
Abstract
Background
Structures and functions of ecosystems and, subsequently, their services for human societies may be influenced by climate change and atmospheric deposition. Jenssen et al. (UBA Texte 87/2013: 1–381, 2013) developed a spatially explicit evaluation system enabling the evaluation of ecosystems’ integrity. This methodology is based on a spatially explicit ecosystem classification of forests. Based on the six ecological functions, the methodology enables to compare the ecosystem type-specific integrity at different levels of ecological hierarchy for a reference state (1961–1990) with the further development of the forest ecosystem types as measured for the years 1991–2010 and as modelled for the period 2011–2070. The present study aimed at deepening the methodology and developing it into a practical system for assessing and mapping forest ecosystem integrity and services. The objectives of this advanced investigation were: (1) to quantify the reference conditions for a total of 61 forest ecosystem types; (2) to test the possibility of supplementing the quantification of ecosystem integrity by information on soil biocenoses as yielded by soil monitoring; (3) to model chemical soil indicators and to compare the respective results with those derived by Ellenberg’s indicator values for nutrient state; and (4) to verify the indicator modelling.
Results
Reference states related to the time prior to 1991 have been quantified for a total of 61 forest ecosystem types covering 85% (81,577 km2) of the mapped forest area of Germany. The reference states comprise statistical indicators for the plant-species diversity (habitat function), for nutrient and water balances and further ecological information as net-primary production and carbon storage. The assignment of lumbricide communities as soil biocenosis indicators was attempted but not succeeded because of insufficient data availability. The nutrient cycle types of the elaborated reference states were characterized by humus form, C/N ratio in topsoil and N indicator values according to Ellenberg et al. (Scr Geobot 18:1–262, 2001). Applying the developed methodology, for 83 out of 105 study plots the reference states prior to 1991 could be determined.
Conclusions
For complementing forest ecosystem reference states by soil biocenosis indicators it is necessary to further evaluate the primary literature looking for missing observation data. The W.I.E. indicator value applied in this paper to determine topsoil C/N ratios in forests is well suited for area-covering mapping of both near-natural forest–soil states and deposition-induced disharmonic state changes, in which C/N value and base saturation are no longer correlated.
Similar content being viewed by others
Background and objectives
Background: Previous method development as framework of the study at hand.
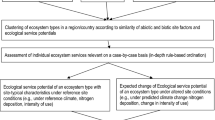
Climate change and atmospheric nitrogen inputs can alter the integrity of ecosystems in terms of their dominant structures and functions, and thus change or even limit their benefits (services) for humans. Jenssen et al. [23] developed the methodology for an ecological, spatially explicit and nationwide applicable assessment of impacts on forest ecosystem integrity [26, 29, 30, 38, 39]. In further extensive, more application-oriented studies, the scientific foundations of the methodology were deepened and developed into a practical integrated assessment system for ecosystem functions. One of these studies is presented in this article.
The methodology developed so far enables an integrative assessment of changes in forest ecosystem integrity taking into account the effects of climate change in combination with atmospheric nitrogen (N) deposition. Thereby, self-organisation, functionality and compliance of abiotic and biotic properties with the natural site potential (identity) were considered as characteristics of ecosystem integrity. The approach comprises the following steps: classification of forest ecosystem types, description of ecosystem type specifics, historical reference states by using selected indicators, dynamic modelling of ecosystem climate and soil conditions and comparisons of current and future states with the reference states to evaluate risk of loss of ecosystem integrity.
The methodology was based on an extensive vegetation database from the Waldkunde Institut Eberswalde (W.I.E., Germany) as well as on nationally available data from maps and long-term monitoring programmes. It is complemented by dynamic modelling of future climate and soil conditions. The methodology enables the identification and mapping of potential natural ecosystem types and current forest ecosystem types. For certain scenarios of the development of climate and atmospheric nitrogen inputs (2011–2070), expectable future ecosystem developments can be estimated and evaluated. The condition of forest ecosystem types is assessed regarding their ecological functionality, their (near) natural biological and chemical characteristics, and their stress tolerance to anthropogenic nitrogen inputs and climate change. The approach directly links changes of abiotic environmental factors with changes of ecosystem structures and functions by evaluation of monitoring data or modelling. This enables identification of the causes of disturbances as early as possible and to derive suitable measures for maintaining and developing a desired ecosystem condition.
A fundamental component of the methodology was the newly developed classification of Germany’s current forest ecosystems. It aimed at distinguishing and mapping of area units which are homogenous in terms of structures and ecological functions. To this end the W.I.E. database is used, which combines abiotic and biotic criteria in a specific indicator model. The spatial and thematic concordance of the new classification with other ecosystem classifications for which no spatial concretisation has been carried out nationwide so far has been achieved. This is especially true for the habitat classification of the European Nature Information System—EUNIS [5], the ecosystem classification used in the German red list of endangered ecosystem types [36] and, respectively, the classification of habitat types according to Annex I to Habitats Directive 92/43EEC. Thus, the developed ecosystem classification is connectable to other assessments, and in contrast to the previous ecosystem classifications in Germany it enables a spatially differentiated, ecologically based interpretation.
Because ecosystem types are really basic entities in the approach we provide a summary on how it was identified already here: the ecosystem type is clearly determined by three ecological coordinates of the forest vegetation, which are lumped to an ecosystem type code. The first coordinate in the coding describes the large-climatic area scaling (altitudinal zonation, horizontal zonation), the second coordinate the water balance, i.e. the humidity level (of the topsoil and the air layer near the ground; very dry to continuously very wet). The third coordinate indicates the type of nutrient cycle, which in turn corresponds to defined intervals of soil chemical indicators (pH value in n/10 KCl, base saturation, C/N ratio) of the soil root space predominantly used by the plants for nutrition and thus also to certain forms of humus (more detail in [26]). Predominantly used soil root space is defined by the upper 5 cm starting with the organic humus layer. Depending on ecosystem type, this space contains either or both organic layer and mineral soil. For examples see the Additional file 1: Tables S1 and S2.
First, for 33 ecosystem types, a historical reference condition was quantified based on the data from the period 1961 to 1990 [23, 26, 38]. The reference condition was defined as a type-specific condition of ecosystems, the characteristics of which were quantified by intervals of variables for historical conditions (1961–1990). This reference period is a historical one and corresponds to the climate normal used for analysing and modelling climate data [42]. Since in Germany systematic monitoring of ecosystem conditions started in the 1960s at the earliest, it was assumed that out of available measurement data conditions from this period show the smallest changes due to atmospheric N deposition and climate change as compared to pre-industrial conditions.
For selected ecosystem functions (habitat function, net primary function, carbon storage, nutrient flow, water flow and adaptability), indicators were selected to compare current and modelled future ecosystem conditions with the reference condition (cf. Table 1 in [38]). The indication was based quantitatively on the data from monitoring programmes, in particular national data of the intensive environment monitoring of forests [17] and from the Waldkunde Institut Eberswalde (W.I.E., Germany) database.
The simulation model Very Simple Dynamic (VSD) Soil Acidification Model [2, 35] was used to derive quantified information on possible future ecosystem development during the years 2011–2040 and 2041–2070. To this end, the STARFootnote 1 II projections for the climate scenarios RCPFootnote 2 2.6 and RCP 8.5 and two scenarios for atmospheric nitrogen inputs (from 2010: 5 kg/ha * a and 15 kg/ha * a N deposition) have been used. The dynamic modelling enables the calculation of future soil conditions in terms of acidification and eutrophication status as well as water supply for each location in Germany described by a soil profile, taking into consideration the scenarios as described. By comparing the modelled development of the indicators organic carbon, C/N ratio, pH and base saturation with respective information on soil characteristics from the W.I.E. indicator model and with the ecosystem classification, possible future ecosystem developments can be estimated site specifically. This could even include a possible future change of the current ecosystem type into another one, which is related with a change or limitation of biodiversity or habitat function, respectively.
Finally, a rough concept for estimating changes in ecosystem integrity of previously defined and mapped forest ecosystems was developed. The consideration of nutrient, water and energy balance should be combined with other professional assessments—e.g. of nature conservation criteria—in order to achieve an even more comprehensive statement on ecosystem integrity from the point of view of potential users.
