. fumigatus spent medium. Results of several spent treatments corroborated with HR-LC–MS results suggested the target molecule to be glucose or GlcNAc. The presence of either GlcNAc or A. fumigatus spent medium is mandatory for the attachment process. The t test between harvesting kinetics of control and GlcNAc-incubated C. pyrenoidosa cells did not show any significant difference. When GlcNAc was used for harvesting studies, it was clearly seen that the harvesting process was significantly faster as compared to glucose. HR-LC–MS and HPLC results confirmed that glucose is converted into GlcNAc by A. fumigatus pellets. Conversions of sugar-like molecules into GlcNAc have been reported by saprophytic fungus using degrading/hydrolytic enzymes [49, 50]. Kinetics of glucosamine formation using glucose and other sugars as substrate by fungus like Aspergillus sp. has also been reported during submerged fermentation [51]. Similar harvesting experiments with glucose anomers (Galactose and Mannose) showed only 30% harvesting which further suggests the role of GlcNAc in the harvesting process since these glucose anomers cannot be converted into GlcNAc (Additional file 4: Figure S3). The present result explains the previously observed pre-requisites for C. pyrenoidosa–A. fumigatus attachment (the presence of metabolically active fungi, temperature of 38 °C and neutral pH) [12], which seem to favor GlcNAc production. Earlier report showed that GlcNAc production by Aspergillus sp. was dependent on the pH of the system, where higher pH inhibited its formation [51]. Comparative study of GlcNAc production among three wild-type fungi viz. A. fumigatus, Rhizopus oligosporus and Monascus pilosus showed that A. fumigatus had the highest production capacity among the three fungi.
We could demonstrate the presence of GlcNAc on the spent medium-incubated and GlcNAc-incubated C. pyrenoidosa using the lectin Concanavalin A (Con A) stain, and FTIR. Although, Con A gives signal for glucose as well as glucosamine, C. pyrenoidosa suspended in BG11 only do not show any signal as seen in the Fig. 4a. When C. pyrenoidosa is incubated with A. fumigatus spent media or glucosamine, Con A shows signals on the algal cell surface (Fig. 4b, c). When C. pyrenoidosa cells were incubated in the presence of glucosamine alone, the concentration of glucosamine decreases over time. The FTIR pattern of glucosamine-incubated cells show peaks similar to glucosamine. The Con A staining results complement the findings of the FTIR data. The mechanism of interaction could be further elaborated by examining the effect of A. fumigatus spent medium on C. pyrenoidosa cell’s morphology, ultrastructure and cell surface roughness. Roughness analysis is one of the parameters which indicates the changes in the cell surface due to changes in the growing environment. The root mean square (rms) value corresponds to the roughness of the sample. AFM analysis showed that the roughness of the A. fumigatus spent medium-incubated C. pyrenoidosa cells was increased compared to the normal C. pyrenoidosa cells. Increase in roughness is an essential step for the cellular attachment process [52]. It has also been reported that in case of a green fouling alga Enteromorpha, increased roughness of cells caused more fouling compared to smooth cells with low roughness [53]. SEM micrographs (Fig. 5c) of spent medium-incubated cells depicted elongated C. pyrenoidosa cells embedded in a sticky matrix. Extracellular release of mucilage for entrapment is well documented for parasitic type of A. fumigatus species [40].
The fact that both A. fumigatus spent medium-incubated C. pyrenoidosa and GlcNAc-incubated algae showed the presence of GlcNAc-specific peaks in the FTIR spectra suggests that similar cell surface changes may be induced by A. fumigatus spent medium and GlcNAc. In this connection, SEM micrographs of GlcNAc-incubated C. pyrenoidosa cells also showed elongation and wrinkles on the cell surface. TEM images clearly show that A. fumigatus spent medium induces the formation of villi-like structures on C. pyrenoidosa surface. It was interesting to note that similar villi-like structures were induced by incubating C. pyrenoidosa cells with GlcNAc. On the other hand, washed A. fumigatus pellets (without A. fumigatus spent medium) failed to induce such changes. The above results strongly suggest that GlcNAc present in A. fumigatus spent medium is responsible for inducing surface changes in C. pyrenoidosa cells that mediates the attachment to A. fumigatus hyphae .
