
Abstract
Background
Organic cation transporters (OCTs) determine not only physiological processes but are also involved in the cellular uptake of anticancer agents. Based on microarray analyses in hepatocellular carcinoma (HCC), SLC22A1/OCT1 mRNA seems to be downregulated, but systematic protein expression data are currently missing. Moreover, the underlying molecular mechanisms responsible for altered SLC22A1 expression in HCC are not fully understood. Therefore, we investigated the role of DNA methylation in the transcriptional regulation of the family members SLC22A1/OCT1, SLC22A2/OCT2 and SLC22A3/OCT3 in HCC.
Methods
Semiquantitative immunohistochemistry of SLC22A1 protein expression was performed in paired HCC and histological normal adjacent liver tissues (n = 71) using tissue microarray analyses, and the results were correlated with clinicopathological features. DNA methylation, quantified by MALDI-TOF mass spectrometry and gene expression of SLC22A1, SLC22A2 and SLC22A3 were investigated using fresh-frozen HCC (n = 22) and non-tumor adjacent liver tissues as well as histologically normal liver samples (n = 120) from a large-scale liverbank.
Results
Based on tissue microarray analyses, we observed a significant downregulation of SLC22A1 protein expression in HCC compared to normal adjacent tissue (P < 0.0001). SLC22A1 expression was significantly inverse correlated with expression of the proliferation marker MIB1/Ki-67 (rs = -0.464, P < 0.0001). DNA methylation of SLC22A1 was significantly higher in HCC compared with non-tumor adjacent liver tissue and was lowest in histologically normal liver tissue. Methylation levels for SLC22A1 in combination with RASSF1A resulted in a specificity of > 90% and a sensitivity of 82% for discriminating HCC and tumor-free liver tissue.
Conclusions
DNA methylation of SLC22A1 is associated with downregulation of SLC22A1 in HCC and might be a new biomarker for HCC diagnosis and prognosis. Moreover, targeting SLC22A1 methylation by demethylating agents may offer a novel strategy for anticancer therapy of HCC.
Similar content being viewed by others


Background
Hepatocellular carcinoma (HCC) is the sixth most common cancer worldwide and the third most common cause of cancer-related death. Understanding of the molecular pathogenesis of HCC has improved, and some progress has been made in translating these findings into clinical practice [1]. Therapeutic options include sorafenib, as first effective systemic treatment against HCC, as well as radioembolization and oncolytic approaches. Currently, there are more than 150 ongoing clinical trials. But, in the main, these are not integrating information about the molecular pathogenesis of HCC [2]. For instance, the cellular uptake of anticancer drugs is an important first step in the mechanism of drug action. Therefore, multidrug resistance can result not only from increased efflux but also reduced uptake of drug into tumor cells. Uptake transport in hepatocytes is mediated by members of the solute carrier (SLC) family. Organic cation transporters (OCTs) are involved in the transport of a variety of endogenous and exogenous organic cationic compounds and thus the expression of these transporters is suggested to be a significant determinant of physiological functions in different organs [3–5]. OCTs are also important drug targets since several clinically relevant agents, including anticancer drugs (for example, platinum agents), are substrates of OCTs.
Expression profiles among the three OCT family members OCT1 (encoded by SLC22A1), OCT2 (SLC22A2) and OCT3 (SLC22A3) are extremely diverse in different tissues [6]. SLC22A1 is expressed mainly in normal human liver, and SLC22A2 is predominately expressed in kidney [3–5, 7, 8]. There is already evidence that OCTs are differentially expressed in tumor tissues, and based on microarray data, SLC22A1 mRNA expression is downregulated in HCC [9–11]. However, systematic analyses of expression of SLC22A1, SLC22A2 and SLC22A3 have not been performed so far in HCC. Moreover, the underlying mechanisms responsible for altered expression of SLC22A1 in HCC compared with normal liver are poorly understood.
Epigenetic alterations, including DNA methylation and histone modifications, are important mechanisms in tumorigenesis. DNA hypermethylation-induced transcriptional silencing of tumor suppressor and DNA repair genes is a frequent phenomenon [12]. There is already proof of principle for the clinical value of methylation markers for classification (for example, colorectal cancer [13, 14]), prognosis [15] and prediction of therapeutic response [12]. Thus, identification of specific expression-regulating core gene regions (that is, CpG dinucleotides) in the promoter is essential. With respect to OCTs, there is evidence that kidney-specific expression of SLC22A2 is regulated by DNA methylation [16]. Currently there are no data on whether downregulation of SLC22A1 in HCC could be explained by hypermethylation of the SLC22A1 promoter. Elucidating the precise mechanism for downregulation of SLC22A1 in HCC is especially important because it is possible to overcome gene silencing with demethylating agents like decitabine, which opens new therapeutic strategies for HCC.
Therefore, in the present study we performed systematic immunohistochemical analysis of SLC22A1 protein in well-characterized, paired HCC and corresponding non-tumor tissues and correlated the results with clinicopathological data. We elucidated systematically the influence of DNA methylation on the transcriptional regulation of SLC22A1, SLC22A2 and SLC22A3 in HCC tissues and in tissues in a large human liver bank.
Materials and methods
Tumor and non-neoplastic human tissues
An overview about different sample sets used in the present study is given in Table S1 in Additional file 1. HCC tissues and corresponding adjacent non-tumor liver tissues from patients who underwent surgical resection between 2001 and 2007 were collected at the Department of Internal Medicine I, University Hospital Regensburg, Regensburg, Germany and used for tissue microarray analysis (TMA), as previously described [17, 18]. Detailed clinicopathological characteristics of patients (n = 71) are given in Table S2 in Additional file 1. Additionally, paired fresh-frozen HCC tissues (n = 15) and adjacent non-tumor liver samples were obtained from the same institution [19]. As additional samples, seven HCCs were obtained from OriGene (OriGene, Rockville, MD, USA) and re-examined by a pathologist. The patient characteristics are summarized in Table S3 in Additional file 1. Finally, as a control group, 120 histologically normal fresh-frozen liver tissues (IKP-liverbank; Table S4 in Additional file 1) were investigated, which were collected at the Department of General, Visceral and Transplantation Surgery (Campus Virchow, University Medical Center Charité, Humboldt University Berlin, Germany) as previously reported [20]. For systematic expression profiling of SLC22A1, SLC22A2 and SLC22A3 in normal and tumor tissues, commercial arrays including cDNA from 381 human tissues normalized to beta-actin were used (TissueScan, Origene technologies, Rockville, MD, USA). The study conforms to the ethical guidelines of the 1975 Declaration of Helsinki and was approved by the ethics committee of the University Hospital Regensburg (Regensburg, Germany), by the ethics committee of University of Tuebingen (Tuebingen, Germany) and the Charite (Berlin, Germany). Written informed consent was provided by each patient.