Objectives
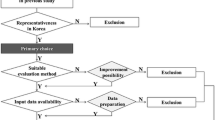
Against this background, the advanced method development presented in the article at hand pursued the following four interlinked goals of methodological development (“Complementing the reference states of forest ecosystem types”, “Supplementing the information on reference conditions by soil biocenosis data”, “Validation and extension of the W.I.E. indicator model”, and “Verification of indicator models” sections): one objective was to supplement the reference conditions with further important forest ecosystem types and with primary data from the period 1960–1990 not yet provided in Jenssen et al. [23], Schröder et al. [38]. The reference conditions quantified so far included 33 ecosystem types. 30 of these are forest ecosystem types of the high mountain and mountain forest locations, the subatlantic, the Central European as well as the subcontinental lowlands, 3 ecosystem types describe near-natural openland ecosystems. For a nationwide application of the concept, this selection should be supplemented by about 30–35 additional representative ecosystem types. The criteria for the selection of these, the databases and methods as well as results are described in “Complementing the reference states of forest ecosystem types” section. Broadening the spatial reference base and the spectrum of ecosystem types should be added by an expansion of the ecological indicators.
Consequently, the second objective of the study presented in the article at hand aimed at supplementing the quantification of ecosystem integrity by information on soil biocenoses as yielded by soil monitoring (“Supplementing the information on reference conditions by soil biocenosis data” section). To this end, it should be investigated whether it is possible to assign lumbricide communities to ecosystem types based on correlations between the biotic data and abiotic topsoil indicators C/N ratio, base saturation and pH and soil moisture. If the outcome was positive, the lumbricide communities could be regarded as characteristic for specific combinations of nutrient cycle type and water balance type, which in turn correspond to certain humus forms under different soil moisture conditions. Then, soil biological reference states described by characteristic species combinations of lumbricides could be assigned to the different ecosystem types.
This kind of additional soil biocoenotic indication of ecosystem types should be corroborated by comparing the results of the statistically based W.I.E. indicator model with the indicator scores as suggested by Ellenberg et al. [7, 8] and, on this basis, to derive recommendations for the application of each model. For the development of the W.I.E. indicator model more than 1600 measured values of the topsoil condition in combination with the corresponding plant community relevés were used, which are documented in the database of the W.I.E. (“Validation and extension of the W.I.E. indicator model” section). Finally, the W.I.E. indicator model should be validated (“Verification of indicator models” section).
Since each of the four objectives of this study is rather comprehensive, each of the respective “Complementing the reference states of forest ecosystem types”, “Supplementing the information on reference conditions by soil biocenosis data”, “Validation and extension of the W.I.E. indicator model”, and “Verification of indicator models” sections were structured according to the IMRaD scheme.
Complementing the reference states of forest ecosystem types
Objective
In the selection of additional ecosystem types, for which reference states should be derived, high priority was set on gaining a high spatial coverage, so that for about half of all 131 ecosystem types mapped in the preceding investigations [23, 38] and the majority of Germany’s territory covered with forest ecosystems descriptions of reference states would become available. Another priority was to consider ecologically relevant ecosystem types in terms of specific ecological functions. Finally, also ecosystem types should be considered, which could gain high relevance in future in Germany because of climate change.
Materials and methods
The ecosystem types studied so far [23, 38] provided the reference conditions for 30 current forest ecosystem types in Germany, which are located on a total of 35% of the mapped area. Of these 35%, 23% relate to forests in the sub-Atlantic and Central European lowlands and 12% to forests in high mountains and mountainous areas. The selection of further ecosystem types in the present study was based on the information from the regions dealt with in the previous study [23, 38], namely the Thuringian Forest, the region South-Brandenburg/North-Saxony and the region North-Brandenburg. It was done using the following criteria:
-
1.
Which spatially representative forest ecosystems are currently lacking?
-
2.
Which ecologically significant forest ecosystems are missing?
-
3.
Which forest ecosystems will be lacking in the future with regard to climate change and assuming continuing enhanced nitrogen deposition?
-
4.
Which selection leads to the greatest possible coverage of the mapped forest ecosystem types?
The analysis with regard to spatial representativity (criterion 1) showed that the lowland spruce forests on 18% of the mapped area are completely missing, lowland beech forests on 16% of the mapped area and lowland pine forests on 9% of the mapped area are neither considered. It was therefore decided to supplement these forests that were lacking in terms of areal proportion.
With regard to the ecological relevance (criterion 2), in particular the wet forest ecosystems (alder forests, ash forests, oak forests) have to be supplemented. Although these account for only 5% of the mapped area, they are relevant for carbon storage therefore of outstanding importance for climate protection [16]. At the same time, these ecosystems are highly dependent on the water balance of the landscape and are therefore particularly threatened by continuing climate change. Wet and humid forest ecosystems are a preferred habitat for earthworm cenoses [21]. Their consideration thus is also in line with the objective to examine a possible inclusion of earthworm cenoses in the assessment of ecosystem integrity. Furthermore, selected ecosystem types were taken into account due to their special relevance for nature conservation, e.g. the ecosystem types to be assigned to the habitat types 9150, 9190 and 91D0 according to the EU Habitats Directive 93/42 EWG, which have not yet been considered.
Accepting the fact that some ecosystems already show irreversible anthropogenic effect and/or are facing future changes of ecological conditions because of climate change and atmospheric N deposition, the methodology should also enable the derivation of possible target ecosystem types. Such adaptations of ecosystem types can develop spontaneously or supported by management measures. For this reason, significant forest ecosystems must be taken into account in the quantification of reference conditions, which currently have a rather subordinate spatial relevance (criterion 3). These include in particular the climate–plastic forest types of the pine sessile oak forests, sessile oak forests, small-leaved lime hornbeam forests and hornbeam beech forests [24], the current areal proportion of which is less than 2%.
The current spatial representation of forest ecosystem types within the selected model regions called at the beginning of this chapter and the ICPFootnote 3 Forests Level II plots available therein has in turn influenced the selection of additional ecosystem types for which reference conditions should be quantified. The aim was to achieve the highest possible area coverage of the mapped ecosystem types in these regions (criterion 4).
Results
As a result of these four selection criteria, the extension of the description of reference conditions was developed for the forest ecosystem types shown in Additional file 1: Table S1. With the 31 new ecosystem types, 50% of the mapped forest area of Germany (47,820 km2) are covered, so that with the total of 61 ecosystem types the quantitative description of the historical reference conditions is provided for more than 85% (81,577 km2) of the mapped total forest area of Germany.
In addition, the reference conditions for all 61 forest ecosystem types were complemented and partly revised. The results are summarized in Additional file 1: Table S1 and detailed in the research data documentation [25]. The following features have been added to the reference states throughout:
-
1.
Supplementing the characteristics of the "habitat function" with the occurrence potential for plant species of the Red List Germany [28] and their frequency and quantity;
-
2.
Addition of nutritional parameters of leafes and needles for sessile oak (Quercus petraea), pedunculate oak (Quercus robur), white fir (Abies alba), small-leaved lime (Tilia cordata), hornbeam (Carpinus betulus), sycamore maple (Acer pseudoplatanus) and ash (Fraxinus excelsior) to the "nutrient balance" characteristics;
-
3.
Inclusion of soil data available in the literature and in the W.I.E. archives, which can be clearly assigned to ecosystem types;
-
4.
Integrating illustrations of typical soil profiles for almost all ecosystem types;
-
5.
Attaching of all images as high-resolution jpg files;
-
6.
References for all information submitted;
-
7.
Excel tables with a total of 3683 vegetation surveys (2707 of them in the reference period 1905–1990), with source references and—if available—topsoil data.
The indicators for the habitat function (maximum Kullback distance [20, 27] of the individual surveys to the mean plant-species abundance distribution, and, additionally, minimum percentage similarity [20] of the individual surveys to the mean plant-species abundance distribution) to describe the reference conditions before 1990 were recalculated for all 61 forest ecosystem types according to the methodology identified in Jenssen et al. [23] (Additional file 1: Table S2). On average across all ecosystem types, an average maximum Kullback distance of 0.83 ± 0.29 of the individual surveys to the mean species abundance distribution and an average minimum percentage similarity of the individual surveys to the mean species abundance distribution of (57 ± 11)% result. This proves a high degree of homogeneity within the 61 forest ecosystem types identified.
Mean values and standard deviations for the indicators of nutrient and water balance (C/N ratio, pH value in 1/10 KCl, base saturation and moisture index) were calculated to describe the reference conditions before 1990 for a total of 61 forest ecosystem types using the indicator models described in Jenssen et al. [23]. The results are presented in Additional file 1: Table S1. For example: the analysis for the ‘Moder beech forests on Bunter’ (code Eb-5n-C2) is based on 120 vegetation samples between 1960 and 1990. The C/N ratio averages out to 17.9 and the standard deviation amounts to 2.1 This leads to a typical range of C/N 15.8–20, which is used to characterize the ecosystem type-specific reference state. The pH value is between 3.71 and 4.91, the base saturation (V) between 24.5 and 32.1 and the soil moisture index (DKF) between 4.4 and 5.4. The indicator models are documented in detail and compared with the approach suggested by Ellenberg et al. [7, 8] and VDI [41].