Role of GlcNAc has also been demonstrated for cell–cell adhesion in bacteria and yeast [54, 55]. Moreover, it has also been established that GlcNAc can function as a signaling molecule or as an inducer of hyphal growth in Candida albicans without undergoing a metabolic reaction [56]. It is also known to induce formation of curly fibers in bacteria [57]. In the present study, we demonstrate that GlcNAc plays a similar role in adhesion of C. pyrenoidosa cells to A. fumigatus pellets by inducing morphological changes on the C. pyrenoidosa cell surface which is not reported till now. In summary, we show that the C. pyrenoidosa attachment to A. fumigatus pellets is mediated by GlcNAc as depicted by biochemical and analytical methods. Based on the results, the probable mechanism of the process is depicted in Fig. 8. When the C. pyrenoidosa cells are incubated with A. fumigatus spent medium or GlcNAc, it attaches with the C. pyrenoidosa surface. This induces surface modifications in C. pyrenoidosa cells as the cells become elongated with development of projections on the surface. The GlcNAc molecule on C. pyrenoidosa might act as a chemical signal for A. fumigatus receptors, thereby attaching the C. pyrenoidosa cells onto A. fumigatus cell wall. Recent reports have shown the role of G protein-coupled receptors (G-PCRs) present in ascomycetes fungi for sensing sugars [58]. Also, it has been reported that the receptors on the A. fumigatus cell surface are dynamic and are made only in the presence of external stimuli [59]. This kind of receptors might play a role in the C. pyrenoidosa–A. fumigatus attachment for which metabolically active fungi are required. Our results indicate that this attachment is triggered by GlcNAc which may act like a quorum-sensing molecule. GlcNAc induces morphological changes on the C. pyrenoidosa cell surface which is clearly visible by TEM micrograph. However, our recent study with such microalgal–fungal attachment has shown that fungus uses this interaction process to ultimately utilize the microalgal cells as source of food [32]. Hence, this microbial interaction is different from other interactions where the emitter and receiver of the signal are both benefited [44]. The existence of A. fumigatus receptors and factors triggering their expression needs to be investigated further.

The figure depicts the probable mechanism of C. pyrenoidosa–A. fumigatus interaction mediated by GlcNAc (mediating molecule)
Methods
Organism and culture conditions
Microalgal species Chlorella pyrenoidosa was obtained from National Collection of Industrial Microorganisms (NCIM), NCL Pune (India). C. pyrenoidosa was maintained in 2% algae culture agar (HiMedia M343-500G) slants supplemented with BG11 (HiMedia, M1541-500G) in a plant growth chamber (Daihan Labtech, LGC-5101). Liquid cultures were maintained in BG11 broth at 120 rpm without CO2 supplementation. For experimental purposes, C. pyrenoidosa culture was grown in 2.5-L flasks under continuous light in a greenhouse maintained at 25 ± 1 °C and a light intensity of ≈ 3500 Lx [12]. The fungal strain Aspergillus fumigatus (Accession no. KY241789), previously isolated from wastewater [60], was used as the harvesting organism. A. fumigatus was maintained on sterile potato dextrose agar (PDA) slants (Himedia M096-500G) at 28 ± 1 °C.
C. pyrenoidosa and A. fumigatus harvesting experiments
Aspergillus fumigatus slant (3 days old) was used to inoculate potato dextrose broth (PDB; Himedia M403-500G) medium for cultivation. A. fumigatus spore suspension prepared with 0.1% Tween-80 (≈ 108 spores ml−1) was inoculated in 100-ml potato dextrose broth and incubated for 24 h at 28 ± 1 °C and 150 rpm in an orbital shaker. C. pyrenoidosa cultures exhibiting optical density (OD680) ≈ 2.5 were used for harvesting experiments. Ten milliliters of overnight-grown A. fumigatus culture was added to 90 ml of C. pyrenoidosa culture having OD680 ≈ 2.5 (1:5 dry weight basis) and kept at 38 ± 1 °C and 100 rpm for 4 h in an incubator shaker. The harvesting efficiency based on the optical density of C. pyrenoidosa was measured after every 30 min, for which the flask was allowed to stand for 3 min followed by drawing of the sample for absorbance measurement (OD680) using a microplate reader (Biotek EON®-C). Harvesting efficiency (HE) was calculated as:
where ODt = optical density at time t and OD0 = initial optical density.
To check the viability of normal C. pyrenoidosa cells and C. pyrenoidosa cells incubated with A. fumigatus spent medium during harvesting (2.5 h), the cells were stained with a nucleic acid stain, i.e., SYTOX green (Cat. No. S7020, Invitrogen), which specifically stains the dead or damaged cells. Following this, dual fluorescence was used to show red autofluorescence for live C. pyrenoidosa cells and green fluorescence of SYTOX green (Excitation/Emission: 504/523 nm) for dead cells [61].
