
Abstract
Dental caries is a major health problem worldwide. This disease results from improper interactions between oral microorganisms and host that stimulate the establishment of cariogenic biofilms on tooth surfaces, whose high and sustained acidogenicity promotes demineralization of tooth tissues. These interactions are modulated by environmental factors, among which diet has significant impact, especially regarding its carbohydrate content. Although microorganisms are recognized as having a central role in dental caries, current approaches for intervening in the establishment of cariogenic biofilms are limited. In this review, we focus on clinical studies supporting the role of mutans streptococci (MS) as promoters of cariogenic microbiota. Functions of MS in this process are analyzed under the current knowledge about the genetic and phenotypic diversity of Streptococcus mutans, the most well-known MS species. Effects of host immune responses on the establishment and pathogenesis of S. mutans are also discussed, further encouraging studies testing anti-MS therapies.
Similar content being viewed by others

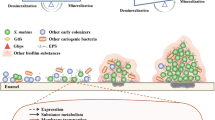
Introduction
Studies using animal models of dental caries have pointed to mutans streptococci (MS) as agents promoting establishment of cariogenic microbiota, and have shown that once transmitted from infected to uninfected rodent, these bacteria induce dental caries in the presence of a sucrose-rich diet (reviewed in [1]). A large number of studies have identified several molecular mechanisms by which MS efficiently enhance biofilm biomass in the presence of sucrose, and promote ecological shifts that lead to emergence of acidogenic and aciduric organisms [2–4, 5••, 6]. Moreover, several clinical studies further indicated the role of MS in caries development in humans [7]. Two species of MS are exclusively detected in humans, Streptococcus mutans (serotypes c, e, f and k), and Streptococcus sobrinus (serotypes d and g) [8, 9]. Most of the molecular and clinical studies analyzed S. mutans, which is the most prevalent species.
However, several observations challenge the notion that MS are the major agents implicated in the pathogenicity of dental caries: 1) MS are ubiquitous in the human populations and over 50 % of MS carriers do not show development of caries lesions in one or two years of follow-up [10], even when at heavy oral levels [11, 12, 13••]; 2) MS are part of a highly diverse group of microorganisms, including cultivable and uncultivable bacteria [14, 15••, 16] and fungi [17]; and 3) MS are not always detected in biofilms associated with a number of initial caries lesions [10, 14, 18].
The ability of MS to promote disease depends on a complex web of events of bacterial interactions with host defenses and host microbiota [19–21]. As analyzed in this review, the outcome of the host encounter with a S. mutans strain seems to depend on: 1) the virulence traits of the strain(s), 2) the host responses at the time that the strain is encountered and becomes established, and 3) the endogenous (e.g., oral microbiota) and external conditions (e.g., sucrose consumption) that can influence the bacterial−host interactions. Thus, for translational application in cariology, two important issues need to be addressed: first, whether it is possible to interfere with MS (or other pathogenic species) establishment in dental biofilms through therapeutic tools that reduce the impact of socio-cultural factors associated with unfavorable health and/or dietary habits; second, whether these anti-MS therapies would indeed lead to control of caries development. The studies reviewed here might provide answers to these questions; further encouraging research to identify the best panel of therapeutic targets, and to analyze the effects of blocking already characterized targets to control the establishment of cariogenic biofilms.
Mutans Streptococci Continue to be the Main Pathogens Linked to Dental Caries Development
Systematic analyses by Tanzer and colleagues of 313 articles published between the years of 1966 and 2000 indicated that MS are associated with dental caries initiation [7]. A 2-year prospective study of 530 intact tooth surfaces in 28 subjects showed development of caries lesions in 53 % of sites with detectable levels of MS, but in only 8 % of sites without detectable MS [22]. The advances in molecular-genetic technologies provided a broader view of the microbial diversity of the oral biofilms from subjects with different statuses of disease activity [14, 15••, 16, 23, 24]. Cross-sectional studies focused on determining the biofilm microbiota on healthy and affected tooth sites (from white-spot to dentin lesions) within subjects with distinct degrees of caries activity [14, 15••, 16, 25]. A significant number of non-MS organisms were associated with caries, including Lactobacillus spp., Actinomyces spp., Bifidobacterium dentium, Scardovia spp., Veillonela spp., Atopobium parvulum, Dialister invisus, and other non-MS streptococci (e.g. Streptococcus vestibularis, Streptococcus salivarius, Streptococcus sanguinis, Streptococcus mitis) [14, 15••, 16, 24, 25]. Levels of MS could not be significantly associated with white-spot lesions in permanent teeth in one study [14]. In contrast, a longitudinal study of microbial communities of tooth biofilms in permanent dentition significantly associated S. mutans with white-spot lesions, along with other 25 taxa [24]. Additionally, levels of MS on intact tooth sites were significantly associated with latter development of lesions in caries-affected young children [15••]. In young children without detectable signs of caries, levels of Veillonella parvula were significantly associated with future development of caries lesions [15••]. Interestingly, V. parvula strain PK1910 strongly promotes S. mutans growth in co-cultures with an initial proportion of ≈2 % S. mutans, 10 % V. parvulla and 88 % S. gordonii [26•]. S. gordonii and S. sanguinis are known to antagonize S. mutans growth by production of H2O2 [27].
The discrepancies regarding the most predominate non-MS species in cariogenic biofilms detected in molecular studies might be due to variations in methods and difficulty to define specific stages of disease. Additionally, there may be differences in the events of biofilm initiation and microbial succession during initial establishment of disease compared with established acute forms of caries, e.g. severe early-childhood caries (S-ECC). Nevertheless, non-MS streptococcal species are frequently found in biofilms of initial lesions [14, 15••, 24], confirming previous studies using culture methods [28, 29]. Non-MS streptococci represent the major pioneer species of teeth in humans [30]; thus, it is not surprising that these organisms are consistently observed with initial lesions. However, isolates from biofilms associated with lesions may be represented by atypical strains with potent acidogenicity and acid-tolerance in cariogenic biofilms [31]. Non-MS streptococci species can induce caries in germ-free rats, but the extent of disease activity is generally low compared to MS [32, 33]. Interestingly, associations were noticed between the ability to produce extracellular polysaccharide in vitro and the ability to cause dental caries in rats among non-MS streptococcal strains [33].
