
Abstract
Aromatic polyketides have attractive biological activities and pharmacological properties. Different from other polyketides, aromatic polyketides are characterized by their polycyclic aromatic structure. The biosynthesis of aromatic polyketides is usually accomplished by the type II polyketide synthases (PKSs), which produce highly diverse polyketide chains by sequential condensation of the starter units with extender units, followed by reduction, cyclization, aromatization and tailoring reactions. Recently, significant progress has been made in characterization and engineering of type II PKSs to produce novel products and improve product titers. In this review, we briefly summarize the architectural organizations and genetic contributions of PKS genes to provide insight into the biosynthetic process. We then review the most recent progress in engineered biosynthesis of aromatic polyketides, with emphasis on generating novel molecular structures. We also discuss the current challenges and future perspectives in the rational engineering of type II PKSs for large scale production of aromatic polyketides.
Similar content being viewed by others


Background
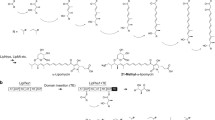
Polyketides are structurally diverse and biologically active secondary metabolites derived from natural sources such as animals, plants, fungi and bacteria [1]. These compounds are widely used as clinical medicines for the treatment of various acute and chronic diseases [2]. Examples include antibacterial (erythromycin and tetracycline) [3], antitumor (anthracycline and doxorubicin) [4], antifungal (amphotericin and griseofulvin) [5], antiparasitic (avermectin) [6] and anti-cholesterol (lovastatin) [7] drugs. Polyketides are synthesized by polyketide synthases (PKSs) which are multi-domain enzymes or enzyme complexes [8], consisting of acyltransferase (AT), ketosynthase (KS), thioesterase (TE) and optional domains. The biosynthesis of polyketides is initiated by loading the starter unit acyl-Coenzyme A (CoA) on the acyl carrier protein (ACP) catalyzed by the AT domain [2]. The KS domain subsequentially elongates the carbon chain by decarboxylative Claisen condensation. The β-keto group can be further modified to generate different polyketide structures by additional domains, including ketoreductase (KR), dehydratase (DH) and enoylreductase (ER) domains. Finally, the TE domain hydrolyzes or cyclizes the completed polyketide chain from the ACP-domain to terminate the elongation [9]. Although sharing a similar synthetic process, PKSs can be classified into three types, type I, II, and III PKSs (Fig. 1) [10, 11]. Type I PKSs are multifunctional peptides containing linearly arranged and covalently fused domains. The type I PKSs can be further classified into iterative type I PKSs (iPKSs) (Fig. 1a) and modular type I PKSs (mPKSs) (Fig. 1b) [12]. iPKSs are mainly found in fungi and the domains are used repeatedly to catalyze multiple rounds of elongation [13, 14]. While, mPKSs are primarily found in bacteria and function in an assembly line similar to the nonribosomal peptide synthases [15]. mPKSs consist of several domains with defined functions that are separated by short spacer regions. The distinct domains work cooperatively and non-iteratively to catalyze the carbon chain elongation and functional group regeneration [16]. Type II PKSs are multi-enzyme complexes composed of monofunctional proteins (Fig. 1c). They are found predominantly in bacteria and produce diverse aromatic polyketides [17]. Different from both type I and type II PKSs, type III PKSs are mainly found in plants as simple homodimers that use CoA rather than ACP as an anchor for chain extension (Fig. 1d) [18], while both type II and type III PKSs are iterative.

Catalytic reaction of PKSs. a Iterative type I PKSs; b Modular type I PKSs; c Type II PKSs; d Typical type III PKSs use acyl-CoA as the starter unit and malonyl-CoA as the extender unit. KS ketoacyl synthase, AT acyltransferase, DH dehydratase, ER enoylreductase, KR ketoreductase, ACP acyl carrier protein, TE thioesterase, CYC cyclase, ARO aromatase, CHS chalcone synthase, STS stilbene synthase, AS acridone synthase
Type II PKSs are mainly responsible for producing aromatic polyketides by catalyzing iterative Claisen condensation reaction usually using acetate as the starter unit [19]. Aromatic polyketides are polycyclic compounds harboring at least one aromatic ring [17]. They are an important type of natural products with antibacterial, anticancer and antiviral bioactivities, with representative examples being the above-mentioned drugs tetracyclines and anthracyclines [3, 4]. The clinical and environmental potential of aromatic polyketides has attracted increasing attention from researchers to conduct studies on the biosynthesis of these polyketides. The production process of aromatic polyketides includes the following reactions. (1) α-Carboxylated precursor loading: acetate is loaded onto ACP, forming acyl-ACP. (2) Iterative chain extension and ketone reduction: acyl-ACP is transferred to the KS subunit and iteratively elongated with the extender unit malonyl-CoA to form the poly-β-keto chain. (3) Cyclization and/or aromatization: the poly-β-keto chain is catalyzed by aromatases (ARO) and oxygenases to generate the aromatic polyketide core. (4) Post-modification: oxygenases, methyltransferases and glycosyltransferases catalyze this aromatic polyketide core to generate the final product [17, 20]. The characteristic of aromatic polyketides biosynthesis is the employment of a set of iteratively used enzymes. This set of enzymes is called minimal type II PKS, which contains KSα, chain length factors (CLF or KSβ) and ACP subunits. The sequences of KSα and CLF components are highly similar, except for a cysteine-containing active site in KSα that is essential for assembling aromatic polyketides [17, 21]. A minimal PKS can solely produce some aromatic polyketides, for instance, SEK4, SEK 4b and dehydro-SEK4b [22].
Although type II PKSs and aromatic polyketides are a tribute to our life, there have been only a few articles focusing on the understanding and engineering of type II PKSs probably due to the complicated catalytic mechanisms. In contrast, type I and type III PKSs have been well-characterized and engineered to produce various bioactive compounds [18, 23, 24]. Many excellent reviews have summarized the research progress of these PKSs [18, 25,26,27,28,29]. With the development of biotechnology and emerged characterization of type II PKSs, great progress has been made in recent years to optimize type II PKSs for high-level production of aromatic polyketides. Meanwhile, with the rising issue of antibiotic and anticancer drug resistance, developing of diverse polyketides with new bioactivity is of particular interest. In this paper, we will briefly review the recent efforts on improving the production and broadening the spectrum of aromatic polyketides.
