
Abstract
Background
The incidence of metabolic syndrome linked to dyslipidemia and insulin resistance has increased; thus, studies must be conducted to elucidate this phenomenon. The present study aimed to investigate the protective effects of chokeberry and dried jujube diet on high-fat and high-fructose diet-induced dyslipidemia in mice.
Methods
Male C57BL/6 J mice were divided into five groups: ND, mice fed normal diet and tap water; HFFD, mice fed 60% high-fat and 10% fructose diet (HFFD) in tap water; HFFD+C, mice fed HFFD with 1% chokeberry powder; HFFD+J, mice fed HFFD with 1% jujube fruit powder; and HFFD+M, mice fed HFFD with 0.5% chokeberry + 0.5% jujube fruit powder mixture.
Results
After 10 weeks of dietary treatment, chokeberry and dried jujube fruits reduced HFFD-induced weight gain and central obesity and decreased liver weight and abdominal and epididymal fat mass. Furthermore, such fruits attenuated HFFD-induced dyslipidemia; decreased triglyceride, total cholesterol, non-high-density lipoprotein-cholesterol, low-density lipoprotein-cholesterol, and very-low-density lipoprotein-cholesterol levels. Insulin resistance was improved via the consumption of dietary chokeberry and dried jujube fruits according to various indicators (serum insulin level, fasting blood glucose level, homeostatic model assessment-insulin resistance score, and oral glucose tolerance test value). These treatments were found to lower serum triglyceride levels. Moreover, the consumption of chokeberry and dried jujube changed the hepatic protein expression of insulin receptor, insulin receptor substrate 1, phosphoinositide 3-kinase, Akt, and catalase, which are associated with insulin resistance.
Conclusions
Chokeberry and dried jujube could be used in the management of dyslipidemia and insulin resistance associated with metabolic syndrome by reducing risk parameters in mice with HFFD.
Similar content being viewed by others


Background
High-fat and high-fructose diet (HFFD), which is a western diet, is generally composed of carbohydrates, such as fructose and sucrose, as well as saturated fat. This diet has been associated with numerous diet-induced complications, such as obesity, type II diabetes mellitus (T2DM), and metabolic syndrome (MetS), which is a pathological state characterized by hypertension, abdominal obesity, dyslipidemia, and hyperglycemia, and such characteristics are considered major public health problems worldwide. Their complex conditions are significantly correlated to overweight or obesity and insulin resistance, which is an important factor in the development of MetS and T2DM. In particular, hepatic insulin resistance is correlated to a decrease in insulin signal transmission for inhibiting glucose production and insulin-stimulated hepatic lipogenesis [1]. In the Republic of Korea, the prevalence of MetS in adults over 30 is reported to be 30% according to a report from Korea Centers for Disease Control and Prevention (KCDC). For this reason, there are continuing needs to prevention study to relieve the MetS in South Korea.
Various studies have been shown that polyphenol-rich diet could reduce the risk of developing MetS [2,3,4,5,6]. Polyphenols have potent antioxidant properties and perform a variety of biological functions such as anticancer and anti-inflammatory properties, and lipid homeostasis [2, 7,8,9]. For this reason, the relationship between polyphenols in fruits and various health problems is being assessed in research studies. According to the previous studies, polyphenols containing flavonoids could contribute the health benefits by improvement of insulin receptor substrate (IRS)/ phosphoinositide 3-kinase (PI3K)/Akt pathway [10] and of insulin sensitivity with reduced markers of inflammation [11]. The association between the HFFD diet and the IRS/PI3K/Akt pathway will be described in the following paragraphs.
Several studies have investigated the actual mechanism of MetS and insulin receptor (IR)/IRS1 with PI3K/Akt pathway, which is a downstream of the insulin signaling pathway. Insulin receptor, a glycoprotein consisting of an extracellular α-subunit (135 kDa) and a transmembrane β-subunit (95 kDa), is an allosteric enzyme in which the α-subunit inhibits the tyrosine kinase activity of the β-subunit [12]. IR combined with insulin initiates a cascade of phosphorylation events, including that of downstream as well as the IRS and PI3K/Akt pathway [13, 14]. Activation of IRS/PI3K/Akt pathway is the main target that alleviating dyslipidemia, insulin resistance and obesity [12, 15,16,17].
Insulin is secreted by the β-cells of the pancreatic islet of Langerhans. The secreted insulin binds to the IR that traverses the cell membrane. Consequently, they transmit insulin signaling across the cell membrane, resulting in a sequence of further phosphorylation reactions, such as that observed for PI3K [14]. Akt, which is also called protein kinase B, is a key downstream effector of the PI3K pathway. Akt mediates most of the metabolic effects of insulin, thereby regulating glucose transport, lipid synthesis, gluconeogenesis, and glycogen synthesis. Moreover, it plays a role in the control of cell cycle and survival [13, 18]. The activation of PI3K generated phosphatidylinositol-3,4,5-triphosphate (PIP3) from phosphatidylinositol-4,5-bisphosphate (PIP2), and as a result, insulin affects metabolism by activating Akt via phosphorylation (at T308 and S473) [12]. Activated Akt promotes the downstream effectors, which regulates glucose metabolism via FoxO protein, particularly FoxO1 and glucose 6-phosphate and glycogen synthase kinase 3 (GSK-3) and lipid metabolism via mTOR complex 1 (mTORC1) and sterol regulatory element-binding proteins (SREBP) [19]. Therefore, activation of IRS/PI3K/Akt pathway modulates the risk of MetS including dyslipidemia and insulin resistance.
Black chokeberry (Aronia melanocarpa) is the main breed of the Aronia melanocarpa species, and it is classified according to color difference between red and black. Chokeberry is a member of the Rosaceae family, and it contains several phytochemicals, such as total polyphenols, procyanidins, anthocyanins, and flavonols [20, 21]. Moreover, it is known as a super food worldwide and has been used as a research material to elucidate the effects of dietary intake. Such fruits have been extensively used to validate protective effects against aging [22], dyslipidemia [23], hyperglycemic state [24, 25], liver damage [23, 26, 27], and hypertension [5]. Chokeberry has been used in limited industrial productions due to its strong sour taste, bitterness, and unripe smell. In relation to this reason, such fruit is blended with foods to compensate for its undesirable taste and smell rather than being used alone [20].
Jujube (Ziziphus jujuba) is considered a health-promoting food in Asia. Such fruit is dried to increase its nutrient contents, and it has been used as a traditional medicine in Asia. According to previous studies, the nutritional contents of jujube could change during drying process [28, 29]. In a review article, the jujube fruit contains minerals, vitamins, polyphenols, flavonoids, anthocyanins, and proanthocyanidins [30]. Dried jujube has beneficial effects on hepatoprotection [31], diabetes [32], dyslipidemia [33], and inflammation [34].
As interest in the consumption of health-promoted food containing fruits increased in Korea, research was needed to establish scientific evidences of protective effect. Therefore, the current study was conducted whether the effects of chokeberry and jujube fruits could reduce the risk of MetS when consumed individually or in combination. Many studies reported chokeberry consumption led to reduce the risk of obesity, however, it is difficult to applicate the individual consumption because of bitterness. Nowadays, functional foods in combination form are being developed. Despite these efforts, the scientific evidences are insufficient. The tests on availability continue, however there are weaknesses. This is because most tests are only been conducting on the antioxidative activities of food without confirming changes of the specific mechanism, which promotes health. This study is the only experiment that directly feeding in form of fruit powder to mice with MetS induced by HFFD diet and have specialty because the present study conducted health-promoting effects with combination form of different fruits. Therefore, there is uniqueness and originality in current study.