Role of extracellular factors
To assess the role of an extracellular substance for the attachment process, A. fumigatus biomass was washed twice with distilled water removing all the spent medium from it. The A. fumigatus medium present after the growth of the fungus has been termed as A. fumigatus spent medium throughout the text. The A. fumigatus biomass was subjected to 5 harvesting experimental sets viz: Set I—Control (unwashed C. pyrenoidosa and A. fumigatus pellets), Set II—Unwashed C. pyrenoidosa and washed A. fumigatus pellets, Set III—Unwashed C. pyrenoidosa and washed A. fumigatus pellets resuspended in its spent medium, Set IV—Unwashed C. pyrenoidosa and washed A. fumigatus pellets resuspended in fresh PDB, Set V—Washed C. pyrenoidosa and unwashed A. fumigatus pellets. Harvesting of all the above sets was performed similarly as described above for 4 h. Experiments were done in triplicates for each and every experimental set. C. pyrenoidosa culture without any A. fumigatus biomass was run as a negative control to check the sedimentation rate of C. pyrenoidosa culture. The experimental setup is graphically shown in Fig. 1a.
To study the effect of an extracellular factor (present in A. fumigatus spent medium) on the C. pyrenoidosa cell surface, scanning electron microscopy (SEM) and High-Resolution Transmission electron microscopy (HR-TEM) were performed before harvesting (normal C. pyrenoidosa cells) and after designated time during harvesting (2.5 h). For comparison, the SEM and HR-TEM of C. pyrenoidosa cells incubated with washed A. fumigatus pellets (without A. fumigatus spent medium) from Set II were also performed. The atomic force microscopy (AFM) of normal C. pyrenoidosa cells and cells during harvesting was performed to evaluate the change in cell height and roughness.
Treatment and characterization of A. fumigatus spent medium
By the above preliminary experiments, the role of some extracellular factors presents in the A. fumigatus spent medium was highlighted. Hence, various treatments of A. fumigatus spent medium like autoclaving, sodium periodate treatment, Methanol: Chloroform extraction and hexane extraction were performed followed by testing the harvesting efficiency of these pre-treated A. fumigatus spent medium when added to washed A. fumigatus pellets and C. pyrenoidosa cells. For every experiment, A. fumigatus pellets were first washed to remove any spent medium present. Harvesting experiments were done with normal C. pyrenoidosa cells and A. fumigatus pellets resuspended in fungal spent medium after treatments (autoclaving, sodium periodate treatment, Methanol: Chloroform extraction and hexane extraction). A mixture of C. pyrenoidosa cells and A. fumigatus pellets with untreated spent was used as a positive control. A negative control comprising C. pyrenoidosa mixed with washed A. fumigatus pellets was run to ensure that the pellets of A. fumigatus did not have the fungal spent media.
Aspergillus fumigatus spent medium was autoclaved for 15 min at 121 °C and 15 psi pressure before using for harvesting experiments. As autoclaving would denature any protein component in the spent A. fumigatus medium, the role of protein molecule (if any) could be found out. Sodium periodate targets any saccharide/polysaccharide leading to the oxidation of these residues. The spent A. fumigatus media (100 ml) were pre-treated with 20-mM sodium periodate (Cat No. S1147, Sigma) solution prepared in an oxidation buffer (50-mM sodium acetate and 50-mM acetic acid, pH-4.5). Since sodium periodate is light sensitive, the preparation of the solution and pre-treatment of A. fumigatus spent medium were performed in the dark at 28 °C and 150 rpm. To stop the activity of sodium periodate, the sample was exposed to light for 15 min to remove the remaining periodate in the system.
The methanol: chloroform extraction was performed by mixing an equal volume of methanol: chloroform mixture (1:1) with the A. fumigatus spent medium for 1 h at 28 °C and 150 rpm to identify any polar molecules responsible for the harvesting process. The aqueous fraction was separated from the mixture by a separating funnel and the organic fraction was discarded. Washed A. fumigatus pellets suspended in this aqueous fraction were then subjected to harvesting of C. pyrenoidosa cells. A similar experiment was done using the hexane-extracted aqueous fraction (extracted following the similar protocol used for Methanol: Chloroform aqueous extract) to identify the presence of any non-polar molecule which aided in the harvesting process. The A. fumigatus spent medium was analyzed using high-resolution liquid chromatography–mass spectrometry (HR-LC–MS).
Determination of mediating molecule
Based on the HR-LC–MS analysis and preceding A. fumigatus spent medium treatments, glucose and N-acetyl glucosamine (GlcNAc) were suspected to play the most important role in harvesting process. C. pyrenoidosa biomass was exposed to 100-mM glucose and GlcNAc, respectively, by incubation for 2 h at 38 °C and 100 rpm, followed by removal of residual glucose and GlcNAc by centrifugation (4000g for 10 min) and resuspension of biomass in fresh BG11. C. pyrenoidosa after glucose/GlcNAc incubation was then subjected to harvesting with washed A. fumigatus pellets (without spent medium) for 5 h as described in “C. pyrenoidosa and A. fumigatus harvesting experiments” section. Another set of C. pyrenoidosa suspension was also incubated under the same conditions without supplementation of glucose/GlcNAc followed by its harvesting with washed A. fumigatus pellets. To analyze the concentration of glucose and GlcNAc at different stages of experiment, high-performance liquid chromatography (HPLC) was performed at three levels: (i) supernatant of C. pyrenoidosa suspension immediately after glucose or GlcNAc addition (initial); (ii) supernatant of C. pyrenoidosa suspension after 2 h of incubation with glucose or GlcNAc and (iii) supernatant of respective sets at the end of harvesting for 5 h. The results were compared with HPLC of glucose and GlcNAc standard as a reference. HR-LC–MS of the supernatants from all the set after harvesting was also done to confirm the result of HPLC.