The reference states refer to the period up to 1990, mainly from 1960 onwards, but in individual cases to data dating back to the beginning of the twentieth century. For each ecosystem type its reference status is indicated by a data sheet with the following information:
-
1.
Ecosystem code: 1st digit = Eco-climatological coordinate, 2nd digit = Water balance type, 3rd digit = Nutrient cycle type, description see identification key ([37], vol. 3),
-
1.
Name of ecosystem type;
-
2.
EUNIS Class;
-
3.
Biotope type BfN [36],
-
4.
Vegetation type according to common plant sociological classifications;
-
5.
Photo;
-
6.
Habitat type according to Council Directive 92/43/EEC of 21 May 1992 on the conservation of natural habitats and of wild fauna and flora;
-
7.
Position in the two-dimensional ecogram with the coordinates soil moisture and base saturation;
-
8.
Site characteristics: Soil form, soil type, terrain, macroclimate;
-
9.
Habitat function: Characteristic species association with continuity and mean quantity development of soil coverage, maximum Kullback distance of the individual surveys to mean species quantity distribution, minimum similarity of the individual surveys with the mean species quantity distribution;
-
10.
Net primary production: Above-ground average annual net primary production (NPP) at the time of culmination of increment in tree wood, leaf/needle mass, ground vegetation and total mass, population top height at age 100 as comparative parameter;
-
11.
Carbon storage: Carbon stock in humus (Corg in humus layer and in soil up to 80 cm depth);
-
12.
Nutrient flow: pH value in 1/10 KCl, base saturation V in % and C/N ratio in the uppermost 5 cm from H to Ah horizon (interval of mean value and standard deviation), humus form, nutritional parameters N%, P%, K%, Ca%, Mg% in the assimilation apparatus of trees in g/100 g of leaf/needle dry matter (time of August, interval of mean value and standard deviation);
-
13.
Water flow: Humidity index scaling from 1 to 9 according to Hofmann [14] (interval of mean value and standard deviation),
-
14.
Adaptation to changing environmental conditions: maximum proportions of tree species belonging to the site in self-organised development stages.
The information in these data sheets is documented by the digitally provided original data [25], consisting of,
-
1.
Total table of vegetation composition (Excel);
-
2.
Soil data/soil profile;
-
3.
High-resolution photo of the respective ecosystem type;
-
4.
Growth data;
-
5.
Literature.
Supplementing the information on reference conditions by soil biocenosis data
Background and objective
Graefe [10, 11], Graefe and Belotti [12] and Graefe and Beylich [13] suggest that it is possible to assign lumbricide communities to ecosystem types, since strong correlations between the topsoil indicators C/N ratio, base saturation and pH and soil moisture have been demonstrated. On the basis of these findings, the lumbricide communities can be assumed to be characteristic for specific combinations of nutrient cycle type and water balance type, which in turn correspond to certain humus forms under different soil moisture conditions. The aim of the investigations here is checking the feasibility, to assign soil biological reference states to the different ecosystem types, which consist of characteristic species combinations of lumbricides in terms of quality and quantity. To this end the extensive Edaphobase, a taxonomic–ecological database system of the Senckenberg Institute in Görlitz (Germany) that combines existing taxonomic primary data on soil organisms from scientific literature and reports etc. [40], was evaluated.
Materials and methods
The Edaphobase was initially evaluated on the basis of biotope types according to Riecken et al. [36]. The assignment of biotope types to ecosystem types as defined in this paper is not clear, however, since several ecosystem types can usually be assigned to one biotope type. This is due to more differentiated homogeneity criteria for ecosystem types (vegetation composition, site conditions and in particular process homogeneity) compared to biotope types (land use, vegetation composition, site conditions), which leads to a more differentiated classification of ecosystems. Therefore, in a second step, the data on physical and chemical soil characteristics as well as vegetation structure available in the Edaphobase should be used to specify the assignment of soil biological recordings within a biotope type. In particular, the data on soil and humus type and chemical topsoil properties, if available, may be considered. However, the data on vegetation going beyond the biotope type are only included in the Edaphobase in exceptional cases. The data assigned to an ecosystem type or—if the unambiguous assignment was not possible—to a biotope type were each stored in an Excel table. The further evaluation then took place outside Edaphobase on the basis of these Excel tables.
After the allocation of the recordings to the ecosystem types, the next step was to summarise these recordings in tables, in which characteristic species combinations are shown in typical quantity. In accordance with the methodology used in merging of vegetation surveys [23], information on continuity (relative frequency of occurrence of a species) and abundance (quantity development, e.g. as mean quantity or dominance class) should be combined wherever possible. In any case, the prerequisite for this is that the data can be evaluated in such a way that different observations can be combined into a single image. These observations necessarily have been made at the same location during the same period, they are characterised by the same vegetation structure and topsoil indicators, and have a similar area reference.
Results and discussion
On the basis of the available data, the classifications of the observations to ecosystem types or groups of ecosystem types shown in Additional file 1: Table S3 were made.
The lack of completeness and very often also a lack of plausibility of the abiotic topsoil data usually only allow an assignment to biotope types of the BfN, which, however, comprise several ecosystem types and in particular also nutrient cycle types. The Edaphobase does not contain any vegetation surveys and also only very few references to vegetation, which would make an ecosystem type classification possible. Another fundamental problem arises from the fact that it is often not possible to assign individual observations to a homogeneous area of a relevé, i.e. to an image. Area references to the sampling are also not possible. The species inventories derived on this basis are little to hardly differentiated, a reliable indication of continuity and/or mean abundance values is not possible.
The analysis shows that only for some ecosystem types is there a sufficient number of observations that could in principle allow characterisation with regard to typical lumbricide communities. This applies in particular to the black alder forest ecosystems and the oak–hornbeam ecosystems. For this purpose, however, it is necessary to evaluate the primary literature sources indicated in Edaphobase for each observation. However, this could not be achieved within the framework of the present feasibility study.
Validation and extension of the W.I.E. indicator model
Background and objective
Under given climatic and stocking conditions, the amount of nitrogen (N) available to plants is a major driver for the differentiation of forest vegetation stands far from groundwater [20]. This is also the reason why the external N supply via atmospheric deposition has led to rapid, large-scale vegetation changes [1, 6, 9, 14, 15, 19, 22].
The N sensitivity of the vegetation, the high aerodynamic roughness of the canopy roofs and the fact that the N budget in forests is mostly not controlled by fertilization or other management measures, make forest plant species an ideal indicator also for anthropogenic N inputs. While N inputs, N availability and N discharges into groundwater can only be measured selectively and with great effort, spatial gradients and temporal changes in N eutrophication can also be detected with bioindication using forest plant species, e.g. within the framework of environmental monitoring programmes. For this purpose, VDI Guideline 3959 Part 1 specifies a procedure [41] based on the use of N indicator values according to Ellenberg et al. [7, 8]. For Central European ferns and flowering plants, these indicator values are defined as ordinal numbers on a scale between 1 (very low) and 9 (very high). For a vegetation survey with at least five plant species, to each of which a N indicator value was assigned, the arithmetic mean (NZm) is calculated and this mean value is assigned to the corresponding study area. Although the averaging of ordinal numbers is not actually permissible from a mathematical point of view, the procedure has proved to be very robust and effective in its application [3]. On this basis, the study areas to be evaluated in this investigation were assigned to the N-availability levels shown in rows 2 and 3 of Table 1.
The indicator values of the plant species represent estimations derived from expert assumptions on the basis of N contents in plant leaves [7, 8]. They thus have only an indirect relation to the available N reserves in the forest soils. They cannot be equated with chemical topsoil indicators and are therefore not directly comparable with measurement or model values. In the calculation of mean values, each occurring plant species is evaluated in the same way, independent of its abundance on the survey plot. In fact, any form of weighting, e.g. with area cover or the number of individuals, would also be arbitrary [3]. With the help of the Ellenberg indicator value model, a large-scale database study confirmed eutrophication of the Central European forest landscape since the middle of the last century [9].