Production of aromatic polyketides by employing different starter units
The first step of aromatic polyketide biosynthesis is loading of a starter unit, mostly being acetate, onto ACP. A typical case is the biosynthesis of kinamycins, a group of bacterial polyketide secondary metabolites containing a diazo group [30], which begins with the condensation of 1 acetyl-CoA and 9 malonyl-CoA molecules to form the intermediate dehydrorabelomycin [31]. While only a few type II PKSs have been identified to use other starter units, the employment of different starter units could provide additional structurally diverse products. Propionyl-CoA has been reported to be the starter unit for several aromatic polyketides. For instance, lomaiviticins, an important class of diazo-containing aromatic polyketides, have gained great interest because of their antibiotic and antitumor activities [32]. Waldman and his colleagues identified a gene cluster responsible for the biosynthesis of lomaiviticin in Salinispora pacifica DPJ-0019. This cluster contains a bifunctional enzyme (encoded by Lom62) that has both acyltransferation and decarboxylation activities and can catalyze the conversion of methylmalonyl-CoA to propionyl-CoA, which serves as a starter unit for lomaiviticin production (Fig. 2a) [19]. In addition, Otten et al. identified a gene cluster dnr encoding type II PKS which can also employ propionyl-CoA as the starter unit for the production of daunorubicin and doxorubicin, two famous anthracycline topoisomerase inhibitors, in Streptomyces spp. [33]. However, expression of this cluster was found restrained by the lack of bldA-tRNA in S. peucetius to read a rare TTA codon in dnrO, which is a transcriptional activator involved in the biosynthesis of daunorubicin. For purpose of activating daunorubicin biosynthesis, bldA-tRNA was heterologously over-expressed in S. peucetius. The productivity of daunorubicin was increased by 45.7% compared to that of wild type S. peucetius strain [34]. To obtain a more efficient doxorubicin producer, Wang et al. employed UV and ARTP (atmospheric and room temperature plasma) approaches to mutate doxorubicin producer strains. With the aid of fermentation condition optimization, the titer of doxorubicin dramatically increased from 119 mg/L to 1.1 g/L in a 5-L fermenter, being the highest reported so far [35]. Those researches offered a new starter unit for the generation of aromatic polyketides and represented the potential of large-scale production of daunorubicin.

Biosynthesis of lomaiviticin and trioxacarcin A using different starter units. Type II PKSs can catalyze different starter units to synthesize different products
In addition to propionyl-CoA, malonamate (3-amino-3-oxopropanoate) has been reported as the starter unit for the biosynthesis of oxytetracycline, a broad-spectrum antibiotic belonging to the tetracycline family [21, 36]. Malonamate is converted from malonyl-CoA by aminotransferase OxyD and thiolase OxyP. Then, oxytetracycline was produced after a series of reactions catalyzed by 19 enzymes encoded by oxy gene cluster, with the addition of extender units malonyl-CoA [37]. Based on these studies, heterologous production of oxytetracycline was achieved recently. Stevens et al. successfully engineered E. coli to produce 2 mg/L oxytetracycline by co-expressing the biosynthesis gene cluster with an alternative sigma factor σ54 from the oxytetracycline native producer S. rimosus [38]. A fast-growing strain S. venezuelae WVR2006 was also chosen as the producer to accumulate oxytetracycline. The titer of oxytetracycline reached 431 mg/L within 48 h by overexpressing the oxy gene cluster [39]. In order to further increase the production of oxytetracycline, S. rimosus strain was engineered for enhanced self-resistance level by overexpression of the ribosomal protection protein and two efflux proteins, leading to a remarkable increase in titer of 7.59 g/L oxytetracycline in shake flasks [40]. By assembling oxytetracycline and chelocardin biosynthetic pathways, Lešnik et al. obtained 2-carboxamido-2-deacetyl-chelocardin (CDCHD), a new class of tetracycline that uses malonamate as the starter unit. Compared to chelocardin, the new antibiotic CDCHD has a carboxamido moiety and showed higher in vitro activities against clinical isolates [41].
In addition to propionyl-CoA and malonamate, several other starter units have been employed to produce novel aromatic polyketides. Recently, 2-methylbutyryl-CoA was found to initiate the scaffold trioxacarcins biosynthesis by Zhang et al. via 13C-labeling experiments (Fig. 2b) [42]. This research provides an opportunity to get more potential drugs by identifying a possible new starter unit. A further study showed that p-nitrobenzoic acid was synthesized from the native intermediate p-aminobenzoic acid catalyzed by a non-heme di-iron monooxygenase AurF [43]. Additionally, the structural diversity of polyketides can also be expanded by utilizing modified amino acids as the starter units. 4,5-Dichloropyrrolyl-S-carrier protein was found to be the starter unit of pyoluteorin biosynthetic in Pseudomonas fluorescens Pf-5 [44]. The discovery of more distinct starter units and their corresponding native producers can facilitate the production of various aromatic polyketides.
Production of aromatic polyketides by varying chain length
In addition to altering starter units, changing the chain length can also yield a variety of polyketides. Different from type I and type III PKSs, type II PKSs are aggregates of mono-functional proteins, containing a KSα-CLF complex that can determine the carbon chain length of aromatic polyketides by controlling the number of Claisen condensations [45]. Previous study showed that act KSα-CLF or tcm KSα-CLF act as the chain length determination subunit for the synthesis of C-16 or C-20 chain length polyketides, respectively [1]. In order to further understand the mechanism of structural determination, both act KSα-CLF and tcm KSα-CLF were separated into 12 cassettes (6 KSα and 6 CLF parts). KSα-CLF hybrids were then constructed by single replacements with mutual counterparts based on sequence homology. The results showed that cassette D in KSα, cassette C and D in CLF are the primary structural determinants for carbon chain length [46].
To date, type II PKSs naturally synthesizing aromatic polyketides with chain length of C-10 to C-30 have been reported [17, 47]. To our surprise, an increasing number of new aromatic polyketides with different carbon chain lengths have been continuously explored in recent years. One C-13 and two new C-18 aromatic polyketides huanglongmycin, HLM A, HLM B and HLM C, were identified from Streptomyces sp. CB09001 which was isolated from Xiangxi, China. HLM A-C showed broad antibacterial spectrum and HLM A also exhibited antitumor activity with moderate cytotoxicity [48]. Saccharothrixones A–D are synthesized by Saccharothrix sp. 10-10. The molecular formula of saccharothrixones A is C23H22O11 and the other three compounds are C-24 polyketides. Remarkably, saccharothrixone D displayed in vitro antitumor activity [49]. These studies enlarged the spectrum of aromatic polyketides and offered new potential drugs. In some cases, KSα-CLF contributes to longer chains formation for production of new aromatic polyketieds. SanFG is the dedicated KSα-CLF to generate A-74528, a C-30 aromatic polyketide. A-74528 is an antiviral drug owing to its inhibitory effect on the regulatory enzyme 2′,5′-oligoadenylate phosphodiesterase (2′-PDE) [50]. The gene cluster in Streptomyces sp. SANK 61196 responsible for the biosynthesis of A-74528 was identified and sequenced. Gene samF and samG encode KSα and CLF, respectively. The introduction of this gene cluster into S. lividans K4-114 led to only trace amount of A-74528 (less than 1 mg/L) [51]. This titer was further increased to 3 mg/L by identifying the essential pathway genes (sanFGH, sanI, sanJ and sanS2C) and refactoring the minimal biosynthetic pathway in the host strain S. coelicolor CH999 [52]. Although the titer of A-74528 is still low, those efforts offer an opportunity for further studying on enhancement of its titer.