RNA isolation and TaqMan analysis
Total RNA was extracted from HCCs and non-tumor tissue using the mirVana™ miRNA Isolation Kit (Applied Biosystems/Ambion, Austin, TX, USA). High quality liver RNA was extracted as previously described [20], and only high-quality RNA preparations with RNA Integrity Number (RIN) assignment (> 7), as determined by Agilent Bioanalyzer (Nano LabChip Kit, Agilent Technologies, Waldbronn, Germany), were used. Total RNA was used for cDNA syntheses using the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA). mRNA expression was quantified by TaqMan technology (for details see additional methods in Additional file 1).
Immunofluorescence microscopy of tissue samples
Localization of OCTs in liver and HCC tissues was analyzed by immunofluorescence confocal laser scanning microscopy (for details see additional methods in Additional file 1).
HCC tissue microarray analysis
TMAs were processed as previously described [17, 18]. SLC22A1 immunohistochemistry was performed using a polyclonal antiserum raised in rabbits against a synthetic peptide corresponding to the carboxy-terminal sequence of human SLC22A1 as described previously [20]. Staining of SLC22A1 was performed according to the following protocol: 5 μm TMA sections were transferred to slides (Superfrost-Plus, Langenbrinck, Teningen, Germany). Tissues were deparaffinized by passing the specimens through xylene and rehydrated through serial dilutions of ethanol (100%, 96% and 70%). Heat-induced antigen retrieval was accomplished using a solution buffer at pH 6.0 (Dako Cytomation, Glostrup, Sweden) for 30 minutes. Endogenous peroxidase was blocked with a peroxidase blocker (0.03% H2O2; Dako Cytomation) for 10 minutes. Slides were incubated for 30 minutes at room temperature with the primary antibody (OCT1/KEN, dilution 1:2,000) in an antibody diluent (Dako Cytomation). For negative control the primary antibody was omitted. The REAL-HRP system (REAL Envision Detection System, Rabbit/Mouse, Dako Cytomation) was used for detection of the antibody binding before sections were counterstained with hematoxylin and mounted. For semiquantitative immunohistochemical assessment first, staining intensity was graded according to the following scoring system: 0 = negative, 1 = low, 2 = medium and 3 = high. As previously described [21, 22], the final immunoreactivity score was calculated by multiplication of the staining intensity score (0 to 3) and the percentage of immunoreactive (stained) cells (0 to 100%), resulting in a numeric value of 0 to 300. MIB1 was analyzed applying the anti-Ki-67 antibody (clone MIB1, 1:10, final concentration of 5 μg/ml; DAKO) to assess the MIB1/Ki-67 proliferation rate as previously described [17]. Evaluation of the staining was done by two investigators blinded to the patient characteristics.
DNA methylation analysis
Quantitative DNA methylation analysis was performed with matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS; Sequenom, San Diego, CA, USA), as described in the additional methods in Additional file 1. Mass spectra were acquired with the MassArray Compact MALDI-TOF MS (Sequenom) and spectra methylation ratios were analyzed using the Epityper 1.0 software (Sequenom).
Statistical methods
Statistical analyses were performed with R-2.13.0, PASW Statistics 17.0.2 and GraphPadPrism 4.03 (GraphPad Software Inc., La Jolla, CA, USA). Statistical significance was defined as P < 0.05. For a detailed description, see the additional methods in Additional file 1.
Results
Expression profiling of SLC22A1, SLC22A2, and SLC22A3 in normal and tumor tissue
A comprehensive analysis using cDNAs from 381 human tissue samples (Table S1 in Additional file 1) from 20 different tissues revealed that SLC22A1 mRNA is expressed most prominently in normal liver and expressed at significantly lower levels in HCC tissues (P = 0.029; Figure 1A). SLC22A2 transcripts were present in human kidney, but almost absent in human normal liver, as well as in HCC tissue. SLC22A3 transcripts were detectable in a variety of human tissues, including human liver (Figure 1a), and SLC22A3 expression did not differ significantly between normal and tumor liver tissue.

Expression analysis in normal and tumor tissues. (a) SLC22A1, SLC22A2, and SLC22A3 mRNA expression in different human normal and tumor tissues of the respective organs (TissueScan, Origene Technologies, Rockville, MD, USA) was quantified using TaqMan technology. (b) SLC22A1/OCT1, SLC22A2/OCT2 and SLC22A3/OCT3 protein expression (green) was analyzed by confocal laser scanning microscopy in HCC tumor tissue (n = 7) compared with histologically normal liver tissue (n = 5, IKP-liverbank) derived from patients without hepatocellular carcinoma (HCC). Specific antibodies were used to detect SLC22A1, SLC22A2 and SLC22A3 proteins. Representative images are shown.
Moreover, specific antibodies were used to detect SLC22A1, SLC22A2 and SLC22A3 protein by immunofluoresence confocal laser scanning microscopy in HCC tumor tissue (n = 7 from HCC cohort 1; Tables S1 and S3 in Additional file 1) and histologically normal liver tissue (n = 5 from IKP-liverbank; Tables S1 and S4 in Additional file 1) derived from patients without HCC. In line with previous data, only SLC22A1 and SLC22A3 were detected in human liver [20]. In HCC tissue, SLC22A3 and, at a lower level, SLC22A1 proteins were detected. SLC22A2 was not detected in either normal or HCC tissue (Figure 1b).