This study aimed to investigate the effects of dietary chokeberry and/or dried jujube fruits on central obesity, lipid homeostasis, glucose intolerance, and insulin resistance that are induced by HFFD in C57BL/6j mice by controlling the IRS/PI3K/Akt pathway. These data would provide sound scientific evidence for the clinical treatment of dyslipidemia and insulin resistance.
Methods
Ethical approval
This experimental design was approved by the Institutional Animal Care and Use Committee (IACUC) of Sookmyung Women’s University (SMWU-IACUC-1701-044).
Experimental scheme
Lipid and glucose metabolism have been documented as important risk factors of metabolic disorder. Thus, this study aimed to investigate the synergistic effects of chokeberry (Aronia melanocarpa) and jujube (Ziziphus jujuba Mill.) diet on lipid metabolism and antioxidant capacity in HFFD-induced metabolic disorders in the C57BL/6 J mice model.
After the acclimation period, 6-week-old C57BL/6 J mice were divided into five groups: ND, mice fed with normal diet and tap water; HFFD, mice fed with HFFD (60% of fat, 10% of fructose) in tap water; HFFD+C, mice fed with HFFD with 1% chokeberry powder and 10% fructose in tap water; HFFD+J, mice fed with HFFD with 1% jujube fruit powder and 10% fructose in tap water; and HFFD+M, mice fed with HFFD with 0.5% chokeberry + 0.5% jujube fruit powder mixture and 10% fructose in tap water (Fig. 1).

Experimental Scheme of Animal and Diet. In the current study, 6-week-old C57BL/6 J mice were divided into five groups; ND (normal diet with tap water), HFFD (60% of fat with 10% of fructose water), HFFD+C (HFFD with 1% of chokeberry powder), HFFD+J (HFFD with 1% jujube fruit powder), and HFFD+M (HFFD with mixture that contains 0.5% chokeberry and 0.5% jujube fruit powder)
The animals were euthanized with CO2 after 10 weeks of treatment. Their serum, organs (heart, kidneys, lung, and liver), and tissues (abdominal fat and epididymal fat) were isolated. Serum was separated by centrifugation at 3000 rpm for 45 min and stored at − 70 °C until analysis. For liver analysis, the liver samples were homogenized, and hepatic triglyceride (TG) level was measured. Moreover, hepatic protein expression of Insulin Receptor (IR), Insulin Receptor Substrate 1 (IRS-1), PI3K, phosphor-PI3K (p-PI3K), Akt, phosphor-Akt (p-Akt), and Catalase (CAT) were measured via Western blot analysis.
Animals and diets
The 5-week-old male C57BL/6 J mice (Saeronbio Inc., Gyeonggi-do, Korea) were housed with controlled temperature (21 ± 1 °C) and humidity (50–60%) conditions in a 12-h light/dark cycle throughout the study. They had free access to water and chow diet.
After 1 week of acclimation, the 6-week-old mice were randomly divided into five groups (n = 7 for each group): [1] ND, mice fed with normal diet (AIN-93G) and tap water; [2] HFFD, mice fed with HFFD (60% fat, 10% fructose) in tap water; [3] HFFD+C, mice fed with HFFD with 1% chokeberry powder and 10% fructose in tap water; [4] HFFD+J, mice fed with HFFD with 1% jujube fruit powder and 10% fructose in tap water; and [5] HFFD+M, mice fed with HFFD with 0.5% chokeberry and 0.5% jujube fruit powder mixture and 10% fructose in tap water. The normal diet was the AIN-93G diet (Research diet, New Brunswick, NJ, the USA), and the high-fat diet was the D12492 diet (Research diet, New Brunswick, NJ, the USA). The experimental scheme of the current study is shown in Fig. 1.
Fresh chokeberries were purchased from Ofresh (Gochang, Jeonbuk, Korea) and freeze dried for 2 days (Bondiro MCFD 8508 Freeze Dryer, Ilshin, Seoul, Korea). Dried jujubes were purchased from Boeun Dule farm (Boeun, Chungbuk, Korea). Freeze-dried chokeberry and dried jujube were grounded and mixed to the diet. All diets, except the normal diet, were isocaloric. The nutritional characterization of chokeberry and jujube fruit powders are listed in Table 1. Table 2 shows the composition of each experimental diets. The percentage of chokeberry (1%) was chosen according to the previous pilot studies which investigated by our laboratory [22, 26, 35]. We decided and tested the 1% of jujube fruits as the same dose with chokeberry. Moreover, we chose to use 0.5% of chokeberry and 0.5% of dried jujube fruits (HFFD+M) in the mixed powder group because we hoped to match that experimental groups were equal to 1% of the total intake.
Measurement of body weight, food intake, and water intake
Body weight for each animal was recorded weekly. Food intake and water intake were measured every 2 days, and body weight was evaluated once a week during the feeding period. The animals were weighed weekly, starting from the day of arrival. The final body weight was measured a day before the sacrifice. The food efficiency ratio (FER) was calculated using the following equation:
FER = total body weight gain (g) / total food intake (g) × 100.
Blood and tissue preparation
After an overnight fast, the final body weight was measured, and the mice were euthanized using CO2. Blood sample was collected via cardiac puncture to determine the serum lipid profiles. Serum was separated by centrifugation at 3000 rpm for 45 min (Combi-450R, Hanil Co. Ltd., Seoul, Korea) and stored at − 70 °C until analysis (DF8517; Ilshin Laboratory Co., Ltd., Seoul, Korea). Hearts, kidneys, lungs, livers, abdominal fats, and epididymal fats were isolated and measured with an electronic balance (OHAUS, NJ, the USA). All organs were stored at − 70 °C until analysis (DF8517; Ilshin Laboratory Co., Ltd., Seoul, Korea). The organ coefficient of each organ was calculated with following equation:
Glucose metabolism profiles
-
1.
Fasting blood glucose level, serum insulin concentration, and HOMA-IR score
Fasting glucose concentration was directly evaluated using the GlucoCard X-Meter (Arkray, Kyoto, Japan). Serum insulin levels were measured using the Insulin Mouse ELISA kit (80-INSMS-E01, ALPCO, Salem, NH). The Homeostatic Model Assessment-Insulin Resistance (HOMA-IR) score, which is called the homeostasis model assessment of insulin resistance, was calculated using fasting blood glucose and insulin concentrations.
-
2.
Oral glucose tolerance tests
After 10 weeks of consuming the experimental diet, according to their respective groups, oral glucose tolerance tests (OGTTs) were performed. All mice were fasted overnight and then provided with 2 g/kg d-glucose solution. Whole blood was collected from the caudal vein, and blood sugar level was measured using a glucose monitoring device (Arkray, Kyoto, Japan) immediately after treatment at 0, 30, 60, 90, and 120 min.
Lipid profiles
-
1.
Serum TG, TC, HDL, LDL, and VLDL levels
Serum TG and total cholesterol (TC) levels were measured with the TG-S kit (3I1570, Asanpharm, Hwaseong, Korea) and T-CHO kit (3I2020, Asanpharm, Hwaseong, Korea), respectively. The HDL-CHO kit (3I2030, Asanpharm, Hwaseong, Korea) was used to measure serum high-density lipoprotein cholesterol (HDL-C) levels.
Low-density lipoprotein and very-low-density lipoprotein cholesterol (LDL-C and VLDL-C) levels were calculated using the Friedewald equation [36] as follows:
LDL-C level (mg/dL) = TC level-(HDL-C level + TG level/5) (mg/dL)
VLDL-C level (mg/dL) = TG level/5 (mg/dL)
-
2.
