
Abstract
Background
The molecular mechanisms underlying the association between increased adiposity and aggressive breast cancer phenotypes remain unclear, but likely involve the adipokines, leptin (LEP) and adiponectin (ADIPOQ), and their receptors (LEPR, ADIPOR1, ADIPOR2).
Methods
We used immunohistochemistry (IHC) to assess LEP, LEPR, ADIPOQ, ADIPOR1, and ADIPOR2 expression in breast tumor tissue microarrays among a sample of 720 women recently diagnosed with breast cancer (540 of whom self-identified as Black). We scored IHC expression quantitatively, using digital pathology analysis. We abstracted data on tumor grade, tumor size, tumor stage, lymph node status, Ki67, estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor 2 (HER2) from pathology records, and used ER, PR, and HER2 expression data to classify breast cancer subtype. We used multivariable mixed effects models to estimate associations of IHC expression with tumor clinicopathology, in the overall sample and separately among Blacks.
Results
Larger proportions of Black than White women were overweight or obese and had more aggressive tumor features. Older age, Black race, postmenopausal status, and higher body mass index were associated with higher LEPR IHC expression. In multivariable models, lower LEPR IHC expression was associated with ER-negative status and triple-negative subtype (P < 0.0001) in the overall sample and among Black women only. LEP, ADIPOQ, ADIPOR1, and ADIPOR2 IHC expression were not significantly associated with breast tumor clinicopathology.
Conclusions
Lower LEPR IHC expression within the breast tumor microenvironment might contribute mechanistically to inter-individual variation in aggressive breast cancer clinicopathology, particularly ER-negative status and triple-negative subtype.
Similar content being viewed by others


Background
In the United States (US), Black women have higher breast cancer incidence at a younger age (e.g., ≤ 45 years) and are more frequently diagnosed with cancers exhibiting more aggressive phenotypes compared to their White counterparts [1,2,3]. US cancer incidence data during 2008 and 2012 show that Black women have the lowest incidence of breast cancers characterized as hormone receptor-positive (positive for estrogen receptor [ER] and/or progesterone receptor [PR]) and human epidermal growth factor receptor 2 (HER2)-negative and the highest incidence of hormone receptor-negative and HER2-negative breast cancers when compared to other racial/ethnic groups [1]. Furthermore, breast cancer mortality among Black women continues to be higher than among White women by at least 41% [1, 3]. Given that Black women also tend to have higher age-adjusted prevalence rates of obesity (approximately 57% among non-Hispanic Black vs. 40% among non-Hispanic White women [4]), we hypothesize that inter-individual variation in adiposity and adiposity-related biomarkers (namely leptin [LEP], leptin receptor [LEPR], adiponectin [ADIPOQ], adiponectin receptor 1 [ADIPOR1], and adiponectin receptor 2 [ADIPOR2]) within the breast tumor microenvironment might be involved in the observed racial differences in aggressive breast cancer phenotypes between Black and White women.
Increased levels of adiposity are consistently associated with elevated risk of postmenopausal breast cancer [5, 6] and poorer breast cancer outcomes, irrespective of menopausal status [6,7,8]. However, the molecular mechanisms underlying these associations are not well understood. The relationship between adiposity and breast cancer might be explained partly by biological effects of the adipokines, ADIPOQ, and LEP, which are secreted by adipocytes [9,10,11,12,13,14]. Epidemiologic data have shown an inverse association of circulating ADIPOQ levels with increasing body mass index (BMI) [15,16,17] and breast cancer risk [18,19,20,21]. Conversely, circulating LEP levels are positively associated with BMI [22, 23] and also breast cancer risk in some studies [18, 24,25,26].
ADIPOQ is the most abundantly produced hormone in adipose tissue [16, 27] and, with its receptors (ADIPOR1 and ADIPOR2), is expressed in both histologically normal and malignant breast tissues [28, 29]. ADIPOQ has anti-inflammatory and anti-atherogenic properties [27, 30], and is known to inhibit cellular proliferation and to promote apoptosis [11, 14, 31], implying a protective role in breast carcinogenesis and progression. ADIPOQ expression is downregulated by elevated adiposity, glucocorticoids, β-adrenergic agonists, and TNF-α, and upregulated by leanness, cold exposure, adrenalectomy, and IGF-1 [32, 33]. LEP, also produced in adipose tissues, is similarly expressed in histologically normal and malignant breast tissues, as is the LEPR [34, 35]. LEP is a key growth factor and may be involved in breast carcinogenesis and progression by promoting cell growth, proliferation, and angiogenesis [36,37,38,39]. Once bound to LEPR, LEP induces the activation of several signaling pathways (including Janus kinase-signal transducer and activator of transcription [JAK/STAT], mitogen-activated protein kinase [MAPK], insulin receptor substrate 1 [IRS1], and suppressor of cytokine signaling 3 [SOCS3] [9, 36, 40]) that modulate cellular proliferation and survival [9], and these activities have been demonstrated in breast epithelial cells [36, 40]. In summary, ADIPOQ and LEP act on breast epithelial tissues through endocrine pathways as well as locally through autocrine and/or paracrine pathways [28, 41], and, along with their receptors, likely contribute to breast carcinogenesis, progression, and aggressiveness through these mechanisms.
The objective of this study was to determine whether immunohistochemical (IHC) expression of LEP, LEPR, ADIPOQ, ADIPOR1, and ADIPOR2 are associated with breast tumor clinicopathological characteristics, namely those that are indicative of aggressive phenotypes among Black and White women newly diagnosed with breast cancer, including poor differentiation (higher tumor grade), larger tumor size, positive lymph node status, unfavorable Ki67 status (Ki67+), ER− status, HER2+ status, and non-luminal HER2-enriched (HER2-E) and triple-negative (TN) subtypes.
Methods
Study sample and data collection
In this study, we conducted a case-only analysis that included 720 incident, primary invasive breast cancer cases diagnosed from 2001 to 2015 and enrolled in the Women’s Circle of Health Study (WCHS). The study design of WCHS is described elsewhere [42]. Briefly, newly diagnosed breast cancer cases with histologically confirmed ductal carcinoma in situ (DCIS, stage 0) or invasive breast cancer (stages I–IV), who self-identified as either Black/African American or White/European American, were 20–75 years of age, able to complete an interview in English, and had no history of cancer except non-melanoma skin cancer, were eligible to participate. Data collection for the WCHS was conducted through in-person assessments and included interviewer-administered questionnaires as well as anthropometric and body composition measurements [43]. The baseline interview ascertained information on sociodemographics, as well as established or probable breast cancer risk factors, including family and personal health history, reproductive history, hormone therapy use, and lifestyle exposures. Anthropometric measurements (height, weight, waist and hip circumference measures) and body composition measures (lean and fat mass, percent body fat) were also taken at the in-person, baseline interview using standardized protocols and instruments [43].