While the molecular mechanisms through which non-MS organisms participate in caries initiation are unclear, there is evidence that some of them may benefit from MS-derived factors present in saliva or in the acquired pellicle of tooth surfaces [34–36]. For example, S. sanguinis is a pioneer colonizer of saliva-coated tooth surfaces, likely due to its large array of adhesins [37], but does not form stable biofilms in vitro in the presence of sucrose, even though several strains produce glucosyltransferases (Gtfs) able to synthesize water-insoluble glucan (WIG) from sucrose [34]. However, biofilm formation is significantly enhanced when S. sanguinis strains are mixed with MS-derived purified Gtfs [34]. Moreover, S. mutans-derived active Gtfs bind to a variety of non-MS oral bacteria (e.g. Streptococci, Lactobacilli and Actinomyces) [38]. Of note, saliva samples from children with S-ECC contain significant amounts of S. mutans Gtfs [36]. Additionally, Gtf-encoding genes are induced in S. mutans by co-cultivation with Streptococcus oralis and A. naeslundii [39•]. Interestingly, even in proportionally low levels in mixed-species cultures, S. mutans is able to alter the three-dimensional structure of biofilm in the presence of sucrose, creating highly acidic microenvironments, which in turn promote local growth of aciduric microorganisms [39•]. More studies are also need to look for the effects of MS factors on other organisms associated with cariogenic biofilms [35, 40•].
Prospective studies of young children during the development of their oral microbiota provide data compatible with a role of MS in dental caries pathogenesis [41]. MS transmission to children could be controlled by suppressing the cariogenic microbiota of mothers until their children were 3 years old. This approach showed extended effects, since at 7 years of age only 46 % of children of treated mothers were MS-colonized, compared to 95 % of children of untreated mothers. MS suppression in mothers was highly dependent on patient commitment, as it involved intense educational approaches to reduce sucrose consumption, to improve oral hygiene, and included antimicrobial therapy [42, 43]. Absent or MS colonization later than 3 years of age significantly reduced development of caries lesions at the ages of 7 [43] to 19 years [44, 45••], compared to early MS-colonized children. Thus, it is possible to control MS transmission in humans, which has protective effects in caries development.
MS are more frequently acquired early in life, mainly during a period of 1.5 to 2.5 years of age, which has been designated the “window of infectivity” [46]. Although MS can be acquired before [11, 12, 13••, 45••, 46] or later than the so-called window of infectivity [48, 49], it seems that the risk for MS establishment decreases when the primary teeth complete eruption (by 2.5 to 3.0 years of age), even in MS-challenged populations [11]. In populations with high sucrose exposure, fluctuations in MS oral levels are observed during the first 3 years of life; about 35 % of 1 to 2.5-year-old children with high levels of MS show a decrease in MS levels 1 year later, in a fashion not explained by differences in sucrose intake [11]. The establishment of a competitive microbiota and the maturation of the host adaptive immune responses [21] could explain those observations. In MS-challenged populations, IgA antibody responses to MS antigens are more intense during 1 to 2.5 years of age compared to older children or even adults [50, 51••]. Common members of the health-associated dental microbiota, e.g. S. sanguinis, are also known to inhibit S. mutans colonization [27, 52].
Characteristics of S. mutans Clinical Isolates: Impact of Genetic and Phenotypic Diversity on S. mutans Virulence
To understand the roles of MS in dental caries development, it is important to consider S. mutans strain diversity and its influence on virulence. Some of the first studies addressing this issue compared the ability of clinical isolates of S. mutans and S. sobrinus to induce dental caries in a hamster model [53]. MS diversity was later shown to influence the ability of individual MS strains to be transmitted from mother to child, as strains producing a more efficient array of bacteriocins were more frequently transmitted between mother−child pairs compared to strains with a lower spectrum of bacteriocin production [54•]. Thus, S. mutans strains differ in their competitive ability to establish in the oral microbiota. The phenotypic diversity of S. mutans was explored when tracing the sources and routes of S. mutans transmission to children during the development of the oral microbiota [55, 56]. Genotyping methods, used to track strains within families, day care schools and other populations [57, 58], identified both vertical and horizontal routes of transmission [12, 47, 55, 59, 60•, 61]. Genotyping studies also demonstrated that S. mutans strains acquired early in life tend to persist in the oral cavity for several years, although specific strains can be lost or acquired during life [48, 62, 63]. These studies raise the question of whether this diversity could influence the disease development within MS-colonized subjects.
The sequencing of the strain UA159 isolated from a caries-active child was critical to understand S. mutans biology [64•]. Further comparative analysis of S. mutans genomes revealed that S. mutans have a core genome formed by genes present in all the strains, and a dispensable genome, composed of strain-specific genes [65, 66]. The S. mutans dispensable genome includes virulence genes involved in biofilm formation, sugar metabolism and bacteriocin production, and a class of laterally acquired genes including genomic islands with atypical C + G contents [66]. The sequencing of the genomes of 57 S. mutans isolates from eight countries confirmed that the S. mutans genomes are dynamically altered mostly by lateral gene transfer, but the core genome includes a set of exclusive genes not present in non-MS streptococci [67•]. Interestingly, the S. mutans-exclusive core genome contains a significant proportion of genes encoding small peptides that are potentially involved in cell signaling and gene regulation [67•], implying that an important trait for S. mutans biology relies on its ability to rapidly respond and adapt to environmental changes. The dynamic adaptation of S. mutans to oral niches most likely involves several processes, including gene expression changes, post-transcriptional regulatory mechanisms, selection of polymorphisms associated with specific enzymatic/protein activities, and acquisition or loss of genes [67•, 68–70, 71•, 72••].