Atherosclerotic index (AI) and cardiac risk factor (CRF)
Atherosclerotic index (AI) and cardiac risk factor (CRF) were calculated using the following formula [36, 37]:
AI = (TC level – HDL-C level)/HDL-C level
CRF = TC level/HDL-C level
Liver TG levels
Total liver lipid levels were obtained using the Folch method [38]; then, liver TG levels were determined. Briefly, the liver samples were homogenized with chloroform/methanol (2:1) solution to a final dilution of 20 folds than the volume of the tissue sample. The samples were mixed for 15 min and were centrifuged at 1000 rpm for 5 min at room temperature. The upper phase was removed as completely as possible with a pipette. Next, the lower phase was evaporated on a hot plate until chloroform disappeared. Liver TG levels were identified using the TG-S kit (3I1570, Asanpharm, Hwaseong, Korea) and were measured.
Western blot analysis
In total, 8 mg of liver tissue was used to extract protein in the liver with the Pro-prep kit (17,081, iNtRON biotechnology, Gyeonggi-do, Korea). Protein concentration was measured using the PRO-MEASURE™ kit (21,011, iNtRON biotechnology, Gyeonggi-do, Korea). Protein samples were resolved on the SDS-PAGE and were transferred to the polyvinylidene difluoride (PVDF) membrane (Merck Millipore, MA, the USA) via electrophoretic transfer (Bio-Rad Laboratories, Inc., Hercules, CA, the USA). The membrane was pre-blocked in 5% skim milk containing phosphate-buffered saline solution and 0.1% tween-20 (PBST) for an hour. Then, the membrane was incubated overnight at 4 °C with the following primary antibodies: Insulin Receptor (IR, 1:1000, Abcam, Cambridge, U.K.), Insulin Receptor Substrate 1 (IRS-1, 1:1000, Cell Signaling Technology Inc., MA, USA), PI3 kinase class III antibody (PI3K, 1:500, Cell Signaling Technology Inc., MA, USA), PI3 kinase p85 alpha + gamma (Tyr467 + Tyr199) antibody (p-PI3K, 1:500, Bioss, MA, USA), Akt polyclonal antibody (Akt, 1:1800, Abnova, Taipei, Taiwan), phospho-Akt (Ser 473), antibody (p-Akt, 1:1000, Cell Signaling Technology, Inc., MA, the USA), CAT (1:1000, Abnova, Taipei, Taiwan). After incubation, the membrane was washed with PBST for 10 min for 3 times in a row. Then, the membrane was incubated in goat anti-rabbit IgG H&L (HRP) secondary antibody (1:7500, Abnova, Taipei, Taiwan) and donkey anti-goat secondary antibody (1:7500, Abnova, Taipei, Taiwan). Protein level was normalized via glyceraldehyde 3-phosphate dehydrogenase (GAPDH) expression with GAPDH polyclonal antibody (1:1000, Abnova, Taipei, Taiwan). Immobilon Western horseradish peroxidase substrate (Merck Millipore, Bedford, MA, the USA) was used for chemiluminescence detection. The immunoreactive band intensities were quantified via densitometric analysis (LAS-3000, Fujifilm Co., Tokyo, Japan).
Statistical analysis
Statistical analysis was performed with SAS 9.4 (SAS Institute Inc., Cary, NC, the USA). All data were presented as mean ± SD. The results for each experimental group were compared using one-way analysis of variance (ANOVA). Differences in mean values between the five groups were tested with the Duncan’s multiple tests. A p value <.05 was considered statistically significant.
Results
Body weight, food intake, and food efficiency ratios
Body weight changes are presented in Fig. 2. At the start of the experiment period, body weight was not significantly different among all five groups. A significant elevation in the final body weight and body weight gain was observed in the group with high-fat and high-fructose diet for 10 weeks compared with the group with normal diet (p < .000 l). Compared with the HFFD group, the final body weight and body weight gain were reduced in the three groups fed with three fruits (p < .0001 for all groups). According to consumption of chokeberry and/or dried jujube fruits, there was significant decreases in HFFD+C, HFFD+J, and HFFD+M group in comparison with HFFD group although it was not up to the level of ND group. The weight changes in HFFD+C, HFFD+J and HFFD+M group were reduced by 31.42, 27.66 and 15.94% respectively, relative to the weight changes of the HFFD group.

Body Weight Changes of Each Group. Values are means ± SD (n = 7). The different letters (a, b, c) within a column indicate significant difference (p < .05) determined by Duncan multiple range test. Abbreviation: ns, not significant. ND: normal diet group, HFFD: high fat and high fructose (HFFD) diet group, HFFD+C: HFFD with 1% chokeberry powder group, HFFD+J: HFFD with 1% jujube fruits powder group, HFFD+M: HFFD with 0.5% chokeberry + 0.5% jujube fruits mixed powders group
Calorie intake per day did not significantly differ in all groups (p = 0.0865). The FER (%) of the HFFD group was statistically higher than the ND group (p < .0001). The HFFD+C, HFFD+J, and HFFD+M groups had a significantly lower FER than the HFFD group (p < .0001). Results of all groups are as follows: ND group with 4.03 ± 0.83%, HFFD group with 10.15 ± 2.62%, HFFD+C group with 7.70 ± 1.58%, HFFD+J group with 8.36 ± 1.91%, and HFFD+M group with 8.67 ± 1.91%. (Table 3).
Organ weight and organ coefficient
Based on the results of organ weight (Table 4) and coefficient of each organ, a significant difference was observed in liver, abdominal, and epididymal fat (p < .0001 for each group). The weight of liver, abdominal, and epididymal fat in the HFFD group increased compared with those of the ND group. The liver weight of the HFFD+C, HFFD+J, and HFFD+M groups was significantly lower than the hepatic weight of the HFFD group (p < .0001), and abdominal fat and epididymal fat mass decreased in the group fed with chokeberry and/or jujube (p < .0001, respectively). Organ coefficients were calculated using the liver, abdominal fat, epididymal fat, heart, kidney, and lung mass.
Serum lipid profiles
The serum lipid profiles are shown in Fig. 3. The administration of HFFD significantly induced higher levels of serum TG, TC, LDL-C, and VLDL-C levels (p < .0001 for all). However, serum HDL-C levels were not affected (p = 0.0614). After 10 weeks of administration, the HFFD+C, HFFD+J, and HFFD+M groups had significantly lower serum TG, TC, and VLDL-C levels relative to those level of HFFD group. The LDL-C levels (Fig. 2 (C)) of the HFFD+C and HFFD+M groups significantly differed from those of the HFFD group. Serum HDL-C levels, as shown in Fig. 2 (E), were not significantly different in all groups; however, a decreasing trend was observed in the HFFD+C, HFFD+J, and HFFD+M groups compared with the HFFD group.

Serum Lipid Profiles. (a) Total cholesterols, TC; (b) Total triglyceride, TG; (c) LDL-cholesterols, LDL-C; (d) VLDL-cholesterols, VLDL-C; (e) HDL-cholesterols, HDL-C; Values are means ± SD (n = 7). The different letters (a, b, c, d) within a column indicate significant difference (p < .05) determined by Duncan multiple range test. Abbreviation: ns, not significant. ND: normal diet group, HFFD: high fat and high fructose (HFFD) diet group, HFFD+C: HFFD with 1% chokeberry powder group, HFFD+J: HFFD with 1% jujube fruits powder group, HFFD+M: HFFD with 0.5% chokeberry + 0.5% jujube fruits mixed powders group
Serum glucose profiles and OGTTs
Figure 4 shows the serum glucose profiles, fasting serum glucose levels, index of HOMA-IR, and serum insulin levels. The HOMA-IR score was calculated with fasting serum glucose and insulin levels. Fasting serum glucose levels of the HFFD+C, HFFD+J, and HFFD+M groups were significantly lower than that of the HFFD group (p = 0.0066). Both HOMA-IR score and serum insulin level were significantly lower in the HFFD+C, HFFD+J, and HFFD+M groups than in the HFFD group (p < .0001).