Upon consent for medical records release, medical and pathology records from all providers and institutions where WCHS participants reported receiving breast cancer care were retrieved [44]. Trained abstractors reviewed and abstracted relevant data from each record and entered data into an electronic database [44]. For quality assurance, values were checked for errors during data entry, and if errors were detected, the original abstractor was contacted with instructions to re-check the medical records/pathology report, allowing for confirmation of the recorded data. For the present analysis, we utilized abstracted data on tumor clinicopathology. Tumor grade was defined as grades 1 through 3: grade 1 denoted well-differentiated tumors, grade 2 denoted moderately differentiated tumors, and grade 3 denoted poorly differentiated tumors. Tumor size (cm) was classified into three categories: < 1.0 cm, 1.0–2.0 cm, and > 2.0 cm. American Joint Committee on Cancer (AJCC) stage data was recorded as stages 0 through IV; we considered four categories in the main analysis, stage 0, I, II, and a combined category that included stages III and IV. Lymph node status was defined as node negative or node positive, based on the presence of cancer cells in axillary lymph nodes. Ki67 staining was classified as Ki67+/unfavorable, Ki67 borderline, or Ki67−/favorable (due to the lack of abstraction of clinically relevant percentages of Ki67 staining). Ki67 status was then dichotomized where cases coded as Ki67−/favorable or borderline were classified as having “favorable” Ki67 status, while cases coded as Ki67+ were classified as having “unfavorable” Ki67 status. We used surrogate classifications of ER status, PR status, and HER2 status, which were based on IHC expression of ER and PR, and overexpression of HER2 (by IHC and/or fluorescence in situ hybridization [FISH]). Using these classifications, we approximated breast cancer subtype into four mutually exclusive, clinically recognized subtypes: luminal A (ER+ and/or PR+/HER2−), luminal B (ER+ and/or PR+/HER2+), HER2-E (ER−/PR−/HER2+), and TN (ER−/PR−/HER2−).
This study was approved by the Institutional Review Boards of all participating institutions, and all study participants provided written informed consent prior to study enrollment.
Collection of archived breast tumor specimens and tissue microarray construction
In the WCHS, tumor blocks and/or slides were collected from hospitals after written consent was obtained from study participants. The retrieval rate of archived breast tumor specimens was approximately 85% in the study. Upon receipt at the Data Bank and BioRepository (DBBR) at Roswell Park Comprehensive Cancer Center, a study pathologist (TK) reviewed the hematoxylin and eosin (H&E) slides and circled areas where cores were taken for tissue microarrays (TMAs). TMA cores ranged in size from 0.6 to 1.2 μm in diameter, and most WCHS participants’ tumors were represented by at least two TMA cores (range, 1 to 6 cores), which were placed into a recipient formalin-fixed paraffin-embedded (FFPE) block. The location of each core was recorded in a detailed TMA map file. The completed TMAs were stored at room temperature.
Immunohistochemistry
All IHC staining was performed using Ventana Discovery XT Automated Slide Stainer (Ventana Medical Systems, Inc., Tucson, AZ, USA). Deparaffinization, antigen retrieval, blocking, DAB detection, counterstain, post-counterstain, and slide cleaning steps were automated on the Discovery XT. Primary antibodies and secondary antibodies were manually applied at programmed steps. The following primary antibodies were used: rabbit monoclonal OB (leptin) antibody (1:40 dilution; Santa Cruz, cat #sc-842), mouse monoclonal Ob-R (leptin receptor) antibody (1:25 dilution; Santa Cruz, cat #sc-8391), mouse monoclonal adiponectin antibody (1:30 dilution; Abcam, cat #ab22554), rabbit monoclonal adiponectin receptor 1 antibody (1:350 dilution; Abcam, cat #ab126611), and goat polyclonal adiponectin receptor 2 antibody (1:25 dilution; Abcam, cat #ab77612). Optimal staining on control slides (human breast tissue TMAs) was obtained for each individual antibody. IHC was then performed using the optimized conditions on the experimental TMA slides constructed from WCHS specimens as well as on additional control slides. Primary antibodies were incubated at 37 °C for 1–2 h; secondary antibodies were incubated at 37 °C for 1 h, followed by either the DAB Map Detection Kit (Ventana, 760-124) or ChromoMap DAB kit (Ventana, 760-159). Slides were counterstained with hematoxylin (Ventana, 760-2021) and bluing reagent (Ventana, 760-2037) before cover slipping.
TMA specimens stained for LEP, LEPR, ADIPOQ, ADIPOR1, and ADIPOR2 were digitized at × 20 on an Olympus VS120 whole slide scanner (Olympus Corporations, Central Valley, PA, USA). Examples of the resultant IHC staining for each biomarker on WCHS TMA specimens are shown in Fig. 1. TMA registration was performed on the software platform TMA-AID, as previously described [45], to correlate each study participant’s unique identification number with the corresponding imaged tissue cores and protein expression information. A digital pathology analysis platform (VisioPharm, Hoersholm Denmark) was used to build a custom workflow and perform quantitative analysis of IHC expression on each tissue core. Quantitative IHC expression results were reported as effective staining intensity (ESI) within the effective staining area (ESA) [45]. Specimen artifacts such as tissue folding were manually excluded from quantification. TMA-AID was also used by a board-certified pathologist (MC) to evaluate IHC expression for LEP and LEPR of each tissue core stained. Semi-quantitative expression results were reported as follows: 0 (negative), 1 (weak expression), 2 (moderate expression), or 3 (strong expression). We confirmed through Pearson’s correlation analysis that there were relatively high concordance rates between the automated/unsupervised (quantitative) and pathologist-generated (semi-quantitative) scores for LEP (r = 0.70; P < 0.0001) and LEPR (r = 0.71, P < 0.0001); therefore, for all five biomarkers examined, only the quantitative data were included in the analysis.