Remarkably, no significant differences were observed in virulence gene content, including those encoding Gtfs (gtfB/C/D), glucan-binding proteins (gbpA/B/C) or involved in acid production and tolerance (atpA, fabM) among S. mutans isolates from caries-free children versus children with S-ECC [68], although genes with putative roles in bacteriocin production, carbohydrate metabolism or other unknown functions were often found to be different between isolates of low-caries and caries-active children [65]. S. mutans strains clearly differ with respect to their ability to form biofilms in vitro [13••, 71•, 73]; these abilities are positively associated with the diversity in amounts of GtfB and GtfC (enzymes that synthesize WIG) produced in vitro [73], as well as with amounts of glucan-binding protein B (GbpB) [71•], a surface protein involved in cell-wall biogenesis and biofilm formation [74]. Patterns of production of these virulence proteins appear to be strain-specific [60•, 71•, 73]. Prospective comparisons among S. mutans isolates from children 1 to 2.5 years of age carrying significant oral levels of MS revealed that isolates from caries-free children produced significantly less WIG in vitro than strains isolated at baseline from children who developed a mean of four caries lesions 1 year later [13••]. The synthesis of WIG in vitro by the colonizing strains was significantly associated with future development of lesions. Although a high diversity in acidogenicity and aciduricity was also detected among strains (final pH ranging from 3.89 to 4.36), no significant associations with disease activity were verified [13••]. Thus, once MS are established, the acidogenesis of biofilms may be sustained by several organisms [15••, 16, 31, 75]. Similar associations between syntheses of WIG with disease were also detected in young adults [76]. Differing capacities of WIG production could not be associated with polymorphisms in genes encoding GtfB and GtfC, because these enzymes show large sequence diversity [73]. However, phylogenetic analyses of gtf genes of S. mutans and non-MS streptococci indicated amino acid changes clustered in the catalytic domains of these enzymes [77], which may explain the differential activity of the enzymes and/or type of glucan produced.
Additionally, analyses of the diversity of bacterial regulatory systems include the characterization of signal transduction systems designated as two-component regulatory systems (TCS) [78], which are widely applied by prokaryotic organisms to rapidly sense and respond to environmental stimuli [79]. The number of TCS vary from 12 to 14 among S. mutans strains [80, 81]. There is also high variation in the S. mutans quorum-sensing pathways that is important for biofilm formation [72••, 82]. The high diversity in the activities of genes encoding transcriptional regulators (vicR, comE, rr11 and covR) of S. mutans virulence genes (including gtfB/C, comCD, gbpB, and gbpC) indicates that the TCS regulatory networks may be strain-specific [69]. By identifying the regulons of TCS involved in biofilm formation through direct regulation of gtfB/C and gbpB (VicRK and CovR), a set of genes involved in cell wall biogenesis was found necessary for S. mutans to initiate biofilms in the presence of sucrose [83]. Those include genes encoding murein hydrolases (LysM, SMU.2146c, SmaA) and a gene putatively involved in the surface attachment of teichoic acids (epsC) [83]. Interestingly, these genes are not involved in planktonic growth phenotypes [83]. The mechanisms of cell wall biogenesis and turnover in S. mutans are largely unknown, but analysis of 15 clinical S. mutans strains revealed a high diversity in patterns of autolysis, likely as result of different activities of murein hydrolases and other cell wall components [72••]. These results indicate ecological relevance of S. mutans cell wall diversity. S. mutans are known to differ from other oral streptococci with respect to cell wall components, including amounts of lipoteichoic acid (LTA) [3]. Soluble LTA seems to modulate Gtfs adsorption to tooth surfaces [3] and is found in high amounts in cariogenic biofilms [84]. LTA might be also involved in S. mutans interactions with other bacteria in biofilms [85•]. Further analyses of S. mutans cell wall components may reveal novel therapeutic targets.
Host Response to MS: Antibody Responses May Affect Susceptibility to S. mutans Colonization
The oral cavity is protected by a large number of soluble and cellular components of the innate and adaptive systems. The unique individual patterns of adaptive immune responses occur in part because of the enormous diversity of antigen-specific receptors of T and B lymphocytes and the complexity of the events involved in the activation of effector functions in these cells [86]. Major components of oral specific immunity are the SIgA antibodies, which are dimeric forms of IgA associated with a secretory component that confers antibody resistance to bacterial and host proteases [87]. The functions of salivary SIgA in controlling bacterial colonization include bacterial agglutination, blocking of adhesins to salivary pellicle or other biofilm components, interference in enzymatic activities important for colonization and immunomodulatory activities [87, 88]. The pioneer colonizers of the mucosa and teeth (S. mitis and S. sanguinis, respectively) produce proteases that can cleave the most abundant SIgA antibody isotype in saliva (SIgA1), perhaps enhancing the colonization potential of these bacteria. Abundant evidence indicates that the mucosal immune response matures rather quickly in the infant. In the oral cavity this phenomenon is evidenced by measureable levels of salivary IgA antibody reactive with pioneer oral organisms within the first weeks of life [87]. Even these earliest responses differ in specificity, amount, timing and IgA subclass among infants, emphasizing the individuality in the development of and response to the emerging microbiota. Interestingly, antibodies reactive with pioneer microbiota and S. mutans are detected in the oral fluids of neonates [89]. The salivary SIgA antibody to virulence antigens of these organisms showed specificities distinct from those of SIgA in the respective maternal milk [89], suggesting active immunological interactions very early in life.
As oral habitats become more diverse with age, so do the consequent microbial challenges to the mucosal immune system. Conditions that favor MS colonization bring with them the potential for responses to virulence-associated components. However, children can respond differently to these organisms, influenced by factors such as extent of colonization or maturation of immune response. In fact, siblings often demonstrate unique patterns of antibody specificities, despite being challenged with the same maternal subspecies [21, Nogueira, unpublished observations]. Although it is clear that most children demonstrate salivary antibody to MS persisting through adulthood, the relationship of antibody levels to MS colonization and disease remains somewhat elusive [21, 90]. Nogueira and colleagues [51••] investigated patterns of specificities of salivary SIgA in a longitudinal study of children initially 6 months old. In this population of children heavily exposed to sucrose and to MS, children with natural immunodominant responses to GbpB (a potential vaccine target) were less susceptible to S. mutans acquisition. About 76 % of the babies in which MS were not detected showed robust amounts of salivary SIgA antibody to GbpB, while only 38.1 % of children already colonized by MS showed SIgA reactive with this antigen [51••]. Early colonized children developed one to seven cavity lesions at the end of the study, while non MS-colonized children remained free of lesions [50]. These studies indicated that children under heavy MS challenge can mount secretory IgA responses to S. mutans-specific antigens in the first year of life, and that vigorous immune responses to antigens important to colonization may influence susceptibility to MS acquisition and caries development.
The origin of early immune responses reactive with MS antigen epitopes may stem from more than one source. Infants may respond to these epitopes following transient exposure from caregiver saliva early in life. In addition, pioneer microbiota may initiate salivary IgA responses that react with, and possibly prime responses to, MS [89]. Several MS virulence components share MHC Class II binding peptides with pioneer colonizers’ protein homologues [91]. Furthermore, culture supernatants of S. mitis and S. salivarius contained antigens reactive with rat IgG or chicken IgY antibody to purified S. mutans GbpB. Mass spectroscopic analysis identified homologues to GbpB in pioneer streptococci.
