
Abstract
Background
Intestinal mucus production by hyperplasic goblet cells is a striking pathological feature of many parasitic helminth infections and is related to intestinal protection and worm expulsion. Induction of goblet cell hyperplasia is associated with TH2 immune responses, which in helminth infections are controlled primarily by IL-13, and also IL-4. In the study presented here we examine the goblet cell hyperplasic response to three experimental parasitic helminth infections; namely Nippostrongylus brasiliensis, Syphacia obvelata and Schistosoma mansoni.
Results
As expected N. brasiliensis infection induced a strong goblet cell hyperplasia dependent on IL-4/IL-13/IL-4Rα expression. In contrast, and despite previously published transiently elevated IL-4/IL-13 levels, S. obvelata infections did not increase goblet cell hyperplasia in the colon. Furthermore, induction of goblet cell hyperplasia in response to S. mansoni eggs traversing the intestine was equivalent between BALB/c, IL-4/IL-13-/- and IL-4Rα-/- mice.
Conclusion
Together these data demonstrate that intestinal goblet cell hyperplasia can be independent of TH2 immune responses associated with parasitic helminth infections.
Similar content being viewed by others
Background
Interleukin (IL)-4 and IL-13 are related cytokines and the dominant mediators of TH2 immune responses [1–3]. Signalling by both cytokines is dependent on binding to heterodimeric receptors containing the IL-4 receptor α chain (IL-4Rα). Ligand binding results in intracellular signalling pathways activating the TH2 defining transcription factors STAT-6 and/or GATA-3 [4, 5]. This polarisation to a TH2 immune response is essential for the successful resolution of a number of helminth infections [6–10].
Actual worm expulsion, in nematode infections, is associated with increased IL-13/IL-4Rα/STAT-6 dependent intestinal smooth muscle contractions, epithelial cell turnover and goblet cell hyperplasia [11–13]. Infections of IL-4-/-, IL-13-/-, IL-4Rα-/- and Stat 6-/- mice with the nematodes Trichuris muris, Heligmosomoides polygyrus and Nippostrongylus brasiliensis have demonstrated a positive relationship between polarisation to a TH2 immune response, goblet cell hyperplasia and worm expulsion [14–19]. In support of a role for goblet cell derived mucus in worm expulsion in vitro experiments have demonstrated increased viscosity of mileu surrounding N. brasiliensis at an equivalent density to intestinal mucus inhibits worm movement [20]. Moreover, isolation of the goblet cell secreted protein RELMβ/FIZZ2 and incubation with parasitic nematodes in vitro results in impaired chemotactic function in the worm [21]. These observations have led to TH2 induced goblet cell hyperplasia being considered a key mechanistic factor in resolving gastrointestinal related nematode infections [22–24]
Intestinal goblet cell hyperplasia in Schistosoma mansoni (S. mansoni) infections is driven by parasite eggs traversing the intestine [8, 25], as opposed to nematode infections where adult worms residing in the intestine induce the goblet cell responses [8, 9, 26]. S. mansoni eggs produced by adults residing in the mesenteric venules move from the blood vessels through the intestine passing to the lumen. This movement of eggs generates considerable tissue damage as well as inducing a strong mucosal response in the intestine [8, 27]. As with nematode infections, S. mansoni induced mucus production has been considered to be TH2 dependant [22, 28–30].
In this study we examined goblet cell hyperplasia in response to infection with the nematodes N. brasiliensis and Syphacia obvelata and the trematode S. mansoni. As already published N. brasiliensis infection induced a goblet cell hyperplasic response dependent on IL-4/IL-13/IL-4Rα expression [9]. However, infection with the nematode S. obvelata did not increase goblet cell hyperplasia in the host colon, irrespective of IL-4Rα expression. Such data demonstrates that IL-4Rα driven goblet cell hyperplasia may not be essential for the clearance of all gastro-intestinal nematode infections. Furthermore, we also show S. mansoni induced goblet cell hyperplasia to be independent of IL-4/IL-13 responsiveness. This data represents the first demonstration of goblet cell hyperplasia and mucus production in response to helminth infections being independent of IL-4/IL-13.