Serum Glucose Profiles. a Fasting blood glucose of each week; b Serum insulin; (c) HOMA-IR; Values are means ± SD (n = 7). The different letters (a, b) within a column indicate significant difference (p < .05) determined by Duncan multiple range test. Abbreviation: ns, not significant. ND: normal diet group, HFFD: high fat and high fructose (HFFD) diet group, HFFD+C: HFFD with 1% chokeberry powder group, HFFD+J: HFFD with 1% jujube fruits powder group, HFFD+M: HFFD with 0.5% chokeberry + 0.5% jujube fruits mixed powders group
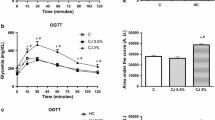
OGTTs were carried out after 9 weeks. The test results are shown in Fig. 5. At 0 min, the fasting blood glucose level of the HFFD group was significantly higher than that of the four groups (p = 0.0066). After the administration of 2 g/kg of D-glucose solution, no significant difference was observed at 30 and 60 min in all groups. The HFFD groups had the highest fasting blood glucose value among all groups after 120 min of glucose administration. The HFFD+C and HFFD+M groups had decreased trends at 120 min compared with the HFFD group; however, the difference was not statistically significant. The HFFD+J group had a significantly lower blood glucose level after 2 h of glucose administration than the HFFD group (p = 0.0029). The HFFD group showed a higher area under the curve (AUC) than the ND group. However, the difference was not significant.

Oral Glucose Tolerance Tests at 0, 30, 60, 90, and 120 min. Values are means ± SD (n = 7). The different letters (a, b) within a column indicate significant difference (p < .05) determined by Duncan multiple range test. Abbreviation: ns, not significant. ND: normal diet group, HFFD: high fat and high fructose (HFFD) diet group, HFFD+C: HFFD with 1% chokeberry powder group, HFFD+J: HFFD with 1% jujube fruits powder group, HFFD+M: HFFD with 0.5% chokeberry + 0.5% jujube fruits mixed powders group
AI and CRF
AI and CRF were calculated using the Haglund method [37], and the results were as follows: [1] AI: ND (0.80 ± 0.48b), HFFD (1.45 ± 0.45a), HFFD+C (0.81 ± 0.44b), HFFD+J (1.66 ± 0.35a), and HFFD+M (1.30 ± 0.39ab) (p = 0.00340) and [2] CRF: ND (1.80 ± 0.48b), HFFD (2.45 ± 0.45a), HFFD+C (1.81 ± 0.44b), HFFD+J (2.66 ± 0.35a), and HFFD+M (2.30 ± 0.39ab) (p = 0.00340). (Fig. 6).

Atherosclerotic Index and Cardiac Risk Factor of Each Group. Values are means ± SD (n = 7). The different letters (a, b) within a column indicate significant difference (p < .05) determined by Duncan multiple range test. Abbreviation: ND: normal diet group, HFFD: high fat and high fructose (HFFD) diet group, HFFD+C: HFFD with 1% chokeberry powder group, HFFD+J: HFFD with 1% jujube fruits powder group, HFFD+M: HFFD with 0.5% chokeberry + 0.5% jujube fruits mixed powders group
Liver TG
The TG levels of the mice are presented in Fig. 7 and Table 5. That of the HFFD group was 508.67 ± 105.07 mg/dL, and it was the highest among all groups. The liver TG values of the HFFD+J and HFFD+M groups were not significantly compared with those of the HFFD group. Moreover, a decreasing trend was observed. The TG level of the HFFD+C group (293.33 ± 211.14 mg/dL) was significantly lower than the HFFD group (p = 0.0192).

Liver Triglycerides of Each Group. Values are means ± SD (n = 7). The different letters (a, b) within a column indicate significant difference (p < .05) determined by Duncan multiple range test. ND: normal diet group, HFFD: high fat and high fructose (HFFD) diet group, HFFD+C: HFFD with 1% chokeberry powder group, HFFD+J: HFFD with 1% jujube fruits powder group, HFFD+M: HFFD with 0.5% chokeberry + 0.5% jujube fruits mixed powders group
Hepatic protein expression of IR, IRS-1, p-PI3K, PI3K, p-Akt, Akt, and CAT
Results of the representative Western blot analysis of IR, IRS-1, PI3K, p-PI3K, Akt, p-Akt, and CAT are shown in Figs. 8 and 9.

Hepatic Protein Expression of IRS-1/PI3K/Akt Pathway of Each Group. (a) IR; (b) IRS-1; (c) p-PI3K/PI3K; (d) p-Akt/Akt; Values are means ± SD (n= 7 ). The different letters (a, b) within a column indicate significant difference (p < .05) determined by Duncan multiple range test. Abbreviation: ns, not significant. ND: normal diet group, HFFD: high fat and high fructose (HFFD) diet group, HFFD+C: HFFD with 1% chokeberry powder group, HFFD+J: HFFD with 1% jujube fruits powder group, HFFD+M: HFFD with 0.5% chokeberry + 0.5% jujube fruits mixed powders group

Hepatic Protein Expression of CAT of Each Group. Values are means ± SD (n = 7). The different letters (a, b) within a column indicate significant difference (p < .05). Abbreviation: ns, not significant. ND: normal diet group, HFFD: high fat and high fructose (HFFD) diet group, HFFD+C: HFFD with 1% chokeberry powder group, HFFD+J: HFFD with 1% jujube fruits powder group, HFFD+M: HFFD with 0.5% chokeberry + 0.5% jujube fruits mixed powders group
IR is acted by insulin, which is secreted by controlling glucose/lipid metabolism. These factors increase insulin secretion, and insulin is then activated by IR tyrosine kinase, resulting to phosphorylated and activated insulin receptor substrate 1 (IRS-1). The IR expressions were lower in the four HFFD-fed groups than the ND group. However, the difference was not statistically significant among all five groups. The hepatic IRS-1 expression showed a compelling change in the difference between the groups. The expression in the HFFD+C and HFFD+M groups increased although the result was not statistically significant. Meanwhile, the increase in the expression in the HFFD+J group was more significant than that of the HFFD group (p < .05).
The PI3K/Akt pathway was analyzed via western blot analysis to monitor the activation of PI3K/Akt proteins by consumption of chokeberry and/or jujube on HFFD-induced metabolic disorders. The phosphorylation level of PI3K and Akt was calculated using the p-PI3K/PI3K and p-Akt/Akt ratio.
The phosphorylated PI3K/PI3K in the liver did not significantly differ among the five groups. A decreasing trend was observed among the HFFD, HFFD+C, HFFD+J, and HFFD+M groups compared with the ND group. Although it was not statistically significant, in terms of phosphorylated PI3K/PI3K, an increasing trend was observed in the HFFD+M group compared to the other groups.
Akt is downstream to PI3K, and it can be phosphorylated and activated via PI3K activation. In Fig. 8, the pAkt-to-Akt ratio of the HFFD groups was significantly lower than that of the ND groups (p = 0.003). A significantly higher expression was observed in the HFFD+M group than in the HFFD control group (p = 0.003).
Figure 9 shows that hepatic protein expression of CAT, and results were statistically significant among the five groups. CAT is considered an important enzyme because it protects the cell from oxidative stress by reactive oxygen species (ROS). The HFFD+M group had a significantly higher expression of CAT than the HFFD group, and this value was comparable to that of the ND group.