Associations between caries development, colonization with caries-associated microbiota, and immunity were explored in children during the initial transition to mixed dentition. Caries-free (CF) and caries-active (CA) children were evaluated at 3−4 years of age and a year later, for total levels of salivary IgA and IgA antibody reactive with S. mutans epitopes [92]. As expected, MS was associated with caries development in this population, and salivary IgA antibody levels to Gtfs, GbpB and Antigen I/II SBR increased in both groups. However, in the CF cohort, salivary antibody levels to seven peptides associated with presumed domains of biological importance increased at follow-up, in contrast to increases to only three peptides in CA-cohort salivas. Similar findings were previously observed in older children [93]. Multivariate modeling showed that a lower baseline level of salivary IgA anti-GbpB was associated with higher dental caries-risk [92], further indicating that the breadth of mucosal IgA response to S. mutans virulence-epitopes may influence the degree to which these microorganisms can cause disease.
Devising Effective Anti-Caries Vaccine Strategies
There are recent reviews regarding possible anti-bacterial therapies to control dental caries [94]. Infectious diseases have long been intercepted by enabling host defense mechanisms through immunization prior to infection, or by providing external sources of antibody directed to the disease-responsible entities. These approaches have often been successful, are generally cost-effective, and can reach a large percentage of the population. Thus, active and passive immunization strategies have been pursued to deal with organisms implicated in dental caries [95•], and have the potential to complement existing anti-caries regimens [41–43]. Although several microorganisms have been associated with various forms of dental caries, MS have been the foci of these immunological approaches. MS offer obvious cell wall and extracellular targets that are central to their attachment and accumulation in the oral biofilm. As the oral microbiota becomes more fully catalogued, other acidogenic microorganisms separately, or in community, may be found to participate in disease development [15••, 16, 23, 24]. Nonetheless, MS remains prominent in most molecular genetic profiles of incipient dental disease; thus, it continues as a reasonable vaccine target.
Adaptive immune regimes using protein-based vaccines have shown that induction of salivary IgA antibody to components of MS that are responsible for adhesion to the tooth surface (antigen I/II), for glucan formation (Gtf), or for aggregation via Gbp’s, inhibits dental caries formation in experimental systems [21]. Thus, proof of principle exists for a dental caries vaccine; at least one approach is under commercial development [personal communication]. Considerable attention has also been focused on effective adjuvants [96] and on creating protein-based or peptide-based constructs that increase the breadth of protection through biochemical or recombinant combinations of putatively protective epitopes from these virulence components. Conjugating Gtf to glucan has also been shown to extend experimental protection to the two MS species pathogenic in human [98; unpublished observations]. Mucosal immunity, as expressed in the saliva as secretory IgA, is considered to offer the most obvious mechanism for provision of potentially protective antibody. Nasal antigen application is being pursued as a primary site for delivery of antigen to inductive sites that result in salivary IgA antibody formation [96, 97]. Inducing protective levels of salivary antibody prior to establishment of cariogenic streptococci continues to be a goal.
DNA vaccines have also been explored for their ability to provoke protective responses. Theoretically, this approach could permit, depending on the construct, expression of a combination of MS virulence components and might result in an extended duration of epitope expression at the desired site. DNA vaccine constructs containing the A-P fragment of the S. mutans pac gene [98], and/or catalytic and glucan binding fragments from S. mutans GtfB or S. sobrinus Gtf genes, cloned into plasmid vectors, have been delivered experimentally via routes similar to those used for protein-based vaccines. Recent experiments have concentrated on optimizing intranasal administration [99], adjuvant composition, and use of combinations of DNA vaccine-protein for immunization [100].
Adopting the adaptive approach in humans has been delayed, in part, by the lack of acceptable adjuvants and reluctance to add another active vaccine to those currently recommended for pediatric use. Passive immune approaches, which do not have these issues, thus hold some promise. IgG or IgY antibodies to intact cells or virulence elements, when introduced into the oral cavity by topical application, diet, drinking water or intravenous transfer, have each reduced dental caries in experimental animals during challenge with MS [21]. Small scale clinical trials using topical application of monoclonal/ transgenic reagents directed to S. mutans cell surface epitopes were met with some success [101]. Kruger and coworkers [102•] designed a single-chain variable fragment (scFv) with anti-adhesin properties and expressed it on the surface of Lactobacilli, which, after colonizing the oral cavity, could inhibit experimental caries. Dietary administration of human scFv antibody to Gtf epitopes was also shown to modify experimental MS infection [91; unpublished observations].
Thus, considerable experimental evidence exists for the vaccine approach. The current challenges of this approach include the likelihood of multiple cariogenic species, the ability to induce sufficient and long-lasting levels of protective antibody in young children, and, especially in the case of passive immunity, the requirement to continuously apply protective levels of antibody to address a chronic infection.
Conclusions
MS oral colonization promotes dental caries development in humans. This disease activity is affected by the timing of MS establishment in the oral microbiota, which is modulated by host responses during the course of colonization and by external and microbial factors that enhance or mitigate MS competitiveness. The most prevalent MS species, S. mutans, shows high genetic diversity, and clinical studies highlight its significance in strain ability to promote disease. The role of S. mutans in cariogenesis is likely not to be entirely dependent on bacterial levels, since virulence factors expressed by this species are shown to alter the biofilm structure and to promote ecological shifts leading to an acidogenic and acid-tolerant microbiota. Other organisms in the biofilm may also participate in caries development, but further studies are needed to identify mechanisms by which non-MS organisms would drive (in combination or not with MS) the ecological shifts leading to cariogenic biofilms. Finally, there is evidence that natural salivary IgA antibody response of children to MS virulence-associated antigens affects susceptibility to MS colonization, supporting the application of anti-MS vaccines. Combination of different therapeutic approaches to control caries disease will likely be necessary to benefit susceptible populations.
References
Papers of particular interest, published recently, have been highlighted as: • Of importance •• Of major importance
Tanzer JM. Dental caries is a transmissible infectious disease: the Keyes and Fitzgerald revolution. J Dent Res. 1995;74:1536–42.