GlcNAc-incubated C. pyrenoidosa biomass was also subjected to Fourier Transform Infra-Red Spectroscopy (FTIR) and was compared to FTIR spectra of C. pyrenoidosa biomass during harvesting (after 2 h) as well as normal C. pyrenoidosa. The FTIR of GlcNAc powder was also performed to correlate the presence of similar peaks on the incubated C. pyrenoidosa biomass. To further confirm the role of GlcNAc and A. fumigatus spent medium in the harvesting process, C. pyrenoidosa cells incubated with GlcNAc (100 mM) and A. fumigatus spent medium was observed under a fluorescent microscope and high-resolution transmission electron microscope (HR-TEM, Sect. 2.7.5) and was compared with normal C. pyrenoidosa cells. For fluorescent microscopy, the cells were stained with Concanavalin A (Con A) conjugated with Alexa Fluor® 594 (Cat No. C11253, ThermoFisher Scientific) to detect the presence of GlcNAc on the C. pyrenoidosa cell surface [62], followed by viewing of cells under a fluorescent microscope (Nikon Eclipse Ti-U). Con A is a lectin which binds d-glucose, d-fructose, d-mannose, N-acetyl-d-glucosamine and related monosaccharides [63]. Since Con A does not have fluorescence, it is tagged with a fluorescent dye Alexa Fluor 594 (Excitation/Emission: 590/617 nm). To 100 µl of C. pyrenoidosa cells, 0.1 µl of the dye was added and kept in the dark for 5 min at room temperature. The stained cells were then observed under the microscope.
Analytical techniques
Sample preparation for high-performance liquid chromatography (HPLC) and high-resolution liquid chromatography–mass spectrometry (HR-LC–MS)
Samples were made salt free by passing through polymeric cartridge Strata-X-CW (Cat No. 8B-S035-JEG, Phenomenex). The cartridge was first conditioned by adding 10-ml methanol followed by 10-ml distilled water. The flow-through was discarded, and then 10 ml of the sample was added to the cartridge. The flow-through from the cartridge was collected, tested for harvesting activity and then analyzed using HPLC and HR-LC–MS.
HPLC
Standards for glucose (Cat No. 47829) and GlcNAc (Cat No. PHR1432-1G) were purchased from Sigma. Standards were dissolved in HPLC grade water (Cat No. AS077-1L, Hi-Media). HPLC was performed according to manufacturer’s protocol using Agilent 1260 series machine with Agilent Hi-Plex H column and RI detector. The mobile phase was 0.005-M H2SO4 (Cat No. 5438270100, Sigma) at a flow rate of 0.6 ml min−1. The column temperature was kept at 60 °C and the run time was 20 min. All samples were degassed prior to analysis.
HR-LC–MS
HR-LC–MS was done to identify the nature of the compounds present in the A. fumigatus spent medium. The A. fumigatus spent medium is a mixture of various types of compounds like proteins, carbohydrates, organic acids, sugars and A. fumigatus metabolites. Hence, HR-LC–MS was done for these compounds. The aliquot was subjected to HR-LC–MS analysis (ACCUCORE RP-MS), and was outsourced to Sophisticated Analytical Instrument Facility (SAIF), Indian Institute of Technology (IIT), Mumbai. The column used was ZORBAX ECLIPSE C-18 (Agilent Technologies). The sample was run isocratically for 30 min using acetonitrile (95%) as a solvent. The compounds were analyzed using Quadrupole—Time of Flight mass spectrometer (Q-TOF MS; Agilent iFunnel G6550A) giving the probable hits from the database library provided by the manufacturer.
Scanning electron microscopy (SEM)
SEM analysis of (i) normal C. pyrenoidosa cells, (ii) C. pyrenoidosa cells incubated with A. fumigatus spent medium, (iii) GlcNAc-incubated C. pyrenoidosa and (iv) washed A. fumigatus pellets were done using a previously described protocol [61]. The samples for SEM analysis were first washed with PBS and then fixed with 1% glutaraldehyde for 4 h at room temperature. The samples were centrifuged and the fixative was discarded. Samples were then lyophilized (Allied Frost FD3) for SEM analysis using a ZEISS EVO 50 instrument under the following analytical condition: EHT = 20.00 kV, WD = 9.5 mm, Signal A = SE1.