For the assessment of the ecosystem integrity of near natural forest and managed forest ecosystems in this study, a model (W.I.E. indicator value model) is used that maps the indicator value of the forest floor vegetation for the C/N ratio in the topsoil [20]. On the one hand, it is used to define the topsoil conditions of the reference conditions (ecosystem types). By mapping the current ecosystem types of forest vegetation, reference states of the C/N ratios in certain type-specific ranges can thus be given with surface coverage. Periodic recording of the vegetation composition at a location, e.g. within the framework of nationwide inventory or monitoring procedures, makes it possible to detect developments of the C/N ratios in the topsoil and thus also of the N-availabilities. Comparison with the ranges of the reference state of the site can be executed with high temporal resolution, depicting whether the site is still within this range or deviates.
The core of the W.I.E. indicator value model is the calculation of probability density functions for the distribution of plant species over the indicator topsoil C/N and pH value. The uppermost 5 cm of the soil profile are taken into account, regardless of whether they are sampled from the organic layer or the mineral soil. This model makes it possible for the first time to quantify both the amplitudes and the focal points of the occurrence of plant species in relation to the chemical conditions of the topsoil. This means, for example, that plant species with a narrow ecological amplitude have a different indicator value with regard to the topsoil C/N ratio than those occurring in a wide C/N range. Both symmetrical and skewed distributions are possible. In addition, both the stratum affiliation (two strata) and the quantity development (up to three cover value classes) of the plant species are taken into account in their indicator value. The distribution functions of the plant species occurring at a location are linked multiplicatively for each class of C/N values and the resulting distribution function is normalized so that the area under the curve is one. In this way, a new distribution function for the study area is obtained, which indicates the probability that this area has a certain C/N ratio in the topsoil. The area can then be assigned the modal value of the resulting distribution or, for practical reasons, the arithmetic mean and standard deviation, the latter because the resulting distribution is usually very symmetrical and approximates a normal distribution.
The aim of this investigation is to compare the applicability of both methods (W.I.E. indicator value model, VDI method according to Ellenberg et al. [7, 8] and to relate them. This is necessary because the VDI procedure is widely used and has a guideline character, and with the indication of the Ellenberg indicator values in most vegetation databases it is also easy to apply. Despite the different prerequisites of both models described above, it is particularly desirable for practical purposes to assign approximate intervals of the topsoil C/N to the N-availability levels according to VDI. In addition, it enables to assign them to the nutrient cycle types used for the description of the reference states in this investigation and mapped throughout Germany.
Materials and methods
To compare both models, those vegetation surveys were selected from the W.I.E. database for which measured topsoil C/N values were available and which contained at least five plant species with Ellenberg–N indicator values. Furthermore, the analysis was limited to forest surveys, i.e. the cover of the tree layers should be at least 30%. The C/N values refer either to the humus layer of the organic layer (Oh) or to the mineral Ah horizon. If both values were available, the value for the Oh layer was calculated. In this way, a data set of 1328 vegetation analyses with associated C/N topsoil values was selected. It should be noted that this data set is largely a subset of the data set with a total of 1643 C/N values that has already been used to derive the probability density functions used in the W.I.E. indicator value model.
First, the C/N measured values were qualitatively checked for normal distribution using a histogram, a Q–Q plot and additionally quantitatively using the Shapiro–Wilk test. In the second step, the mean N indicator values NZm were calculated according to VDI and also tested for normality. Since there was no indication of normal distribution in either case, a χ2 adjustment test was used to check whether both populations can be attributed to at least the same non-normal distribution function. Since the data sets are not bi-normally distributed, the correlation between the C/N readings and the N indicator values was calculated using Spearman's rank correlation coefficient. Since nonlinear regression models did not yield significantly better results, the C/N measured values were modelled using linear regression from the N nitrogen numbers NZm and the linear coefficient of determination r2 was calculated. Subsequently, both the W.I.E. indicator value model and the linear regression model were used to model C/N topsoil values from a total of 17,703 vegetation analyses of the W.I.E. database with at least five N indicator species each.
In a final step, a total of 2499 vegetation surveys (survey year before 1991), which were used and documented in this study for the derivation and documentation of 61 reference states (ecosystem types) and which each contain at least five N indicator species, were used to relate the nutrient cycle types of the reference states on the one hand and the C/N topsoil ratios and the N availability levels derived from the Ellenberg indicator values on the other hand. The C/N model values and mean N indicator values NZm assigned to the different nutrient cycle types of the ecosystem reference states were checked with the parameter-free Wilcoxon–Mann–Whitney U test to determine whether the central trend measures between the nutrient cycle types differ from one another.
Results and discussion
The frequency distribution shows a markedly right-skewed distribution of the C/N values and a left-skewed distribution of the indicator values NZm (Fig. 1). Both distributions deviate clearly from a normal distribution. The hypothesis derived from this finding that both distributions (C/N ratios in reverse order of the classes) approximate a common (non-normal) distribution function could be rejected at the significance level p = 0.05.

The ranking correlation coefficient between the 1328 selected C/N measurements and the values obtained from the vegetation surveys using the W.I.E. indicator model amounts to r = 0.92. The correlation between the measured values and the calculated mean N indicator values NZm is r = − 0.72. If the C/N measured values are replaced by the W.I.E. model values C/N, a ranking correlation to the N indicator values NZm of r = − 0.83 is obtained.
A calculation of topsoil C/N values from N indicator values determined according to the VDI guideline can be carried out with the linear relation (Fig. 2).
whereby the coefficient of determination amounts to r2 = 0.53.

C/N ratios measured in the topsoil and plotted against the mean N indicator values NZm [8] calculated from the species composition
As the N-indicator value and C/N values have a different distribution structure according to the investigations described, and in particular are not bi-normally distributed, the quantity of all vegetation analyses covering the entire ecological breadth of Central European forests and woodlands was stratified according to the nutrient cycle types of the reference states [23] (Fig. 3; Additional file 1: Table S4). All those ecosystems are grouped together in the nutrient cycle types which—irrespective of their climatic conditionality and their water balance—are largely homogeneous and different from each other with regard to the determining characteristics of the nutrient balance. As expected, the C/N ratios of the topsoil show a clear differentiation between the nutrient cycle types, which is predominantly statistically significant. The N-indicator values also result in a ranking of the mean values, which, however, are often not significant due to the higher variabilities (overlaps of the standard ranges). With regard to C/N, the types B2 (moder-raw humus) and C1 (raw humus-moder) are not distinguishable. They do show, however, different indicator values. The morphological humus forms raw humus, moder-raw humus (B1, B2) and raw humus-moder, moder (C1, C2) can be combined with regard to the indicator values, but they are more heterogeneous with regard to the C/N distribution. Also not clearly distinguished (both with regard to C/N and indicator values) are the nutrient cycle types E1 (mull) and E2 (calcareous mull) as well as T5 (calcareous peat) and Tangel (TA1) (only with regard to C/N).

The results very clearly demonstrate the qualitative differences between the two compared approaches described in “Background and objective” section. This refers both to the nature of the quantities modelled and to the different methods of modelling. With the proposed indicator value model, one third more variance of the topsoil C/N ratios can be explained than with N indicator values according to Ellenberg. This is plausible insofar as our indicator value model was parameterized with measured values of C/N in soil, while the indicator values represent ordinal estimates derived from N contents in leaves. Provided that the absolute amounts of carbon in the topsoil are known, the direct relation to quantitative topsoil indicators of the proposed model allows quantitative statements on the fixation of N in forest soils as well as the parameterisation of and comparison with dynamic material balance models and measured values. The better ranking correlation of the indicator values with the C/N model values as compared to the C/N measured values is also interesting. This can be interpreted as an indication of a high random variability of the C/N ratio (laboratory error, representativeness of the sample name for the entire study area), which is partially compensated by both models.
It also becomes clear that the N-indicator values by their nature can be interpreted more strongly as nutrient indicator values, which is plausible insofar as the N uptake into the leaves (as well as the morphological humus form) depends more on the total nutrient supply than the C/N ratio. This allows the different selectivity of both models to be explained with regard to nutrient cycle types/morphological humus forms. From this it can be in turn concluded that our indicator value model can map in particular atmospheric deposition-induced disharmonic state changes in which C/N value and base saturation are no longer correlated as in the natural humus forms.