The variation of actinorhodin chain length has been studied by engineering its KSα-CLF complex. Actinorhodin, a C-16 polyketide with multiple bioactivities, is the most-characterized type II PKSs product found in S. coelicolor A3 [53]. The gene cluster act is responsible for the synthesis of actinorhodin and act KSα-CLF contributes to the chain length determination [53, 54]. On this basis, a set of act KSα-CLF mutants were constructed to change amino acid (residues 109, 112, and 116) residue size by employing site-directed mutagenesis. The results showed a smaller size of those amino acid residues in act CLF led to longer reaction channel and made two additional of Claisen condensations possible. New aromatic polyketides with the carbon chain length of C-20 and C-24 were produced by the host strain carrying the obtained act CLF mutants [55]. This research provided a new direction for the rational design of CLF subunit to alter the chain length of aromatic polyketides. In recent years, some efforts were focused on developing strategies of improvement the production of actinorhodin. For example, the TraA-like protein ZouA and its recombination sites RsA and RsB that are essential for the amplification of kan gene cluster in S. kanamyceticus, were inserted into S. coelicolor genome flanking act gene cluster. The strains with increased copy numbers of act gene cluster were selected by stress-based screening and resulted in 20-fold-increase of actinorhodin yield [56]. An xdhR-encoded inhibitor, which negatively regulates actinorhodin biosynthesis through binding the intergenic region of xdhABC, were deleted for enhanced production of actinorhodin by 2.5 fold [57]. Lysoquinone-TH1 has antibacterial activity as well as pharmaceutical effects towards asthma (by inhibiing the target protein, phosphodiesterase 4) and chronic obstructive pulmonary disease. With the expression of llpE, encoding for lysolipin CLF, and other minimal type II PKSs genes of lysolipin, lysoquinone-TH1 was successfully synthesized in S. albus after condensing 13 acetate extenders. When condensing 15 acetate extenders, lysoquinone-TH2 was produced [58]. This study provides a promising strategy for the production of non-natural analogs derived from natural compounds utilizing different numbers of extenders. In addition to engineering the KSα-CLF complex, other approaches to achieve variant chain length of aromatic polyketides have been explored. For instance, oxytetracycline is a C-19 aromatic polyketide, whose chain length can be altered by disrupting an anhydrotetracycline oxygenase to generate a new C-17 polyketide [59]. Recently, Du et al. reported that IgaPKS, which belonged to a new subfamily of type II PKSs, can synthesize unsaturated carboxylic acids with carbon chain lengths of C-8, C-10, C-12 and C-14 [60]. This research demonstrates a new opportunity to employ type II PKSs for the production of carboxylic acids. The discovery and characterization of aromatic polyketide synthesis complexes for different chain lengths is a promising strategy to created more novel aromatic polyketides.
Production of aromatic polyketides by combining tailoring reactions
The complexity and diversity of aromatic polyketides with new bioactivities can be endowed by tailoring reactions. The common tailoring reactions involved in the biosynthesis of aromatic polyketides are oxidation, methylation and glycosylation. Understanding and engineering of such modification processes can help derive different new aromatic polyketides by rationally combining the tailoring reactions.
Hydroxylation, epoxidation, dehydrogenation, quinone formation and oxidative rearrangement are the major oxidative tailoring reactions. The corresponding oxygenases include flavin-dependent oxygenases, anthrone-type oxygenases and cytochrome P450-dependent monooxygenases [17, 20]. EncM, a FAD-dependent oxygenase, catalyzes oxidative Favorskii-type rearrangement reaction which is a pivotal step in enterocin biosynthesis. This enzyme has a broad substrate spectrum and can also generate 5-deoxyenterocin and wailupemycin analogues [61]. The antitumor drug mithramycin A can be converted from premithramycin B, a tetracyclic intermediate, through Baeyer–Villiger oxygenation catalyzed by a FAD-dependent oxygenase, MtmOIV [62]. To improve the production of mithramycin A, the whole gene cluster was constructed into S. lividans TK24 along with the deletion of interfering secondary metabolite genes. The best strain produced 3 g/L mithramycin A [63]. Seven mithramycin analogues were generated by using combinatorial biosynthesis strategy in S. argillaceus M7C1 [64]. One of these new compounds, EC-8042, was identified to enhance the inhibitory activity towards Ewing sarcoma and showed higher inhibition activity and lower toxicity than mithramycin A [65]. This study exhibited the promising application of aromatic polyketide derivatives.
Anthrone-type oxygenases are responsible for the oxidation of anthrone-type intermediates to the related quinones without cofactors or metal ions. Shen et al. first reported an anthrone-type monooxygenase (TcmH) that can convert naphthacenone tetracenomycin F1 into 5,12-naphthacenequinone tetracenomycin D3 [66]. ActVA-orf6 and AknX are well-characterized anthrone-type monooxygenases involved in quinone-forming reactions [67, 68]. The crystal structure of ActVA-orf6 was determined by Sciara and co-workers. Based on the structural similarity, ForS and ForU from Lechevalieria fradiae and Pseudonocardiales bacteria belonging to ABM superfamily were characterized as the monooxygenase for the biosynthesis of fasamycin [69]. This study uncovered a new role of monooxygenases in the metabolism of aromatic polyketides.
On the other hand, there have been few cytochrome P450-dependent monooxygenases identified in bacteria until now. EncR and DoxA from S. maritimus and S. peucetius, respectively, have been reported to assist in the conversion of deoxyenterocin into enterocin. To catalyze this reaction, EncR requires ferredoxin and DoxA needs NADPH as cofactors [70, 71]. Furthermore, DoxA also contributes to the biosynthesis of doxorubicin, daunorubicin as well as anthracycline and its analogues [71].