Protein expression analysis of SLC22A1 in HCC
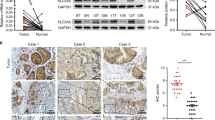
To systematically evaluate expression of SLC22A1 protein in HCC tissues, levels were analyzed by semi-quantitative immunohistochemistry using TMA of HCC and corresponding adjacent non-tumor tissues [17, 18]. Investigation of SLC22A1 protein expression was informative in 68 HCC and 67 adjacent tissues of this HCC study population (set 1; for details see Tables S1 and S2 in Additional file 1), because some of the cores on TMAs were lost or damaged during processing of the TMAs. The expression of SLC22A1 was determined by applying an immunohistochemical score, as described in the Materials and methods. Statistical analysis revealed significantly lower SLC22A1 immunoreactivity in HCC tissues compared with adjacent tissues (P < 0.0001; Figure 2a), irrespective of the presumed etiology of the HCC (for example, hepatitis B virus or hepatitis C virus; a paired comparison is shown in Figure S1 in Additional file 1). Examples of different staining intensities (0 = negative, 1 = low, 2 = medium, 3 = high) in non-tumor as well as in HCC tissue are shown in Figure 2b. SLC22A1 expression in HCC tissues, when present, was variable, ranging from at least 20 to 90% of the cells, and showed a more diffuse pattern compared with non-tumor tissue (Figure 2c). In addition, expression of SLC22A3 was analyzed for a subset of HCC samples, used for immunostaining of SLC22A1. As shown in Figure 2c, immunostaining in these samples clearly demonstrates SLC22A3 expression at the protein level.

Immunohistochemical staining of SLC22A1 protein in hepatocellular carcinoma and histological non-tumor adjacent tissue. (a) Expression of SLC22A1 is significantly decreased in hepatocellular carcinoma (HCC) versus adjacent non-tumor tissue (P < 0.0001) and individual paired results are given. For each patient, score values from both tissues are connected by a line. (b) Examples of different staining intensities (0 = negative, 1 = low, 2 = medium, 3 = high) in non-tumor as well as HCC tissues. SLC22A1 was present in all non-tumor tissues. (c) Exemplary weak immunohistochemical staining of SLC22A1 in two HCC samples is shown compared with strong staining in adjacent non-tumor liver tissue. In addition, representative examples of SLC22A3 staining in HCC tissue as well as in adjacent non-tumor liver tissue is shown. SLC22A1 and SLC22A3 are detected in the sinusoidal membrane of the hepatocytes (black arrows).
When we correlated the SLC22A1 immunoreactivity score with clinicopathological characteristics, a significant inverse correlation was found only for the MIB1/Ki-67 proliferation rate (rs = -0.464, P < 0.0001), which was determined in HCC tissue samples by immunohistochemical staining as previously described (Figure S2A in Additional file 1) [18]. Moreover, a significant difference in SLC22A1 expression between samples with low (≤5%) and high (> 5%) proliferation rate was found (P = 0.01; Figure S2B in Additional file 1). In contrast, no association was detected with etiological factors (for example, hepatitis B virus or hepatitis C virus), age at surgery or sex. Moreover, tumor grade or histological stage was not significantly associated with SLC22A1 expression, although both stage 4 tumors revealed the lowest levels of protein expression (Figure S3A,B in Additional file 1).
DNA methylation of SLC22A1, SLC22A2 and SLC22A3 in HCC and adjacent non-tumor tissues
To confirm the finding of a significantly lower expression of SLC22A1 in HCC, mRNA levels of SLC22A1, SLC22A2, and SLC22A3 were determined by real-time PCR analysis in a cohort of 22 fresh-frozen HCC tissues and adjacent non-tumor liver samples (HCC study population set 2; Tables S1 and S3 in Additional file 1). Moreover, expression levels were compared with histologically normal liver tissue from patients without primary liver tumors (n = 100 pooled cDNAs, IKP-liver bank; Tables S1 and S4 in Additional file 1). As shown in Figure 3a, SLC22A1 mRNA expression in HCC samples was significantly lower (P = 0.0007) compared with adjacent tissue. Generally, SLC22A3 mRNA did not differ significantly between HCC and adjacent tissue (Figure 3b). Again, the SLC22A2 mRNA levels in HCC and adjacent tissue was very low (data not shown).

Expression and DNA methylation of SLC22A1 and SLC22A3 in hepatocellular carcinoma and adjacent non-tumor tissue. (a,b) Quantitative mRNA expression of SLC22A1 (a) and SLC22A3 (b) in hepatocellular carcinoma (HCC), adjacent non-tumor tissue and histologically normal liver tissue (pooled n = 100, IKP-liverbank). Data represent median and range. (c,d) DNA methylation profiles of SLC22A1 (c) and SLC22A3 (d) in HCC and adjacent non-tumor tissue. DNA methylation (y-axis) at each individual CpG site (x-axis) is given. Methylation profiles show median levels (diamonds) at each CpG position (shaded areas are defined by 25%/75% quantiles). P-values are given for differences in methylation level for CpG sites. *P < 0.05, **P < 0.01 are given on the x-axis. (e) Correlation of SLC22A1 mRNA expression in the 22 HCC samples with DNA methylation of one individual CpG site (SLC22A1_02_CpG_5) in SLC22A1.
To evaluate whether SLC22A1 is silenced in HCC due to hypermethylation of SLC22A1 in tumor tissue, we studied tissue-specific DNA methylation. DNA methylation of SLC22A1, SLC22A2, and SLC22A3 gene promoters was studied by gene-specific amplification of bisulfite-treated DNA followed by in vitro transcription and MALDI-TOF MS analysis [23, 24]. The amplification regions were designed to cover the surrounding 5' UTRs of the target genes. In addition, promoter regions were analyzed in silico for the presence of CpG islands. In contrast to SLC22A2 and SLC22A3, no CpG island was predicted in the promoter region of SLC22A1, which is supported by genome-wide CpG island mapping analysis [25]. We were able to quantify 18 CpG sites for SLC22A1, 6 sites for SLC22A2 and 57 sites for SLC22A3 (Table S5 in Additional file 1). As described in detail in the additional methods in Additional file 1 the MALDI-TOF MS method does not always allow detection of methylation levels for each single CpG site [24], and some of the methylation values represent the methylation state of subsequent CpG sites. As shown in Figure 3c, SLC22A1 methylation was significantly increased in HCC tissues, consistent with the low level of mRNA expression. For both HCC and adjacent non-tumor liver tissue, DNA methylation of SLC22A1 was highly variable between individuals. In contrast, SLC22A3 methylation was generally not different between HCC tissues and adjacent non-tumor liver tissue (Figure 3d). DNA methylation of SLC22A2 in HCC and adjacent non-tumor liver tissue was high and showed low inter-individual variability (Figure S4A in Additional file 1).