These images represent relatively average immunohistochemical expression of the adipokines and adipokine receptors in invasive breast tumor tissues. The specimens shown in a through e are imaged at × 20 magnification and show LEP (a), LEPR (b), ADIPOQ (c), ADIPOR1 (d), and ADIPOR2 (e) immunoreactivity in invasive breast tumor tissues. The images in f through j are further magnified images of each core shown in a through e. TMA cores were mostly 0.6 μm (with a range of 0.6 to 1.2 μm). Presence of granular cytoplasmic pattern of staining of the adipokines and adipokine receptors in the breast tumor tissues ranged in intensity from weakly positive (shown in images d and e—and the corresponding magnified images i and j) to moderate and strong (shown in images a–c—and the corresponding magnified images f–h)
Statistical analysis
Select sociodemographics, breast tumor clinicopathologic characteristics, and IHC expression of LEP, LEPR, ADIPOQ, ADIPOR1, and ADIPOR2 were described overall and by race using means (± standard deviations [SDs]) and frequencies (proportions) for continuous and categorical variables, respectively. Student’s t tests and analysis of variance (ANOVA) were used to test IHC expression of each biomarker by select characteristics. Pearson’s correlation analysis was used to assess pairwise correlations of adipokine and adipokine receptor IHC expression, overall and separately by race. Multivariate mixed effects models were used to describe the associations of breast tumor clinicopathologic features with adipokine and adipokine receptor IHC expression in order to incorporate the correlation among the adipokine and adipokine receptor IHC expression. Differences of least squares means were generated using multivariate mixed effects models controlling for race, BMI, and menopausal status. We decided a priori (based on background knowledge and a review of the literature) to control for these factors in the multivariate mixed effects models. In addition, each model was mutually controlled for the IHC expression of the other four biomarkers. Subsequent analysis focused on assessing the multivariate associations (controlling for BMI and menopausal status) between adipokine and adipokine receptor IHC expression with breast tumor clinicopathologic features among Black WCHS participants only.
Analyses were performed using SAS (v9.4 SAS Institute, Cary, NC). All reported P values are two-sided, and P < 0.05 was considered statistically significant. To address concerns of multiple comparisons in the multivariate mixed effects models assessing associations between adipokine IHC expression and tumor features, the Bonferroni correction was used, with a criterion for statistical significance of P < 0.003 (i.e., 0.05/15).
Results
Select characteristics of the study sample
Select characteristics of the WCHS participants included in the analysis are shown in Table 1, overall and with comparisons by race. The proportion of Black women in the sample is larger than the proportion of White women (540 Black vs. 159 White) because the study stopped recruiting White breast cancer cases in 2012, while enrollment of new Black breast cancer cases continued (through 2020). The current analysis included White cases diagnosed in 2001 through 2009 and Black cases diagnosed in 2001 through 2015, who had tumor specimens in the TMAs constructed by the start of the IHC analysis. Mean age at diagnosis was 52.6 ± 10.8 years, more than half of the sample was postmenopausal, and mean BMI was 30.7 ± 7.0 kg/m2. Several differences were observed by race: larger proportions of Black women were overweight or obese and had breast tumor features indicative of more aggressive phenotypes (e.g., tumors that were poorly differentiated, ER−, TN subtype) compared to White women.
Summary of adipokine and adipokine receptor IHC expression in WCHS
Distributions of IHC expression of each biomarker by select characteristics are shown in Table 2. Some notable findings were that LEPR IHC expression was higher among postmenopausal women compared to premenopausal women and increased with increasing BMI. We also found that IHC expression of LEPR, ADIPOQ, and ADIPOR2 was significantly higher among Black women than White women, which was likely attributable to the finding that Black women were more frequently postmenopausal and overweight or obese. Variation in LEPR IHC expression was observed by breast tumor clinicopathologic features, while the expression of LEP, ADIPOQ, ADIPOR1, and ADIPOR2 did not appear to vary by tumor clinicopathology. Among Black women, older age, postmenopausal status, and increasing BMI were associated with higher LEPR IHC expression, and higher LEPR IHC expression was associated with Ki67−/favorable status, ER+ status, PR− status, and luminal A and luminal B subtypes (see Additional file 1). Among White women, only increasing BMI was associated with higher LEPR IHC expression, and higher LEPR IHC expression was associated with larger tumor size, positive lymph node status, and ER− status (see Additional file 2).
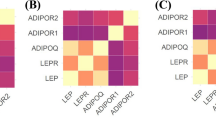
Figure 2 depicts the pairwise Pearson correlation matrix for IHC expression of LEP, LEPR, ADIPOQ, ADIPOR1, and ADIPOR2 in the overall sample (Fig. 2a), and separately among Black (Fig. 2b) and White study participants (Fig. 2c). There were significant positive correlations, although weak (r < 0.25), for LEP IHC expression with LEPR, ADIPOQ, ADIPOR1, and ADIPOR2 IHC expression. There was a moderate positive correlation for ADIPOQ with ADIPOR1 (r = 0.59) and a weak-to-moderate positive correlation for ADIOPQ with ADIPOR2 (r = 0.34). IHC expression of ADIPOR1 and ADIPOR2 were also moderately correlated (r = 0.44). It is worth noting that IHC expression of LEPR and ADIPOR1 were not significantly correlated. These findings were generally consistent in the subgroup analyses by race.

The pairwise Pearson correlations between IHC expression of LEP, LEPR, ADIPOQ, ADIPOR1, and ADIPOR2 among the overall study sample (N = 720) (a), among Black study participants only (n = 540) (b), and among White study participants only (n = 159) (c)
Associations of adipokine and adipokine receptor IHC expression with breast cancer clinicopathology in WCHS
Table 3 shows the mixed effects models of the associations of LEP, LEPR, ADIPOQ, ADIPOR1, and ADIPOR2 IHC expression with breast tumor clinicopathologic features. LEP IHC expression was not significantly associated with any tumor clinicopathologic feature apart from breast cancer subtype. Higher LEP IHC expression was marginally associated with luminal B subtype (vs. luminal A, P = 0.05). Lower LEPR IHC expression was associated with ER− and PR− status, and HER2-E and TN subtypes. Lower ADIPOQ IHC expression was associated with larger tumor size and unfavorable Ki67 status. No statistically significant associations were observed between any tumor clinicopathologic feature and ADIPOR1 or ADIPOR2 IHC expression. After the Bonferroni correction (P < 0.003), only the observed associations of lower LEPR IHC expression with ER− status and TN subtype remained statistically significant.
