
Abstract
Well-preserved external moulds of articulated brittle stars from the middle to late Cenomanian (early–Late Cretaceous) “Lower formation” of the Mifune Group on the island of Kumamoto, southern Japan, are described as a new species of the genus Stegophiura, S. miyazakii. Extinct (Late Cretaceous) species previously assigned to Stegophiura are now shown to represent other genera; here, we transfer S.? hagenowi and S.? nekvasilovae to the extant ophiopezid genus Ophiopeza and S.? trispinosa to the extinct ophiacanthid Sabinacantha. The present specimens are thus inferred to be the sole wholly extinct representative and the oldest record of the genus Stegophiura.
Similar content being viewed by others


Introduction
Our knowledge of ophiuroid systematics and evolution is currently increasing at an impressive rate thanks to concerted advances in molecular and morphological phylogenies (e.g., O’Hara et al. 2014, 2017, 2018; Thuy and Stöhr 2016) and to an ongoing systematic survey of the fossil record of brittle stars (e.g., Hess 1975; Jagt 2000; Thuy 2013; Thuy and Stöhr 2018). Despite this, fossil ophiuroids are still poorly known in comparison with their Recent relatives (e.g., Stöhr et al. 2012). However, in view of their pivotal role in reconstructing and dating phylogenies (e.g., O’Hara et al. 2014; Thuy and Stöhr 2016), extinct brittle stars are a valuable sources of data. Therefore, to explore the evolutionary history of the Ophiuroidea in greater detail, attempts should be made to assess their fossil record as exhaustively as possible.
Here, we describe new finds of ophiuroids from the lower Upper Cretaceous of southern Japan and identify these as a new species that is assignable to the extant ophiopyrgid genus Stegophiura H. Matsumoto, 1915. To date, 16 living species of this genus have been recorded worldwide (Stöhr et al. 2018). Fossil forms previously assigned to this genus are all Late Cretaceous in age and include S.? hagenowi (Rasmussen, 1950) from the lower Campanian to upper Maastrichtian of England, Denmark, Belgium, Germany, France and Poland (Jagt 2000; Jagt and Odin 2001), S.? nekvasilovae Štorc and Žítt, 2008 from the upper Turonian of the Bohemian Basin (Czech Republic), and S.? trispinosa Jagt and Kutscher, in Jagt, 2000 from the upper Campanian of Belgium. They are known exclusively from dissociated lateral arm plates and/or radial shields, except for S.? hagenowi which is also known from arm fragments and other skeletal plates; there is also a more or less articulated specimen with arms from the upper Maastrichtian of the Maastricht area (the Netherlands; J. W. M. Jagt, pers. comm., June 2018). Fully articulated specimens of the extant species, S. sterea (H. L. Clark, 1908), were recorded from the Pliocene of central Japan by Ishida et al. (1996).
The present study aims to provide a detailed morphological description of the new finds from southern Japan and a systematic interpretation with respect to previously recorded fossil occurrences of Stegophiura.
Material and geological setting
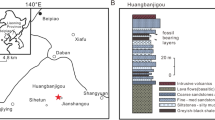
The two individuals described herein were collected from siltstone of Matsumoto’s (1939) “Lower formation” of the Cretaceous Mifune Group. These strata are exposed along the Akai River at Kawauchida (Mashiki Town, Kumamoto Prefecture), approximately 3.5 km northeast of Funano Mountain (co-ordinates 32°46′44″N, 130°52′2″E; see Fig. 1) (Matsumoto 1939; Tamura 1979). The specimens expose the ventral and dorsal sides, respectively (Fig. 2a), and are preserved as external moulds that lackall original skeletal calcite. To enable a detailed morphological assessment, a synthetic resin cast of the specimens was made.