When we correlated DNA methylation with SLC22A1 mRNA expression in the total group of 22 HCC samples (HCC study population, set 2; Tables S1 and S3 in Additional file 1), a significant inverse correlation between SLC22A1 mRNA expression and individual CpG sites within the SLC22A1 promoter region was found (Figure 3e; Table 1). There was no correlation between SLC22A1 mRNA expression and DNA methylation with clinicopathological variables like age at surgery, sex, tumor grade and histological stage and tumor size (Figure S5A-D in Additional file 1).
DNA methylation of SLC22A1, SLC22A2, and SLC22A3 in histologically normal liver tissue
Generally, aberrant DNA methylation has been demonstrated in tumor tissue and adjacent, histologically normal tissue of tumor patients [26]. We therefore examined SLC22A1, SLC22A2 and SLC22A3 DNA methylation in histologically normal liver tissue from patients without primary liver tumors (n = 100, IKP-liverbank; Tables S1 and S4 in Additional file 1) compared with histologically normal tissues derived from patients with HCC (n = 20; Tables S1 and S4 in Additional file 1). Significantly higher levels of methylation of SLC22A1 (Figures 4A and S6A in Additional file 1), but not SLC22A3 (Figure S6C in Additional file 1) were observed in adjacent tissue from primary liver tumors (n = 20) compared with normal livers without primary liver tumors (n = 100). The SLC22A2 gene region was again highly methylated (Figure S6B in Additional file 1).

DNA methylation of SLC22A1 and RASSF1A in histologically normal liver tissue. (a,b) DNA methylation profiles of SLC22A1 (a) and RASSF1A (b) in normal liver tissue (n = 100, IKP-liverbank), in histologically normal tissues derived from patients with hepatocellular carcinoma (HCC; n = 20, adjacent to tumor) and in HCC tumor tissue (n = 22). DNA methylation (y-axis) at each individual CpG site (x-axis) is given. Methylation profiles show median levels (diamonds) at each CpG position (shaded areas are defined by 25%/75% quantiles). P-values are given for differences in methylation level for CpG sites between HCC (n = 22) and histologically normal liver tissue (n = 100). *P < 0.05, **P < 0.01.
In summary, SLC22A1 DNA methylation was highest in HCC tissue (HCC study population set 2) and decreased progressively from adjacent to histologically normal tissue of patients without primary liver tumors from the IKP-liverbank (Figure 4a). Since a similar behavior has been reported for the tumor suppressor gene RASSF1A [27–29], DNA methylation levels of RASSF1A were determined in our study. As indicated in Figure 4b, we confirmed DNA hypermethylation of the tumor suppressor gene RASSF1A in HCC samples of our study. Comparable to our data on SLC22A1 methylation, DNA methylation levels of RASSF1A were higher in HCC tissue compared with adjacent non-tumor tissue (Figure 4b). However, in contrast to SLC22A1 (Figure 4a), methylation of RASSF1A was not lower in histologically normal tissue of patients without primary liver tumors (n = 100) compared with normal tissues derived from patients with HCC (n = 20, IKP-liverbank; Figure 4b).
To investigate whether DNA methylation status of SLC22A1 and RASSF1A could effectively distinguish HCC and histologically normal tissue, we determined sensitivity and specificity (see statistical methods in Additional file 1). The methylation levels of 11 selected CpG sites in SLC22A1 and two in RASSF1A together could classify HCC and normal tissues with 82% sensitivity and 96% specificity. For CpG sites of RASSF1A alone, specificity was only 86% in contrast to 96% for CpG sites of SLC22A1. The sensitivity was similar (64%) for both genes.
Discussion
Altered expression of OCT transporters is important for physiological processes and therapeutic responses to drugs. Indeed, many clinically important drugs (for example, metformin, cisplatin) are substrates for OCTs [3–5, 30]. Independent of genetic reasons for inter-individual differences in SLC22A1 and SLC22A3 expression in histologically normal liver tissue [20], there is first evidence from microarray studies that SLC22A1 mRNA expression is decreased in HCC [9–11]. This is supported by our experiments using cDNAs from 381 human tissue samples from 20 different tissues (Table S1 in Additional file 1). SLC22A1 mRNA is expressed most prominently in liver and at significantly lower levels in HCC tissues (P = 0.029; Figure 1a), which corresponds with weaker immunofluoresence staining by confocal laser scanning microscopy. There are studies to suggest that SLC22A1 is one of the most downregulated genes in HCC cells, with a gene pattern correlated to poor prognosis [10, 31, 32]. SLC22A1 also is expressed differentially in EpCAM positive, stem-cell-like-HCC and EpCAM negative mature hepatocyte-like HCC [33]. However, SLC22A1 protein expression in HCC has so far not been studied systematically and the molecular mechanisms underlying the tumor-specific expression of SLC22A1 are poorly understood.
Therefore, we investigated first protein expression in a set of 71 HCC tissues and adjacent non-tumor liver tissues by semiquantiative immunohistochemistry. These experiments indicated significantly decreased expression of SLC22A1 protein in HCC (Figure 2a). When we correlated SLC22A1 expression with clinicopathological characteristics, the most striking result was a significant inverse correlation between expression of SLC22A1 protein and the tumor proliferation rate MIB1/Ki-67. Previous findings support our observation since MIB1/Ki-67 staining is reduced in protein kinase JNK-/- knockout mice [34] and microarray expression profiling clearly indicated that HCC tissues with high activation status of JNK1 showed reduced SLC22A1 mRNA expression and poorer prognosis [10].