Subgroup analysis of the multivariate associations of LEP, LEPR, ADIPOQ, ADIPOR1, and ADIPOR2 IHC expression with breast tumor clinicopathologic features among Black women yielded similar findings (Table 4). Some notable qualitative differences from the associations observed in the overall sample (shown in Table 3) suggested that the association of lower LEPR IHC expression with poorly differentiated tumors (vs. well differentiated) and unfavorable Ki67 status (vs. favorable) was stronger in magnitude among Black women as compared to the overall sample, while the association of lower LEPR IHC expression with PR− status was attenuated among Blacks. Additionally, the association of lower ADIPOQ IHC expression with unfavorable Ki67 status was slightly stronger, while the association of ADIPOQ IHC expression with increasing tumor size was attenuated among Black women.
Discussion
Given observed associations of increasing levels of adiposity with poorer breast cancer survival, we examined relationships between breast tumor IHC expression of adipokine biomarkers (LEP, LEPR, ADIPOQ, ADIPOR1, ADIPOR2) with tumor clinicopathological features that are associated with poorer prognosis. Our overarching hypothesis was that higher IHC expression of LEP, LEPR, ADIPOQ, ADIPOR1, and ADIPOR2 within the breast tumor microenvironment is associated with breast cancer aggressiveness, but we generally observed the opposite. While we found that Black women, postmenopausal women, and those with BMI > 30.0 kg/m2 had significantly higher LEPR IHC expression in their breast tumors, these factors were not significantly associated with IHC expression of LEP, ADIPOQ, ADIPOR1, or ADIPOR2. In multivariable models, which controlled for race, BMI, and menopausal status, we found that lower LEPR IHC was associated with ER− status and TN subtype, and these associations were particularly strong among Black women. These findings suggest that lower expression of LEPR (rather than higher expression as hypothesized) is an important indicator of more aggressive breast cancer, independent of race, BMI, and menopausal status, and might serve as an important biomarker associated with disparate outcomes, particularly among Black women. This seemingly counterintuitive observation is partially supported by data suggesting complex associations of obesity/adiposity, LEP/LEPR’s activation of various signaling pathways, and breast cancer progression, which is further complicated by ER expression [46, 47]. We hypothesize that central adiposity (rather than general obesity, as measured by BMI) is the etiologic mechanism linking LEPR expression in the breast tumor microenvironment with aggressive tumor clinicopathology and ultimately poorer prognosis. This might explain the lack of strong significant associations between BMI and IHC expression of the adipokine biomarkers examined in this study. While not the focus of the present analysis, our next steps will be to examine other anthropometric measures (e.g., waist circumference, hip circumference, waist-to-hip ratio), as well as body composition measures (percent body fat), as predictors of adipokine expression in the tumor microenvironment, with consideration to differences by ER status.
While numerous epidemiologic studies have explored and confirmed significant associations between circulating levels of the adipokines with risk of obesity-related cancers [48], fewer studies have examined the associations of IHC expression of the adipokines and their receptors with breast cancer clinicopathology [49,50,51,52,53,54,55,56,57], and most studies to date have been conducted in predominantly non-Black study samples. Findings from some of these studies suggest that LEPR expression might be downregulated in invasive tumors [50, 55] (and those with more aggressive features, including TN subtypes and unfavorable Ki67 status [52]) as compared to DCIS and normal-adjacent tissues. These observations indicate an association between LEPR downregulation and breast cancer invasion and progression. In contrast to our findings, several studies examining LEPR expression in breast cancer did not report significant associations of lower LEPR IHC expression with tumor features indicative of more aggressive phenotype [49, 51, 55, 56]. These inconsistencies could be due to differences in the analytical approach used (e.g., semi-quantitative, discrete vs. quantitative, continuous assessment of IHC expression, which directly influences the selection of statistical methodology), differences in the characteristics of the study participants examined (our analysis of adipokine IHC expression included the largest number of Black breast cancer cases to date), and/or a lack of consideration of the associations studied herein. For instance, our study explored the associations of interest using an unsupervised, digital analysis platform allowing for continuous, quantitative measurement of LEP, LEPR, ADIPOQ, ADIPOR1, and ADIPOR2 IHC expression, which are more objective. Conversely, other studies [49,50,51, 53, 55, 57] have examined IHC expression of these biomarkers using semi-quantitative methods, where discrete scores were assigned based on staining intensity (e.g., negative vs. positive, or none, mild, moderate, intense) and staining distribution (e.g., 0, < 10%; 1+, 10–50%; 2+, 50–80%; 3+, > 80%). These methods tend to yield more subjective results. Additionally, our analytic approach allowed us to control for important confounders (BMI, menopausal status, and race).
Similar to the observations reported herein, there is evidence that lower LEPR IHC is associated with clinicopathologic features indicative of tumor progression or aggressiveness in other obesity-related cancers [58,59,60,61,62,63]. A study of colorectal cancer showed that downregulation of LEPR IHC expression was associated with aggressive tumor features (namely late stage, high grade) as well as with shorter survival time [61]. Osorio and colleagues [60] showed that while there was no significant association of LEP IHC expression with prostate tumor aggressiveness, quantitative IHC expression of LEPR was significantly lower in prostate tumors exhibiting prognostic factors indicative of aggressive phenotype (namely, urethral margin involvement, surgical margin involvement, and seminal vesicle involvement). Studies of thyroid cancer [58, 59] have also shown that downregulation of LEPR was associated with increased risk of thyroid cancer recurrence and metastasis, particularly in the anaplastic thyroid cancer (ATC) subtype [59]. In endometrial cancer [62, 63], although LEP and LEPR were more highly expressed in endometrial tumor tissues than normal tissues, there was a suggestion that rates of LEPR positivity were significantly lower among poorly differentiated endometrial tumors. Notably, one study [62] showed that downregulation of the short form of LEPR was significantly associated with poor tumor differentiation in endometrial cancer. This finding indicates that there could be differences in the association of LEPR expression based on the isoform analyzed. Moreover, the complexity of the relationship of adiposity and deregulation of adipokine signaling with breast cancer (as mentioned above) might be similarly complex in other obesity/adiposity-related cancers [60, 64, 65], warranting additional research.