Map showing the position of the locality (marked by X) that has yielded the new Stegophiura specimens described herein. The map on the left is part of the “Mifune” 1:25,000 topographic map of the Geographical Survey Institute

Synthetic resin casts of Stegophiura miyazakii sp. nov. from the “Lower formation” of the Mifune Group (middle-to-late Cenomanian; Upper Cretaceous). a Two partly overlapping individuals showing dorsal (holotype; MDM 15017) and ventral (paratype: MDM 15018) sides of the disc. b Holotype, dorsal view of disc and basal arm portions. c Paratype, ventral view of disc and basal arm portions. d Holotype, dorsal view of basal arm segments. e Holotype, dorso-lateral view of basal arm segments. Acpa, arm comb papilla; Agpl, abradial genital plate (arm comb plate); Aos, adoral shield; Apa, apical papilla; Asa, arm spine articulation; Asp1, primary arm spine; Asp2, secondary arm spine; Dap, dorsal arm plate; Dsc, disc scale; Dsp, disc spine; Gsc, genital scale; Lap, lateral arm plate; Opa, Oral papilla, Opl, oral plate; Osh, oral shield; Rsh, radial shield; Otp2, second oral tentacle pore; Spu, Spur; Tpo, tentacle pore; Tsc, tentacle scale; Vap, ventral arm plate. All scale bars equal 1 mm
The “Lower formation” of the Mifune Group is a succession of mainly sandstone and siltstone beds, including several coal seams and attains an overall thickness of 450–700 m (Matsumoto 1939). This unit has yielded the ammonite Eucalycoceras cf. spathi (Collignon) and the bivalve Actinoceramus tamurai (Matsumoto and Noda); both taxa are indicative of a middle-to-late Cenomanian age (Tamura and Matsumura 1974; Matsumoto et al. 1982, 1991; Saito et al. 2005).
The “Lower formation” comprises brackish to shallow-marine deposits and yields numerous bivalves such as Tetoria inflata, Nipponicorbula mifunensis, Crassostrea japonica, Matsumotoa unisulcata, Brachidontes mashikensis, Anomia foldia, Ceratostreon japonica, Eomiodon matsubasensis, and Pulsides okadai (Tamura 1979). It overlies the “Basal formation” that consists mainly of basal conglomerates and is overlain by terrestrial deposits of the “Upper formation” (Matsumoto 1939) which have yielded vertebrate fossils, including dinosaur bones and teeth (Tamura et al. 1991).
Specimens of the Recent Stegophiura sterea contained in the collections of the National Museum of Nature and Science, Tokyo (NSMT E-9136) were used for morphological comparisons with the fossil material.
Systematic palaeontology (section by YI, BT, and LDN-T)
Order Ophiurida Müller and Troschel, 1840
Suborder Ophiurina Müller and Troschel, 1840
Family Ophiopyrgidae Perrier, 1893
Genus Stegophiura H. Matsumoto, 1915
Stegophiura miyazakii sp. nov. (Figs. 2, 3)

Diagnosis Species of Stegophiura with tiny spines fringing the edges of dorsal and ventral disc sides (Figs. 2b, c, 3a–c) and long primary arm spines that correspond in length to arm segments (Figs. 2d, 3c).
Derivation of name Named in honour of Hayao Miyazaki, co-founder of Studio Ghibli, an animation studio in Japan, to pay tribute to his anime lifework, in particular the masterpiece “Ponyo” that celebrates marine biodiversity. In addition, Hayao Miyazaki`s favourite novelist, Soseki Natsume, lived in Kumamoto Prefecture, where the new ophiuroid fossils were found.
Holotype MDM 15017 (Mifune Dinosaur Museum, Kumamoto, Japan), an articulated skeleton exposing the dorsal side.
Paratype MDM 15018 (Mifune Dinosaur Museum, Kumamoto, Japan), an articulated skeleton exposing the ventral side.
Type locality and stratum Siltstone within the “Lower formation” of the Mifune Group (middle-to-upper Cenomanian, Upper Cretaceous) at Kawauchida (Mashiki Town, Kumamoto Prefecture), southern Japan (Fig. 1) (Matsumoto 1939).
Measurements Holotype: disc diameter, 6.1 mm, maximum preserved arm length, 10.0 mm, basal arm width, 1.5 mm. Paratype: disc diameter, 5.0 mm, maximum preserved arm length, 8.7 mm, and basal arm width, 1.1 mm.
Description
Of the holotype, the dorsal side is exposed (Figs. 2a, b, d, e, 3c, d). The disc is circular in outline, being covered by moderately large, oval or elongate, and thick scales, that either abut or overlap, with a row of tiny, contiguous papilla-like spines fringing the disc edge (Figs. 2b, 3c). Radial shields are quadrangular with rounded angles, being separated from the disc edge by very large abradial genital plates and in contact along their straight perradial edge. Abradial genital plates are fully exposed on the dorsal side of the disc and are conspicuous, large, oval, and longer than wide, being separated from each other by a row of two or three plates that are assumed to correspond to dorsal arm plates (Figs. 2b, 3c). The distal edge of the abradial genital plate bears a row of tiny, squarish papillae, in contact with each other and forming an arm comb. Arms are robust, taller than wide at the base. Dorsal arm plates are trapezoidal, wider than long in proximal segments and of equal width and length in median segments, widely separating the lateral arm plates on all observable arm segments (Fig. 2d, e). Lateral arm plates are swollen, with moderately coarse outer surface stereom and with at least four small, well-defined, prominent spurs grouped in the centre of the outer proximal edge (Fig. 2d). The spine articulations are integrated into the stereom at the distal edge of the lateral arm plates and consist of large muscle openings separated from a much smaller nerve opening by a low vertical ridge (Figs. 2e, 3d). The lateral arm plates carry a single, large, slender arm spine that is of equal length to or a little shorter than an arm segment; the spine is located in the lower part of the lateral arm plates, under a small angle to the arm (Figs. 2d, 3c). In addition to the single arm spine, lateral arm plates carry at least seven very short, squarish, contiguous, addressed papillae (Fig. 3c).