Since altered DNA methylation is a common mechanism in tumor development, downregulation of SLC22A1 might be caused by hypermethylation of the SLC22A1 promoter region. This prompted us to systematically quantify DNA methylation levels of SLC22A1, SLC22A2, and SLC22A3 in the surrounding 5' UTRs of the target genes by MALDI-TOF MS. Consistent with both low expression of mRNA and protein, SLC22A1 methylation was significantly increased in HCC tissues compared with adjacent non-tumor (Figure 3c) or non-tumor liver tissues derived from patients without primary liver tumors (Figure 4a). Methylation of SLC22A3 was generally not different.
Of note, altered methylation has been reported not only for tumor tissue, but also for histologically normal tissue adjacent to the tumor, and is supposed to be an early and ubiquitous event in cancer development. Interestingly, we found significantly higher levels of SLC22A1 methylation (Figure 4a) but not SLC22A3 methylation in normal liver tissue adjacent to HCC tumors. This important finding may explain the discrepancy between our data and the results of Aoki et al. [16], who reported high methylation of SLC22A1 in a collection of 11 liver samples. Since Aoki et al. used only tumor-free tissue adjacent to HCC, these data strongly support our observation that SLC22A1 is hypermethylated in HCC and to a lesser extent in the adjacent, histologically tumor-free tissue.
Thus, based on our results, methylation of SLC22A1 decreased from high levels in HCC to lower levels in adjacent, histologically normal liver and was lowest in normal tissue from livers that were not derived from patients with HCC (Figure 4a). The same methylation pattern has been reported already for some tumor suppressor genes like RASSF1A [27]. RASSF1A was the best discriminator between HCC and tumor-free liver tissue [35, 36], which is confirmed by our data showing highest methylation levels of RASSF1A in HCC compared with adjacent tumor-free liver tissue. Since early detection of aberrant DNA methylation might be beneficial for diagnosis of HCC, we suggest that SLC22A1 methylation might be a new biomarker for the diagnosis of HCC or for estimating risk for HCC in following patients at risk. This idea is supported by the observation that the MIB1/Ki-67 proliferation index correlated inversely with SLC22A1 expression in our HCC cohort. Additional analysis of whether DNA methylation effectively discriminates between HCC and histologically normal liver tissue indicated that the combination of the methylation status of SLC22A1 and RASSF1A had a higher specificity and sensitivity for this compared with each of the two markers alone. Further studies will be needed to verify these data.
Several causes for aberrant DNA methylation in HCC have been discussed. A recent study demonstrated that the methylation state in specific genes in HCC is influenced by viral infection or alcohol intake, thereby promoting hepatocarcinogenesis [28]. Genome-wide methylation analysis of human colon cancer showed that cancer-related methylation changes occur predominately at sites that distinguish normal tissues [37]. Interestingly, the same region in the SLC22A1 gene that is differentially methylated in HCC tissue in our study showed tissue-specific differences in SLC22A1 DNA methylation [38].
Since there is emerging evidence on the importance of the location of CpG dinucleotide hypermethylation in relation to gene expression but also clinicopathologic characteristics of tumors, we next performed a systematic correlation analysis between SLC22A1 mRNA expression and quantitative DNA methylation of individual CpG sites within the SLC22A1 promoter region. Using the total set of 22 fresh-frozen HCC tissues (Table S2 in Additional file 1), the CpG sites in SLC22A1 with the strongest inverse correlation with mRNA expression in HCC (Table 1) were remarkably located upstream of the translational start site in exon 1. However, the overall methylation profile for the total SLC22A1 region investigated in our study was quite similar. The underlying mechanism for this phenomenon is unknown, but recent findings indicate that especially DNA methylation of the first exon correlates with transcriptional silencing [39]. Thus, DNA methylation within the first exon may block effective transcription.
Since OCTs are important targets for various therapeutic agents, such as metformin or the platinum drugs cisplatin or picoplatin [30, 40, 41], the finding that SLC22A1 is downregulated in liver cancer due to hypermethylation may be important for alternative therapeutic approaches for treatment of HCC. We speculate that DNA methylation of SLC22A1 in human liver contributes to differences in drug response for drugs that are substrates of SLC22A1. By contrast with genetic variants, epigenetic modifications are reversible, which offers the possibility to modulate the expression of genes with demethylating agents (for example, decitabine). As a consequence, for instance, pretreatment with decitabine may increase the intracellular concentration of the SLC22A1 substrate cisplatin, thereby resulting in enhanced cytotoxicity. Of course, treatment with decitabine will not only lead to re-expression of OCTs and additional factors might be important for cisplatin efficacy, for example, the expression of the copper transporter CTR1, which alters sensitivity to cisplatin [42]. In addition, demethylation may impair expression of efflux transporters, thereby leading to an enhanced efflux and chemoresistance (reviewed in [43]). For instance, the promoter of the efflux transporter gene ABCB1, encoding P-glycoprotein, is hypomethylated in several cancer cell lines, thereby leading to a multidrug-resistance phenotype compared to drug-sensitive cell lines [44]. Moreover, the effect of decitabine treatment might lead to increased risk of cisplatin cytotoxicity in other organs, since the demethylating effect is not restricted to liver tissue.
Our study has some limitations that need to be considered. First, our study design of HCC samples was retrospective and, for instance, confounding of SLC22A1 expression cannot be excluded with certainty. Second, the number of HCC samples was small, and the relevance of the correlation analyses of distinct clinicopathological factors (for example, tumor stage, histological grade) with DNA methylation or expression is thus limited and needs further study. Nevertheless, our findings indicate for the first time that downregulation of the uptake transporter SLC22A1 in HCC is associated with DNA methylation as an epigenetic phenomenon. This finding has potential consequences for the diagnosis of HCC and for future therapeutic concepts.
Conclusions
SLC22A1 protein expression is significantly decreased in HCC compared with tumor-free adjacent liver tissue. DNA hypermethylation of individual CpG sites within the SLC22A1 gene is associated with downregulation of SLC22A1 expression in HCC. Moreover, SLC22A1 methylation decreased from high levels in HCC to lower levels in histological tumor-free adjacent liver tissue and was lowest in tumor-free liver tissue not derived from patients with HCC. Therefore, early detection of aberrant DNA methylation of SLC22A1 might be beneficial for the diagnosis of HCC or for estimating risk for HCC in following patients at risk. Since platinum drugs are substrates of SLC22A1, the modulation of gene expression by pretreatment with demethylating agents may offer novel therapeutic options in HCC, which has to be proven in further studies.