Discussion
Daily consumption of fruits can improve various biomarkers related to obesity because of their antioxidant properties. Previous studies have reported that chokeberry and jujube fruits have antioxidative properties because they contain polyphenols and flavonoids [20, 29, 30, 39, 40]. We previously measured the total phenol, flavonoids, and DPPH radical scavenging activity. The results of them were as follows: total phenols levels (chokeberry, 968.47 mg GAE/100 g; jujube fruits, 518.65 mg GAE/100 g; mixture of chokeberry and jujube fruit, 521.81 mg GAE/100 g), flavonoid levels (chokeberry, 374.08 mg QE/100 g; jujube fruits, 199.92 mg QE/100 g; mixture of chokeberry and jujube fruit, 244.08 1 mg QE/100 g), and DPPH radical scavenging activity (chokeberry, 78.59%; jujube fruit, 85.30%; mixture of chokeberry and jujube fruit, 82.32%. all of them compared with Ascorbic acid, 92.59%). We administered the same chokeberry and jujube powder to animals used in the current study. Therefore, this current study investigated the protective effects of chokeberry and/or jujube on lipid and glucose metabolism by attenuating the metabolic alteration associated with HFFD in C57BL/6 J mice. The high fat and high fructose diet-induced MetS mouse model has been used as a valuable rodent model [41, 42]. In previous studies, most HFFDs contained 45–60% of fat and up to 30% of fructose with drinking water [41,42,43,44,45,46,47,48]. Results showed that HFFD increased body weight and abdominal fat mass and reduced glucose function with insulin sensitivity [42]. Moreover, HFFD was correlated to insulin resistance and impaired lipid metabolism [48, 49].
To the best of our knowledge, this study first investigated the effects of dietary chokeberry and/or jujube fruits in mice models with HFFD-induced metabolic disorder. The present study confirmed that HFFD induced obesity and increased the level of body weight changes. Moreover, HFFD caused increased fat accumulation in abdominal and epididymal regions. These results demonstrated that HFFD is correlated to central obesity in mice.
The body weight of the ND and HFFD groups did not differ at the start of the study. However, differences were observed 10 weeks after the administration of HFFD. That is, the body weight of the HFFD group was 1.45-fold higher than that of the ND group. The changes in body weight of HFFD+C, HFFD+J, and HFFD+M group had persuasive decline in their weight compared to that of HFFD group. In this regard, dietary consumption of chokeberry and jujube fruits lead to lose the weight effectively in dyslipidemia and insulin resistance mice model induced by HFFD. Compared with the results of a review article [50], polyphenol rich foods have anti-obesity effects via several mechanisms involving suppression of adipocyte differentiation and proliferation, and inhibition of de novo lipogenesis and fatty acid oxidation. Our results also indicated that weight and organ weight were reduced when fruits containing polyphenols were consumed.
Based on our results, calorie intake per day was not different among all groups. Our results showed that the increase in body weight was not attributed to an increase in energy intake. The FER (%) increased the value of the HFFD group than the ND group. That is, the mice fed with HFFD gained more weight per equal feed intakes than mice fed with normal diet. Chokeberry and/or jujube suppressed FER (%) in this study.
In mice fed with HFFD, the higher levels of lipid profiles in the serum were associated with dyslipidemia [48, 51]. The present study showed increased serum TG, TC, HDL-C, non HDL-C, LDL-C, and VLDL-C levels, which is in accordance with previous studies. Chokeberry and/or jujube fruit diet had beneficial effects on attenuating serum lipid profiles but not HDL-C levels. Previous studies have shown that polyphenol-rich products can decrease serum lipid parameters leading to relieve the risk of dyslipidemia [6, 52,53,54]. Our findings indicated that dietary chokeberry and/or jujube have protective effects against dyslipidemia.
Both fasting blood glucose and serum insulin levels are important in HFFD-induced mice model. HFFD feeding is an important dietary pattern associated with abdominal adiposity and insulin resistance [55]. High caloric diet with fructose-enriched drinks is a proper method for inducing hepatic insulin resistance in rodent models [43]. This diet is a major contributor to insulin resistance because rodents that were fed with such diet had a higher HOMA-IR score. Shuang Mei et al. [56] have shown that a small amount of dietary carbohydrate can promote HFFD-induced insulin resistance to induce maximal levels. This study showed that HFFD increased fasting glucose concentration, HOMA-IR score, and serum insulin level in the HFFD group compared with the ND group. That is, HFFD feeding causes higher insulin resistance, hyperinsulinemia, and hyperglycemia. Chokeberry and/or dried jujube fruit diet significantly lowers the levels of fasting serum glucose and insulin and HOMA-IR score. Our findings indicated that the consumption of chokeberry and jujube fruit with HFFD might lower serum glucose profiles associated with insulin resistance due to their rich polyphenol contents.
OGTTs are performed to examine the effect of dietary chokeberry and/or dried jujube fruits on insulin sensitivity and glucose homeostasis. High fat and high carbohydrate (sucrose) treatment in mice resulted in increased plasma insulin level and units of AUC during OGTTs compared with normal chow diet [6]. In this study, the HFFD group had the highest point among the five groups after 120 min of D-glucose solution administration. In addition, the consumption of chokeberry and/or dried jujube fruits lowered blood glucose levels. In relation to this, our results showed that dietary chokeberry and/or dried jujube fruits may have protective effects against HFFD-related hyperglycemia and insulin sensitivity.
In a review article [57], insulin signaling is associated with the regulation of glucose and lipid metabolism by activating the downstream, such as the PI3K/Akt pathway. After secreted insulin combines with insulin receptor, which is located in the cell membrane, it activates IRS-1 and IRS-2, leading to the subsequent activation of the PI3K/Akt pathway. The aforementioned signaling cascade positively regulated cell growth, metabolism, and survival [12]. The results of the current study are similar to those of a review paper showing that the loss of IRS-1 is indicative of insulin resistance, and this loss is correlated to the inactivation of hepatic PI3K and Akt resulting in diabetes (hyperglycemia and hyperinsulinemia) and hypolipidemia [12]. Incidentally, the HFFD+C, HFFD+J, and HFFD+M groups showed an increasing trend compared with the HFFD group. In particular, the IRS-1 expression of HFFD+J group was significantly higher than the HFFD group. Therefore, the intake of fruits reduces the severity of insulin resistance via induction of IRS-1 expression in the liver.
PI3K interacts with Akt mainly in insulin signaling. The serine/threonine kinase Akt, which is also called protein kinase B (PKB), is an important protein in cell signaling downstream of insulin stimuli due to glucose transporter 4 (GLUT4) [18]. Studies that examined Akt expression in models with insulin resistance have shown increased Akt phosphorylation compared with normal groups [58, 59]. Another study has revealed reduction of p-Akt/Akt ratio in the HFFD group compared with the normal diet group [60]. According to an Akt article, Akt inactivation resulted in the impairment of systemic glucose and lipid homeostasis and body weight control in the development of MetS [12]. In relation to this, our study results about Akt expression in the liver were similar to those of a previous study. The HFFD+J and HFFD+M groups had activated expression of p-Akt/Akt in liver compared with HFFD group, but the expression of HFFD+M group only showed a convincing increase in comparison with HFFD group. In summary, the IRS-1 and p-Akt/Akt expressions decreased in the HFFD group and increased in the fruit-fed group, it suggested that the intake of the fruits can reduce the risk of developing MetS in mice fed with HFFD. The molecular mechanisms by which flavonoids and phenolic acids stimulate the expression of proteins involving IRS/PI3K/Akt pathway were reported by previous studies [10, 19, 59, 61,62,63]. Activation of IRS/PI3K/Akt pathway promotes the downstream markers such as FoxO1, GSK-3, mTORC1, and SREBP, which are major regulators of glucose and lipid metabolism. Our results indicated that chokeberry and jujube fruit may have therapeutic potency for alleviating the risk of MetS.