Results
N. brasiliensis infection induces IL-4/IL-13 dependent goblet cell hyperplasia while S. obvelata infection does not induce goblet cell hyperplasia
Examination of IL-4/IL-13 dependent goblet cell hyperplasic responses in the intestinal niches utilised by the nematodes N. brasiliensis and S. obvelata infections was carried out in BALB/c, IL-4/IL-13-/-, IL-4-/- and IL-4Rα-/- mice.
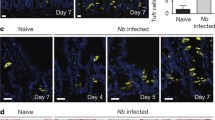
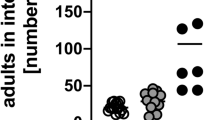
N. brasiliensis infected BALB/c mice demonstrated significantly higher levels of goblet cell hyperplasia in the small intestine at both days 7 and 10 post infection (PI) when compared to naïve mice (Figure 1A). However, no significant increase in the number of goblet cells in the intestine could be detected in infected and naïve IL-4/IL-13-/- or IL4Rα-/- mice when compared to naïve controls (Figure 1A). Examination of intestinal worm burdens in BALB/c mice showed resolution of infection by day 10 PI. Both IL-4/IL-13-/- and IL-4Rα-/- mice failed to expel adult worms by day 10 PI (Figure 1B). These data confirm N. brasiliensis clearance to be associated with an IL-4/13/IL-4Rα dependent goblet cell hyperplasia.

Goblet cell hyperplasia in N. brasiliensis infection. A) Goblet cell quantification naïve mice and N. brasiliensis infected BALB/c (solid bar), IL-4Rα-/- (open bar) and IL-4/IL-13-/- (gray bar) mice 7 days and 10 days PI in small intestine sections. B) Intestinal adult worm burden 7 and 10 days post infection. Data representative of two experiments are shown. Data are means of four mice per group ± SEM. * P < 0.05; *** P < 0.001 (significantly different from naïve mice).
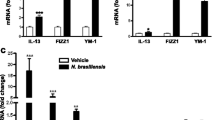
S. obvelata adult worm burdens in infected wild type mice are only detectable by day 28 PI (approx. 1.25 worms/caecum), this burden peaks by day 35 PI to approximately 21 worms/caecum. Infection of IL-4Rα-/- mice results in considerably higher worm burdens (approx. 251 worms/caecum at day 28 PI and 400 worms/caecum at day 35 PI) when compared to wild type mice [10]. In contrast to N. brasiliensis infected mice, no induction of goblet cell hyperplasia in the colon of S. obvelata infected BALB/c, IL-4Rα-/- and IL-4-/- mice was seen (Figure 2A and 2B). This lack of a goblet cell response was irrespective of heightened levels of the TH2 cytokine IL-4 at day 7 PI (p < 0.01) in BALB/c restimulated splenocytes isolated from infected mice (Figure 2C) BALB/c IL-4 levels declined to that found in naïve mice at day 14 PI. IL-4Rα-/- mice failed to demonstrate any significant increase in IL-4 production when compared to naïve mice.

Goblet cell hyperplasia in S. obvelata infection. (A) Photomicrograph of colon from BALB/c, IL-4Rα-/- and IL-4-/-. Representative pictures of colon sections are shown from both naïve (i) and pinworm infected mice at 7 (ii) and 14 days PI (iii). Sections were stained with PAS to identify goblet cells. 100× magnification. (B) Quantification of goblet cells per crypt in colon 7 and 14 days PI in BALB/c (solid bar), IL-4Rα-/- (open bar) and IL-4/IL-13-/- (gray bar). Data representative of two experiments are shown. Data are means of four mice per group ± SEM. (C) S. obvelata dependent IL-4 secretion from anti-CD3 restimulated splenocytes. BALB/c (solid bar), IL-4Rα-/- (open bar). **, P < 0.01 (significantly different from naive mice). Data representative of two experiments showing means for four mice/group ± SD.