Methylation is another major type of tailoring reaction that aids in expanding the diversity of aromatic polyketides. The methyltransferases (MTs)-mediated reactions usually transfer a methyl group from a donor such as S-adenosyl-l-methionine (SAM) to the C, N or O atoms on acceptor targets. MTs are categorized into C-MTs, O-MTs and N-MTs by which of those atoms are modified. DutMT1, located in the dut gene cluster was identified from S. minoensis NRRL B-5482 by genome sequencing. To further study the function of this gene, aiming gene deletion combined with intermediates identification experiments were performed. This enzyme was then identified as a C-MT that transfers a methyl group to 12-desmethyl-dutomycin to yield dutomycin [72]. Two C-MTs were found to involve in the biosynthesis of polyketomycin (POK), an antitumor aromatic polyketide in S. diastatochromogenes Tü6028 (Fig. 3). PokMT2 is able to methylate the substrate at C6 position, while PokMT1 is responsible for methylation of another intermediate 6-methylsalicylyl-CoA to form 3,6-DMSA moiety of the POK molecule [73] (Fig. 3). In order to understand the reaction process of POK biosynthesis, Guo et al. characterized PokMT2 and an ATP-dependent CoA ligase, PokM3, and provided an unusual way for the generation of POK [74] (Fig. 3). This study paved the way for investigation of the catalytic mechanisms of C-MTs in polyketides biosynthesis.

A new pathway logic for biosynthesis of polyketomycin. SAM S-adenosyl methionine, PokC1-C3 cyclase, PokMT2 C-methyltransferase, PokM3 6-methylsalicylic acid-AMP ligase, PokMT1 C-methyltransferase
Since O-MTs from microorganisms commonly have a wide substrate scope, various substrates can be introduced to expand the library of aromatic polyketides. Three O-MTs: TcmO, TcmN and TcmP were reported to participate in tetracenomycin (Tcm) biosynthesis. TcmO methylates C8 hydroxy group of Tcm D3 to generate 8-O-methyl-Tcm D3, which can then be modified by TcmN and TcmP individually. TcmN adds a methyl group on C3 hydroxy group of 8-O-methyl-Tcm D3 forming Tcm E. TcmN can also methylate Tcm D3 to generate Tcm B3. While TcmP transfers a methyl group to C9 carboxyl yielding 9-carboxymethyl-8-O-methyl-Tcm D3, respectively. Likewise, TcmP can also accept Tcm B3 as the substrate and convert it to 9-carboxymethyl-Tcm B3. The broad substrate spectrum of these O-MTs shows the potential for producing diverse Tcms [75].
DnrK is another well-studied O-MT that is SAM-dependent and was isolated from S. peucetius. It transfers a methyl group to the 4-hydroxyl group of carminomycin and its analogues to synthesize the famous anticancer drugs, daunorubicin and its derivatives. The crystal structure of DnrK has been determined by Jansson et al. [76], which provides important information for rational protein engineering of this enzyme. Based on the crystal structure, a mutant DnrK was constructed by replacing its R1 region with the counterpart of a hydroxylase RdmB. The mutant DnrK exhibited both methylase and monooxygenase activities. This chimera was further used to produce a novel anthracycline [77]. Meanwhile, DnrK was employed to produce novel flavonoids depending on the structural similarity of substrates. Surprisingly, DnrK can accept luteolin, luteolin 4′-glucoside, kaempferol, kaempferol-3-O-glucoside and apigenin as the substrate to form their corresponding methylated compounds [78]. These results represented the flexibility of DnrK towards substrates and expanded the spectrum of polyketides. More O-MTs were identified along with new aromatic polyketides discovery. Three O-MTs were deduced to participate in the biosynthesis of four paramagnetoquinones (Pmq A, Pmq B, Pmq C and Pmq D) from Actinoallomurus by genome sequencing and amino acids sequence alignment [79]. In order to alleviate the issue of drug-resistant bacteria, a new antibiotic enduracyclinone was discovered from Nonomuraea sp. ID40491. This molecule contains methoxy groups and three putative O-MTs were proposed to form the methoxy groups [80]. Although further characterization of these new O-MTs is pending, these studies provided possibilities for expanding the variety of aromatic polyketides by employing new O-MTs.
Compared to O-methylation, N-methylation is less common in the biosynthesis of aromatic polyketides. In 2006, Zhang et al. identified a gene cluster that is responsible for the biosynthesis of oxytetracycline. OxyT was described as a N-MT putatively, because it exhibited high amino acid sequence identity to O-MTs [37]. To further understand the catalytic property of OxyT, 4-amino anhydrotetracycline (ATC) was used as the substrate and incubated with or without purified OxyT in the reaction system. The HPLC/MS result showed that ATC was produced only in the presence of OxyT (Fig. 4a). Remarkably, monomethylated 4-amino ATC was formed (Fig. 4a) as an intermediate when the reaction mixture was stopped at 10 min. Those experiments demonstrated that OxyT is a N-MT that can catalyze both mono- and di-methylating reactions [81] (Fig. 4a). Lukežic et al. designed biosynthetic pathways of 5 novel chelocardin (CHD) analogues in Amycolatopsis sulphurea via combining OxyT with an aminotransferase and a glycosyltransferase (Fig. 4b and Fig. 4c) [82]. This research showed the potential of OxyT in the biosynthesis of innovative aromatic polyketides.

N-methyltransferase OxyT catalyzes mono- and di- methylation. a OxyT involved in biosynthesis of ATC. b OxyT was employed to catalyze epi-CHD generating new compound N,N-dimethyl-epi-CHD. c New compound was produced by employing OxyT and GT. ATC anhydrotetracycline, SAM S-adenosyl methionine, OxyT N-methyltransferase, CHD chelocardin, GT glycosyltransferase
In addition to oxidation and methylation, glycosylation also tremendously enhances the diversity of aromatic polyketides. The major glycosylation enzyme is glycosyltransferases (GTs) [83]. UrdGTs were isolated from S. fradiae Tü2717 for the biosynthesis of urdamycin. Among the identified UrdGTs encoding genes (urdGT1a, urdGT1b, urdGT1c and urdGT2) by Trefzer et al., UrdGT2 transfers an olivose moiety from dNDP-olivose to rabelomycin and its analogue to generate urdamycinone B and aquayamycin, respectively [84]. This is the first glycosylating step during urdamycin biosynthesis. Then, UrdGT1c and UrdGT1b transfer a rhodinose moiety and an olivose moiety to urdamycinone B sequentially for the generation of compound 100-1 and urdamycin B. UrdGT1c glycosylate aquayamycin to form 100-2, which is the second step involved in the biosynthesis of urdamycin A. Then, UrdGT1c along with UrdGT1b add a rhodinose moiety and an olivose moiety on 100-2 to generate urdamycin A. Four more novel urdamycin derivatives can be synthesized by using various UrdGT1s combinations [85]. This work discovered the function of UrdGT1s and exhibited great potential for expanding the scope of aromatic polyketides by glycosylation. Based on this research, nine novel urdamycins N1–N9 were isolated from S. diastaticus subsp. SCSIO GJ056, among which urdamycins N7 and N8 containing (5R, 6R)-angucycline glycosides were discovered for the first time [86]. Saprolmycin A-E are antibiotics isolated from Streptomyces sp. TK08046. Three GTs, SprGT1-3, were identified from this producer strain that transfer aculose, rhodinose and olivose moiety to UWM6 (the core structure of saprolmycin), respectively. However, the substrates of these three GTs have not yet to be discovered [87]. As mentioned above, dnr gene cluster contributes to biosynthesize daunorubicin. DnrS (encoded by dnrS) transfers a daunosamine moiety to rhodomycinne. The latter was converted to daunorubicin catalyzed by DnrP and DnrK [88]. To improve the diversity of daunorubicin family, Chu et al. designed one-pot enzymatic UDP-recycling glycosylation. Two new daunorubicinone glucosides, 7-O-alpha-D glucoside and 7,9-di-O-alpha-D glucoside were generated. These two compounds showed higher thermal stability at 100 °C and tolerance to pH range 4.5–8.5 than daunorubicin [89].