Abbreviations
- HCC:
-
hepatocellular carcinoma
- MALDI-TOF MS:
-
matrix-assisted laser desorption/ionization time-of-flight mass spectrometry
- OCT:
-
organic cation transporter
- PCR:
-
polymerase chain reaction
- SLC:
-
solute carrier
- TMA:
-
tissue microarray
- UTR:
-
untranslated region.
References
Farazi PA, DePinho RA: Hepatocellular carcinoma pathogenesis: from genes to environment. Nat Rev Cancer. 2006, 6: 674-687. 10.1038/nrc1934.
Breuhahn K, Gores G, Schirmacher P: Strategies for hepatocellular carcinoma therapy and diagnostics: Lessons learned from high throughput and profiling approaches. Hepatology. 2011, 53: 2112-2121. 10.1002/hep.24313.
Ciarimboli G: Organic cation transporters. Xenobiotica. 2008, 38: 936-971. 10.1080/00498250701882482.
Koepsell H, Lips K, Volk C: Polyspecific organic cation transporters: structure, function, physiological roles, and biopharmaceutical implications. Pharm Res. 2007, 24: 1227-1251. 10.1007/s11095-007-9254-z.
Urban TJ, Giacomini KM: Organic cation transporters. Drug Transporters: Molecular Characterization and Role in Drug Disposition. Edited by: You G, Morris ME. 2010, Hoboken, NJ: John Wiley & Sons, 11-33.
Nies AT, Koepsell H, Damme K, Schwab M: Organic cation transporters (OCTs, MATEs), in vitro and in vivo evidence for the importance in drug therapy. Handb Exp Pharmacol. 2011, 201: 105-167. 10.1007/978-3-642-14541-4_3.
Jonker JW, Schinkel AH: Pharmacological and physiological functions of the polyspecific organic cation transporters: OCT1, 2, and 3 (SLC22A1-3). J Pharmacol Exp Ther. 2004, 308: 2-9.
Hilgendorf C, Ahlin G, Seithel A, Artursson P, Ungell AL, Karlsson J: Expression of thirty-six drug transporter genes in human intestine, liver, kidney, and organotypic cell lines. Drug Metab Dispos. 2007, 35: 1333-1340. 10.1124/dmd.107.014902.
Okabe H, Satoh S, Kato T, Kitahara O, Yanagawa R, Yamaoka Y, Tsunoda T, Furukawa Y, Nakamura Y: Genome-wide analysis of gene expression in human hepatocellular carcinomas using cDNA microarray: identification of genes involved in viral carcinogenesis and tumor progression. Cancer Res. 2001, 61: 2129-2137.
Chang Q, Chen J, Beezhold KJ, Castranova V, Shi X, Chen F: JNK1 activation predicts the prognostic outcome of the human hepatocellular carcinoma. Mol Cancer. 2009, 8: 64-
Park T, Yi SG, Shin YK, Lee S: Combining multiple microarrays in the presence of controlling variables. Bioinformatics. 2006, 22: 1682-1689. 10.1093/bioinformatics/btl183.
Esteller M: Epigenetics in cancer. N Engl J Med. 2008, 358: 1148-1159. 10.1056/NEJMra072067.
Melotte V, Lentjes MH, van den Bosch SM, Hellebrekers DM, de Hoon JP, Wouters KA, Daenen KL, Partouns-Hendriks IE, Stessels F, Louwagie J, Smits KM, Weijenberg MP, Sanduleanu S, Khalid-de Bakker CA, Oort FA, Meijer GA, Jonkers DM, Herman JG, de Bruine AP, van EM: N-Myc downstream-regulated gene 4 (NDRG4): a candidate tumor suppressor gene and potential biomarker for colorectal cancer. J Natl Cancer Inst. 2009, 101: 916-927. 10.1093/jnci/djp131.
Wong IH, Lo YM, Zhang J, Liew CT, Ng MH, Wong N, Lai PB, Lau WY, Hjelm NM, Johnson PJ: Detection of aberrant p16 methylation in the plasma and serum of liver cancer patients. Cancer Res. 1999, 59: 71-73.
Brock MV, Hooker CM, Ota-Machida E, Han Y, Guo M, Ames S, Glockner S, Piantadosi S, Gabrielson E, Pridham G, Pelosky K, Belinsky SA, Yang SC, Baylin SB, Herman JG: DNA methylation markers and early recurrence in stage I lung cancer. N Engl J Med. 2008, 358: 1118-1128. 10.1056/NEJMoa0706550.
Aoki M, Terada T, Kajiwara M, Ogasawara K, Ikai I, Ogawa O, Katsura T, Inui K: Kidney-specific expression of human organic cation transporter 2 (OCT2/SLC22A2) is regulated by DNA methylation. Am J Physiol Renal Physiol. 2008, 295: F165-F170. 10.1152/ajprenal.90257.2008.
Amann T, Maegdefrau U, Hartmann A, Agaimy A, Marienhagen J, Weiss TS, Stoeltzing O, Warnecke C, Scholmerich J, Oefner PJ, Kreutz M, Bosserhoff AK, Hellerbrand C: GLUT1 expression is increased in hepatocellular carcinoma and promotes tumorigenesis. Am J Pathol. 2009, 174: 1544-1552. 10.2353/ajpath.2009.080596.
Hellerbrand C, Amann T, Schlegel J, Wild P, Bataille F, Spruss T, Hartmann A, Bosserhoff AK: The novel gene MIA2 acts as a tumour suppressor in hepatocellular carcinoma. Gut. 2008, 57: 243-251. 10.1136/gut.2007.129544.
Kirovski G, Stevens AP, Czech B, Dettmer K, Weiss TS, Wild P, Hartmann A, Bosserhoff AK, Oefner PJ, Hellerbrand C: Down-regulation of methylthioadenosine phosphorylase (MTAP) induces progression of hepatocellular carcinoma via accumulation of 5'-deoxy-5'-methylthioadenosine (MTA). Am J Pathol. 2011, 178: 1145-1152. 10.1016/j.ajpath.2010.11.059.