Together these data demonstrate that S. obvelata infections do not induce a colonic mucus response even though levels of IL-4 and other TH2 cytokines are significantly increased [10]
Schistosoma mansoni induces goblet cell hyperplasia in the intestine in an IL-4/IL-13 independent manner
S. mansoni infection induces a strong TH2 immune response and goblet cell hyperplasia related to parasite egg production [25, 31] In order to confirm the role of parasite eggs in induction of goblet cell hyperplasia, we analysed the hyperplasic response at 5 weeks PI (before the peak of egg production) and at the peak of parasite egg production; 8 weeks PI. While no difference in the number of goblet cells could be detected in the intestine of naïve or infected BALB/c at 5 weeks PI (data not shown), a strong induction of goblet cell hyperplasia was detected at 8 weeks PI (Figure 3B and 3C). To establish whether this hyperplasic response was dependent on IL-4/IL-13/IL-4Rα responsiveness we examined the intestines of infected IL-4/IL-13-/- and IL-4Rα-/- mice at 8 weeks PI. Here we found no difference in the numbers of eggs accumulating in the small intestine and large intestine between BALB/c, IL-4/IL-13-/- and IL-4Rα-/- mice (Figure 3A and 4A). Goblet cell hyperplasia in the small intestine of all infected mouse groups was significantly elevated above naïve controls (Figures 3B, C(i) and 3C(ii)). Furthermore, equivalent levels of goblet cell hyperplasia were found in the intestine of all infected mice groups (Figures 3B and 3C). To demonstrate if IL-4/IL-13/IL-4Rα independent goblet cell hyperplasia occurred throughout the intestine we also examined the colon of both naïve and infected mice. As with the small intestine goblet cell hyperplasia was elevated above naïve controls in all mouse groups and no differences were found between infected groups (Figure 4B and 4C). Together these results demonstrate IL-4/IL-13/IL-4Rα independent goblet cell hyperplasia in the intestine of mice infected with S. mansoni.

Goblet cell hyperplasia in ileum during S. mansoni infection. (A) S. mansoni egg content in the ileum of BALB/c (solid bar), IL-4Rα-/- (open bar) and IL-4/IL-13-/- (gray bar) mice at 8 weeks PI. Data are pooled from 2 to 4 individual experiments. Data are means of these experiments ± SEM. (B) Quantification of goblet cells per villus in ileum of naïve mice and 8 weeks PI from BALB/c (black bars), IL-4Rα-/- (open bar) and IL-4/IL-13-/- (gray bar). Data representative of three experiments are shown. * P < 0.05 (significantly different from naïve mice). (C) Photomicrograph of ileum from BALB/c, IL-4Rα-/- and IL-4/IL-13-/-. Representative pictures of ileum sections are shown from both naïve (i) and S. mansoni infected mice at 8 weeks PI (ii). Sections were stained with PAS to identify goblet cells. 100× magnification.

Goblet cell hyperplasia in the colon during S. mansoni infection. (A) Tissue egg content of the colon from S. mansoni infected BALB/c (solid bar), IL-4Rα-/- (open bar) and IL-4/IL-13-/- (gray bar) mice at 8 weeks PI. Data are pooled from 2 to 4 individual experiments. Data are means of these experiments ± SEM. (B) Quantification of goblet cell number per villus in colon of naïve mice and 8 weeks PI from BALB/c (solid bar), IL-4Rα-/- (open bar) and IL-4/IL-13-/- (gray bar) mice. Data representative of three experiments are shown. * P < 0.05 (significantly different from naïve mice). (C) Photomicrograph of colon from BALB/c, IL-4Rα-/- and IL-4/IL-13-/-. Representative pictures of colon sections are shown from both naïve (i) and S. mansoni infected mice at 8 weeks PI (ii). Sections were stained with PAS to identify goblet cells. 100× magnification
Discussion
Our data demonstrates that (i) goblet cell hyperplasia is dependent on helminth species and (ii) IL-4/IL-13 responsiveness is not required for induction of S. mansoni egg induced goblet cell hyperplasia.
It has previously been demonstrated that N. brasiliensis [9] and S. obvelata [10] infected IL-4-/-, IL-13-/- and IL-4Rα-/- mice have impaired worm expulsion, while in S. mansoni infections IL-4/IL-13 signalling is essential for host survival [8]. A common feature of both N. brasiliensis and S. mansoni infections is the hosts' goblet cell hyperplasic response to the parasite. Such responses have previously been considered to be dependent, in part at least, on the hosts TH2 polarised immune response [9, 32]. From the data presented here and in other studies this does indeed appear to be the case in N. brasiliensis infections[6, 8, 9]. Work on other parasitic nematode models such a T. muris also show a TH2 dependent worm expulsion and goblet cell response [16]. However, in this study we have demonstrated that this may not be the case for all intestinal nematode infections.