Our findings that lower ADIPOQ IHC expression was associated with ER− status and TN subtype, which although were not of statistical significance after the Bonferroni correction, are nonetheless worth noting. Several properties of ADIPOQ [11, 14, 27, 30, 31] imply a protective role in breast carcinogenesis. However, evidence suggests that there is a dichotomy observed in the relationship between ADIPOQ and breast cancer progression by ER status [66, 67]. In vitro studies have shown ADIPOQ stimulates growth in ER+ breast cancer cells and inhibits cellular proliferation in ER− breast cancer cells [66, 67]. So, lower IHC expression of ADIPOQ might be consistent with breast tumors progressing towards increasing aggressiveness (but only in some instances). The significant inverse associations of ADIPOQ IHC expression observed with larger tumor size and unfavorable Ki67 status also support this hypothesis. Additional research, particularly in large, diverse samples, is essential to elucidate the role of the ADIPOQ in carcinogenesis (along with the other adipokines and adipokine receptors examined herein), specifically in the tissue-specific microenvironment, which could contribute to the identification of new targets for obesity-related cancers. Moreover, as ADIPOQ exerts its effects through ADIPOR1 and ADIPOR2, more in-depth analysis of the predictors of ADIPOQ, ADIPOR1, and ADIPOR2 IHC expression in the tumor microenvironment and the impacts of their expression on tumor clinicopathology and subsequent survival outcomes, by adiposity type (e.g., overall adiposity, visceral) as well as by ER status, are critically needed.
A major strength of this study was the assessment of quantitative IHC expression and analysis of the associations of interest in a sample of breast cancer cases with well-defined clinicopathologic annotations, including > 500 Black breast cancer cases with TMA specimens and comprehensive clinicopathology data. Another strength was the relatively large number of samples included in the IHC analysis, which is one of, if not, the largest studies of adipokine and adipokine receptor IHC expression in breast cancer to date. There were also some limitations that should be considered. The first concern is the relatively small sample size of White study participants, which limited the power to detect meaningful differences in the associations of interest by race. The lack of complete and detailed clinical information on Ki67 expression was also a concern. Ki67 status was not available in the medical records of 217 participants (30.1% of the analytic sample). Unfortunately, we were also unable to retrieve the exact percentage of Ki67 staining in the medical records of those participants among whom the data were available, so we crudely classified Ki67 status as either clinically favorable (negative or borderline) or clinically unfavorable (positive) in the analysis. Despite these limitations, our findings support the hypothesis that IHC expression of the adipokines and adipokine receptors, particularly LEPR, is associated with tumor features that are indicative of more aggressive breast cancer phenotypes.
Conclusions
In summary, findings from this study suggest that lower LEPR IHC expression within the breast tumor microenvironment might serve as an indicator of increased breast tumor aggressiveness. This study focused primarily on breast cancer clinicopathologic features as a first step in assessing the association of adipokine and adipokine receptor expression on breast cancer prognosis, as breast clinicopathologic features that might be indicative of more aggressive phenotypes are likely to affect survival. Additional studies are needed to clarify the clinical implications of LEPR expression and the mechanisms involved in the regulation of LEPR expression, to ultimately determine the utility of this biomarker in understanding breast tumor aggressiveness.
Availability of data and materials
The datasets generated and analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- ADIPOQ:
-
Adiponectin
- ADIPOR1:
-
Adiponectin receptor 1
- ADIPOR2:
-
Adiponectin receptor 2
- AJCC:
-
American Joint Committee on Cancer
- ANOVA:
-
Analysis of variance
- BMI:
-
Body mass index
- DBBR:
-
Data Bank and BioRepository
- DCIS:
-
Ductal carcinoma in situ
- ER:
-
Estrogen receptor
- ESA:
-
Effective staining area
- ESI:
-
Effective staining intensity
- FFPE:
-
Formalin-fixed paraffin-embedded
- H&E:
-
Hematoxylin and eosin
- HER2:
-
Human epidermal growth factor receptor 2
- HER2-E:
-
Non-luminal HER2-enriched subtype
- IGF-1:
-
Insulin-like growth factor 1
- IHC:
-
Immunohistochemistry
- IRS1:
-
Insulin receptor substrate 1
- JAK/STAT:
-
Janus kinase-signal transducer and activator of transcription
- LEP:
-
Leptin
- LEPR:
-
Leptin receptor
- MAPK:
-
Mitogen-activated protein kinase
- PR:
-
Progesterone receptor
- SD:
-
Standard deviation
- SOCS3:
-
Suppressor of cytokine signaling 3
- TMA:
-
Tissue microarray
- TN:
-
Triple-negative subtype
- TNF-α:
-
Tumor necrosis factor alpha
- US:
-
United States
- WCHS:
-
Women’s Circle of Health Study
References
DeSantis CE, Fedewa SA, Goding Sauer A, Kramer JL, Smith RA, Jemal A. Breast cancer statistics, 2015: convergence of incidence rates between black and white women. CA Cancer J Clin. 2016;66(1):31–42.
DeSantis CE, Ma J, Goding Sauer A, Newman LA, Jemal A. Breast cancer statistics, 2017, racial disparity in mortality by state. CA Cancer J Clin. 2017;67(6):439–48.
DeSantis CE, Miller KD, Goding Sauer A, Jemal A, Siegel RL. Cancer statistics for African Americans, 2019. CA Cancer J Clin. 2019;69(3):211–233.
Flegal KM, Kruszon-Moran D, Carroll MD, Fryar CD, Ogden CL. Trends in obesity among adults in the United States, 2005 to 2014. JAMA. 2016;315(21):2284–91.
Pearson-Stuttard J, Zhou B, Kontis V, Bentham J, Gunter MJ, Ezzati M. Worldwide burden of cancer attributable to diabetes and high body-mass index: a comparative risk assessment. Lancet Diabetes Endocrinol. 2018;6(6):e6–e15.
Sung H, Siegel RL, Torre LA, Pearson-Stuttard J, Islami F, Fedewa SA, Goding Sauer A, Shuval K, Gapstur SM, Jacobs EJ, et al. Global patterns in excess body weight and the associated cancer burden. CA Cancer J Clin. 2019;69(2):88–112.
Seiler A, Chen MA, Brown RL, Fagundes CP. Obesity, dietary factors, nutrition, and breast cancer risk. Curr Breast Cancer Rep. 2018;10(1):14–27.