CAT is associated with obesity or insulin resistance, and it is a decomposing enzyme from H2O2 to O2 and H2O, resulting in oxidative stress plays an important role in the development of MetS [64]. In a review article [65], CAT polymorphisms were associated with the risk of developing diabetes mellitus since CAT decreased due to other factors such as genetics and environments. In our results, a valid value was only observed in the HFFD+M group. It indicated that mixed fruit feeding along with western-style HFFD diet may reduce the severity of insulin resistance. Since S. Wang et al. [66] have shown that the combination of different foods could have synergistic effects on total antioxidant capacities because mixed foods have more various bioactive compounds than single food. Taken together, our results would have been relatively better in the HFFD+M group. According to the previous study [67], activation of CAT is main to the induction of cellular antioxidant system, which can improve insulin resistance in in-vivo study [68]. They also confirmed that expression of inflammatory factors such as CAT by HFD-induced via PI3K/AKT/JNK pathway [67].
In summary, the current study investigated to elucidate the effect of individual or combined fruits consumption on dyslipidemia and insulin resistance-induced mice by high fat and high fructose diet. By daily intake of fruits for 10 weeks, there were significant reduction in body weight, organ weight, serum lipid profiles, serum glucose profiles, leading to upregulate hepatic protein expression of IRS/PI3K/Akt and CAT protein expression. Our results might be suggested that daily consumption of fruits has the potential to provide therapeutic help to alleviate dyslipidemia and insulin resistance by HFFD diet. In addition, this study showed that consumption of chokeberry with jujube has not only better taste but also more metabolic benefits than eating alone. The development of functional food with chokeberry, as known as superfood, has been attempted in South Korea. Our research provides scientific evidences and proposes as a potential therapeutic agent for functional foods. Based on the results of the present study, commercial availability for functional food development might be provided. There are limitations in this study due to lack of downstream effectors on the IRS/PI3K/Akt pathway. Therefore, direct or indirect findings will need to be developed through further studies.
Conclusion
The present study showed that the HFFD diet caused obesity, insulin resistance, hyperinsulinemia, and dyslipidemia associated with MetS through impaired parameters such as increased body weight, cholesterol levels (TC, TG, LDL, VLDL, and non HDL), fasting glucose concentration, and insulin level. The consumption of dietary chokeberry and dried jujube fruits improved the aforementioned indicators in this study. Dietary chokeberry and dried jujube fruits had protective effects in HFFD-fed mice via activation of IRS-1, p-Akt/Akt, and CAT in the liver. Hence, based on these results, the combination of chokeberry and jujube fruits may have therapeutic effects against obesity, insulin resistance, and MetS (Fig. 10).

The Proposed Mechanism for Chokeberry and Jujube’s action in liver. Chokeberry and jujube activate IRS/PI3K/Akt pathways on dyslipidemia and insulin resistance mice induced HFFD diet
Abbreviations
- AI:
-
Atherosclerotic index
- CAT:
-
Catalase
- CRF:
-
Cardiac risk factor
- FER:
-
Food efficiency ratio
- HDL-C:
-
High-density lipoprotein cholesterol
- HFFD:
-
High fat and high fructose diet
- HFFD+C:
-
HFFD with chokeberry powder
- HFFD+J:
-
HFFD with dried jujube fruit powder
- HFFD+M:
-
HFFD with chokeberry and dried jujube fruit mixed powder
- HOMA-IR:
-
Homeostatic model assessment-insulin resistance
- IR:
-
Insulin receptor
- IRS-1:
-
Insulin receptor substrate 1
- LDL-C:
-
Low-density lipoprotein cholesterol
- MetS:
-
Metabolic syndrome
- OGTTs:
-
Oral glucose tolerance tests
- PI3K:
-
Phosphoinositide 3-kinase
- PVDF:
-
Polyvinylidene difluoride
- TC:
-
Total cholesterol
- TG:
-
Total triglyceride
- VLDL-C:
-
Very-low-density lipoprotein cholesterol
References
Savage DB, Petersen KF, Shulman GI. Disordered lipid metabolism and the pathogenesis of insulin resistance. Physiol Rev. 2007;87(2):507–20. https://doi.org/10.1152/physrev.00024.2006.
Vendrame S, Del Bo’ C, Ciappellano S, Riso P, Klimis-Zacas D. Berry fruit consumption and metabolic syndrome. Antioxidants. 2016;5(4):34. https://doi.org/10.3390/antiox5040034.
Lipińska P, Jóźwik A. Hepatoprotective , hypoglycemic , and Hypolipidemic effect of chokeberry pomace on polish merino lambs * Pomace on Polish Merino Lambs * Anim Biotechnol 2018;29(2):136–141. doi:https://doi.org/10.1080/10495398.2017.1330209.
Chuang CC, Shen W, Chen H, et al. Differential effects of grape powder and its extract on glucose tolerance and chronic inflammation in high-fat-fed obese mice. J Agric Food Chem. 2012;60(51):12458–68. https://doi.org/10.1021/jf3028107.
Kardum N, Milovanović B, Šavikin K, et al. Beneficial effects of polyphenol-rich chokeberry juice consumption on blood pressure level and lipid status in hypertensive subjects. J Med Food. 2015;18(11):1231–8. https://doi.org/10.1089/jmf.2014.0171.
Anhê FF, Roy D, Pilon G, et al. A polyphenol-rich cranberry extract protects from diet-induced obesity, insulin resistance and intestinal inflammation in association with increased Akkermansia spp. population in the gut microbiota of mice. Gut. 2015;64(6):872–83. https://doi.org/10.1136/gutjnl-2014-307142.
Nile SH, Park SW. Edible berries: bioactive components and their effect on human health. Nutrition. 2014;30(2):134–44. https://doi.org/10.1016/j.nut.2013.04.007.
Vinson JA, Zubik L, Bose P, Samman N, Proch J. Dried fruits: excellent in vitro and in vivo antioxidants. J Am Coll Nutr. 2005;24(1):44–50. https://doi.org/10.1080/07315724.2005.10719442.
Manganaris GA, Goulas V, Vicente AR, Terry LA. Berry antioxidants: small fruits providing large benefits. J Sci Food Agric. 2014;94(5):825–33. https://doi.org/10.1002/jsfa.6432.
Vareda PMP, Saldanha LL, Camaforte NADP, Violato NM, Dokkedal AL, Bosqueiro JR. Myrcia bella leaf extract presents hypoglycemic activity via PI3k/Akt insulin signaling pathway. Evidence-Based Complement Altern Med. 2014;2014:1–11. https://doi.org/10.1155/2014/543606.
Collins B, Hoffman J, Martinez K, et al. A polyphenol-rich fraction obtained from table grapes decreases adiposity, insulin resistance and markers of inflammation and impacts gut microbiota in high-fat-fed mice. J Nutr Biochem. 2016;31:150–65. https://doi.org/10.1016/j.jnutbio.2015.12.021.
Guo S. Insulin signaling, resistance, and the metabolic syndrome: insights from mouse models into disease mechanisms. J Endocrinol. 2014;220(2):T1–T23. https://doi.org/10.1530/JOE-13-0327.
Boucher J, Kleinridders A, Kahn CR. Insulin receptor signaling in Normal and insulin-resistant states. Cold Spring Harb Perspect Biol. 2014;6(1):a009191. https://doi.org/10.1101/cshperspect.a009191.
McCracken E, Monaghan M, Sreenivasan S. Pathophysiology of the metabolic syndrome. Clin Dermatol. 2018;36(1):14–20. https://doi.org/10.1016/j.clindermatol.2017.09.004.