Following oral infection with S. obvelata eggs, larvae emerge in the hosts small intestine at 7 day PI [33]. From here the larvae migrate, mature and establish the definitive infection in the hosts cecum and colon. We found the hosts TH2 immune response to peak at day 7 PI and then decreases from at least day 14 PI. Previous work has shown that by day 35 PI this response is undetectable [10]. Together, these data demonstrate a transient TH2 response to this infection. TH2 responses in other intestinal nematode infections result in strong goblet cell hyperplasic responses [14, 17–19]. However, mice infected with S. obvelata failed to generate hyperplasic goblet cell responses, suggesting that TH2 induction of intestinal mucus responses is not a common feature of intestinal nematode infections, or that the TH2 response needs to be sustained. Other factors such as prostaglandins [34], cholinergic [13] and non-cholinergic [13] agonist may also play a role. Additionally, the different niches occupied by various species of parasitic nematodes could effect the host response to them [35]. S. obvelata infections do not cause major pathology in the intestine [36] as opposed to N. brasiliensis and T. muris which cause considerable histological damage to the hosts intestinal architecture [9, 22]. Such differences in worm pathogenicity may explain the lack of a goblet cell response in S. obvelata infections, irrespective of the hosts TH2 polarisation [10].
S. mansoni infection induces a strong TH2 response initiated by worm egg production at week 4 PI and persists throughout the infection [37]. Associated with this are significant levels of goblet cell hyperplasia in the intestine [25, 31]. S. mansoni egg antigens have previously been shown to also induce goblet cell hyperplasia in the lung in a IL-4Rα dependent manner [32]. However the role of IL-4Rα in goblet cell hyperplasia in the intestine during the live infection has not been shown. An explanation for the IL-4Rα independent hyperplasia described here could be the mode of S. mansoni infection and its interaction with the hosts' tissue. S. mansoni eggs cause pathology from the adventitial surface of the intestine, as opposed to nematodes driving the pathology from the lumen. We propose that the severe tissue damage resulting from the eggs migration from the adventitial surface to the lumen is capable of initiating a goblet cell response, independently of IL-4 and IL-13 signalling during S. mansoni infection.
In addition to IL-4/IL-13 other cytokines may act to induce goblet cells hyperplasia. IL-9 and IL-5 have previously been shown to play a role in directly inducing IL-4/IL-13 independent goblet cell hyperplasia in lung models [38, 39]. IL-9 overexpressing transgenic mice infected with S. mansoni do have increased goblet cell hyperplasia [40]. However IL-9 transgenic mice also had increased IL-4 and IL-13 compared to wild type mice, and therefore it cannot be concluded that IL-9 directly increases goblet cell hyperplasia. Furthermore IL-9 levels are decreased in N. brasiliensis infected IL-4Rα-/- mice [9]. As such IL-4/IL-13 independent intestinal goblet cell hyperplasia may not be due to increased IL-9. No clear reports linking IL-5 to goblet cell hyperplasia during S. mansoni infection have reported. As IL-4Rα-/- mice have decreased IL-5 expression it is also unlikely that IL-5 induces intestinal goblet cell hyperplasia in S. mansoni infections [8].
Conclusion
Our results demonstrate for the first time that intestinal goblet cell hyperplasia in response to parasitic helminth infections can occur independently of IL-4/IL-13 signalling and that intestinal nematode infections may not always induce a goblet cell response.
Methods
Mouse strains
IL-4-/- [41] IL-4/13-/-[2] and IL-4Rα-/- [3] mice were generated on a BALB/c background. BALB/c mice were used as controls in all experiments. All mice were age and sex matched. Mice were kept in the Health Science Faculty animal unit of the University of Cape Town (UCT), in individually ventilated cages under specific-pathogen-free (SPF) conditions. All experiments were performed in accordance with guidelines laid down by the Animal Ethics Research Board of UCT (Cape Town, South Africa).