At the same time, however, the results also show that in principle both model approaches are comparable and can be parallelized (Table 1). Thus, reference states assigned with the type code A (raw humus, meager) correspond in rough approximation to dystrophic states according to VDI guideline, those with the type code B or b (raw humus, moder-raw humus) to oligotrophic states, C or c types (raw humus-moder, moder and calcareous moder) to eutrophic, D and E1 types to eutrophic (brown-mull, calcareous brown-mull and mull) and E2 types to naturally very eutrophic states (calcareous mull). On the basis of these reference states, deviations of the current state, for instance due to anthropogenic deposition, are interpreted and compared.
Verification of indicator models
Background and objective
The modules developed in “Validation and extension of the W.I.E. indicator model” section should finally be applied to quantify the physical and chemical topsoil indicators with the aid of vegetation indicator values and to determine the ecosystem type. Further, they are used to assign the reference states required for the assessment of ecosystem integrity with the aid of vegetation–structural distance measures. For this purpose, vegetation surveys are to be used which have not yet been considered in the derivation of reference conditions and model modules.
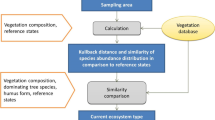
The ecosystem type is clearly determined by the three ecological coordinates as described in chapter 1. On the one hand, the ecosystem type can be determined by applying vegetation–structural distance or similarity measures, which allow a quantitative assignment to the reference conditions of these types via a single measure. On the other hand, the classification of the ecosystem type can be done by separate determination of its ecological coordinates. The indicator value models for vegetation can be used to determine the type of water balance and nutrient cycle. The large-scale climatic coordinate can be determined by locating the study area on the nationwide map of the current semi-natural vegetation.
The determination of the ecosystem type by applying vegetation–structural distance or similarity measures implies a reduction of vegetation structures to a single measure and is inevitably associated with a loss of information. First, the most probable ecosystem type was determined from the agreement of different distance and similarity measures as well as the indicator values of the vegetation. Regarding future applications, it was investigated with which individual vegetation–structural distance or similarity measure the highest accuracy can be achieved in determining the current ecosystem type. Furthermore, agreement or deviation of the ecological coordinates, calculated by the indicator value of the vegetation, from the ecological coordinates, determined by assigning the ecosystem type, was investigated. Finally, possible disharmonies in the nutrient balance, namely the mismatch between the acid–base status described by the pH value and the C/N ratio, were investigated using the indicator value models.
Materials and methods
A total of 105 vegetation relevés from the Kellerwald National Park (Hesse, federal state of Germany) were available for validation purposes. These were digitised so that they could be evaluated numerically using the indicator model developed. Physical or chemical measured values were not available for further verification of the results, so that the investigations were limited exclusively to an evaluation of the vegetation–structural indicators collected by use of different methods.
For each study area, the developed models were used to calculate indicator values for the chemical indicators C/N ratio, pH value (1/n KCl) and base saturation (V value) for the uppermost 5 cm of the topsoil. Furthermore, the water balance and moisture levels scaled between 1 (dry) and 9 (very wet) according to Hofmann [14] were modelled.
The vegetation–structural distance measures were calculated with a computer program developed at W.I.E. [23], which was developed in the scientific programming language IDL 8 (EXELIS). This program was used to calculate the similarity or distance between the vegetation composition of the study areas and the mean coverage of soil in % development of 61 forest ecosystem types (reference conditions from the period up to 1990) that have been parameterised to date [37]. The Kullback information [20, 27] was calculated as the first vegetation–structural distance measure:
The pi denotes the percentage development in coverage of soil in % of the species occurring on the area (indicated with i), the \(p_{i}^{{\text{O}}}\) denote the reference condition of the vegetation. The coverage values totalled for all types are standardized to 1, that is
In addition to this measure, which is further referred to as the absolute Kullback distance, a relative Kullback distance was determined, which was defined as the quotient between the absolute Kullback distance of the study area to the mean species quantity distribution of the reference state and the maximum Kullback distance of the respective ecosystem type. The maximum Kullback distance of the ecosystem type results from the mean value of all individual relevés assigned to the reference condition plus the standard deviation of the individual values [23]. The relative Kullback distance of a recording is less than or equal to 1, if it lies within the interval defined by the standard deviation of the respective ecosystem type. With the help of this standardization, the different variance of different ecosystem types, which results primarily from their definition, is taken into account.
As a third vegetation structural measure, the modified Sörensen index was calculated, which represents the percentage agreement of the current quantity development of vegetation with the mean quantity development of the type [20, 23]:
To each study plot the ecosystem type was assigned with the smallest absolute Kullback distance, the smallest relative Kullback distance and the largest percentage agreement of similarity indices. Furthermore, a nutrient cycle type ([23]: Annex A3) was derived from the indicator values of the topsoil condition and compared with the nutrient cycle coordinates of the determined ecosystem types. Finally, the water balance type determined from the indicator value model was compared with the water balance coordinate of the ecosystem types determined. The most likely ecosystem type was the one with the highest agreement in the five characteristics mentioned above.
Results and discussion
For 83 of the 105 study plots for which digitised vegetation surveys were available, the current ecosystem type could be determined by comparison with the 61 reference conditions parameterised to date. The remaining 22 study areas refer either to nonforest areas (woody or open land areas, no tree layer recorded in the vegetation surveys) or to forest ecosystem types for which the reference condition could not yet be quantified. Within the 83 study areas that could be assigned to an ecosystem type, there are several areas that are in succession towards potential natural vegetation. These were assigned to the type for which the highest agreement was found with regard to the three vegetation–structural measures and the two indicator values calculated by the model proposed above.
Indicator values for the nutrient cycle type were determined for 105 (C/N ratio and pH value) or 102 (V value) and 101 (moisture index) areas, respectively. For 57% of all investigated areas, agreement was found in four or five of the investigated characteristics (Fig. 4) and thus a high degree of certainty in determining the most probable ecosystem type. Only for 9% of all investigated areas the most probable ecosystem type was determined from only two coinciding characteristics.

Determination of the most likely ecosystem type from vegetation structure distance or similarity measures as well as from modelled topsoil parameters
Figure 5 shows that all applied vegetation–structural distance and similarity measures are basically suitable for identifying the ecosystem type and thus for assigning the respective reference condition. In two-thirds to three-quarters of all cases, complete agreement between the identified ecosystem type and the most likely ecosystem type is achieved. If only one vegetation–structural measure is used, in almost all cases where there is no complete agreement, there is a deviation of at most one household and/or moisture level, i.e. a directly adjacent ecosystem type in the ecogram is assigned. The highest accuracy is achieved with the absolute Kullback distance.

Correspondence or deviation of the ecosystem type, determined with different vegetation structure distance or similarity measures with the most likely ecosystem type
The comparison of the moisture balance levels determined by the indicator value of the vegetation with the water balance type of the ecosystem type reveals complete agreement in 60% of all cases investigated, and a deviation by one level in 31% (scaling of the water balance levels between 1 and 9). The deviations scatter evenly in the direction of drier or wetter, a systematic deviation is not discernible (Fig. 6).

Differences between the soil moisture characteristic of the ecosystem type and the soil moisture value determined by our model
A comparison of the nutrient cycle types determined by our model with the nutrient cycle type corresponding to the ecosystem type shows complete agreement in 84% of all cases investigated (Fig. 7). The statistical deviations of the distribution show a slight right skewness, i.e. in almost all cases in which a deviation occurs, the nutritional status determined by modelled indicator values is above the reference status of the ecosystem type. In these cases, the ecosystem type was determined from all three indicator values of the chemical topsoil condition (C/N, pH, V value, indicator ranges according to Jenssen et al. [23], whereby in cases of nonconformity the pH value was not taken into account due to its higher variability. However, if only the deviation of the pH indicator values from the respective indicator ranges defined for the nutrient cycle levels is considered, a clear left skew of the distribution results (Fig. 8). In 69% of all cases there is agreement, in 27% of all cases the pH value determined by indicator value is lower than the reference condition of the ecosystem type, in 4% of all cases only it is higher.

Difference between the nutrient cycle code of the ecosystem type and the nutrient cycle type determined by our model

Differences between typical pHs(1/n KCl) of the ecosystem types and pH of the topsoil, determined by our indicator value model
The deviations of the pH indicator values of the topsoils from the nutrient-cycle level corresponding to the indicator value of the C/N ratio were analysed (Fig. 9). In this case there is a very pronounced left skewness of the distribution, i.e. more than half of all investigated topsoil conditions show a pronounced disharmony between pH value and C/N ratio in relation to the nutrient cycle types serving as reference.