High-resolution Transmission electron microscopy (HR-TEM)
The C. pyrenoidosa cells ((i) normal C. pyrenoidosa cells, (ii) C. pyrenoidosa cells incubated with A. fumigatus spent medium, (iii) GlcNAc-incubated C. pyrenoidosa and (iv) washed A. fumigatus pellets) were centrifuged at 4000g for 10 min and prepared for viewing as outlined by Gola et al. [64]. The samples were examined by high-resolution transmission electron microscope (Tecnai G2 20) operated at 200 kV.
Atomic force microscopy (AFM)
To confirm the observations of TEM analysis, AFM studies of the C. pyrenoidosa cells (normal cells and A. fumigatus spent medium-incubated C. pyrenoidosa cells) were done. C. pyrenoidosa cells were centrifuged and suspended in 50-mM citrate–phosphate buffer (pH 3) for conditioning. The buffer was removed by centrifugation, and the pellets were kept on glass cover slip for 30 min followed by washing with de-ionized water and then air drying. AFM micrographs were obtained using Bruker INNOVAA2 Sys instrument in tapping mode.
Fourier transform infrared spectroscopy (FTIR)
For FTIR measurements, the samples were first washed with phosphate buffer saline (PBS) and then lyophilized. The lyophilized powder was then used for FTIR analysis using a Nicolet Is50 (Thermo Scientific) instrument.
Statistical analysis
All experiments were performed in triplicates and results were represented as mean ± S.D wherever applicable. Graphs were drawn using Microsoft Excel® (Part of Microsoft Office 2013 package). Significance test has been done using one-way ANOVA (p < 0.05). t Test (two-tailed) was also performed to check the significance between two data sets.
Availability of data and materials
Data supporting the results of the article are included within this manuscript and additional information.
Abbreviations
- AF:
-
Aspergillus fumigatus
- AFM:
-
atomic force microscopy
- CP:
-
Chlorella pyrenoidosa
- FTIR:
-
Fourier transform infrared spectroscopy
- GlcNAc:
-
N-acetyl glucosamine
- HPLC:
-
high-performance liquid chromatography
- HR-LC–MS:
-
high-resolution liquid chromatography–mass spectrometry
- OD:
-
optical density
- PDA:
-
potato dextrose agar
- PDB:
-
potato dextrose broth
- SEM:
-
scanning electron microscopy
- TEM:
-
transmission electron microscopy
References
Sharma YC, Singh B, Korstad J. A critical review on recent methods used for economically viable and eco-friendly development of microalgae as a potential feedstock for synthesis of biodiesel. Green Chem. 2011;13:2993.
Ngangkham M, Ratha SK, Prasanna R, Saxena AK, Dhar DW, Sarika C, et al. Biochemical modulation of growth, lipid quality and productivity in mixotrophic cultures of Chlorella sorokiniana. SpringerPlus. 2012;1:33. https://doi.org/10.1186/2193-1801-1-33.
Tango MD, Calijuri ML, Assemany PP, Couto E. Microalgae cultivation in agro-industrial effluents for biodiesel application: effects of the availability of nutrients. Water Sci Technol. 2018;78:2018180. https://doi.org/10.2166/wst.2018.180.
Molina Grima E, Belarbi EH, Acién Fernández FG, Robles Medina A, Chisti Y. Recovery of microalgal biomass and metabolites: process options and economics. Biotechnol Adv. 2003;20:491–515.
Gomez JA, Höffner K, Barton PI. From sugars to biodiesel using microalgae and yeast. Green Chem. 2015;18:461–75.
Rashid N, Rehman MSU, Han J-I. Use of chitosan acid solutions to improve separation efficiency for harvesting of the microalga Chlorella vulgaris. Chem Eng J. 2013;226:238–42.
Şirin S, Trobajo R, Ibanez C, Salvadó J. Harvesting the microalgae Phaeodactylum tricornutum with polyaluminum chloride, aluminium sulphate, chitosan and alkalinity-induced flocculation. J Appl Phycol. 2012;24:1067–80. https://doi.org/10.1007/s10811-011-9736-6.
Granados MR, Acién FG, Gómez C, Fernández-Sevilla JM, Molina Grima E. Evaluation of flocculants for the recovery of freshwater microalgae. Bioresour Technol. 2012;118:102–10.
Fasaei F, Bitter JH, Slegers PM, van Boxtel AJB. Techno-economic evaluation of microalgae harvesting and dewatering systems. Algal Res. 2018;31:347–62.