Loesche WJ. Role of Streptococcus mutans in human dental decay. Microbiol Rev. 1986;50:353–80.
Hamada S, Slade HD. Biology, immunology, and cariogenicity of Streptococcus mutans. Microbiol Rev. 1980;44:331–84.
Banas JA. Virulence properties of Streptococcus mutans. Front Biosci. 2004;9:1267–77.
Bowen WH, Koo H. Biology of Streptococcus mutans-derived glucosyltransferases: role in extracellular matrix formation of cariogenic biofilms. Caries Res. 2011;45:69–86. Comprehensive review about the extracellular matrix of cariogenic biofilms.
Lemos JA, Burne RA. A model of efficiency: stress tolerance by Streptococcus mutans. Microbiology. 2008;154:3247–55.
Tanzer JM, Livingston J, Thompson AM. The microbiology of primary dental caries in humans. J Dent Educ. 2001;65:1028–37.
Whiley RA, Beighton D. Current classification of the oral streptococci. Oral Microbiol Immunol. 1998;13:195–216.
Nakano K, Nomura R, Nakagawa I, Hamada S, Ooshima T. Demonstration of Streptococcus mutans with a cell wall polysaccharide specific to a new serotype, k, in the human oral cavity. J Clin Microbiol. 2004;42:198–202.
van Houte J. Microbiological predictors of caries risk. Adv Dent Res. 1993;7:87–96.
Mattos-Graner RO, Correa MS, Latorre MR, Peres RC, Mayer MP. Mutans streptococci oral colonization in 12-30-month-old Brazilian children over a one-year follow-up period. J Public Health Dent. 2001;61:161–7.
Alves AC, Nogueira RD, Stipp RN, Pampolini F, Moraes AB, Goncalves RB, et al. Prospective study of potential sources of Streptococcus mutans transmission in nursery school children. J Med Microbiol. 2009;58:476–81.
Mattos-Graner RO, Smith DJ, King WF, Mayer MP. Water-insoluble glucan synthesis by mutans streptococcal strains correlates with caries incidence in 12- to 30-month-old children. J Dent Res. 2000;79:1371–7. This study shows the influence of diversity in virulence-associated factors of S. mutans with disease activity in humans, further indicating that diversity in production of water-insoluble glucan influences S. mutans virulence.
Aas JA, Griffen AL, Dardis SR, Lee AM, Olsen I, Dewhirst FE, et al. Bacteria of dental caries in primary and permanent teeth in children and young adults. J Clin Microbiol. 2008;46:1407–17.
Gross EL, Beall CJ, Kutsch SR, Firestone ND, Leys EJ, Griffen AL. Beyond Streptococcus mutans: dental caries onset linked to multiple species by 16S rRNA community analysis. PLoS One. 2012;7:e47722. This paper includes longitudinal analysis of biofilm composition of young children to identify predictors of caries onset. Veillonella spp. was a predictor of caries lesions among initially healthy children.
Tanner AC, Kent Jr RL, Holgerson PL, Hughes CV, Loo CY, Kanasi E, et al. Microbiota of severe early childhood caries before and after therapy. J Dent Res. 2011;90:1298–305.
de Carvalho FG, Silva DS, Hebling J, Spolidorio LC, Spolidorio DM. Presence of mutans streptococci and Candida spp. in dental plaque/dentine of carious teeth and early childhood caries. Arch Oral Biol. 2006;51:1024–8.
van Ruyven FO, Lingstrom P, van Houte J, Kent R. Relationship among mutans streptococci, “low-pH” bacteria, and lodophilic polysaccharide-producing bacteria in dental plaque and early enamel caries in humans. J Dent Res. 2000;79:778–84.
Jenkinson HF, Lamont RJ. Oral microbial communities in sickness and in health. Trends Microbiol. 2005;13:589–95.
Marsh PD. Microbial ecology of dental plaque and its significance in health and disease. Adv Dent Res. 1994;8:263–71.
Smith DJ, Mattos-Graner RO. Secretory immunity following mutans streptococcal infection or immunization. Curr Top Microbiol Immunol. 2008;319:131–56.
Kristoffersson K, Grondahl HG, Bratthall D. The more Streptococcus mutans, the more caries on approximal surfaces. J Dent Res. 1985;64:58–61.
Corby PM, Lyons-Weiler J, Bretz WA, Hart TC, Aas JA, Boumenna T, et al. Microbial risk indicators of early childhood caries. J Clin Microbiol. 2005;43:5753–9.
Torlakovic L, Klepac-Ceraj V, Ogaard B, Cotton SL, Paster BJ, Olsen I. Microbial community succession on developing lesions on human enamel. J Oral Microbiol. 2012;4. doi:10.3402/jom.v4i0.16125
Becker MR, Paster BJ, Leys EJ, Moeschberger ML, Kenyon SG, Galvin JL, et al. Molecular analysis of bacterial species associated with childhood caries. J Clin Microbiol. 2002;40:1001–9.
Liu J, Wu C, Huang IH, Merritt J, Qi F. Differential response of Streptococcus mutans towards friend and foe in mixed-species cultures. Microbiology. 2011;157:2433–44. This paper proposes that Veillonella parvula strain enhances S. mutans ability to establish in biofilms. Transcriptomic analyses of S. mutans in co-cultures with V. parvula and its typical competitor species, S. gordonii indicate that V. parvula relieves S. mutans from the inhibitory functions of S. gordonii.
Kreth J, Zhang Y, Herzberg MC. Streptococcal antagonism in oral biofilms: Streptococcus sanguinis and Streptococcus gordonii interference with Streptococcus mutans. J Bacteriol. 2008;190:4632–40.
Nyvad B, Kilian M. Comparison of the initial streptococcal microflora on dental enamel in caries-active and in caries-inactive individuals. Caries Res. 1990;24:267–72.
Sansone C, van Houte J, Joshipura K, Kent R, Margolis HC. The association of mutans streptococci and non-mutans streptococci capable of acidogenesis at a low pH with dental caries on enamel and root surfaces. J Dent Res. 1993;72:508–16.
Diaz PI, Chalmers NI, Rickard AH, Kong C, Milburn CL, Palmer Jr RJ, et al. Molecular characterization of subject-specific oral microflora during initial colonization of enamel. Appl Environ Microbiol. 2006;72:2837–48.