Chan DS, Vieira AR, Aune D, Bandera EV, Greenwood DC, McTiernan A, Navarro Rosenblatt D, Thune I, Vieira R, Norat T. Body mass index and survival in women with breast cancer-systematic literature review and meta-analysis of 82 follow-up studies. Ann Oncol. 2014;25(10):1901–14.
Cirillo D, Rachiglio AM, la Montagna R, Giordano A, Normanno N. Leptin signaling in breast cancer: an overview. J Cell Biochem. 2008;105(4):956–64.
Cleary MP, Ray A, Rogozina OP, Dogan S, Grossmann ME. Targeting the adiponectin:leptin ratio for postmenopausal breast cancer prevention. Front Biosci. 2009;1:329–57.
Dos Santos E, Benaitreau D, Dieudonne MN, Leneveu MC, Serazin V, Giudicelli Y, Pecquery R. Adiponectin mediates an antiproliferative response in human MDA-MB 231 breast cancer cells. Oncol Rep. 2008;20(4):971–7.
Fischer-Posovszky P, Wabitsch M, Hochberg Z. Endocrinology of adipose tissue - an update. Horm Metab Res. 2007;39(5):314–21.
Jarde T, Caldefie-Chezet F, Goncalves-Mendes N, Mishellany F, Buechler C, Penault-Llorca F, Vasson MP. Involvement of adiponectin and leptin in breast cancer: clinical and in vitro studies. Endocr Relat Cancer. 2009;16(4):1197–210.
Wang Y, Lam KS, Xu JY, Lu G, Xu LY, Cooper GJ, Xu A. Adiponectin inhibits cell proliferation by interacting with several growth factors in an oligomerization-dependent manner. J Biol Chem. 2005;280(18):18341–7.
Arita Y, Kihara S, Ouchi N, Takahashi M, Maeda K, Miyagawa J, Hotta K, Shimomura I, Nakamura T, Miyaoka K, et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem Biophys Res Commun. 1999;257(1):79–83.
Haluzik M, Parizkova J, Haluzik MM. Adiponectin and its role in the obesity-induced insulin resistance and related complications. Physiol Res. 2004;53(2):123–9.
Weyer C, Funahashi T, Tanaka S, Hotta K, Matsuzawa Y, Pratley RE, Tataranni PA. Hypoadiponectinemia in obesity and type 2 diabetes: close association with insulin resistance and hyperinsulinemia. J Clin Endocrinol Metab. 2001;86(5):1930–5.
Chen DC, Chung YF, Yeh YT, Chaung HC, Kuo FC, Fu OY, Chen HY, Hou MF, Yuan SS. Serum adiponectin and leptin levels in Taiwanese breast cancer patients. Cancer Lett. 2006;237(1):109–14.
Mantzoros C, Petridou E, Dessypris N, Chavelas C, Dalamaga M, Alexe DM, Papadiamantis Y, Markopoulos C, Spanos E, Chrousos G, et al. Adiponectin and breast cancer risk. J Clin Endocrinol Metab. 2004;89(3):1102–7.
Miyoshi Y, Funahashi T, Kihara S, Taguchi T, Tamaki Y, Matsuzawa Y, Noguchi S. Association of serum adiponectin levels with breast cancer risk. Clin Cancer Res. 2003;9(15):5699–704.
Tworoger SS, Eliassen AH, Kelesidis T, Colditz GA, Willett WC, Mantzoros CS, Hankinson SE. Plasma adiponectin concentrations and risk of incident breast cancer. J Clin Endocrinol Metab. 2007;92(4):1510–6.
DeLellis HK, Rinaldi S, Kaaks R, Kolonel L, Henderson B, Le Marchand L. Lifestyle and dietary correlates of plasma insulin-like growth factor binding protein-1 (IGFBP-1), leptin, and C-peptide: the multiethnic cohort. Nutr Cancer. 2007;58(2):136–45.
Ragin CC, Dallal C, Okobia M, Modugno F, Chen J, Garte S, Taioli E. Leptin levels and leptin receptor polymorphism frequency in healthy populations. Infect Agent Cancer. 2009;4(Suppl 1):S13.
Han C, Zhang HT, Du L, Liu X, Jing J, Zhao X, Yang X, Tian B. Serum levels of leptin, insulin, and lipids in relation to breast cancer in China. Endocrine. 2005;26(1):19–24.
Liu CL, Chang YC, Cheng SP, Chern SR, Yang TL, Lee JJ, Guo IC, Chen CP. The roles of serum leptin concentration and polymorphism in leptin receptor gene at codon 109 in breast cancer. Oncology. 2007;72(1–2):75–81.
Wu MH, Chou YC, Chou WY, Hsu GC, Chu CH, Yu CP, Yu JC, Sun CA. Circulating levels of leptin, adiposity and breast cancer risk. Br J Cancer. 2009;100(4):578–82.
Maeda K, Okubo K, Shimomura I, Funahashi T, Matsuzawa Y, Matsubara K. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). Biochem Biophys Res Commun. 1996;221(2):286–9.
Jarde T, Caldefie-Chezet F, Damez M, Mishellany F, Perrone D, Penault-Llorca F, Guillot J, Vasson MP. Adiponectin and leptin expression in primary ductal breast cancer and in adjacent healthy epithelial and myoepithelial tissue. Histopathology. 2008;53(4):484–7.
Takahata C, Miyoshi Y, Irahara N, Taguchi T, Tamaki Y, Noguchi S. Demonstration of adiponectin receptors 1 and 2 mRNA expression in human breast cancer cells. Cancer Lett. 2007;250(2):229–36.
Kershaw EE, Flier JS. Adipose tissue as an endocrine organ. J Clin Endocrinol Metab. 2004;89(6):2548–56.
Dieudonne MN, Bussiere M, Dos Santos E, Leneveu MC, Giudicelli Y, Pecquery R. Adiponectin mediates antiproliferative and apoptotic responses in human MCF7 breast cancer cells. Biochem Biophys Res Commun. 2006;345(1):271–9.
Fasshauer M, Klein J, Neumann S, Eszlinger M, Paschke R. Tumor necrosis factor alpha is a negative regulator of resistin gene expression and secretion in 3T3-L1 adipocytes. Biochem Biophys Res Commun. 2001;288(4):1027–31.