Parasites and infection
Syphacia obvelata
Infection and recovery of S. obvelata were performed as previously described [33]. Briefly, eggs of S. obvelata used for infection were collected from the caeca of naturally infected mice (IL-4/13-/-, and IL-4Rα-/-) maintained in barrier facilities. The caeca were collected in 0.65% NaCl, cut open, and submerged in a gauze mesh at the mouth of a conical flask for 1 to 2 h at 37°C to allow the worms to migrate out. Worm burdens were assessed on various days post infection. After being washed in 0.65% NaCl, worms were crushed and their eggs were isolated by passage through 70 μm nylon cell strainers (BD Falcon, BD Biosciences, Belgium). Each mouse was inoculated orally with 500 eggs using oral dosing cannulae (VetTech, Cheshire, United Kingdom).
Nippostrongylus brasiliensis
N. brasiliensis nematodes were kindly provided by Klaus Erb, (Wurzberg, Germany). Mice were subcutaneously injected with 750 L3 larvae of N. brasiliensis. Analysis of numbers of adult worm numbers in the intestine was determined as previously described [9].
Schistosoma mansoni
Naïve sex-matched mice from 6 to 10 weeks of age were percutaneously infected with 70 to 80 live cercariae of a Puerto Rican strain of S. mansoni obtained from infected Biomphalaria glabrata snails. Eight weeks post infection the intestine was surgically removed. Ileum and colon were removed 2 cm proximal and 0.5 cm distal to the caecum, respectively [42]. Approximately 2 cm of tissue was weighed and digested in 5 ml of 5% potassium hydroxide overnight at 37°C. The digests were vortexed and centrifuged at 100 g for 5 min to pellet eggs. The supernatant was aspirated until 1–2 mls remained. The eggs were vortexed and counted in 50 μl in triplicate. The counts were presented as eggs per gram of tissue as previously described [43, 44]
Histology
Tissue samples were fixed in a neutral buffered formalin solution. Following embedding in paraffin, samples were cut into 5–7 μm sections. Sections were stained with periodic acid Schiff reagent (PAS). The number of positively stained cells per five villi or crypts was counted by light microscopy for small intestine or colon, respectively. All samples were randomized and counted in a blinded manner. Photomicrographs were captured using a Nikon 5.0 Mega Pixels Color Digital Camera (Digital SIGHT DS-SMc).
Splenocyte restimulation and IL-4 cytokine ELISA
Single cell splenocyte suspensions were prepared from spleens removed from infected (days 7 and 14 PI) and uninfected mice. 1 × 106 splenocytes per ml were cultured in IMDM (Gibco) media supplemented with 10% fetal calf serum (Gibco) for 72 h at 37°C in 96 well plates pre-coated with either PBS or 20 mg/ml anti-CD3 (clone 145-2C11). Cells were then centrifuged at 1200 rpm for 5 min and the supernatants collected. Supernatent IL-4 concentrations were then determined by ELISA as described previously [3].
Statistics
Data are presented as means ± standard error of the mean (SEM), and the significant differences were determined using Student's t test (Prism software [45]).
Abbreviations
- IL:
-
interleukin
- KO:
-
knockout
- PAS:
-
periodic acid Schiff
- PI:
-
post infection
- WT:
-
wild type.
References
McKenzie GJ, Emson CL, Bell SE, Anderson S, Fallon P, Zurawski G, Murray R, Grencis R, McKenzie AN: Impaired development of Th2 cells in IL-13-deficient mice. Immunity. 1998, 9 (3): 423-432. 10.1016/S1074-7613(00)80625-1.
McKenzie GJ, Fallon PG, Emson CL, Grencis RK, McKenzie AN: Simultaneous disruption of interleukin (IL)-4 and IL-13 defines individual roles in T helper cell type 2-mediated responses. J Exp Med. 1999, 189 (10): 1565-1572. 10.1084/jem.189.10.1565.
Mohrs M, Ledermann B, Kohler G, Dorfmuller A, Gessner A, Brombacher F: Differences between IL-4- and IL-4 receptor alpha-deficient mice in chronic leishmaniasis reveal a protective role for IL-13 receptor signaling. J Immunol. 1999, 162 (12): 7302-7308.
Andrews RP, Ericksen MB, Cunningham CM, Daines MO, Hershey GK: Analysis of the life cycle of stat6. Continuous cycling of STAT6 is required for IL-4 signaling. J Biol Chem. 2002, 277 (39): 36563-36569. 10.1074/jbc.M200986200.