Svensater G, Borgstrom M, Bowden GH, Edwardsson S. The acid-tolerant microbiota associated with plaque from initial caries and healthy tooth surfaces. Caries Res. 2003;37:395–403.
Willcox MD, Knox KW, Green RM, Drucker DB. An examination of strains of the bacterium Streptococcus vestibularis for relative cariogenicity in gnotobiotic rats and adhesion in vitro. Arch Oral Biol. 1991;36:327–33.
Willcox MD, Drucker DB, Green RM. In vivo dental plaque-forming ability and cariogenicity of the bacterium Streptococcus bovis in gnotobiotic rats. Arch Oral Biol. 1990;35:163–6.
Hamada S, Torii M, Kotani S, Tsuchitani Y. Adherence of Streptococcus sanguis clinical isolates to smooth surfaces and interactions of the isolates with Streptococcus mutans glucosyltransferase. Infect Immun. 1981;32:364–72.
Tamesada M, Kawabata S, Fujiwara T, Hamada S. Synergistic effects of streptococcal glucosyltransferases on adhesive biofilm formation. J Dent Res. 2004;83:874–9.
Vacca Smith AM, Scott-Anne KM, Whelehan MT, Berkowitz RJ, Feng C, Bowen WH. Salivary glucosyltransferase B as a possible marker for caries activity. Caries Res. 2007;41:445–50.
Xu P, Alves JM, Kitten T, Brown A, Chen Z, Ozaki LS, et al. Genome of the opportunistic pathogen Streptococcus sanguinis. J Bacteriol. 2007;189:3166–75.
Vacca-Smith AM, Bowen WH. Binding properties of streptococcal glucosyltransferases for hydroxyapatite, saliva-coated hydroxyapatite, and bacterial surfaces. Arch Oral Biol. 1998;43:103–10.
Xiao J, Klein MI, Falsetta ML, Lu B, Delahunty CM, Yates 3rd JR, et al. The exopolysaccharide matrix modulates the interaction between 3D architecture and virulence of a mixed-species oral biofilm. PLoS Pathog. 2012;8:e1002623. This study demonstrates that a small population of S. mutans can modulate the development of a 3D matrix scaffold in mixed-species biofilms when sucrose is available. The presence of the matrix is essential for the creation of localized and highly acidic microenvironments throughout the biofilm 3D architecture. The acidic niches promote ecological shifts (favoring aciduric organisms such as S. mutans), while creating demineralizing sites at the tooth-enamel surface; these are critical for the onset of the dental caries disease.
Gregoire S, Xiao J, Silva BB, Gonzalez I, Agidi PS, Klein MI, et al. Role of glucosyltransferase B in interactions of Candida albicans with Streptococcus mutans and with an experimental pellicle on hydroxyapatite surfaces. Appl Environ Microbiol. 2011;77:6357–67. This paper proposes that S. mutans-derived Gtfs may mediate the association of C. albicans and cariogenic biofilm.
Alaluusua S, Renkonen OV. Streptococcus mutans establishment and dental caries experience in children from 2 to 4 years old. Scand J Dent Res. 1983;91:453–7.
Kohler B, Andreen I, Jonsson B, Hultqvist E. Effect of caries preventive measures on Streptococcus mutans and lactobacilli in selected mothers. Scand J Dent Res. 1982;90:102–8.
Kohler B, Andreen I. Influence of caries-preventive measures in mothers on cariogenic bacteria and caries experience in their children. Arch Oral Biol. 1994;39:907–11.
Kohler B, Andreen I. Mutans streptococci and caries prevalence in children after early maternal caries prevention: a follow-up at eleven and fifteen years of age. Caries Res. 2010;44:453–8.
Kohler B, Andreen I. Mutans streptococci and caries prevalence in children after early maternal caries prevention: a follow-up at 19 years of age. Caries Res. 2012;46:474–80. This study describes the outcomes of the preventive program capable to interfere in MS transmission to children during the phase of increased risk of MS colonization that was described in reference 43.
Caufield PW, Cutter GR, Dasanayake AP. Initial acquisition of mutans streptococci by infants: evidence for a discrete window of infectivity. J Dent Res. 1993;72:37–45.
Fujiwara T, Sasada E, Mima N, Ooshima T. Caries prevalence and salivary mutans streptococci in 0-2-year-old children of Japan. Community Dent Oral Epidemiol. 1991;19:151–4.
Cheon K, Moser SA, Wiener HW, Whiddon J, Momeni SS, Ruby JD, et al. Characteristics of Streptococcus mutans genotypes and dental caries in children. Eur J Oral Sci. 2013;121:148–55.
Okada M, Taniguchi Y, Hayashi F, Doi T, Suzuki J, Sugai M, et al. Late established mutans streptococci in children over 3 years old. Int J Dent. 2010;2010:732468.
Nogueira RD, Alves AC, King WF, Goncalves RB, Hofling JF, Smith DJ, et al. Age-specific salivary immunoglobulin A response to Streptococcus mutans GbpB. Clin Vaccine Immunol. 2007;14:804–7.
Nogueira RD, Alves AC, Napimoga MH, Smith DJ, Mattos-Graner RO. Characterization of salivary immunoglobulin A responses in children heavily exposed to the oral bacterium Streptococcus mutans: influence of specific antigen recognition in infection. Infect Immun. 2005;73:5675–84. This study shows that intense and immunodominant responses to GbpB of S. mutans are highly prevalent among children who do not acquire S. mutans early in life in an SM-challenged population.
Caufield PW, Dasanayake AP, Li Y, Pan Y, Hsu J, Hardin JM. Natural history of Streptococcus sanguinis in the oral cavity of infants: evidence for a discrete window of infectivity. Infect Immun. 2000;68:4018–23.
Kohler B, Krasse B. Human strains of mutans streptococci show different cariogenic potential in the hamster model. Oral Microbiol Immunol. 1990;5:177–80.
Gronroos L, Saarela M, Matto J, Tanner-Salo U, Vuorela A, Alaluusua S. Mutacin production by Streptococcus mutans may promote transmission of bacteria from mother to child. Infect Immun. 1998;66:2595–600. This study indicates the importance of bacteriocin production phenotypes in S. mutans competitiveness during oral colonization in humans.
Berkowitz RJ, Jordan HV, White G. The early establishment of Streptococcus mutans in the mouths of infants. Arch Oral Biol. 1975;20:171–4.
Berkowitz RJ. Mutans streptococci: acquisition and transmission. Pediatr Dent. 2006;28:106–9.