Nies AT, Koepsell H, Winter S, Burk O, Klein K, Kerb R, Zanger UM, Keppler D, Schwab M, Schaeffeler E: Expression of organic cation transporters OCT1 (SLC22A1) and OCT3 (SLC22A3) is affected by genetic factors and cholestasis in human liver. Hepatology. 2009, 50: 1227-1240. 10.1002/hep.23103.
Kruck S, Bedke J, Hennenlotter J, Ohneseit PA, Kuehs U, Senger E, Sievert KD, Stenzl A: Activation of mTOR in renal cell carcinoma is due to increased phosphorylation rather than protein overexpression. Oncol Rep. 2010, 23: 159-163.
Theodorescu D, Broder SR, Boyd JC, Mills SE, Frierson HF: Cathepsin D and chromogranin A as predictors of long term disease specific survival after radical prostatectomy for localized carcinoma of the prostate. Cancer. 1997, 80: 2109-2119. 10.1002/(SICI)1097-0142(19971201)80:11<2109::AID-CNCR10>3.0.CO;2-2.
Ehrich M, Nelson MR, Stanssens P, Zabeau M, Liloglou T, Xinarianos G, Cantor CR, Field JK, van den BD: Quantitative high-throughput analysis of DNA methylation patterns by base-specific cleavage and mass spectrometry. Proc Natl Acad Sci USA. 2005, 102: 15785-15790. 10.1073/pnas.0507816102.
Coolen MW, Statham AL, Gardiner-Garden M, Clark SJ: Genomic profiling of CpG methylation and allelic specificity using quantitative high-throughput mass spectrometry: critical evaluation and improvements. Nucleic Acids Res. 2007, 35: e119-10.1093/nar/gkm662.
Bock C, Walter J, Paulsen M, Lengauer T: CpG island mapping by epigenome prediction. PLoS Comput Biol. 2007, 3: e110-10.1371/journal.pcbi.0030110.
Lee S, Lee HJ, Kim JH, Lee HS, Jang JJ, Kang GH: Aberrant CpG island hypermethylation along multistep hepatocarcinogenesis. Am J Pathol. 2003, 163: 1371-1378. 10.1016/S0002-9440(10)63495-5.
Gao W, Kondo Y, Shen L, Shimizu Y, Sano T, Yamao K, Natsume A, Goto Y, Ito M, Murakami H, Osada H, Zhang J, Issa JP, Sekido Y: Variable DNA methylation patterns associated with progression of disease in hepatocellular carcinomas. Carcinogenesis. 2008, 29: 1901-1910. 10.1093/carcin/bgn170.
Lambert MP, Paliwal A, Vaissiere T, Chemin I, Zoulim F, Tommasino M, Hainaut P, Sylla B, Scoazec JY, Tost J, Herceg Z: Aberrant DNA methylation distinguishes hepatocellular carcinoma associated with HBV and HCV infection and alcohol intake. J Hepatol. 2011, 54: 705-715. 10.1016/j.jhep.2010.07.027.
Um TH, Kim H, Oh BK, Kim MS, Kim KS, Jung G, Park YN: Aberrant CpG island hypermethylation in dysplastic nodules and early HCC of hepatitis B virus-related human multistep hepatocarcinogenesis. J Hepatol. 2011, 54: 939-947. 10.1016/j.jhep.2010.08.021.
Giacomini KM, Huang SM, Tweedie DJ, Benet LZ, Brouwer KL, Chu X, Dahlin A, Evers R, Fischer V, Hillgren KM, Hoffmaster KA, Ishikawa T, Keppler D, Kim RB, Lee CA, Niemi M, Polli JW, Sugiyama Y, Swaan PW, Ware JA, Wright SH, Yee SW, Zamek-Gliszczynski MJ, Zhang L: Membrane transporters in drug development. Nat Rev Drug Discov. 2010, 9: 215-236. 10.1038/nrd3028.
Lee JS, Chu IS, Heo J, Calvisi DF, Sun Z, Roskams T, Durnez A, Demetris AJ, Thorgeirsson SS: Classification and prediction of survival in hepatocellular carcinoma by gene expression profiling. Hepatology. 2004, 40: 667-676. 10.1002/hep.20375.
Lee JS, Heo J, Libbrecht L, Chu IS, Kaposi-Novak P, Calvisi DF, Mikaelyan A, Roberts LR, Demetris AJ, Sun Z, Nevens F, Roskams T, Thorgeirsson SS: A novel prognostic subtype of human hepatocellular carcinoma derived from hepatic progenitor cells. Nat Med. 2006, 12: 410-416. 10.1038/nm1377.
Yamashita T, Ji J, Budhu A, Forgues M, Yang W, Wang HY, Jia H, Ye Q, Qin LX, Wauthier E, Reid LM, Minato H, Honda M, Kaneko S, Tang ZY, Wang XW: EpCAM-positive hepatocellular carcinoma cells are tumor-initiating cells with stem/progenitor cell features. Gastroenterology. 2009, 136: 1012-1024. 10.1053/j.gastro.2008.12.004.
Hui L, Zatloukal K, Scheuch H, Stepniak E, Wagner EF: Proliferation of human HCC cells and chemically induced mouse liver cancers requires JNK1-dependent p21 downregulation. J Clin Invest. 2008, 118: 3943-3953. 10.1172/JCI37156.
Moribe T, Iizuka N, Miura T, Kimura N, Tamatsukuri S, Ishitsuka H, Hamamoto Y, Sakamoto K, Tamesa T, Oka M: Methylation of multiple genes as molecular markers for diagnosis of a small, well-differentiated hepatocellular carcinoma. Int J Cancer. 2009, 125: 388-397. 10.1002/ijc.24394.
Lou C, Du Z, Yang B, Gao Y, Wang Y, Fang S: Aberrant DNA methylation profile of hepatocellular carcinoma and surgically resected margin. Cancer Sci. 2009, 100: 996-1004. 10.1111/j.1349-7006.2009.01138.x.
Irizarry RA, Ladd-Acosta C, Wen B, Wu Z, Montano C, Onyango P, Cui H, Gabo K, Rongione M, Webster M, Ji H, Potash JB, Sabunciyan S, Feinberg AP: The human colon cancer methylome shows similar hypo- and hypermethylation at conserved tissue-specific CpG island shores. Nat Genet. 2009, 41: 178-186. 10.1038/ng.298.