Metwali A, Blum A, Elliott DE, Weinstock JV: Interleukin-4 receptor alpha chain and STAT6 signaling inhibit gamma interferon but not Th2 cytokine expression within schistosome granulomas. Infect Immun. 2002, 70 (10): 5651-5658. 10.1128/IAI.70.10.5651-5658.2002.
Barner M, Mohrs M, Brombacher F, Kopf M: Differences between IL-4R alpha-deficient and IL-4-deficient mice reveal a role for IL-13 in the regulation of Th2 responses. Curr Biol. 1998, 8 (11): 669-672. 10.1016/S0960-9822(98)70256-8.
Dehlawi MS, Mahida YR, Hughes K, Wakelin D: Effects of Trichinella spiralis infection on intestinal pathology in mice lacking interleukin-4 (IL-4) or intestinal trefoil factor (ITF/TFF3). Parasitol Int. 2006, 55 (3): 207-211. 10.1016/j.parint.2006.05.002.
Herbert DR, Holscher C, Mohrs M, Arendse B, Schwegmann A, Radwanska M, Leeto M, Kirsch R, Hall P, Mossmann H, Claussen B, Forster I, Brombacher F: Alternative macrophage activation is essential for survival during schistosomiasis and downmodulates T helper 1 responses and immunopathology. Immunity. 2004, 20 (5): 623-635. 10.1016/S1074-7613(04)00107-4.
Horsnell WG, Cutler AJ, Hoving CJ, Mearns H, Myburgh E, Arendse B, Finkelman FD, Owens GK, Erle D, Brombacher F: Delayed Goblet Cell Hyperplasia, Acetylcholine Receptor Expression, and Worm Expulsion in SMC-Specific IL-4Ralpha-Deficient Mice. PLoS Pathog. 2007, 3 (1): e1-10.1371/journal.ppat.0030001.
Michels C, Goyal P, Nieuwenhuizen N, Brombacher F: Infection with Syphacia obvelata (pinworm) induces protective Th2 immune responses and influences ovalbumin-induced allergic reactions. Infect Immun. 2006, 74 (10): 5926-5932. 10.1128/IAI.00207-06.
Akiho H, Blennerhassett P, Deng Y, Collins SM: Role of IL-4, IL-13, and STAT6 in inflammation-induced hypercontractility of murine smooth muscle cells. Am J Physiol Gastrointest Liver Physiol. 2002, 282 (2): G226-32.
Cliffe LJ, Humphreys NE, Lane TE, Potten CS, Booth C, Grencis RK: Accelerated intestinal epithelial cell turnover: a new mechanism of parasite expulsion. Science. 2005, 308 (5727): 1463-1465. 10.1126/science.1108661.
Zhao A, McDermott J, Urban JF, Gause W, Madden KB, Yeung KA, Morris SC, Finkelman FD, Shea-Donohue T: Dependence of IL-4, IL-13, and nematode-induced alterations in murine small intestinal smooth muscle contractility on Stat6 and enteric nerves. J Immunol. 2003, 171 (2): 948-954.
Else KJ, Finkelman FD, Maliszewski CR, Grencis RK: Cytokine-mediated regulation of chronic intestinal helminth infection. J Exp Med. 1994, 179 (1): 347-351. 10.1084/jem.179.1.347.
Finkelman FD, Shea-Donohue T, Goldhill J, Sullivan CA, Morris SC, Madden KB, Gause WC, Urban JF: Cytokine regulation of host defense against parasitic gastrointestinal nematodes: lessons from studies with rodent models. Annu Rev Immunol. 1997, 15: 505-533. 10.1146/annurev.immunol.15.1.505.
Schopf LR, Hoffmann KF, Cheever AW, Urban JF, Wynn TA: IL-10 is critical for host resistance and survival during gastrointestinal helminth infection. J Immunol. 2002, 168 (5): 2383-2392.
Shea-Donohue T, Sullivan C, Finkelman FD, Madden KB, Morris SC, Goldhill J, Pineiro-Carrero V, Urban JF: The role of IL-4 in Heligmosomoides polygyrus-induced alterations in murine intestinal epithelial cell function. J Immunol. 2001, 167 (4): 2234-2239.