The paratype exposes the ventral disc side (Figs. 2a, c, 3a, b). The outline of the disc is circular, being covered by moderately large oval or elongate scales, abutting or overlapping, with a row of tiny, contiguous, papilla-like spines fringing the disc edge (Figs. 2c, 3a). The oral plates are slender and carry at least four squarish, contiguous lateral oral papillae. There is at least one short, conical apical papilla (Figs. 2c, 3a). Adoral shields are elongate, slender proximally, wider distally and in contact proximally (Figs. 2c, 3a). Oral shields are small, with a right to slightly obtuse proximal angle, slightly notched lateral edges and a rounded distal edge (Figs. 2c, 3a). Genital scales are small, squarish, and contiguous. Genital slits are narrow, extending from the oral shield to the disc edge. Second oral tentacle pores open outside the mouth slit and are bordered by small, squarish, and contiguous papillae similar to the lateral oral papillae, but forming a separate row. Successive tentacle pores are large, obliquely elongate, almost completely encompassed by consecutive lateral arm plates, with at least seven small, contiguous, and squarish tentacle scales on both edges of the pores in proximal arm segments (Figs. 2c, 3a). Lateral arm plates are narrow (Figs. 2c, 3a, b). Ventral arm plates are trapezoidal/pentagonal, widest distally, in contact on all observable arm segments, wider than long in proximal arm segments and as long as wide in median ones; the first ventral arm plates are trapezoidal, of comparable length and width (Figs. 2c, 3a, b).
Discussion
The Mifune specimens are here assigned to the genus Stegophiura on account of the thick plates and scales that cover the disc, the stout and relatively tall arms, the contiguous rectangular oral papillae, the second oral tentacle pores opening outside the mouth slit and being bordered by papillae similar to the lateral oral papillae, but forming a separate row, the large tentacle pores that are almost completely encompassed by successive lateral arm plates and bordered by numerous scales, the wide, contiguous dorsal and ventral arm plates, the swollen lateral arm plates, and the two types of arm spines. Features in common with the Recent Stegophiura sterea (Fig. 4), with emphasis on the unusual shape and position of the radial shields, also corroborate placement in this genus. However, it should be noted that Stegophiura ranks amongst ophiopyrgid genera that probably represent a polyphyletic complex of species in urgent need of a thorough re-evaluation. Therefore, our present assignment of these specimens to Stegophiura is tentative at best.

Recent Stegophiura sterea, NSMT E-9136. a, b Dorsal view. c ventral view. d–f Lateral view. a–c and d–f are same specimens, respectively. a Whole view. b, c Disc and basal arm portions. d, e Basal arm segments. f Lateral arm plate (median portion) by SEM (see Fig. 2 for abbreviations). Scale bars of a–e and f equal 1 mm and 0.1 mm, respectively
The Mifune specimens differ from extant species of Stegophiura (Stöhr et al. 2018), viz. S. brachyactis (H. L. Clark, 1911), S. elevata (Lyman, 1878), S. hainanensis Liao, in Liao and Clark, 1995, S. macrarthra H. L. Clark, 1915, S. nodosa (Lütken, 1855), S. ponderosa (Lyman, 1878), S. rhabdotoplax Murakami, 1942, S. singletoni McKnight, 1975, S. sladeni (Duncan, 1879), S. sterea, S. sterilis Koehler, 1922, S. striata (Duncan, 1879), S. stuwitzii (Lütken, 1857), S. vivipara H. Matsumoto, 1915 and S. wilhelmi Manso, 2010, in having tiny spines that fringe the disc edge (Figs. 2b, 3c) and long, slender primary arm spines (Figs. 2d, 3c). Stegophiura carinata Djakonov, 1954 has primary arm spines that are as long as the corresponding arm segments in basal arm portions, similar to those of the present Mifune specimens. However, it differs form the latter in having keeled ventral arm plates and long oral shields (Djakonov 1954) and in lacking the fringing disc spines.