Makimura H, Mizuno TM, Bergen H, Mobbs CV. Adiponectin is stimulated by adrenalectomy in ob/ob mice and is highly correlated with resistin mRNA. Am J Physiol Endocrinol Metab. 2002;283(6):E1266–71.
Hu X, Juneja SC, Maihle NJ, Cleary MP. Leptin--a growth factor in normal and malignant breast cells and for normal mammary gland development. J Natl Cancer Inst. 2002;94(22):1704–11.
O'Brien SN, Welter BH, Price TM. Presence of leptin in breast cell lines and breast tumors. Biochem Biophys Res Commun. 1999;259(3):695–8.
Dieudonne MN, Machinal-Quelin F, Serazin-Leroy V, Leneveu MC, Pecquery R, Giudicelli Y. Leptin mediates a proliferative response in human MCF7 breast cancer cells. Biochem Biophys Res Commun. 2002;293(1):622–8.
Cao R, Brakenhielm E, Wahlestedt C, Thyberg J, Cao Y. Leptin induces vascular permeability and synergistically stimulates angiogenesis with FGF-2 and VEGF. Proc Natl Acad Sci U S A. 2001;98(11):6390–5.
Park HY, Kwon HM, Lim HJ, Hong BK, Lee JY, Park BE, Jang Y, Cho SY, Kim HS. Potential role of leptin in angiogenesis: leptin induces endothelial cell proliferation and expression of matrix metalloproteinases in vivo and in vitro. Exp Mol Med. 2001;33(2):95–102.
Somasundar P, McFadden DW, Hileman SM, Vona-Davis L. Leptin is a growth factor in cancer. J Surg Res. 2004;116(2):337–49.
Laud K, Gourdou I, Pessemesse L, Peyrat JP, Djiane J. Identification of leptin receptors in human breast cancer: functional activity in the T47-D breast cancer cell line. Mol Cell Endocrinol. 2002;188(1–2):219–26.
Jarde T, Perrier S, Vasson MP, Caldefie-Chezet F. Molecular mechanisms of leptin and adiponectin in breast cancer. Eur J Cancer. 2011;47(1):33–43.
Ambrosone CB, Ciupak GL, Bandera EV, Jandorf L, Bovbjerg DH, Zirpoli G, Pawlish K, Godbold J, Furberg H, Fatone A, et al. Conducting molecular epidemiological research in the age of HIPAA: a multi-institutional case-control study of breast cancer in African-American and European-American women. J Oncol. 2009;2009:871250.
Bandera EV, Chandran U, Zirpoli G, Gong Z, McCann SE, Hong CC, Ciupak G, Pawlish K, Ambrosone CB. Body fatness and breast cancer risk in women of African ancestry. BMC Cancer. 2013;13:475.
George P, Chandwani S, Gabel M, Ambrosone CB, Rhoads G, Bandera EV, Demissie K. Diagnosis and surgical delays in African American and white women with early-stage breast cancer. J Women's Health (Larchmt). 2015;24(3):209–17.
Chen W, Reiss M, Foran DJ. A prototype for unsupervised analysis of tissue microarrays for cancer research and diagnostics. IEEE Trans Inf Technol Biomed. 2004;8(2):89–96.
Zimta AA, Tigu AB, Muntean M, Cenariu D, Slaby O, Berindan-Neagoe I. Molecular links between central obesity and breast cancer. Int J Mol Sci. 2019;20(21):5364.
Raut PK, Kim SH, Choi DY, Jeong GS, Park PH. Growth of breast cancer cells by leptin is mediated via activation of the inflammasome: critical roles of estrogen receptor signaling and reactive oxygen species production. Biochem Pharmacol. 2019;161:73–88.
Yoon YS, Kwon AR, Lee YK, Oh SW. Circulating adipokines and risk of obesity related cancers: a systematic review and meta-analysis. Obes Res Clin Pract. 2019.
Ishikawa M, Kitayama J, Nagawa H. Enhanced expression of leptin and leptin receptor (OB-R) in human breast cancer. Clin Cancer Res. 2004;10(13):4325–31.
Caldefie-Chezet F, Damez M, de Latour M, Konska G, Mishellani F, Fusillier C, Guerry M, Penault-Llorca F, Guillot J, Vasson MP. Leptin: a proliferative factor for breast cancer? Study on human ductal carcinoma. Biochem Biophys Res Commun. 2005;334(3):737–41.
Garofalo C, Koda M, Cascio S, Sulkowska M, Kanczuga-Koda L, Golaszewska J, Russo A, Sulkowski S, Surmacz E. Increased expression of leptin and the leptin receptor as a marker of breast cancer progression: possible role of obesity-related stimuli. Clin Cancer Res. 2006;12(5):1447–53.
Kienesberger PC, Lee D, Pulinilkunnil T, Brenner DS, Cai L, Magnes C, Koefeler HC, Streith IE, Rechberger GN, Haemmerle G, et al. Adipose triglyceride lipase deficiency causes tissue-specific changes in insulin signaling. J Biol Chem. 2009;284(44):30218–29.
Kim Y, Kim SY, Lee JJ, Seo J, Kim YW, Koh SH, Yoon HJ, Cho KS. Effects of the expression of leptin and leptin receptor (OBR) on the prognosis of early-stage breast cancers. Cancer Res Treat. 2006;38(3):126–32.
Fiorio E, Mercanti A, Terrasi M, Micciolo R, Remo A, Auriemma A, Molino A, Parolin V, Di Stefano B, Bonetti F, et al. Leptin/HER2 crosstalk in breast cancer: in vitro study and preliminary in vivo analysis. BMC Cancer. 2008;8:305.
Jeong YJ, Bong JG, Park SH, Choi JH, Oh HK. Expression of leptin, leptin receptor, adiponectin, and adiponectin receptor in ductal carcinoma in situ and invasive breast cancer. J Breast Cancer. 2011;14(2):96–103.
Colbert LS, Wilson K, Kim S, Liu Y, Oprea-Ilies G, Gillespie C, Dickson T, Newman G, Gonzalez-Perez RR. NILCO biomarkers in breast cancer from Chinese patients. BMC Cancer. 2014;14:249.
Jarde T, Caldefie-Chezet F, Damez M, Mishellany F, Penault-Llorca F, Guillot J, Vasson MP. Leptin and leptin receptor involvement in cancer development: a study on human primary breast carcinoma. Oncol Rep. 2008;19(4):905–11.