Differences between typical C/N ratios of the ecosystem types and C/N ratios of the topsoil, determined by our indicator value model
Finally, the climate coordinate of the most probable ecosystem type was compared with the climatic assignment resulting from the location of the investigated areas on the map of current forest ecosystem types. The study areas are divided into the category “mountain forest locations” (ecosystem coding Dg, D1) and the category “lower mountain forest locations” (ecosystem coding Eg, Eb). In 39% of all cases, the classification made by the mapping was confirmed by the determination of the ecosystem type. For 55% of all investigated areas, the mapped altitude “lower mountain forest locations” was replaced by an ecosystem type of the category “mountain forest locations”. Conversely, in 6% of all cases, classifications mapped as “mountain forest locations” were replaced by an ecosystem type of the category “lower mountain forest locations”. Overall, the mapping revealed a focus on the category “lower mountain forest locations” with 73% of all areas, while 75% of all areas were assigned to “mountain forest locations” as a result of ecosystem determination using vegetation–structural distance measures.
The present investigations are the first to apply the numerical identification method of the current ecosystem type, which is used to establish a reference condition for the assessment of ecosystem integrity, and the models to calculate the vegetation indicator values for the topsoil condition over a larger contiguous area. Comprehensive proof of practical applicability could be provided if, in addition to the vegetation surveys, measurements of the topsoil condition and the climatic conditions on the experimental plots were also included in the evaluation for the investigated plots. As such data were not available, this was not yet possible in this evaluation. Therefore, we recommend to complete vegetation surveys with measurement of the relevant soil parameters wherever possible.
Although the investigations of this study refer exclusively to an evaluation of the vegetation relevés, they are appropriate to demonstrate the practical applicability and validity of the approach. The determination of the ecosystem type, which is clearly defined by its ecological coordinates in forest vegetation, was carried out using different methodological approaches. On the one hand, the ecosystem type was determined using structural measures of the vegetation. These measures are nonspecific with regard to the climatic and edaphic characteristics and process characteristics of ecosystems. The location in the ecogram, the multidimensional ecological state space, is based exclusively on the classification system applied in deriving the ecosystem types in this study, which is based on a combination of tabulation and expert knowledge and aims at the homogeneity of vegetation–structural characteristics [23, 37]. The differentiation of the intervals of the chemical topsoil indicators as reported by Jenssen et al. [23] was not carried out specifically for individual ecosystem types, but with the help of measured values and expert knowledge on the overall vegetation of forests and woodlands. On the other hand, the ecological coordinates of the ecosystem type were determined on the basis of indicator value models of the vegetation. These indicator value models are based on a data set of approx. 1600 measured values of the topsoil condition or, in the case of the water balance levels, on moisture indicators determined by expert opinions, which were assigned to individual plant species, but not to vegetation units or ecosystem types. The synthesis of the physical and chemical state indicators assigned to the study areas is carried out by multiplicatively linking the intervals of indicator values assigned to the individual plant species [20]. The high consistency of the results obtained with these independent methods, as shown in Figs. 4, 5, 6, 7, proves the consistency of the applied methods and confirms the validity and practicality of both the ecosystem classification and the indicator value models. The results obtained are dependent on the quality of the vegetation surveys, which could not be verified within the framework of our investigations. As mentioned above, additional validation using physical and chemical measurement data is desirable.
Different structural measures of the vegetation were used to identify the current ecosystem type. The absolute Kullback distance is an information–theoretical measure whose use does not require any algebraic, topological or order structure of the state space [4]. With this distance measure, differences in the range of medium volume development are emphasized, while species with low abundance and low consistency as well as dominant species of high area coverage have less influence. In the consequence, this measure mainly includes characteristic combinations of species with medium abundance. With the relative Kullback distance, a combination is carried out with a statistical measurement number, the standard deviation of the individual images on which the reference condition is based. Thus, the absolute distances in the high-dimensional state space of the vegetation are normalized with respect to the respective type, whose variance depends on the homogeneity of the respective type and thus also on the number of individual images underlying the reference state. Compared with the absolute Kullback distance, this measure prefers broadly defined and thus rather inhomogeneous ecosystem types. The third measure, the modified Sörensen index, describes the percentage agreement in the area coverage of the different plant species. The species with high abundance are particularly important here, so that this measure is particularly suitable, for example, for distinguishing between forests with potential natural vegetation and cultivated forests with a vegetation deviating from potential natural one or between main groups of forests that differ in the composition of the tree layer, while edaphic or climate-related differences, which are reflected above all in the characteristic species combination of the ground vegetation, are valued less than applying the Kullback distance.
The high degree of agreement in all three structural measures expressed in Fig. 5 indicates above all that the homogeneity of ecosystem types is relatively high. It thus reaffirms that the ecosystem typing carried out can be regarded as suitable for a nationwide assessment of ecosystem integrity. Since the absolute Kullback distance is used to achieve the highest accuracy, the identification of the ecosystem type in practical application can only be carried out with this measure. With the emphasis on characteristic combinations of species of medium abundance, the climatic and edaphic differences which are responsible for the differentiation of vegetation at the chosen level of classification are apparently best captured.
The agreement of the household levels derived from the indicator values of the vegetation is more pronounced in the case of the nutrient balance (Fig. 7) than for the water balance (Fig. 6). On the one hand, this is due to the fact that a total of three indicators are used for the derivation of the nutrient balance type, while only one indicator is used for the derivation of the water balance type. On the other hand, the amplitudes of the topsoil chemical indicators were calculated using statistical methods as probability distributions of measured values, while the amplitudes of the moisture indicators were derived as intervals of estimated figures.
The findings shown in Figs. 7, 8, 9 indicate pronounced disharmonies in the condition of the topsoil on a large part of the investigated areas. These disharmonies manifest that C/N ratio and pH value correspond to different nutrient balance levels (Fig. 9). Possible causes may be atmospheric deposition in the past. This could be both acid inputs such as sulphur compounds and nitrogen inputs, which have both an eutrophic and acidifying effect. Since a narrowing of the C/N ratio has a greater effect on vegetation than pure acidification, eutrophic inputs in the past could also have led to a change in ecosystem types, so that there is currently a higher correlation between C/N ratio and ecosystem type (Fig. 7) than between pH value and ecosystem type (Fig. 8). More detailed information on the causes of disharmonies could be obtained with the methods presented from time series of the vegetation development of individual areas.
In the ecosystem classification carried out in previous studies by Jenssen et al. [23], the lowlands were combined with the lower highlands on a large-climatic scale and separated from the highlands. In the vegetation composition, the majority of the areas showed a stronger affinity to the mountain forests than to the lowland forests, whereby often similar vegetation-structural distances to lowland and mountain forests were found. It turned out that the relative Kullback distance as compared to the absolute Kullback distance was much more frequently smaller than that to the mountain forests. This indicates a lower homogeneity of the group of lowland and lower mountain forests (level E) as compared to the mountain forests (level D), as the relative Kullback distance favours inhomogeneous ecosystem types in the selection. This finding can be explained by the combination of lowland forests and forests of the Lower Highlands in stage E. If the vegetation surveys included in the definition of the reference condition show a clear focus on the lowland forests, a smaller distance in particular of the absolute Kullback distance to the mountain forests (level D) becomes very probable. It should therefore be reconsidered whether a differentiation of the current level E into lowland forests and forests of the lower mountain range would be sensible.
Conclusions and outlook
The present study details and deepens the methodology of ecological integrity assessment developed by Jenssen et al. [23], Schröder et al. [38] and develops it into a practical system for assessing and mapping ecosystem integrity and services [37]. This article demonstrates how the reference status for a total of 61 forest ecosystem types was quantified (“Complementing the reference states of forest ecosystem types” section), indicators of soil biocenoses have been tested for possible supplementing and the indicator models for quantifying soil indicators have been extended and validated. This work has been complemented by the following investigations (documented in Schröder et al. [37] and other publications as listed below):
-
1.
105 vegetation samplings were taken to determine ecosystem types and soil indicators in the Kellerwald National Park.
-
2.
By use of the Very Simple Dynamics (VSD) model soil indicators were modelled at 15 sites in Germany each representing one ecosystem type.
-
3.
A fuzzy rule-based model for spatio-temporal estimations of ecological soil moisture according to Hofmann [14] was developed and tested at the federal and regional level [31,32,33].
-
4.
The map of the current near natural ecosystems in Germany (1: 500,000, [23]) was validated by means of empirical–statistical methods. The method for assessing ecosystem integrity has been enhanced and tested at the site and regional level. Possibilities for linking forest ecosystem types with habitat types according to the Habitats Directive are shown.