Arunkumar E, Karthik D, Anuradha CV. Genistein sensitizes hepatic insulin signaling and modulates lipid regulatory genes through p70 ribosomal S6 kinase-1 inhibition in high-fat–high-fructose diet-fed mice. Pharm Biol. 2013;51(7):815–24. https://doi.org/10.3109/13880209.2013.766896.
Shao J, Yamashita H, Qiao L, Friedman JE. Decreased Akt kinase activity and insulin resistance in C57BL/KsJ-Leprdb/db mice. JEndocrinol. 2000;167(0022–0795 (Print)):107–115.
Huang X-F, Chen J-Z. Obesity, the PI3K/Akt signal pathway and colon cancer. Obes Rev. 2009;10(6):610–6. https://doi.org/10.1111/j.1467-789X.2009.00607.x.
Mackenzie R, Elliott B. Akt/PKB activation and insulin signaling: a novel insulin signaling pathway in the treatment of type 2 diabetes. Diabetes, Metab Syndr Obes Targets Ther. 2014;7:55. https://doi.org/10.2147/DMSO.S48260.
Huang X, Liu G, Guo J, Su Z. The PI3K/AKT pathway in obesity and type 2 diabetes. Int J Biol Sci. 2018;14(11):1483–96. https://doi.org/10.7150/ijbs.27173.
Kulling SE, Rawel HM. Chokeberry (Aronia melanocarpa) - a review on the characteristic components and potential health effects. Planta Med. 2008;74(13):1625–34. https://doi.org/10.1055/s-0028-1088306.
Denev PN, Kratchanov CG, Ciz M, Lojek A, Kratchanova MG. Bioavailability and antioxidant activity of black chokeberry (Aronia melanocarpa) polyphenols: in vitro and in vivo evidences and possible mechanisms of action: a review. Compr Rev Food Sci Food Saf. 2012;11(5):471–89. https://doi.org/10.1111/j.1541-4337.2012.00198.x.
Jeong H, Liu Y, Kim HS. Dried plum and chokeberry ameliorate D-galactose-induced aging in mice by regulation of Pl3k/Akt-mediated Nrf2 and Nf-kB pathways. Exp Gerontol. 2017;95:16–25. https://doi.org/10.1016/j.exger.2017.05.004.
Lipińska P, Jóźwik A. Hepatoprotective, hypoglycemic, and Hypolipidemic effect of chokeberry pomace on polish merino lambs*. Anim Biotechnol. 2018;29(2):136–41. https://doi.org/10.1080/10495398.2017.1330209.
Qin B, Anderson RA. An extract of chokeberry attenuates weight gain and modulates insulin, adipogenic and inflammatory signalling pathways in epididymal adipose tissue of rats fed a fructose-rich diet. Br J Nutr. 2012;108(04):581–7. https://doi.org/10.1017/S000711451100599X.
Jurikova T, Mlcek J, Skrovankova S, et al. Fruits of black chokeberry aronia melanocarpa in the prevention of chronic diseases. Molecules. 2017;22(6):1–23. https://doi.org/10.3390/molecules22060944.
Park H, Liu Y, Kim H-S, Shin J-H. Chokeberry attenuates the expression of genes related to de novo lipogenesis in the hepatocytes of mice with nonalcoholic fatty liver disease. Nutr Res. 2016;36(1):57–64. https://doi.org/10.1016/j.nutres.2015.10.010.
Valcheva-Kuzmanova S, Borisova P, Galunska B, Krasnaliev I, Belcheva A. Hepatoprotective effect of the natural fruit juice from Aronia melanocarpa on carbon tetrachloride-induced acute liver damage in rats. Exp Toxicol Pathol. 2004;56(3):195–201. https://doi.org/10.1016/j.etp.2004.04.012.
Guo S, Duan JA, Zhang Y, et al. Contents changes of triterpenic acids, nucleosides, nucleobases, and saccharides in jujube (Ziziphus jujuba) fruit during the drying and steaming process. Molecules. 2015;20(12):22329–40. https://doi.org/10.3390/molecules201219852.
Kim SH, Choi YJ, Lee H, et al. Physicochemical properties of jujube powder from air, vacuum, and freeze drying and their correlations. J Korean Soc Appl Biol Chem. 2012;55(2):271–9. https://doi.org/10.1007/s13765-012-1039-3.
Gao Q-H, Wu C-S, Wang M. The jujube (Ziziphus Jujuba mill.) fruit: a review of current knowledge of fruit composition and health benefits. J Agric Food Chem. 2013;61(14):3351–63. https://doi.org/10.1021/jf4007032.
Shen X, Tang Y, Yang R, Yu L, Fang T, Duan J. Ao. The protective effect of Zizyphus jujube fruit on carbon tetrachloride-induced hepatic injury in mice by anti-oxidative activities. J Ethnopharmacol. 2009;122(3):555–60. https://doi.org/10.1016/j.jep.2009.01.027.
Goli-malekabadi N, Asgary S, Rashidi B, et al. The protective effects of Ziziphus vulgaris L. fruits on biochemical and histological abnormalities induced by diabetes in rats. J Complement Integr Med. 2014;11(3):171–7. https://doi.org/10.1515/jcim-2014-0010.
Solati J, Soleimani N. Antihyperglycemic and antihyperlipidemic effects of Ziziphus vulgaris L. onreptozocin-induced diabetic adult male Wistar rats. Acta Diabetol. 2010;47(S1):219–23. https://doi.org/10.1007/s00592-009-0166-8.
Abdoul-Azize S, Souleymane. Potential benefits of jujube ( Zizyphus Lotus L.) bioactive compounds for nutrition and health. J Nutr Metab 2016;2016:1–13. doi:https://doi.org/10.1155/2016/2867470.
Song E, Park H, Kim H. Additive effect of walnut and chokeberry on regulation of antioxidant enzyme gene expression and attenuation of lipid peroxidation in d-galactose-induced aging-mouse model. Nutr Res. 2018. https://doi.org/10.1016/j.nutres.2018.09.011.
Friedewald WT, Levy RI, Fredrickson DS. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin Chem. 1972;18(6):499–502 http://clinchem.aaccjnls.org/content/18/6/499.long.
Haglund O, Luostarinen R, Wallin R, Wibell L, Saldeen T. The effects of fish oil on triglycerides, cholesterol, fibrinogen and malondialdehyde in humans supplemented with vitamin E. J Nutr. 1991;121(2):165–9. https://doi.org/10.1093/jn/121.2.165.
Folch J, Lees M, Sloane Stanley GH. A simple method for the isolation and purification of total lipides from animal tissues. J Biol Chem. 1957;226(1):497–509. http://www.jbc.org/content/226/1/497.long.
Kokotkiewicz A, Jaremicz Z, Luczkiewicz M. Aronia plants: a review of traditional use, biological activities, and perspectives for modern medicine. J Med Food. 2010;13(2):255–69. https://doi.org/10.1089/jmf.2009.0062.
Li JW, Fan LP, Ding SD, Ding XL. Nutritional composition of five cultivars of chinese jujube. Food Chem. 2007;103(2):454–60. https://doi.org/10.1016/j.foodchem.2006.08.016.
Panchal SK, Brown L. Rodent models for metabolic syndrome research. J Biomed Biotechnol. 2011;2011:1–14. https://doi.org/10.1155/2011/351982.
Zhuhua Z, Zhiquan W, Zhen Y, Yixin N, Weiwei Z, Xiaoyong L. A novel mice model of metabolic syndrome : the high-fat-high-fructose diet-fed ICR mice. Exp Anim. 2015;64(4):435–42.
Alwahsh SM, Gebhardt R. Dietary fructose as a risk factor for non-alcoholic fatty liver disease (NAFLD). Arch Toxicol. 2017;91(4):1545–63. https://doi.org/10.1007/s00204-016-1892-7.