Vasconcelos Fernandes T, Shrestha R, Sui Y, Papini G, Zeeman G, Vet LEM, et al. Closing domestic nutrient cycles using microalgae. Environ Sci Technol. 2015;49:12450–6. https://doi.org/10.1021/acs.est.5b02858.
Al-Hothaly KA, Adetutu EM, Taha M, Fabbri D, Lorenzetti C, Conti R, et al. Bio-harvesting and pyrolysis of the microalgae Botryococcus braunii. Bioresour Technol. 2015;191:117–23. https://doi.org/10.1016/j.biortech.2015.04.113.
Bhattacharya A, Mathur M, Kumar P, Prajapati SK, Malik A. A rapid method for fungal assisted algal flocculation: critical parameters and mechanism insights. Algal Res. 2017;21:42–51.
Salim S, Gilissen L, Rinzema A, Vermuë MH, Wijffels RH. Modeling microalgal flocculation and sedimentation. Bioresour Technol. 2013;144:602–7.
Sukenik A, Shelef G. Algal autoflocculation—verification and proposed mechanism. Biotechnol Bioeng. 1984;26:142–7.
Oh HM, Lee SJ, Park MH, Kim HS, Kim HC, Yoon JH, et al. Harvesting of Chlorella vulgaris using a bioflocculant from Paenibacillus sp. AM49. Biotechnol Lett. 2001;23:1229–34. https://doi.org/10.1023/a:1010577319771.
Rodolfi L, Zittelli GC, Barsanti L, Rosati G, Tredici MR. Growth medium recycling in Nannochloropsis sp. mass cultivation. Biomol Eng. 2003;20:243–8.
Vandamme D, Foubert I, Fraeye I, Muylaert K. Influence of organic matter generated by Chlorella vulgaris on five different modes of flocculation. Bioresour Technol. 2012;124:508–11.
Cho K, Hur SP, Lee CH, Ko K, Lee YJ, Kim KN, et al. Bioflocculation of the oceanic microalga Dunaliella salina by the bloom-forming dinoflagellate Heterocapsa circularisquama, and its effect on biodiesel properties of the biomass. Bioresour Technol. 2016;202:257–61.
Hu YR, Wang F, Wang SK, Liu CZ, Guo C. Efficient harvesting of marine microalgae Nannochloropsis maritima using magnetic nanoparticles. Bioresour Technol. 2013;138:387–90.
Zhang B, Lens PNL, Shi W, Zhang R, Zhang Z, Guo Y, et al. Enhancement of aerobic granulation and nutrient removal by an algal–bacterial consortium in a lab-scale photobioreactor. Chem Eng J. 2018;334:2373–82.
Xu L, Guo C, Wang F, Zheng S, Liu CZ. A simple and rapid harvesting method for microalgae by in situ magnetic separation. Bioresour Technol. 2011;102:10047–51.
Xie S, Sun S, Dai SY, Yuan J. Efficient coagulation of microalgae in cultures with filamentous fungi. Algal Res. 2013;2:28–33.
Zhang J, Hu B. A novel method to harvest microalgae via co-culture of filamentous fungi to form cell pellets. Bioresour Technol. 2012;114:529–35.
Papagianni M. Fungal morphology and metabolite production in submerged mycelial processes. Biotechnol Adv. 2004;22:189–259.
Zhou W, Cheng Y, Li Y, Wan Y, Liu Y, Lin X, et al. Novel fungal pelletization-assisted technology for algae harvesting and wastewater treatment. Appl Biochem Biotechnol. 2012;167:214–28. https://doi.org/10.1007/s12010-012-9667-y.
Gultom SO, Zamalloa C, Hu B. Microalgae harvest through fungal pelletization—co-culture of Chlorella vulgaris and Aspergillus niger. Energies. 2014;7:4417–29.
Miranda AF, Taha M, Wrede D, Morrison P, Ball AS, Stevenson T, et al. Lipid production in association of filamentous fungi with genetically modified cyanobacterial cells. Biotechnol Biofuels. 2015;8:179.
Muradov N, Taha M, Miranda AF, Wrede D, Kadali K, Gujar A, et al. Fungal-assisted algal flocculation: application in wastewater treatment and biofuel production. Biotechnol Biofuels. 2015;8:24.
Zhou W, Min M, Hu B, Ma X, Liu Y, Wang Q, et al. Filamentous fungi assisted bio-flocculation: a novel alternative technique for harvesting heterotrophic and autotrophic microalgal cells. Sep Purif Technol. 2013;107:158–65.
Wrede D, Taha M, Miranda AF, Kadali K, Stevenson T, Ball AS, et al. Co-cultivation of fungal and microalgal cells as an efficient system for harvesting microalgal cells, lipid production and wastewater treatment. PLoS ONE. 2014;9:e113497.