Saarela M, Alaluusua S, Takei T, Asikainen S. Genetic diversity within isolates of mutans streptococci recognized by an rRNA gene probe. J Clin Microbiol. 1993;31:584–7.
Kozai K, Nakayama R, Tedjosasongko U, Kuwahara S, Suzuki J, Okada M, et al. Intrafamilial distribution of mutans streptococci in Japanese families and possibility of father-to-child transmission. Microbiol Immunol. 1999;43:99–106.
Li Y, Caufield PW. The fidelity of initial acquisition of mutans streptococci by infants from their mothers. J Dent Res. 1995;74:681–5.
Mattos-Graner RO, Li Y, Caufield PW, Duncan M, Smith DJ. Genotypic diversity of mutans streptococci in Brazilian nursery children suggests horizontal transmission. J Clin Microbiol. 2001;39:2313–6. Provides evidence of horizontal transmission of MS. This paper shows that isolates from different children but with equal genotypic pattern show a nearly identical production of GtfB and GtfC in vitro. Thus, the genomic traits affecting virulence phenotypes are conserved after colonization of different hosts.
Emanuelsson IR, Li Y, Bratthall D. Genotyping shows different strains of mutans streptococci between father and child and within parental pairs in Swedish families. Oral Microbiol Immunol. 1998;13:271–7.
Emanuelsson IR, Thornqvist E. Genotypes of mutans streptococci tend to persist in their host for several years. Caries Res. 2000;34:133–9.
Klein MI, Florio FM, Pereira AC, Hofling JF, Goncalves RB. Longitudinal study of transmission, diversity, and stability of Streptococcus mutans and Streptococcus sobrinus genotypes in Brazilian nursery children. J Clin Microbiol. 2004;42:4620–6.
Ajdic D, McShan WM, McLaughlin RE, Savic G, Chang J, Carson MB, et al. Genome sequence of Streptococcus mutans UA159, a cariogenic dental pathogen. Proc Natl Acad Sci U S A. 2002;99:14434–9. Landmark to study S. mutans biology.
Zhang L, Foxman B, Drake DR, Srinivasan U, Henderson J, Olson B, et al. Comparative whole-genome analysis of Streptococcus mutans isolates within and among individuals of different caries status. Oral Microbiol Immunol. 2009;24:197–203.
Waterhouse JC, Russell RR. Dispensable genes and foreign DNA in Streptococcus mutans. Microbiology. 2006;152:1777–88.
Cornejo OE, Lefebure T, Bitar PD, Lang P, Richards VP, Eilertson K, et al. Evolutionary and population genomics of the cavity causing bacteria Streptococcus mutans. Mol Biol Evol. 2013;30:881–93. This study shows that the S. mutans genomes are dynamically altered but have a core genome not present in non-MS streptococci.
Argimon S, Caufield PW. Distribution of putative virulence genes in Streptococcus mutans strains does not correlate with caries experience. J Clin Microbiol. 2011;49:984–92.
Stipp RN, Goncalves RB, Hofling JF, Smith DJ, Mattos-Graner RO. Transcriptional analysis of gtfB, gtfC, and gbpB and their putative response regulators in several isolates of Streptococcus mutans. Oral Microbiol Immunol. 2008;23:466–73.
Saxena D, Li Y, Caufield PW. Identification of unique bacterial gene segments from Streptococcus mutans with potential relevance to dental caries by subtraction DNA hybridization. J Clin Microbiol. 2005;43:3508–11.
Mattos-Graner RO, Jin S, King WF, Chen T, Smith DJ, Duncan MJ. Cloning of the Streptococcus mutans gene encoding glucan binding protein B and analysis of genetic diversity and protein production in clinical isolates. Infect Immun. 2001;69:6931–41. This study associates biofilm formation with amounts of GbpB produced in clinical strains.
Palmer SR, Miller JH, Abranches J, Zeng L, Lefebure T, Richards VP, et al. Phenotypic heterogeneity of genomically-diverse isolates of Streptococcus mutans. PLoS One. 2013;8:e61358. Analyses of 15 clinical S. mutans strains revealed that this species developed several strategies to thrive in the oral cavity that can influence dental caries pathogenesis.
Mattos-Graner RO, Napimoga MH, Fukushima K, Duncan MJ, Smith DJ. Comparative analysis of Gtf isozyme production and diversity in isolates of Streptococcus mutans with different biofilm growth phenotypes. J Clin Microbiol. 2004;42:4586–92.
Duque C, Stipp RN, Wang B, Smith DJ, Hofling JF, Kuramitsu HK, et al. Downregulation of GbpB, a component of the VicRK regulon, affects biofilm formation and cell surface characteristics of Streptococcus mutans. Infect Immun. 2011;79:786–96.
Nakajo K, Takahashi N, Beighton D. Resistance to acidic environments of caries-associated bacteria: Bifidobacterium dentium and Bifidobacterium longum. Caries Res. 2010;44:431–7.
Napimoga MH, Kamiya RU, Rosa RT, Rosa EA, Hofling JF, Mattos-Graner R, et al. Genotypic diversity and virulence traits of Streptococcus mutans in caries-free and caries-active individuals. J Med Microbiol. 2004;53:697–703.
Argimon S, Alekseyenko AV, DeSalle R, Caufield PW. Phylogenetic analysis of glucosyltransferases and implications for the coevolution of mutans streptococci with their mammalian hosts. PLoS One. 2013;8:e56305.
Smith EG, Spatafora GA. Gene regulation in S. mutans: complex control in a complex environment. J Dent Res. 2012;91:133–41.
Stephenson K, Hoch JA. Virulence- and antibiotic resistance-associated two-component signal transduction systems of Gram-positive pathogenic bacteria as targets for antimicrobial therapy. Pharmacol Ther. 2002;93:293–305.
Biswas I, Drake L, Erkina D, Biswas S. Involvement of sensor kinases in the stress tolerance response of Streptococcus mutans. J Bacteriol. 2008;190:68–77.
Song L, Sudhakar P, Wang W, Conrads G, Brock A, Sun J, et al. A genome-wide study of two-component signal transduction systems in eight newly sequenced mutans streptococci strains. BMC Genomics. 2012;13:128.
Klein MI, Bang S, Florio FM, Hofling JF, Goncalves RB, Smith DJ, et al. Genetic diversity of competence gene loci in clinical genotypes of Streptococcus mutans. J Clin Microbiol. 2006;44:3015–20.