On the contrary of introducing and combining tailoring reactions, inactivation of the corresponding enzymes can also provide some derivatives. The above-mentioned mithramycin can be oxygenated by oxygenase to produce new derivatives. Surprisingly, four new mithramycin analogs could also be obtained by disruption of two genes encoding GTs [90]. Similarly, to explore novel compounds belonging to the trioxacarcin family, Zhang et al. deactivated seven tailoring enzymes including cytochrome P450-dependent oxygenases, MTs and GTs. Eleven trioxacarcin derivatives were generated by the mutants. To further appraise the bioactivity of these analogues, cytotoxicity assays were performed by employing Jurkat cells. The new compound 14a showed similar IC50 value with trioxacarcin [42]. These researches provided some potential drug candidates and represent a new direction to improve the diversity of aromatic polyketides. Frigocyclinone was found in S. griseus NTK 97 which has antitumor and antibacterial activities. Recently, Mo et al. successfully identified its biosynthetic gene cluster by gene inactivation. An anti-MRSA anthraquinone tetrangomycin was produced by inactivating the gene frig1 encoding the GT [91]. This study offered another pathway to produce tetrangomycin and its analogs.
Conclusions
Natural products derived from plants, animals and microorganisms are widely used in food, cosmetic, chemical and pharmaceutical industries. Aromatic polyketides are a special group of natural products synthesized by PKSs and have remarkable bioactivities. The last few decades have witnessed a rapid growth of genetic information acquirement on type II PKSs and an improved understanding of the biological and chemical mechanisms of aromatic polyketide biosynthesis. All the knowledge not only provides exciting opportunities for the rational design of microbial cell factories for high-level biosynthesis of polyketides, but also facilitates the generation of more novel polyketides including “unnatural” derivatives. In order to achieve economically viable production and continually expending the palette of aromatic polyketides, we anticipate the future efforts focused on the following research directions: (1) thorough characterization of catalytic machinery of aromatic polyketides on different levels, such as structural and genetic levels; (2) discovery of more promising aromatic polyketides based on enzyme and gene bioprospecting; (3) production of new aromatic polyketides with improved or novel bioactivities by using combinatorial biosynthesis approach; (4) improvement of catalytic activity of PKSs through protein engineering; (5) exploration and activation of the positive regulatory factor of gene cluster by genetic engineering. We expect that with the continued and rapid development of advanced technologies and tools, more robust and efficient microbial cell factories can be established to achieve economically biosynthesis of type II aromatic polyketides.
Availability of data and materials
Not applicable.
Abbreviations
- PKSs:
-
Polyketide synthases
- AT:
-
Acyltransferase
- KS:
-
Ketosynthase
- TE:
-
Thioesterase
- CoA:
-
Coenzyme A
- ACP:
-
Acyl carrier protein
- KR:
-
Ketoreductase
- DH:
-
Dehydratase
- ER:
-
Enoylreductase
- iPKSs:
-
Iterative type I PKSs
- mPKSs:
-
Modular type I PKSs
- ARO:
-
Aromatases
- CLF or KSβ :
-
Chain length factors
- CDCHD:
-
2-carboxamido-2-deacetyl-chelocardin
- HLM:
-
Huanglongmycin
- 2′-PDE:
-
2′,5′-Oligoadenylate phosphodiesterase
- MTs:
-
Methyltransferases
- SAM:
-
S-adenosyl-l-methionine
- GTs:
-
Glycosyltransferases
- POK:
-
Polyketomycin
- Pmq:
-
Paramagnetoquinones
References
Hong F, Hopwood DA, Khosla C. Engineered biosynthesis of novel polyketides. Science. 1993;262:1546–50.
Piel J. Biosynthesis of polyketides by trans-AT polyketide synthases. Nat Prod Rep. 2010;27:996–1047.
Fang L, Guell M, Church GM, Pfeifer BA. Heterologous erythromycin production across strain and plasmid construction. Biotechnol Prog. 2018;34:271–6.
Katz L, Baltz RH. Natural product discovery: past, present, and future. J Ind Microbiol Biotechnol. 2016;43:155–76.
Klaus M, Grininger M. Engineering strategies for rational polyketide synthase design. Nat Prod Rep. 2018;35:1070–81.
Barajas JF, Blake-Hedges JM, Bailey CB, Curran S, Keasling JD. Engineered polyketides: synergy between protein and host level engineering. Synth Syst Biotechnol. 2017;2:147–66.
Sundaram S, Hertweck C. On-line enzymatic tailoring of polyketides and peptides in thiotemplate systems. Curr Opin Chem Biol. 2016;31:82–94.
Curran S, Hagen A, Poust S, Chan LJG, Garabedian BM, Rond TD, Baluyot MJ, Vu JT, Lau AK, Yuzawa S. Probing the flexibility of an iterative modular polyketide synthase with non-native substrates in Vitro. ACS Chem Biol. 2018;13:2261–8.
Robbins T, Liu YC, Cane DE, Khosla C. Structure and mechanism of assembly line polyketide synthases. Curr Opin Struct Biol. 2016;41:10–8.
Staunton J, Weissman KJ. Polyketide biosynthesis: a millennium review. Nat Prod Rep. 2001;18:380–416.
Yu D, Xu F, Zeng J, Zhan J. Type III polyketide synthases in natural product biosynthesis. Lubmb Life. 2012;64:285–95.
Keatingeclay AT. The structures of type I polyketide synthases. Nat Prod Rep. 2012;29:1050–73.
Faille A, Gavalda S, Slama N, Lherbet C, Maveyraud L, Guillet V, Laval F, Quémard A, Mourey L, Pedelacq JD. Insights into substrate modification by dehydratases from type I polyketide synthases. J Mol Biol. 2017;429:1554–69.
Heimerl N, Hommel E, Westermann M, Meichsner D, Lohr M, Hertweck C, Grossman AR, Mittag M, Sasso S. A giant type I polyketide synthase participates in zygospore maturation in Chlamydomonas reinhardtii. Plant J Cell Mol Biol. 2018;95:268–81.