Prajapati SK, Kumar P, Malik A, Choudhary P. Exploring pellet forming filamentous fungi as tool for harvesting non-flocculating unicellular microalgae. Bioenergy Res. 2014;7:1430–40. https://doi.org/10.1007/s12155-014-9481-1.
Prajapati SK, Bhattacharya A, Kumar P, Malik A, Vijay VK. A method for simultaneous bioflocculation and pretreatment of algal biomass targeting improved methane production. Green Chem. 2016;18:5230–8.
Stratford M, Keenan MHJ. Yeast flocculation: kinetics and collision theory. Yeast. 1987;3:201–6.
Pan G, Zhang MM, Chen H, Zou H, Yan H. Removal of cyanobacterial blooms in Taihu Lake using local soils. I. Equilibrium and kinetic screening on the flocculation of Microcystis aeruginosa using commercially available clays and minerals. Environ Pollut. 2006;141:195–200.
Sanders WB. Lichens: the Interface between mycology and plant morphology. Source Biosci. 2001;51:1025–36.
Hom EFY, Schaeme D, Mittag M, Sasso S. A chemical perspective on microalgal—microbial interactions. Trends Plant Sci. 2015;10:1–4.
Frenken T, Alacid E, Berger SA, Bourne EC, Gerphagnon M, Grossart H-P, et al. Integrating chytrid fungal parasites into plankton ecology: research gaps and needs. Environ Microbiol. 2017;19:3802–22. https://doi.org/10.1111/1462-2920.13827.
Debeaupuis JP, Sarfati J, Goris A, Stynen D, Diaquin M, Latgé JP. Exocellular Polysaccharides from Aspergillus Fumigatus and Related Taxa. Mod Concepts Penicillium Aspergillus Classif. 1990;209:23. https://doi.org/10.1007/978-1-4899-3579-3_18.
Holder DJ, Keyhani NO. Adhesion of the entomopathogenic fungus Beauveria (Cordyceps) bassiana to Substrata. Appl Environ Microbiol. 2005;71:5260–6.
Jones EBG. Fungal adhesion. Mycol Res. 1994;98:961–81.
Mori JF, Ueberschaar N, Lu S, Cooper RE, Pohnert G, Küsel K. Sticking together: inter-species aggregation of bacteria isolated from iron snow is controlled by chemical signaling. ISME J. 2017;11:1075–86.
Chen L, Wang C, Wang W, Wei J. Optimal conditions of different flocculation methods for harvesting Scenedesmus sp. Cultivated in an open-pond system. Bioresour Technol. 2013;133:9–15.
Phuong K, Kakii K, Nikata T. Intergeneric coaggregation of non-flocculating Acinetobacter spp. isolates with other sludge-constituting bacteria. J Biosci Bioeng. 2009;107:394–400.
Keller L, Surette MG. Communication in bacteria: an ecological and evolutionary perspective. Nat Rev Microbiol. 2006;4:249–58.
Hom EFY, Murray AW. Niche engineering demonstrates a latent capacity for fungal-algal mutualism. Science (80 −). 2014;345:94–8.
Aljuboori AHR, Idris A, Abdullah N, Mohamad R. Production and characterization of a bioflocculant produced by Aspergillus flavus. Bioresour Technol. 2013;127:489–93.
Subudhi S, Bisht V, Batta N, Pathak M, Devi A, Lal B. Purification and characterization of exopolysaccharide bioflocculant produced by heavy metal resistant Achromobacter xylosoxidans. Carbohydr Polym. 2016;137:441–51.
Gravelat FN, Beauvais A, Liu H, Lee MJ, Snarr BD, Chen D, et al. Aspergillus galactosaminogalactan mediates adherence to host constituents and conceals hyphal β-glucan from the immune system. PLoS Pathog. 2013;9:e1003575. https://doi.org/10.1371/journal.ppat.1003575.
Contreras R, Maras M, Bruyn A, Vervecken W, Uusitalo J, Penttil M, et al. In vivo synthesis of complex N-glycans by expression of human N-acetylglucosaminyltransferase I in the filamentous fungus Trichoderma reesei. FEBS Lett. 1999;452:365–70.
Steiger MG, Mach-Aigner AR, Gorsche R, Rosenberg EE, Mihovilovic MD, Mach RL. Synthesis of an antiviral drug precursor from chitin using a saprophyte as a whole-cell catalyst. Microb Cell Fact. 2011;10:1–9.
Hsieh J-W, Wu H-S, Wei Y-H, Wang SS. Determination and kinetics of producing glucosamine using fungi. Biotechnol Prog. 2007;1:1. https://doi.org/10.1021/bp070037o.
Hallab NJ, Bundy KJ, O’Connor K, Moses RL, Jacobs JJ. Evaluation of metallic and polymeric biomaterial surface energy and surface roughness characteristics for directed cell adhesion. Tissue Eng. 2001;7:55–71. https://doi.org/10.1089/107632700300003297.