Stipp RN, Boisvert H, Smith DJ, Hofling JF, Duncan MJ, Mattos-Graner RO. CovR and VicRK regulate cell surface biogenesis genes required for biofilm formation in Streptococcus mutans. PLoS One. 2013;8:e58271.
Rolla G, Iversen OJ, Bonesvoll P. Lipoteichoic acid - the key to the adhesiveness of sucrose grown Streptococcus mutans. Adv Exp Med Biol. 1978;107:607–17.
Klein MI, Xiao J, Lu B, Delahunty CM, Yates III JR, Koo H. Streptococcus mutans protein synthesis during mixed-species biofilm development by high-throughput quantitative proteomics. PLoS One. 2012;7:e45795. The assembly of the exopolysaccharide-rich matrix and the concomitant development of acidic niches appear to modulate the regulation of stress survival pathways by S. mutans. GtfB, GtfC and glucan-binding proteins are all induced, leading to continuous biofilm accretion and development of a favorable milieu for the survival and growth of cariogenic organisms.
Abbas AK, Lichtman AH, Pillai S. Cellular and molecular immunology. 6th ed. Saunders Elsevier; 2007.
Brandtzaeg P. Do salivary antibodies reliably reflect both mucosal and systemic immunity? AnnNY Acad Sci. 2007;1098:288–311.
Robinette RA, Oli MW, McArthur WP, Brady LJ. Beneficial immunomodulation by Streptococcus mutans anti-P1 monoclonal antibodies is Fc independent and correlates with increased exposure of a relevant target epitope. J Immunol. 2009;183:4628–38.
Nogueira RD, Sesso ML, Borges MC, Mattos-Graner RO, Smith DJ, Ferriani VP. Salivary IgA antibody responses to Streptococcus mitis and Streptococcus mutans in preterm and fullterm newborn children. Arch Oral Biol. 2012;57:647–53.
Ito T, Maeda T, Senpuku H. Roles of salivary components in Streptococcus mutans colonization in a new animal model using NOD/SCID.e2f1−/− mice. PLoS One. 2012;7:e32063.
King WF, Chen T, Nogueira R, Mattos-Graner R, Smith DJ. Epitopes shared among pioneer oral flora and Streptococcus mutans GbpB. In: Sasano T, Suzuki O, editors. Interface oral health. Tokyo: Springer; 2009. p. 416–7.
Parisotto TM, King WF, Duque C, Mattos-Graner RO, Steiner-Oliveira C, Nobre-Dos-Santos M, et al. Immunological and microbiologic changes during caries development in young children. Caries Res. 2011;45:377–85.
Bratthall D, Serinirach R, Hamberg K, Widerstrom L. Immunoglobulin A reaction to oral streptococci in saliva of subjects with different combinations of caries and levels of mutans streptococci. Oral Microbiol Immunol. 1997;12:212–8.
ten Cate JM, Zaura E. The numerous microbial species in oral biofilms: how could antibacterial therapy be effective? Adv Dent Res. 2012;24:108–11.
Taubman MA, Nash DA. The scientific and public-health imperative for a vaccine against dental caries. Nat Rev Immunol. 2006;6:555–63. This paper states the case for the potential benefit of active immunization, especially in a public health context.
Liang S, Hosur KB, Nawar HF, Russell MW, Connell TD, Hajishengallis G. In vivo and in vitro adjuvant activities of the B subunit of Type IIb heat-labile enterotoxin (LT-IIb-B5) from Escherichia coli. Vaccine. 2009;27:4302–8.
Taubman MA, Smith DJ, Lees A. Preparation and immunogenicity of monoepitopic and polyepitopic tetanus toxoid-glucosyltransferase peptide conjugate vaccines. IADR 81st General Session, Goteborg, Sweden, J Dent Res. 2003, 82B, Abst. 227.
Jia R, Guo JH, Fan MW, Bian Z, Chen Z, Peng B, et al. Mucosal immunization against dental caries with plasmid DNA encoding pac gene of Streptococcus mutans in rats. Vaccine. 2004;22:2511–6.
Liu F, Yang J, Zhang Y, Zhou D, Chen Y, Gai W, et al. Recombinant flagellins with partial deletions of the hypervariable domain lose antigenicity but not mucosal adjuvancy. Biochem Biophys Res Commun. 2010;392:582–7.
Shi W, Li YH, Liu F, Yang JY, Zhou DH, Chen YQ, et al. Flagellin enhances saliva IgA response and protection of anti-caries DNA vaccine. J Dent Res. 2012;91:249–54.
Ma JK, Hikmat BY, Wycoff K, Vine ND, Chargelegue D, Yu L, et al. Characterization of a recombinant plant monoclonal secretory antibody and preventive immunotherapy in humans. Nat Med. 1998;4:601–6.
Kruger C, Hultberg A, van Dollenweerd C, Marcotte H, Hammarstrom L. Passive immunization by lactobacilli expressing single-chain antibodies against Streptococcus mutans. Mol Biotechnol. 2005;31:221–31. This paper presents a novel method for passive immunization against S. mutans infection.
Acknowledgments
The authors’ research in this area was supported in part by research R03-TW-06324 (R.O.MG.), R90DE022529 (M.I.K.), R37 DE06153 (D.J.S.), grants from the National Institute for Dental and Craniofacial Research (NIDCR/NIH). R.O.MG also received support from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, proc. 07/56100-2 and 09/54182-7). The authors declare no conflicts of interest with respect to the authorship and/or publication of this article.
Compliance with Ethics Guidelines
ᅟ
Conflict of Interest
Dr. Renata O. Mattos-Graner declares no potential conflicts of interest relevant to this article.
Dr. Marlise I. Klein has received a grant and support for travel to meetings from NIDCR/NIH.
Dr. Daniel J. Smith has received a grant and support for travel to meetings from NIDCR.
Human and Animal Rights and Informed Consent
This article does not contain any studies with human or animal subjects performed by any of the authors.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Mattos-Graner, R.O., Klein, M.I. & Smith, D.J. Lessons Learned from Clinical Studies: Roles of Mutans Streptococci in the Pathogenesis of Dental Caries. Curr Oral Health Rep 1, 70–78 (2014). https://doi.org/10.1007/s40496-013-0008-1
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40496-013-0008-1