-
5.
Finally, a feasibility study was conducted to link information on conditions of forest ecosystems and the provision of ecosystem services.
As shown in “Supplementing the information on reference conditions by soil biocenosis data” section, a reliable indication of continuity and abundance values for ecosystem types was not possible. A sufficient number of observations are available for some ecosystem types. This applies above all to the black alder forest ecosystems and the oak and hornbeam ecosystems. For a derivation of reference states, however, it is necessary to evaluate the primary literature indicated for each observation. While with the W.I.E. indicator model one third more of the variance of the forest top soil C/N ratio could be explained compared to the assessment with Ellenberg indicator values, both approaches are comparable and can be parallelized (“Validation and extension of the W.I.E. indicator model” section).
The results of the investigation of vegetation analyses in the Kellerwald National Park (Hesse, federal state of Germany) have confirmed the large-scale applicability of the vegetation indicator value models and the methods for determining the ecosystem type using vegetation–structural distance measures (“Verification of indicator models” section). The absolute Kullback distance proves to be a suitable measure for identifying the current ecosystem type, which allows an assignment of the reference states for an assessment of ecosystem integrity and services. The results also show the significance and potential of the methods presented for cost-effective area-wide forest condition monitoring with early warning function on the basis of periodic vegetation analyses.
Availability of data and materials
The datasets generated and/or analysed during the current study are publicly available on ZENODO® [25].
Notes
Statistical Regional climate model [34].
Representative Concentration Pathways [18].
The International Co-operative Programme on Assessment and Monitoring of Air Pollution Effects on Forests is aiming at a comprehensive compilation of information on the condition of forests in Europe and beyond under the Convention on Long-range Transboundary Air Pollution of the United Nations Economic Commission for Europe. ICP Forests monitors forest condition at two monitoring intensity levels: The Level I monitoring is based on 5852 observation plots (as at 2019) on a systematic transnational grid of 16 × 16 km throughout Europe and beyond to gain insight into the geographic and temporal variations in forest condition. The Level II intensive monitoring comprises 623 plots (as at 2018) in selected forest ecosystems with the aim to clarify cause-effect relationships. At present 42 countries in Europe and beyond participate in ICP Forests (http://icp-forests.net).
Abbreviations
- BfN:
-
German Federal Agency for Nature Conservation
- Ca:
-
Calcium
- Corg :
-
Organic carbon
- DKF:
-
Soil moisture index
- EUNIS:
-
European Nature Information System
- HabT:
-
Habitat types
- ICP Forest:
-
International Cooperative Programme on Assessment and Monitoring of Air Pollution Effects on Forests
- K:
-
Potassium
- Mg:
-
Magnesium
- N:
-
Nitrogen
- NPP:
-
Net primary production
- NZm:
-
Arithmetic mean of Ellenberg–N indicator values
- P:
-
Phosphorus
- RCP:
-
Representative Concentration Pathways
- STAR:
-
Statistical Regional climate model
- V :
-
Base saturation
- W.I.E.:
-
Waldkunde-Institut Eberswalde (Forest Science Institute Eberswalde
References
Bücking W (1993) Stickstoff-Immissionen als neuer Standortfaktor in Waldgesellschaften (Nitrogen immissions as a new location factor in forest communities). Phytocoenologia 23:65–94
CCE (2012) Modelling and mapping of atmospherically-induced ecosystem impacts in Europe, CCE status report 2012. Coordination Centre for Effects, RIVM, Bilthoven, The Netherlands, p 144
Diekmann M (2003) Species indicator values as an important tool in applied plant ecology—a review. Basic Appl Ecol 4:493–506
Ebeling W, Freund J, Schweizer F (1998) Komplexe Strukturen: Entropie und Information (Complex structures: entropy and information). Teubner, Stuttgart, Leipzig
EEA (2013) EUNIS habitat classification 2012—a revision of the habitat classification descriptions. https://eunis.eea.europa.eu/references/2416. Accessed 24 Sept 2020
Ellenberg H (1985) Veränderungen der Flora Mitteleuropas unter dem Einfluß von Düngungen und Immissionen (Changes in Central European flora under the influence of fertilization and immissions). Schweizerische Zeitschrift für Forstwesen 136(1):19–39
Ellenberg H, Weber HC, Düll R, Wirth V, Werner W, Paulissen D (1992) Zeigerwerte von Pflanzen in Mitteleuropa (Indicator values of plants in Central Europe). Scr Geobot 18:1–258
Ellenberg H, Weber HE, Düll R, Wirth V, Werner W, Paulissen D (2001) Zeigerwerte von Pflanzen in Mitteleuropa (Indicator values of plants in Central Europe). Scr Geobot 18:1–262
Ewald J, Hennekens S, Conrad S, Wohlgemuth T, Jansen F, Jenssen M, Cornelis J, Michiels HG, Kayser J, Chytrý M, Gégout JC, Breuer M, Abs C, Walentowski H, Starlinger F, Godefroid S (2013) Spatial and temporal patterns of Ellenberg nutrient values in forests of Germany and adjacent regions—a survey based on phytosociological databases. Tuexenia 33:93–109
Graefe U (1993) Die Gliederung von Zersetzergesellschaften für die standortsökologische Ansprache (The structure of decomposer societies for site-specific ccological evaluation). Mitteilungen der Deutschen Bodenkundlichen Gesellschaft 69:95–98
Graefe U (1995) Gibt es bodentyp-spezifische Tiergesellschaften? (Are there soil type specific animal societies?). Mitteilungen der Deutschen Bodenkundlichen Gesellschaft 75:11–14
Graefe U, Belotti E (1999) Strukturmerkmale der Bodenbiozönose als Grundlage für ein natürliches System der Humusformen (Structural characteristics of soil biocoenosis as a basis for a natural system of humus forms). Mitteilungen der Deutschen Bodenkundlichen Gesellschaft 89:181–184
Graefe U, Beylich A (2006) Humus forms as tool for upscaling soil biodiversity data to landscape level? Mitteilungen der Deutschen Bodenkundlichen Gesellschaft 108(6–7108):6–7
Hofmann G (2002) Entwicklung der Waldvegetation des nordostdeutschen Tieflandes unter den Bedingungen steigender Stickstoffeinträge in Verbindung mit Niederschlagsarmut. Mit Anlagen (Development of the forest vegetation of the north-eastern German lowlands under the conditions of rising nitrogen inputs combined with low precipitation. With supplements). In: Anders S et al (eds) Ökologie und Vegetation der Wälder Nordostdeutschlands (Ecology and vegetation of the forests of north-eastern Germany), vol 24–41. Dr. Kessel, Oberwinter, pp 201–283. www.forstbuch.de
Hofmann G, Heinsdorf D, Krauss HH (1990) Wirkung atmogener Stickstoffeinträge auf Produktivität und Stabilität von Kiefern-Forstökosystemen (Effect of atmogenic nitrogen inputs on productivity and stability of pine forest ecosystems). Beiträge für die Forstwirtschaft 24(2):59–73
Hofmann G, Jenssen M (2005) Potentielle natürliche Waldvegetation und Naturraumpotentiale: Quantifizierung natürlicher Potentiale der Nettoprimärproduktion und der Kohlenstoffspeicherung (Potential natural forest vegetation and natural landscape potential: quantification of natural potential of net primary production and carbon storage). In: Bohn U, Hettwer C, Gollub G (eds) Anwendung und Auswertung der Karte der natürlichen Vegetation Europas (Application and analysis of the map of the natural vegetation of Europe), vol 156. (Bundesamt für Naturschutz)–BfN-Skripten, Bonn, pp 411–428
ICP Forests (2014) International co-operative programme on assessment and monitoring of air pollution. Data request on Level II data. http://icp-forests.net/page/data-requests. Accessed 03 Apr 2014
IPCC (2014) Climate change 2014: synthesis report. In: Pachauri RK, Meyer LA (eds) Contribution of working groups I, II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change. IPCC, Geneva, p 151
Jenssen M (2009) Assessment of the effects of top-soil changes on plant species diversity in forests, due to nitrogen deposition. In: Hettelingh JP, Posch M, Slootweg J (eds) Progress in the modelling of critical thresholds, impacts to plant species diversity and ecosystem services in Europe: CCE Status report 2009. Coordination Centre for Effects, pp 83–100. www.rvim.nl/cce
Jenssen M (2010) Modellierung und Kartierung räumlich differenzierter Wirkungen von Stickstoffeinträgen in Ökosysteme im Rahmen der UNECE-Luftreinhaltekonvention. Teilbericht IIII: Modellierung der Wirkung der Stickstoff-Deposition auf die biologische Vielfalt der Pflanzengesellschaften von Wäldern der gemäßigten Breiten (Modelling and mapping of spatially differentiated effects of nitrogen inputs into ecosystems within the framework of the UNECE Air Pollution Control Convention. Substudy IIII: Modelling the effect of nitrogen deposition on the biological diversity of plant communities in temperate forests). UBA-Texte 09/2010_1-72.