Zheng J, Shen N, Wang S, Zhao G. Oat beta-glucan ameliorates insulin resistance in mice fed on high-fat and high-fructose diet. Food Nutr Res. 2013;57(1):22754. https://doi.org/10.3402/fnr.v57i0.22754.
Jearapong N, Chatuphonprasert W, Jarukamjorn K. Effect of tetrahydrocurcumin on the profiles of drug-metabolizing enzymes induced by a high fat and high fructose diet in mice. Chem Biol Interact. 2015;239:67–75. https://doi.org/10.1016/j.cbi.2015.06.022.
Mamikutty N, Thent ZC, Sapri SR, Sahruddin NN, Mohd Yusof MR, Haji Suhaimi F. The establishment of metabolic syndrome model by induction of fructose drinking water in male Wistar rats. Biomed Res Int. 2014;2014:1–8. https://doi.org/10.1155/2014/263897.
Mamikutty N, Thent ZC, Haji Suhaimi F. Fructose-drinking water induced nonalcoholic fatty liver disease and ultrastructural alteration of hepatocyte mitochondria in male Wistar rat. Biomed Res Int. 2015;2015:1–7. https://doi.org/10.1155/2015/895961.
Dissard R, Klein J, Caubet C, et al. Long term metabolic syndrome induced by a high fat high fructose diet leads to minimal renal injury in C57BL/6 mice. Feng W, ed. PLoS One. 2013;8(10):e76703. https://doi.org/10.1371/journal.pone.0076703.
Zaman M-Q, Leray V, Le Bloc’h J, Thorin C, Ouguerram K, Nguyen P. Lipid profile and insulin sensitivity in rats fed with high-fat or high-fructose diets. Br J Nutr. 2011;106(S1):S206–10. https://doi.org/10.1017/S0007114511004454.
Meydani M, Hasan ST. Dietary polyphenols and obesity. Nutrients. 2010;2(7):737–51. https://doi.org/10.3390/nu2070737.
Wada T, Kenmochi H, Miyashita Y, et al. Spironolactone improves glucose and lipid metabolism by ameliorating hepatic steatosis and inflammation and suppressing enhanced gluconeogenesis induced by high-fat and high-fructose diet. Endocrinology. 2010;151(5):2040–9. https://doi.org/10.1210/en.2009-0869.
Peng C-H, Liu L-K, Chuang C-M, Chyau C-C, Huang C-N, Wang C-J. Mulberry water extracts possess an anti-obesity effect and ability to inhibit hepatic lipogenesis and promote lipolysis. J Agric Food Chem. 2011;59(6):2663–71. https://doi.org/10.1021/jf1043508.
Park CH, Kim JH, Lee EB, et al. Aronia melanocarpa extract ameliorates hepatic lipid metabolism through PPARγ2 downregulation. PLoS One. 2017;12(1). https://doi.org/10.1371/journal.pone.0169685.
Shao W, Yu Z, Chiang Y, et al. Curcumin prevents high fat diet induced insulin resistance and obesity via attenuating lipogenesis in liver and inflammatory pathway in adipocytes. Schneider-stock R. PLoS One. 2012;7(1):e28784. https://doi.org/10.1371/journal.pone.0028784.
Riccardi G, Giacco R, Rivellese A. Dietary fat, insulin sensitivity and the metabolic syndrome. Clin Nutr. 2004;23(4):447–56. https://doi.org/10.1016/j.clnu.2004.02.006.
Mei S, Yang X, Guo H, et al. A small amount of dietary carbohydrate can promote the HFD-induced insulin resistance to a maximal level. Zang M, ed PLoS One 2014;9(7):e100875. doi:https://doi.org/10.1371/journal.pone.0100875.
Saltiel AR, Kahn CR. Insulin signalling and the regulation of glucose and lipid metabolism. Nature. 2001;414(6865):799–806. https://doi.org/10.1038/414799a.
Liu H-Y, Hong T, Wen G-B, et al. Increased basal level of Akt-dependent insulin signaling may be responsible for the development of insulin resistance. AJP Endocrinol Metab. 2009;297(4):E898–906. https://doi.org/10.1152/ajpendo.00374.2009.
Xu H, Zhou Y, Liu Y, et al. Metformin improves hepatic IRS2/PI3K/Akt signaling in insulin-resistant rats of NASH and cirrhosis. J Endocrinol. 2016;229(2):133–44. https://doi.org/10.1530/JOE-15-0409.
Arunkumar E, Karthik D, Anuradha CV. Genistein sensitizes hepatic insulin signaling and modulates lipid regulatory genes through p70 ribosomal S6 kinase-1 inhibition in high-fat-high-fructose diet-fed mice. Pharm Biol. 2013;51(7):815–24. https://doi.org/10.3109/13880209.2013.766896.
Lee S, Kim M-B, Kim C, Hwang J-K. Whole grain cereal attenuates obesity-induced muscle atrophy by activating the PI3K/Akt pathway in obese C57BL/6N mice. Food Sci Biotechnol. 2018;27(1):159–68. https://doi.org/10.1007/s10068-017-0277-x.
Rodriguez Lanzi C, Perdicaro DJ, Antoniolli A, et al. Grape pomace and grape pomace extract improve insulin signaling in high-fat-fructose fed rat-induced metabolic syndrome. Food Funct. 2016;7(3):1544–53. https://doi.org/10.1039/c5fo01065a.
Matsuda S, Kobayashi M, Kitagishi Y. Roles for PI3K/AKT/PTEN pathway in cell signaling of nonalcoholic fatty liver disease. ISRN Endocrinol. 2013;2013(Figure 1):472432. https://doi.org/10.1155/2013/472432.
Shin S-K, Cho H-W, Song S-E, Song D-K. Catalase and nonalcoholic fatty liver disease. Pflügers Arch - Eur J Physiol. 2018;470(12):1721–37. https://doi.org/10.1007/s00424-018-2195-z.
Góth L, Rass P, Páy A. Catalase enzyme mutations and their association with diseases. Mol Diagn 2004;8(3):141–149. doi:1084–8592.
Wang S, Meckling KA, Marcone MF, Kakuda Y, Tsao R. Synergistic, additive, and antagonistic effects of food mixtures on total antioxidant capacities. J Agric Food Chem. 2011;59(3):960–8. https://doi.org/10.1021/jf1040977.
Yang Y, Li W, Li Y, Wang Q, Gao L, Zhao J. Dietary Lycium barbarum polysaccharide induces Nrf2/ARE pathway and ameliorates insulin resistance induced by high-fat via activation of PI3K/AKT signaling. Oxidative Med Cell Longev. 2014;2014:1–10. https://doi.org/10.1155/2014/145641.
Ceriello A, Motz E. Is oxidative stress the pathogenic mechanism underlying insulin resistance, diabetes, and cardiovascular disease? The common soil hypothesis revisited. Arterioscler Thromb Vasc Biol. 2004;24(5):816–23. https://doi.org/10.1161/01.ATV.0000122852.22604.78.
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
Authors of this research paper have directly participated in the planning, execution, or analysis of this study. Author contributions are as follows: study design, OJ and HK; experiment, OJ; data analysis, OJ; and writing of the manuscript, OJ and HK.. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
All animal procedures were approved by the Animal Ethical Committee of Sookmyung Women’s University for the care and use of laboratory animals.
Consent for publication
Not applicable.
Competing interests
The authors declare they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Jeong, O., Kim, HS. Dietary chokeberry and dried jujube fruit attenuates high-fat and high-fructose diet-induced dyslipidemia and insulin resistance via activation of the IRS-1/PI3K/Akt pathway in C57BL/6 J mice. Nutr Metab (Lond) 16, 38 (2019). https://doi.org/10.1186/s12986-019-0364-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12986-019-0364-5