Eckhardt F, Lewin J, Cortese R, Rakyan VK, Attwood J, Burger M, Burton J, Cox TV, Davies R, Down TA, Haefliger C, Horton R, Howe K, Jackson DK, Kunde J, Koenig C, Liddle J, Niblett D, Otto T, Pettett R, Seemann S, Thompson C, West T, Rogers J, Olek A, Berlin K, Beck S: DNA methylation profiling of human chromosomes 6, 20 and 22. Nat Genet. 2006, 38: 1378-1385. 10.1038/ng1909.
Brenet F, Moh M, Funk P, Feierstein E, Viale AJ, Socci ND, Scandura JM: DNA methylation of the first exon is tightly linked to transcriptional silencing. PLoS One. 2011, 6: e14524-10.1371/journal.pone.0014524.
Shu Y, Sheardown SA, Brown C, Owen RP, Zhang S, Castro RA, Ianculescu AG, Yue L, Lo JC, Burchard EG, Brett CM, Giacomini KM: Effect of genetic variation in the organic cation transporter 1 (OCT1) on metformin action. J Clin Invest. 2007, 117: 1422-1431. 10.1172/JCI30558.
More SS, Li S, Yee SW, Chen L, Xu Z, Jablons DM, Giacomini KM: Organic cation transporters modulate the uptake and cytotoxicity of picoplatin, a third-generation platinum analogue. Mol Cancer Ther. 2010, 9: 1058-1069. 10.1158/1535-7163.MCT-09-1084.
Gupta A, Lutsenko S: Human copper transporters: mechanism, role in human diseases and therapeutic potential. Future Med Chem. 2009, 1: 1125-1142. 10.4155/fmc.09.84.
Kacevska M, Ivanov M, Ingelman-Sundberg M: Perspectives on epigenetics and its relevance to adverse drug reactions. Clin Pharmacol Ther. 2011, 89: 902-907. 10.1038/clpt.2011.21.
Baker EK, El-Osta A: MDR1, chemotherapy and chromatin remodeling. Cancer Biol Ther. 2004, 3: 819-824. 10.4161/cbt.3.9.1101.
Nies AT, Herrmann E, Brom M, Keppler D: Vectorial transport of the plant alkaloid berberine by double-transfected cells expressing the human organic cation transporter 1 (OCT1, SLC22A1) and the efflux pump MDR1 P-glycoprotein (ABCB1). Naunyn Schmiedebergs Arch Pharmacol. 2008, 376: 449-461. 10.1007/s00210-007-0219-x.
MethPrimer. [http://www.urogene.org/methprimer]
Ehrich M, Turner J, Gibbs P, Lipton L, Giovanneti M, Cantor C, van den Boom D: Cytosine methylation profiling of cancer cell lines. Proc Natl Acad Sci USA. 2008, 105: 4844-4849. 10.1073/pnas.0712251105.
The R Project for Statistical Computing. [http://www.r-project.org]
Holm S: A simple sequentially rejective multiple test procedure. Scand J Stat. 1979, 6: 65-70.
Acknowledgements
We gratefully acknowledge Heidi Köhler for preparing figures and tables and Monika Elbl, Silvia Hübner, Andrea Jarmuth, Monika Seiler and Ursula Waldherr for excellent technical assistance. We thank Professor Dietrich Keppler for helpful discussion. We gratefully thank Professor David Zakim, USA, for helpful advice and discussions. This work was supported by the Robert Bosch Foundation (Stuttgart, Germany), the Federal Ministry for Education and Research (BMBF, Berlin, Germany) grants 03 IS 2061C and 0315755 and the FP7-grant PITN-GA-2009-238132 and the Wilhelm-Sander-Stiftung (München, Germany; no.2010.059.1).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
MS was responsible for study design. ES was responsible for study design and collected, analyzed and interpreted data. Data were collected, analyzed and interpreted by ATN, CH and SK. Statistical analysis was performed by SW. All other authors contributed samples for the study, and/or contributed to drafting the manuscript. All authors have read and approved the manuscript for publication.
Electronic supplementary material
13073_2011_311_MOESM1_ESM.PDF
Additional file 1: Additional methods, tables and figures. Methods: additional description of methods. Immunofluorescence microscopy of tissue samples was performed as previously described [20, 45]. Primers used for DNA methylation analysis were designed with Methprimer software [46]. DNA methylation of RASSF1A was analyzed as described previously [47]. Statistical analyses were performed using the statistical software R, version 2.13.0 [48]. Where indicated, P-values were adjusted according to Holm's multiple testing correction procedure [49]. Table S1: overview of different sample sets used in the present study. Table S2: clinicopathological characteristics of 71 patients with HCC. Table S3: clinicopathological characteristics of HCC study population. Table S4: characteristics of histologically normal liver tissues (IKP-liverbank). Table S5: DNA methylation analysis of OCT1/SLC22A1, OCT2/SLC22A2 and OCT3/SLC22A3. Figure S1: differences in the SLC22A1 immunohistochemical score between the corresponding adjacent non-tumor tissue and HCC tissue, regarding the underlying etiology for HCC. Figure S2: Ki-67/MIB1 data for HCC tumor samples. Figure S3: association between SLC22A1 protein expression and tumor grade or tumor stage. Figure S4: DNA methylation profiles of SLC22A2 and RASSF1A in HCC and adjacent non-tumor tissue. Figure S5: association between SLC22A1 mRNA expression or DNA methylation and tumor stage or histological tumor grade. Figure S6: DNA methylation profiles of SLC22A1, SLC22A2, and SLC22A3 in non-tumor (normal) liver tissues, as well as in non-tumor liver tissue derived from patients with HCC. (PDF 397 KB)
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article
Cite this article
Schaeffeler, E., Hellerbrand, C., Nies, A.T. et al. DNA methylation is associated with downregulation of the organic cation transporter OCT1 (SLC22A1) in human hepatocellular carcinoma. Genome Med 3, 82 (2011). https://doi.org/10.1186/gm298
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1186/gm298