Jenssen M, Hofmann G (2004) Erste Ergebnisse der Ökosystemaren Umweltbeobachtung in Waldökosystemen des Biosphärenreservates Spreewald (First results of ecosystem environmental monitoring in forest ecosystems of the Spreewald biosphere reserve). Beiträge für Forstwirtschaft und Landschaftsökologie 38:201–210
Jenssen M, Hofmann G (2005) Einfluss atmogener Stickstoffeinträge auf die Diversität der Vegetation im Nordosten Deutschlands (Influence of atmogenic nitrogen inputs on the diversity of vegetation in north-eastern Germany). Beiträge für Forstwirtschaft und Landschaftsökologie 39:132–141
Jenssen M, Hofmann G, Nickel S, Pesch R, Riediger J, Schröder W (2013) Bewertungskonzept für die Gefährdung der Ökosystemintegrität durch die Wirkungen des Klimawandels in Kombination mit Stoffeinträgen unter Beachtung von Ökosystemfunktionen und -dienstleistungen (Assessment concept for the threat to ecosystem integrity posed by the effects of climate change in combination with substance inputs, taking into account ecosystem functions and services). UBA Texte 87/2013:1–381
Jenssen M, Hofmann G, Pommer U (2007) Die natürlichen Vegetationspotentiale Brandenburgs als Grundlage klimaplastischer Zukunftswälder (Brandenburg's natural vegetation potentials as the basis for climate–plastic future forests). Beiträge zur Gehölzkunde, pp 17–29
Jenssen M, Nickel S, Schröder W (2019) 61 reference states for assessing the ecological integrity of forest and forest ecosystems, link to research data (version v1). ZENODO. https://doi.org/10.5281/zenodo.2582888
Jenssen M, Schröder W, Nickel S (2015) Typisierung von Wald- und Forstökosystemen als Grundlage zur Einstufung ihrer Integrität. Integrität von Wald- und Forstökosystemen unter dem Einfluss von Klimawandel und atmosphärischen Stickstoffeinträgen (Classification of forest and forest ecosystems as a basis for classifying their integrity. Integrity of forest and forest ecosystems under the influence of climate change and atmospheric nitrogen inputs)—Teil I. Naturschutz und Landschaftsplanung 47(12):391–399
Kullback S (1951) Information theory and statistics. Wiley, New York
Ludwig G, Schnittler M (1996) Rote Liste gefährdeter Pflanzen Deutschlands (Red list of endangered plant species in Germany). Schriftenreihe für Vegetationskunde 28:744
Nickel S, Schröder W, Jenssen M (2015) Veränderungen deutscher Wälder durch Klimawandel und Stickstoffdeposition (Changes in German forests due to climate change and nitrogen deposition). Schweizerische Zeitschrift für Forstwesen 166(5):325–334
Nickel S, Schröder W, Jenssen M (2016) Prädiktive Kartierung und Analyse klimawandelbedingter Veränderungen von Wald- und Forstökosystemen. Integrität von Ökosystemen unter dem Einfluss von Klimawandel und atmosphärischen Stickstoffeinträgen—Teil III (Predictive mapping and analysis of climate change-related changes in forest and forest ecosystems. Integrity of ecosystems under the influence of climate change and atmospheric nitrogen inputs—Part III). Naturschutz und Landschaftsplanung 48(2):46–52
Nickel S, Schröder W (2017) Fuzzy modelling and mapping soil moisture for observed periods and climate scenarios. An alternative for dynamic modelling at the national and regional scale? Ann For Sci 74:71
Nickel S, Schröder W (2017) Fuzzy modelling and mapping soil moisture in Germany, link to research data and scientific software. ZENODO. https://doi.org/10.5281/zenodo.1320219
Nickel S, Schröder W (2018) Ecological soil moisture data across Germany, link to shape files. PANGAEA. https://doi.org/10.1594/PANGAEA.891249
Orlowsky B, Gerstengarbe FW, Werner P (2008) A resampling scheme for regional climate simulations and its performance compared to a dynamical RCM. Theor Appl Climatol 92:209–223
Posch M, Reinds GJ (2009) A very simple dynamic soil acidification model for scenario analyses and target load calculations. Environ Model Softw 24:329–340
Riecken U, Finck P, Raths U, Schröder E, Ssymank A (2006) Rote Liste der gefährdeten Biotoptypen Deutschlands (Red list of endangered biotope types in Germany). Naturschutz und Biologische Vielfalt 34:1–318
Schröder W, Nickel S, Jenssen M, Hofmann G, Schlutow A, Nagel HD, Burkhard B, Dworczyk C, Elsasser P, Lorenz M, Meyerhoff J, Weller P, Altenbrunn K (2019) Anwendung des Bewertungskonzepts für die Ökosystemintegrität unter Berücksichtigung des Klimawandels in Kombination mit Stoffeinträgen (Application of the ecosystem integrity assessment concept for climate change in combination with substance inputs). UBA-Texte 97/2019:1–504
Schröder W, Nickel S, Jenssen M, Riediger J (2015) Methodology to assess and map the potential development of forest ecosystems exposed to climate change and atmospheric nitrogen deposition: a pilot study in Germany. Sci Total Environ 521–522:108–122
Schröder W, Riediger J, Nickel S, Jenssen M (2016) Projektion zukünftiger Ökosystemzustände unter dem Einfluss von Klimawandel und atmosphärischen Stickstoffeinträgen. Integrität von Wald- und Forstökosystemen unter dem Einfluss von Klimawandel und atmosphärischen Stickstoffeinträgen—Teil II (Projection of future ecosystem conditions under the influence of climate change and atmospheric nitrogen inputs. Integrity of forest and forest ecosystems under the influence of climate change and atmospheric nitrogen inputs—Part II). Naturschutz und Landschaftsplanung 48(1):22–28
Senckenberg Museum für Naturkunde Görlitz (2018) Edaphobase. Occurrence dataset. https://doi.org/10.15468/rk9xc7. Accessed 20 Apr 2018
VDI (Verein Deutscher Ingenieure) (2008) Vegetation als Indikator für Stickstoffeinträge - Bewertung der Stickstoffverfügbarkeit durch Ellenberg-Zeigerwerte der Waldbodenvegetation, Kommission Reinhaltung der Luft im VDI und DIN-Normenausschuss KRdL (Vegetation as an indicator for nitrogen inputs—evaluation of nitrogen availability by Ellenberg indicator values of forest floor vegetation). Beuth, Berlin, pp 1–3
WMO (World Meteorological Organisation) (2017) WMO guidelines on the calculation of climate normals. WMO-No. 1203, 2017 edition. Geneva, Switzerland, pp 1–18
Acknowledgements
We would like to thank the German Environment Agency (Dessau-Roßlau) for its financial support and technical advice.
Funding
Open Access funding enabled and organized by Projekt DEAL. Federal Environment Agency, Dessau-Roßlau, Germany.
Author information
Authors and Affiliations
Contributions
MJ conducted the investigations presented. WS headed the study and drafted the manuscript. All authors participated in writing the article. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Means and standard deviations for nutrient and water balance indicators (C/N ratio, pH in 1/10 KCl, base saturation (V-value) and soil moisture index), describing reference states for 61 forest ecosystem types prior to 1990. Table S2. Maximum of Kullback distance of the single vegetation samples to the mean species distribution and minimum of percentage similarity [20] of the single vegetation samples with the average species distribution to describe the reference states before 1990 for 61 forest ecosystem types. Table S3. Assignment of observations to forest ecosystem types and groups. Table S4. Characteristics of the nutrient cycle types (third digit in the ecosystem type code, [23]): humus form, C/N ratios in the topsoil and N indicator values according to Ellenberg et al. [8].
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Jenssen, M., Nickel, S., Schütze, G. et al. Reference states of forest ecosystem types and feasibility of biocenotic indication of ecological soil condition as part of ecosystem integrity and services assessment. Environ Sci Eur 33, 18 (2021). https://doi.org/10.1186/s12302-021-00458-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12302-021-00458-2