Shelest E, Heimerl N, Fichtner M, Sasso S. Multimodular type I polyketide synthases in algae evolve by module duplications and displacement of AT domains in trans. BMC Genomics. 2015;16:1015–28.
Herbst DA, Townsend CA, Maier T. The architectures of iterative type I PKS and FAS. Nat Prod Rep. 2018;35:1046–69.
Hertweck C, Luzhetskyy A, Rebets Y, Bechthold A. Type II polyketide synthases: gaining a deeper insight into enzymatic teamwork. Nat Prod Rep. 2007;24:162–90.
Shimizu Y, Ogata H, Goto S. Type III polyketide synthases: functional classification and phylogenomics. ChemBioChem. 2017;18:50–65.
Waldman AJ, Balskus EP. Lomaiviticin biosynthesis employs a new strategy for starter unit generation. Org Lett. 2014;16:640–3.
Zhang Z, Pan H, Tang G. New insights into bacterial type II polyketide biosynthesis. Research. 2017;6:172.
Pickens LB, Tang Y. Oxytetracycline biosynthesis. J Biol Chem. 2010;285:27509–15.
Matharu A-L, Cox RJ, Crosby J, Byrom KJ, Simpson TJ. MCAT is not required for in vitro polyketide synthesis in a minimal actinorhodin polyketide synthase from Streptomyces coelicolor. Chem Biol. 1998;5:699–711.
Eng CH, Backman TWH, Bailey CB, Magnan CN, Martin HG, Katz L, Baldi P, Keasling JD. ClusterCAD: a computational platform for type I modular polyketide synthase design. Nucleic Acids Res. 2018;46:D509–15.
Zargar A, Barajas JF, Lal R, Keasling JD. Polyketide synthases as a platform for chemical product design. AIChE J. 2018;64:4201–7.
Weissman KJ. Uncovering the structures of modular polyketide synthases. Nat Prod Rep. 2015;32:436–53.
Ziemert N, Alanjary M, Weber T. The evolution of genome mining in microbes – a review. Nat Prod Rep. 2016;33:988–1005.
Yuzawa S, Zargar A, Pang B, Katz L, Keasling JD. Chapter fourteen—commodity chemicals from engineered modular type I polyketide synthases. Method Enzymol. 2018;608:393–415.
Morita H, Wong CP, Abe I. How structural subtleties lead to molecular diversity for the type III polyketide synthases. J Biol Chem. 2019;294:15121–36.
Palmer CM, Alper HS. Expanding the chemical palette of industrial microbes: metabolic engineering for type III PKS-derived polyketides. Biotechnol J. 2019;14:1700463.
Bin W, Jinwei R, Liyuan L, Fang G, Guohui P, Guomin A, Bertrand A, Keqiang F, Keqian Y. Kinamycin biosynthesis employs a conserved pair of oxidases for B-ring contraction. Chem Commun. 2015;51:8845–8.
Herzon SB, Woo CM. The diazofluorene antitumor antibiotics: structural elucidation, biosynthetic, synthetic, and chemical biological studies. Nat Prod Rep. 2011;29:87–118.
Woo CM, Li Z, Paulson EK, Herzon SB. Structural basis for DNA cleavage by the potent antiproliferative agent (-)-lomaiviticin A. Proc Natl Acad Sci U S A. 2016;113:2851–6.
Otten S, Stutzmanengwall KJ, Hutchinson CR. Cloning and expression of daunorubicin biosynthesis genes from Streptomyces peucetius and S. peucetius subsp. caesius. J Bacteriol. 1990;172:3427–34.
Pokhrel AR, Chaudhary AK, Nguyen HT, Dhakal D, Le TT, Shrestha A, Liou K, Sohng JK. Overexpression of a pathway specific negative regulator enhances production of daunorubicin in bldA deficient Streptomyces peucetius ATCC 27952. Microbiol Res. 2016;192:96–102.
Wang X, Tian X, Wu Y, Shen X, Yang S, Chen S. Enhanced doxorubicin production by Streptomyces peucetius using a combination of classical strain mutation and medium optimization. Prep Biochem Biotechnol. 2018;48:514–21.
Yin S, Wang W, Wang X, Zhu Y, Jia X, Li S, Yuan F, Zhang Y, Yang K. Identification of a cluster-situated activator of oxytetracycline biosynthesis and manipulation of its expression for improved oxytetracycline production in Streptomyces rimosus. Microb Cell Fact. 2015;14:46.
Zhang W, Ames BD, Tsai S, Tang Y. Engineered biosynthesis of a novel amidated polyketide, using the malonamyl-specific initiation module from the oxytetracycline polyketide synthase. Appl Environ Microbiol. 2006;72:2573–80.
Stevens DC, Conway KR, Pearce N, Aranda LRV-P, Garza AG, Boddy CN. Alternative sigma factor over-expression enables heterologous expression of a type II polyketide biosynthetic pathway in Escherichia coli. PLoS ONE. 2013;8:e64858. https://doi.org/10.61371/journal.pone.0064858.
Yin S, Li Z, Wang X, Wang H, Jia X, Ai G, Bai Z, Shi M, Yuan F, Liu T. Heterologous expression of oxytetracycline biosynthetic gene cluster in Streptomyces venezuelae WVR2006 to improve production level and to alter fermentation process. Appl Microbiol Biot. 2016;100:10563–72.
Yin S, Wang X, Shi M, Yuan F, Wang H, Jia X, Yuan F, Sun J, Liu T, Yang K. Improvement of oxytetracycline production mediated via cooperation of resistance genes in Streptomyces rimosus. Sci China Life. 2017;60:1–8.
Lešnik U, TLukežic T, Podgoršek A, Horvat J, Tomaž P, Šala M, Jenko B, Harmrolfs K, Ocampo‐Sosa A, Martínez‐Martínez L, et al. Construction of a new class of tetracycline lead structures with potent antibacterial activity through biosynthetic engineering. Angew Chem Int Ed Engl. 2015;54:3937–40.
Zhang M, Hou XF, Qi LH, Yin Y, Li Q, Pan HX, Chen XY, Tang GL. Biosynthesis of trioxacarcin revealing a different starter unit and complex tailoring steps for type II polyketide synthase. Chem Sci. 2015;6:3440–7.
Choi YS, Zhang H, Joseph BS, Satish NK, Zhao H. In vitro reconstitution and crystal structure of p-aminobenzoate N-oxygenase (AurF) involved in aureothin biosynthesis. Proc Natl Acad Sci USA. 2008;105:6858–63.
Thomas MG, Burkart MD, Walsh CT. Conversion of l-proline to pyrrolyl-2-carboxyl–PCP during undecylprodigiosin and pyoluteorin biosynthesis. Chem Biol. 2002;9:171–84.