Granhag L, Finlay J, Jonsson P, Callow J, Callow M. Roughness-dependent removal of settled spores of the green alga Ulva (syn Enteromorpha) exposed to hydrodynamic forces from a water jet. Biofouling. 2004;20:117–22. https://doi.org/10.1080/08927010410001715482.
Prasadarao NV, Wass CA, Kim KS. Endothelial cell GlcNAc beta 1-4GlcNAc epitopes for outer membrane protein A enhance traversal of Escherichia coli across the blood–brain barrier. Infect Immun. 1996;64:154–60.
Cormack BP, Ghori N, Falkow S. An adhesin of the yeast pathogen Candida glabrata mediating adherence to human epithelial cells. Science. 1999;285:578–82.
Naseem S, Gunasekera A, Araya E, Konopka JB. N-Acetylglucosamine (GlcNAc) induction of hyphal morphogenesis and transcriptional responses in Candida albicans are not dependent on its metabolism. J Biol Chem. 2011;286(33):28671–80.
Barnhart MM, Lynem J, Chapman MR. GlcNAc-6P levels modulate the expression of curli fibers by Escherichia coli. J Bacteriol. 2006;188:5212–9.
Xue C, Hsueh YP, Heitman J. Magnificent seven: roles of G protein-coupled receptors in extracellular sensing in fungi. FEMS Microbiol Rev. 2008;32:1010–32.
Grice CM, Bertuzzi M, Bignell EM. Receptor-mediated signaling in Aspergillus fumigatus. Front Microbiol. 2013;4:26.
Mathur M, Gola D, Panja R, Malik A, Ahammad SZ. Performance evaluation of two Aspergillus spp. for the decolourization of reactive dyes by bioaccumulation and biosorption. Environ Sci Pollut Res. 2018;25:345–52. https://doi.org/10.1007/s11356-017-0417-0.
Prajapati SK, Bhattacharya A, Malik A, Vijay VK. Pretreatment of algal biomass using fungal crude enzymes. Algal Res. 2015;8:8–14.
Nobile CJ, Fox EP, Hartooni N, Mitchell KF, Hnisz D, Andes DR, et al. A histone deacetylase complex mediates biofilm dispersal and drug resistance in Candida albicans. MBio. 2014;5:e01201.
Gonçalves GRF, Gandolfi ORR, Santos CMS, Bonomo RCF, Veloso CM, Fontan R. Development of supermacroporous monolithic adsorbents for purifying lectins by affinity with sugars. J Chromatogr B. 2016;1033–1034:406–12.
Gola D, Dey P, Bhattacharya A, Mishra A, Malik A, Namburath M, et al. Multiple heavy metal removal using an entomopathogenic fungi Beauveria bassiana. Bioresour Technol. 2016;218:388–96. https://doi.org/10.1016/j.biortech.2016.06.096.
Acknowledgements
The authors are grateful to Nano Research Facility (NRF) and Central Research Facility (CRF), IIT Delhi, for help with HPLC, FTIR, AFM,SEM and HR-TEM analysis. Sophisticated Analytical Instrument Facility (SAIF), IIT Bombay, for HR-LCMS analysis and SAIF AIIMS, New Delhi for TEM section cutting.
Funding
The present work was carried out with financial support from Science and Engineering Research Board (SERB), Department of Science and Technology, Government of India (SB/S3/CEE/0002/2014). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
AB and AM designed the experiments. AB, MM and PK conducted the experiments. AB and MM have written the manuscript. AM has thoroughly scrutinized the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable for this study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1: Figure S1.
Harvesting kinetics of C. pyrenoidosa with A. fumigatus pellets after incubation with glucose and GlcNAc showing significant difference in process kinetics (p < 0.05) up to 4 h. Non-significant difference (p < 0.05) was observed between harvesting kinetics of control and GlcNAc.
Additional file 2: Table S1.
HR-LC–MS analysis of supernatants after harvesting of algal–fungal mixtures using algae with different pre-incubations.
Additional file 3: Figure S2.
Representative brightfield (left) and fluorescent (right) micrographs of SYTOX Green stained C. pyrenoidosa cells incubatedwith A. fumigatus spent medium.
Additional file 4: Figure S3.
Harvesting efficiency of C. pyrenoidosa cells with A. fumigatus when incubated with different sugars.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Bhattacharya, A., Mathur, M., Kumar, P. et al. Potential role of N-acetyl glucosamine in Aspergillus fumigatus-assisted Chlorella pyrenoidosa harvesting. Biotechnol Biofuels 12, 178 (2019). https://doi.org/10.1186/s13068-019-1519-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13068-019-1519-3