Urban JF, Schopf L, Morris SC, Orekhova T, Madden KB, Betts CJ, Gamble HR, Byrd C, Donaldson D, Else K, Finkelman FD: Stat6 signaling promotes protective immunity against Trichinella spiralis through a mast cell- and T cell-dependent mechanism. J Immunol. 2000, 164 (4): 2046-2052.
Webb RA, Hoque T, Dimas S: Expulsion of the gastrointestinal cestode, Hymenolepis diminuta by tolerant rats: evidence for mediation by a Th2 type immune enhanced goblet cell hyperplasia, increased mucin production and secretion. Parasite Immunol. 2007, 29 (1): 11-21. 10.1111/j.1365-3024.2006.00908.x.
Lee DL, Biggs WD: Two- and three-dimensional locomotion of the nematode Nippostrongylus brasiliensis. Parasitology. 1990, 101 Pt 2: 301-308.
Artis D, Wang ML, Keilbaugh SA, He W, Brenes M, Swain GP, Knight PA, Donaldson DD, Lazar MA, Miller HR, Schad GA, Scott P, Wu GD: RELMbeta/FIZZ2 is a goblet cell-specific immune-effector molecule in the gastrointestinal tract. Proc Natl Acad Sci U S A. 2004, 101 (37): 13596-13600. 10.1073/pnas.0404034101.
Onah DN, Nawa Y: Mucosal immunity against parasitic gastrointestinal nematodes. Korean J Parasitol. 2000, 38 (4): 209-236.
Allen A, Leonard AJ, Sellers LA: The mucus barrier. Its role in gastroduodenal mucosal protection. J Clin Gastroenterol. 1988, 10 Suppl 1: S93-8.
Miller HR: Gastrointestinal mucus, a medium for survival and for elimination of parasitic nematodes and protozoa. Parasitology. 1987, 94 Suppl: S77-100.
Fallon PG, Richardson EJ, McKenzie GJ, McKenzie AN: Schistosome infection of transgenic mice defines distinct and contrasting pathogenic roles for IL-4 and IL-13: IL-13 is a profibrotic agent. J Immunol. 2000, 164 (5): 2585-2591.
McKenzie GJ, Bancroft A, Grencis RK, McKenzie AN: A distinct role for interleukin-13 in Th2-cell-mediated immune responses. Curr Biol. 1998, 8 (6): 339-342. 10.1016/S0960-9822(98)70134-4.
Weinstock JV, Boros DL: Heterogeneity of the granulomatous response in the liver, colon, ileum, and ileal Peyer's patches to schistosome eggs in murine schistosomiasis mansoni. J Immunol. 1981, 127 (5): 1906-1909.
Yamauchi J, Kawai Y, Yamada M, Uchikawa R, Tegoshi T, Arizono N: Altered expression of goblet cell- and mucin glycosylation-related genes in the intestinal epithelium during infection with the nematode Nippostrongylus brasiliensis in rat. Apmis. 2006, 114 (4): 270-278. 10.1111/j.1600-0463.2006.apm_353.x.
Ishikawa N, Horii Y, Nawa Y: Immune-mediated alteration of the terminal sugars of goblet cell mucins in the small intestine of Nippostrongylus brasiliensis-infected rats. Immunology. 1993, 78 (2): 303-307.
Ishikawa N, Horii Y, Oinuma T, Suganuma T, Nawa Y: Goblet cell mucins as the selective barrier for the intestinal helminths: T-cell-independent alteration of goblet cell mucins by immunologically 'damaged' Nippostrongylus brasiliensis worms and its significance on the challenge infection with homologous and heterologous parasites. Immunology. 1994, 81 (3): 480-486.
Grzych JM, Pearce E, Cheever A, Caulada ZA, Caspar P, Heiny S, Lewis F, Sher A: Egg deposition is the major stimulus for the production of Th2 cytokines in murine schistosomiasis mansoni. J Immunol. 1991, 146 (4): 1322-1327.
Leeto M, Herbert DR, Marillier R, Schwegmann A, Fick L, Brombacher F: TH1-dominant granulomatous pathology does not inhibit fibrosis or cause lethality during murine schistosomiasis. Am J Pathol. 2006, 169 (5): 1701-1712. 10.2353/ajpath.2006.060346.
Stahl W: Studies on the life cycle of Syphacia muris, the rat pinworm. Keio J Med. 1963, 12: 55-60.