Hillenmeyer ME, Vandova GA, Berlew EE, Charkoudian LK. Evolution of chemical diversity by coordinated gene swaps in type II polyketide gene clusters. Proc Natl Acad Sci US A. 2015;112:13952–7.
Burson KK, Khosla C. Dissecting the chain length specificity in bacterial aromatic polyketide synthases using chimeric genes. Tetrahedron. 2000;56:9401–8.
Mcdaniel R, Ebertkhosla S, Hopwood DA, Khosla C. Engineered biosynthesis of novel polyketides: manipulation and analysis of an aromatic polyketide synthase with unproven catalytic specificities. J Am Chem Soc. 1993;115:11671–5.
Jiang L, Pu H, Xiang J, Su M, Yan X, Yang D, Zhu X, Shen B, Duan Y, Huang Y. Huanglongmycin A-C, cytotoxic polyketides biosynthesized by a putative type ii polyketide synthase from Streptomyces sp. CB09001. Front Chem. 2018;6:254.
Gan M, Liu B, Tan Y, Wang Q, Zhou H, He H, Ping Y, Yang Z, Wang Y, Xiao C. Saccharothrixones A-D, Tetracenomycin-Type Polyketides from the Marine-Derived Actinomycete Saccharothrix sp. 10-10. J Nat Prod. 2015;78:2260–5.
Silverman RH. Implications for RNase L in prostate cancer biology. Biochem. 2003;42:1805–12.
Kathia ZR, Charkoudian LK, Ridley CP, Chaitan K. Cloning, sequencing, heterologous expression, and mechanistic analysis of A-74528 biosynthesis. J Am Chem Soc. 2010;132:9122–8.
Fitzgerald JT, Charkoudian LK, Watts KR, Khosla C. Analysis and refactoring of the A-74528 biosynthetic pathway. J Am Chem Soc. 2013;135:3752–5.
Okamoto S, Taguchi T, Ochi K, Ichinose K. Biosynthesis of actinorhodin and related antibiotics: discovery of alternative routes for quinone formation encoded in the act gene cluster. Chem Biol. 2009;16:226–36.
Shen B, Summers RG, Wendtpienkowski E, Hutchinson CR. The Streptomyces glaucescens tcmKL polyketide synthase and tcmN polyketide cyclase genes govern the size and shape of aromatic polyketides. J Am Chem Soc. 1995;117:6811–21.
Tang Y, Tsai S, Khosla C. Polyketide chain length control by chain length factor. J Am Chem Soc. 2003;125:12708–9.
Murakami T, Burian J, Yanai K, Bibb MJ, Thompson CJ. A system for the targeted amplification of bacterial gene clusters multiplies antibiotic yield in Streptomyces coelicolor. Proc Natl Acad Sci USA. 2011;108:16020–5.
Fu J, Zong G, Zhang P, Zhao Z, Ma J, Pang X, Cao G. XdhR negatively regulates actinorhodin biosynthesis in Streptomyces coelicolor M145. FEMS Microbiol Lett. 2017;364:fnx226.
Hofeditz T, Unsin C, Wiese J, Imhoff JF, Wohlleben W, Grond S, Weber T. Lysoquinone-TH1, a new polyphenolic tridecaketide produced by expressing the lysolipin minimal PKS II in Streptomyces albus. J Antibiot (Tokyo). 2018;7:53.
Natasa PC, Branko B, Long PF, Daslav H, Waterman PG, Hunter IS. Ablation of the otcC gene encoding a post-polyketide hydroxylase from the oxytetracyline biosynthetic pathway in Streptomyces rimosus results in novel polyketides with altered chain length. J Biol Chem. 2005;280:37455–60.
Du D, Katsuyama Y, Shinya K, Ohnishi Y. Reconstitution of a type II polyketide synthase that catalyzes polyene formation. Angew Chem. 2018;130:1954–7.
Xu D, Ma M, Deng Z, Hong K. Genotype-driven isolation of enterocin with novel bioactivities from mangrove-derived Streptomyces qinglanensis 172205. Appl Microbiol Biotechnol. 2015;99:5825–32.
Prado L, Fernandez E, Weißbach U, Blanco G, Quiros LM, Brana AF, Mendez C, Rohr J, Salas JA. Oxidative cleavage of premithramycin B is one of the last steps in the biosynthesis of the antitumor drug mithramycin. Chem Biol. 1999;6:19–30.
Novakova R, Nunez LE, Homerova D, Knirschova R, Feckova L, Rezuchova B, Sevcikova B, Menendez N, Moris F, Cortes J. Increased heterologous production of the antitumoral polyketide mithramycin a by engineered Streptomyces lividans TK24 strains. Appl Microbiol Biotechnol. 2018;102:871.
Núñez LE, Nybo SE, González-Sabín J, Pérez M, Menéndez N, Braña AF, Shaaban KA, He M, Morís F, Salas JA, et al. A novel mithramycin analogue with high antitumor activity and less toxicity generated by combinatorial biosynthesis. J Med Chem. 2012;55:5813–25.
Osgood CL, Maloney N, Kidd CG, Kitchengoosen SM, Segars LE, Gebregiorgis M, Woldemichael GM, He M, Sankar S, Lessnick SL. Identification of mithramycin analogues with improved targeting of the EWS-FLI1 transcription factor. Clin Cancer Res. 2016;22:4105–18.
Shen B, Hutchinson CR. Tetracenomycin F1 monooxygenase: oxidation of a naphthacenone to a naphthacenequinone in the biosynthesis of tetracenomycin C in Streptomyces glaucescens. Biochemistry. 1993;32:6656–63.
Sciara G, Kendrew SG, Miele AE, Marsh N, Federici L, Malatesta F, Schimperna G, Savino C, Vallone B. The structure of ActVA-Orf6, a novel type of monooxygenase involved in actinorhodin biosynthesis. EMBO J. 2003;22:205–15.
Chung J, Fujii I, Harada S, Sankawa U, Ebizuka Y. Expression, purification, and characterization of AknX anthrone oxygenase, which is involved in aklavinone biosynthesis in Streptomyces galilaeus. J Bacteriol. 2002;184:6115–22.
Qin Z, Devine R, Hutchings MI, Wilkinson B. A role for antibiotic biosynthesis monooxygenase domain proteins in fidelity control during aromatic polyketide biosynthesis. Nat Commun. 2019;10:3611.
Piel J, Hertweck C, Shipley PR, Hunt DM, Newman MS, Moore BS. Cloning, sequencing and analysis of the enterocin biosynthesis gene cluster from the marine isolate ‘Streptomyces maritimus’: evidence for the derailment of an aromatic polyketide synthase. Chem Biol. 2000;7:943–55.