Cheng SP, Chi CW, Tzen CY, Yang TL, Lee JJ, Liu TP, Liu CL. Clinicopathologic significance of leptin and leptin receptor expressions in papillary thyroid carcinoma. Surgery. 2010;147(6):847–53.
Fan YL, Li XQ. Expression of leptin and its receptor in thyroid carcinoma: distinctive prognostic significance in different subtypes. Clin Endocrinol. 2015;83(2):261–7.
Osorio CF, Souza DB, Gallo CB, Costa WS, Sampaio FJ. Leptin and leptin receptor expressions in prostate tumors may predict disease aggressiveness? Acta Cir Bras. 2014;29(Suppl 3):44–8.
Uddin S, Bavi PP, Hussain AR, Alsbeih G, Al-Sanea N, Abduljabbar A, Ashari LH, Alhomoud S, Al-Dayel F, Ahmed M, et al. Leptin receptor expression in Middle Eastern colorectal cancer and its potential clinical implication. Carcinogenesis. 2009;30(11):1832–40.
Yuan SS, Tsai KB, Chung YF, Chan TF, Yeh YT, Tsai LY, Su JH. Aberrant expression and possible involvement of the leptin receptor in endometrial cancer. Gynecol Oncol. 2004;92(3):769–75.
Zhang Y, Liu L, Li C, Ai H. Correlation analysis between the expressions of leptin and its receptor (ObR) and clinicopathology in endometrial cancer. Cancer Biomark. 2014;14(5):353–9.
Gelsomino L, Naimo GD, Catalano S, Mauro L, Ando S. The emerging role of adiponectin in female malignancies. Int J Mol Sci. 2019;20(9):2127.
Drew JE. Molecular mechanisms linking adipokines to obesity-related colon cancer: focus on leptin. Proc Nutr Soc. 2012;71(1):175–80.
Mauro L, Pellegrino M, De Amicis F, Ricchio E, Giordano F, Rizza P, Catalano S, Bonofiglio D, Sisci D, Panno ML, et al. Evidences that estrogen receptor alpha interferes with adiponectin effects on breast cancer cell growth. Cell Cycle. 2014;13(4):553–64.
Mauro L, Pellegrino M, Giordano F, Ricchio E, Rizza P, De Amicis F, Catalano S, Bonofiglio D, Panno ML, Ando S. Estrogen receptor-alpha drives adiponectin effects on cyclin D1 expression in breast cancer cells. FASEB J. 2015;29(5):2150–60.
Acknowledgements
We are sincerely appreciative of the breast cancer advocates, community partners, and all participants of the Women’s Circle of Health Study for generously donating their time to participate in the study. We are equally grateful to the highly motivated, hardworking research personnel at the Rutgers School of Public Health, Rutgers Cancer Institute of New Jersey, Roswell Park Cancer Institute, Mount Sinai School of Medicine (now Icahn School of Medicine at Mount Sinai), and New Jersey State Cancer Registry who have been involved in this research. Services, results, and/or products in support of the research project were generated by the Rutgers Cancer Institute of New Jersey Biomedical Informatics Shared Resource (supported, in part, with funding from NCI-CCSG P30CA072720-5917) and the Biospecimen Repository and Histopathology Service Shared Resource (supported, in part, with funding from NCI-CCSG P30CA072720-5919).
Funding
Research reported in this publication was supported by the National Cancer Institute of the National Institutes of Health under the following award numbers: P01CA151135 (awarded to C.B. Ambrosone), P30CA072720 (awarded to S. Libutti), R01CA100598 (awarded to C.B. Ambrosone), R01CA185623 (awarded to E.V. Bandera, K. Demissie, and C.C. Hong), K01CA193527 (awarded to A.A.M. Llanos), and K08CA172722 (awarded to C. Omene). Research in this publication was also supported by the US Army Medical Research and Development Command under award number DAMD-17-01-1-0334 (awarded to D.H. Bovbjerg), the Breast Cancer Research Foundation (awarded to C.B. Ambrosone and C.C. Hong), and a gift from the Philip L. Hubbell Family (awarded to K. Demissie). Tumor samples were received, processed, and tracked under the auspices of the Roswell Park Comprehensive Cancer Center Data Bank and BioRepository Shared Resource, with funding from NCI-CCSG P30CA16056. Services, results, and/or products in support of this research project were generated using the Rutgers Cancer Institute of New Jersey Biomedical Informatics Shared Resource (P30CA072720-5917) and the Biospecimen Repository and Histopathology Service Shared Resource (P30CA072720-5919). The New Jersey State Cancer Registry, Cancer Epidemiology Services, New Jersey Department of Health, is funded by the Surveillance, Epidemiology and End Results (SEER) Program of the National Cancer Institute under contract HHSN261201300021I and control no. N01-PC-2013-00021, the National Program of Cancer Registries (NPCR), Centers for Disease Control and Prevention under grant NU5U58DP006279-02-00 as well as the State of New Jersey and the Rutgers Cancer Institute of New Jersey.
Author information
Authors and Affiliations
Contributions
AAML designed the study, analyzed the data, and wrote the manuscript. YL contributed to the study design and statistical analysis. MAC, LC, ARO, and TK contributed to the tissue microarray construction and histology and immunohistochemistry methodology development. WC, JN, MAC, and DJF developed and/or contributed to the digital pathology analysis for quantitative assessment of IHC expression. AAML, SY, CO, CCH, SG, MH, CBA, EVB, and KD provided material support and study supervision, contributed to data collection, and were major contributors in reviewing and editing the manuscript. All authors reviewed and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was approved by the Institutional Review Boards of all participating institutions, and all study participants provided written informed consent prior to study enrollment.
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1:
Table S1. Mean adipokine and adipokine receptor IHC expression, by select factors, among Black participants only.
Additional file 2:
Table S2. Mean adipokine and adipokine receptor IHC expression, by select factors, among White participants only.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Llanos, A.A.M., Lin, Y., Chen, W. et al. Immunohistochemical analysis of adipokine and adipokine receptor expression in the breast tumor microenvironment: associations of lower leptin receptor expression with estrogen receptor-negative status and triple-negative subtype. Breast Cancer Res 22, 18 (2020). https://doi.org/10.1186/s13058-020-1256-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13058-020-1256-3