Madden KB, Whitman L, Sullivan C, Gause WC, Urban JF, Katona IM, Finkelman FD, Shea-Donohue T: Role of STAT6 and mast cells in IL-4- and IL-13-induced alterations in murine intestinal epithelial cell function. J Immunol. 2002, 169 (8): 4417-4422.
Grencis R: Resistance to intestinal nematode infection is mediated through IL-13. Interleukin-13. Edited by: Brombacher F. 2003, Georgetown , Eurekah, 51-53.
Michels CE: Investigating the immunological response elicited to the gastrointestinal nematode pinworm (Syphacia obvelata). Immunology Infectious Disease. 2006, Cape Town , University of Cape Town, 33-44.
Pearce EJ, MacDonald AS: The immunobiology of schistosomiasis. Nature reviews. 2002, 2 (7): 499-511. 10.1038/nri843.
Nath P, Yee Leung S, Williams AS, Noble A, Xie S, McKenzie AN, Chung KF: Complete inhibition of allergic airway inflammation and remodelling in quadruple IL-4/5/9/13-/- mice. Clin Exp Allergy. 2007, 37 (10): 1427-1435.
Longphre M, Li D, Gallup M, Drori E, Ordonez CL, Redman T, Wenzel S, Bice DE, Fahy JV, Basbaum C: Allergen-induced IL-9 directly stimulates mucin transcription in respiratory epithelial cells. J Clin Invest. 1999, 104 (10): 1375-1382. 10.1172/JCI6097.
Fallon PG, Smith P, Richardson EJ, Jones FJ, Faulkner HC, Van Snick J, Renauld JC, Grencis RK, Dunne DW: Expression of interleukin-9 leads to Th2 cytokine-dominated responses and fatal enteropathy in mice with chronic Schistosoma mansoni infections. Infect Immun. 2000, 68 (10): 6005-6011. 10.1128/IAI.68.10.6005-6011.2000.
Noben-Trauth N, Kohler G, Burki K, Ledermann B: Efficient targeting of the IL-4 gene in a BALB/c embryonic stem cell line. Transgenic Res. 1996, 5 (6): 487-491. 10.1007/BF01980214.
McDermott JR, Humphreys NE, Forman SP, Donaldson DD, Grencis RK: Intraepithelial NK cell-derived IL-13 induces intestinal pathology associated with nematode infection. J Immunol. 2005, 175 (5): 3207-3213.
Keiser J, El Ela NA, El Komy E, El Lakkany N, Diab T, Chollet J, Utzinger J, Barakat R: Triclabendazole and its two main metabolites lack activity against Schistosoma mansoni in the mouse model. Am J Trop Med Hyg. 2006, 75 (2): 287-291.
Kloetzel K: Egg and pigment production in Schistosoma mansoni infections of the white mouse. Am J Trop Med Hyg. 1967, 16 (3): 293-299.
Acknowledgements
This work was supported by grants from the Welcome Trust, (UK), National Research Foundation (South Africa), The Royal Society, (UK) and MRC (South Africa). B.D. is a postdoctoral researcher of the Fonds National de la Recherche Scientifique (FNRS). For technical assistance we would like to thank Mrs Marilyn Tyler, Mrs Wendy green, Mr Reagan Peterson and Mrs Zenaria Abbas. Dr Natalie Nieuwenhuizen and Mrs J. Claire Hoving are thanked for critically readying the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Authors' contributions
RGM designed the study, carried out the S. mansoni infections, histological and statistical analysis of all infection studies and drafted the manuscript. CM carried out the S. obelata infections and the associated histological analysis. EMS carried out N. brasiliensis infections and the associated histological analysis. LCEF conducted histological stains and contributed to the analysis of all infection studies. ML participated in design of the study and assisted with infections. BD participated in carried out S. mansoni infections and drafting of the manuscript. WGCH participated in N. brasiliensis infections, participated in interpretation of the data and with drafting of the manuscript, FB conceived of the study, participated in its design, coordination, interpretation of the data and drafting the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Marillier, R.G., Michels, C., Smith, E.M. et al. IL-4/IL-13 independent goblet cell hyperplasia in experimental helminth infections. BMC Immunol 9, 11 (2008). https://doi.org/10.1186/1471-2172-9-11
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1471-2172-9-11