Walczak RJ, Hines JV, Strohl WR, Priestley ND. Bioconversion of the anthracycline analogue desacetyladriamycin by recombinant DoxA, a P450-monooxygenase from Streptomyces sp. strain C5. Org Lett. 2001;3:2277–9.
Sun L, Wang S, Zhang S, Shao L, Zhang Q, Skidmore C, Chang CT, Yu D, Zhan J. Characterization of three tailoring enzymes in dutomycin biosynthesis and generation of a potent antibacterial analogue. ACS Chem Biol. 2016;11:1992–2001.
Daum M, Peintner I, Linnenbrink A, Frerich A, Weber M, Paululat T, Bechthold A. Organisation of the biosynthetic gene cluster and tailoring enzymes in the biosynthesis of the tetracyclic quinone glycoside antibiotic polyketomycin. ChemBioChem. 2009;10:1073–83.
Guo X, Crnovcic I, Chang CY, Luo J, Lohman JR, Papinski M, Bechthold A, Horsman GP, Shen B. PokMT1 from the Polyketomycin Biosynthetic Machinery of Streptomyces diastatochromogenes Tü6028 Belongs to the Emerging Family of C-Methyltransferases That Act on CoA-Activated Aromatic Substrates. Biochem. 2018;57:1003–11.
Decker H, Motamedi H, Hutchinson CR. Nucleotide sequences and heterologous expression of tcmG and tcmP, biosynthetic genes for tetracenomycin C in Streptomyces glaucescens. J Bacteriol. 1993;175:3876–86.
Jansson A, Koskiniemi H, Mantsala P, Niemi J, Schneider G. Crystal structure of a ternary complex of DnrK, a methyltransferase in daunorubicin biosynthesis, with bound products. J Biol Chem. 2004;279:41149–56.
Metsaketela M. Evolution inspired engineering of antibiotic biosynthesis enzymes. Org Biomol Chem. 2017;15:4036–41.
Kim N, Kim J, Lee Y, Lee E, Kim JY, Lim Y, Chong Y, Ahn J. O-Methylation of flavonoids using DnrK based on molecular docking. J Microbiol Biotechnol. 2007;17:1991–5.
Iorio M, Cruz JCS, Simone M, Bernasconi A, Brunati C, Sosio M, Donadio S, Maffioli SI. Antibacterial paramagnetic quinones from Actinoallomurus. J Nat Prod. 2017;80:819–27.
Monciardini P, Bernasconi A, Iorio M, Brunati C, Sosio M, Campochiaro L, Landini P, Maffioli SI, Donadio S. Antibacterial aromatic polyketides incorporating the unusual amino acid enduracididine. J Nat Prod. 2019;82:35–44.
Zhang W, Watanabe K, Cai X, Jung ME, Tang Y, Zhan J. Identifying the minimal enzymes required for anhydrotetracycline biosynthesis. J Am Chem Soc. 2008;130:6068–9.
Lukežic T, Fayad AA, Bader CD, Harmrolfs K, Bartuli J, Gros S, Lesnik U, Hennessen F, Herrmann J, Pikl S. Engineering atypical tetracycline formation in Amycolatopsis sulphurea for the production of modified chelocardin antibiotics. ACS Chem Biol. 2019;14:468–77.
Ohtsubo K, Marth JD. Glycosylation in cellular mechanisms of health and disease. Cell. 2006;126:855–67.
Kunzel E, Faust B, Oelkers C, Weissbach U, Bearden DW, Weitnauer G, Westrich L, Rohr J. Inactivation of the urdGT2 Gene, which encodes a glycosyltransferase responsible for the c-glycosyltransfer of activated d-olivose, leads to formation of the novel urdamycins I, J, and K. J Am Chem Soc. 1999;121:11058–62.
Trefzer A, Hoffmeister D, Kunzel E, Stockert S, Weitnauer G, Westrich L, Rix U, Fuchser J, Bindseil KU, Rohr J. Function of glycosyltransferase genes involved in urdamycin A biosynthesis. Chem Biol. 2000;7:133–42.
Gui C, Liu Y, Zhou Z, Zhang S, Hu Y, Gu Y, Huang H, Ju J. Angucycline glycosides from mangrove-derived Streptomyces diastaticus subsp SCSIO GJ056. Mar Drugs. 2018;16:185.
Basnet DB, Oh T, Vu TT, Sthapit B, Liou K, Lee HC, Yoo JC, Sohng JK. Angucyclines Sch 47554 and Sch 47555 from Streptomyces sp. SCC-2136: cloning, sequencing, and characterization. Mol Cells. 2006;22:154.
Otten S, Liu X, Ferguson J, Hutchinson CR. Cloning and characterization of the Streptomyces peucetius dnrQS genes encoding a daunosamine biosynthesis enzyme and a glycosyl transferase involved in daunorubicin biosynthesis. J Bacteriol. 1995;177:6688–92.
Chu LL, Pandey RP, Shin JY, Jung HJ, Sohng JK. Synthetic analog of anticancer drug daunorubicin from daunorubicinone using one-pot enzymatic UDP-recycling glycosylation. J Mol Catal B Enzym. 2016;124:1–10.
Remsing LL, Garcia-Bernardo J, Gonzalez A, Künzel E, Rix U, Braña AF, Bearden DW, Méndez C, Salas JA, Rohr J. Ketopremithramycins and ketomithramycins, four new aureolic acid-type compounds obtained upon inactivation of two genes involved in the biosynthesis of the deoxysugar moieties of the antitumor drug mithramycin by streptomyces argillaceus, reveal novel insights into post-PKS tailoring steps of the mithramycin biosynthetic pathway. J Am Chem Soc. 2002;124:1606–14.
Mo J, Ye J, Chen H, Hou B, Wu H, Zhang H. Cloning and identification of the Frigocyclinone biosynthetic gene cluster from Streptomyces griseus strain NTK 97. Biosci Biotechnol Biochem. 2019;83:2082–9.
Acknowledgements
This work was funded by National Key Research and Development Program of China (2018YFA0903000, 2018YFA0901400 and 2018YFA0901800), the National Natural Science Foundation of China (Grants 21908003, 21636001), the China Postdoctoral Science Foundation (2019M650454), the Fundamental Research Funds for Central Universities (buctrc201911, XK1802-8, buctylkxj03).
Author information
Authors and Affiliations
Contributions
JW and XS overall coordinated and wrote the manuscript; JW, RZ, XC X. Sun and XS interpreted of illustrations and other information; RZ, YY, XS and QY revised the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, J., Zhang, R., Chen, X. et al. Biosynthesis of aromatic polyketides in microorganisms using type II polyketide synthases. Microb Cell Fact 19, 110 (2020). https://doi.org/10.1186/s12934-020-01367-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-020-01367-4