Three extinct (Late Cretaceous) species have previously been tentatively assigned to Stegophiura but, in the light of recent progress in ophiuroid morphology and systematics, these should now be critically revised. Stegophiura? hagenowi from the lower Campanian to upper Maastrichtian of England, Denmark, France, Belgium, Germany, and Poland (Jagt 2000; Jagt and Odin 2001) is known from articulated arm fragments, partially articulated discs (J.W.M. Jagt, pers. comm., June 2018) and dissociated lateral, ventral and dorsal arm plates and radial shields with a conspicuous outer surface stereom consisting of coarse knobs surrounded by smaller tubercles and with small spine articulations comprised of curved dorsal and ventral lobes. The latter feature precludes assignment to the genus Stegophiura and even to the superorder Euryophiurida for that matter. In combination with the conspicuous outer surface ornamentation, the structure of spine articulations suggests transfer to the extant ophiopezid genus Ophiopeza Peters, 1851 within the superfamily Ophiodermatoidea. The same holds true for Stegophiura? nekvasilovae from the upper Turonian of the Czech Republic. Stegophiura? trispinosa from the upper Campanian of Belgium reveals the greatest similarities to the extinct ophiacanthid Sabinacantha archetypa Thuy, 2013 from the Oxfordian (Upper Jurassic) of Germany with respect to overall shape of the lateral arm plates, their outer surface ornamentation and the shape and position of the arm spine articulations, thus suggesting transfer to Sabinacantha.
The individuals described herein differ from all currently known species of Stegophiura and we, therefore, introduce a new species, S. miyazakii, to accommodate them. The only other confirmed fossil occurrence of Stegophiura to date involves articulated specimens identified as S. sterea from the Pliocene of central Japan (Ishida et al. 1996). Stegophiura miyazakii sp. nov. thus represents the first truly extinct species of the genus and significantly extends its stratigraphical range into the lower Upper Cretaceous.
Based on the associated molluscan fossils (Tamura 1979), Stegophiura miyazakii sp. nov. probably lived in shallow-marine environments with brackish influence. This is in stark contrast to the distribution of Recent relatives that mainly inhabit outer shelf to bathyal environments (Irimura et al. 1995).
References
Clark, H. L. (1908). Some Japanese and East Indian echinoderms. Bulletin of the Museum of Comparative Zoology, 51(11), 279–311.
Clark, H. L. (1911). North Pacific ophiurans in the collection of the United States National Museum. Smithsonian Institution, United States National Museum Bulletin, 75, 1–302.
Clark, H. L. (1915). Catalogue of Recent ophiurans. Memoirs of the Museum of Comparative Zoology, 25(4), 163–376. (20 pls).
Djakonov, A. M. (1954). Ophiuroids of the USSR seas. Izdatel’stvo Akademii Nauk SSSR, Moskva-Leningrad, 55, 1–136. (1 pl).
Duncan, P. M. (1879). On some Ophiuroidea from the Korean Seas. Journal of the Linnean Society, 14(77), 445–482.
Hess, H. (1975). Mikropaläontologische Untersuchungen an Ophiuren VI: Die Ophiuren aus den Günsberg-Schichten (oberes Oxford) vom Guldental (Kt. Solothurn). Eclogae Geologicae Helvetiae, 68, 591–601.
Irimura, S., Kubodera, T., & Ishida, Y. (1995). Catalogue of specimens of the Class Ophiuroidea (Echinodermata) donated by Dr. Seiichi Irimura in the National Science Museum, Tokyo (p. 181). Tokyo: National Science Museum.
Ishida, Y., Tanabe, T., Ito, T., & Hachiya, K. (1996). Fossil ophiuroids from the Plio-Pleistocene Hijikata Formation of the Kakegawa Group, Shizuoka, Central Japan. Bulletin of the National Science Museum, Tokyo, Series C, 22(3.4), 63–89.
Jagt, J. W. M. (2000). Late Cretaceous-Early Palaeocene echinoderms and the K/T boundary in the southeast Netherlands and the northeast Belgium—Part 3: Ophiuroids, with a Chapter on Early Maastrichtian ophiuroids from Rügen (Germany) and Møn (Denmark) by Manfred Kutscher and John W. M. Jagt. Scripta Geologica, 121, 1–179.
Jagt, J. W. M., & Odin, G. S. (2001). Campanian-Maastrichtian ophiuroids (Echinodermata) from Tercis les Bains (Landes, France). In G. S. Odin (Ed.), The Campanian-Maastrichtian stage boundary. Characterisation at Tercis les Bains (France) and correlation with Europe and other continents [Developments in Palaeontology and Stratigraphy, 19] (pp. 414–420). Amsterdam: Elsevier.
Koehler, R. (1922). Ophiurans of the Philippine Seas and adjacent waters. Smithsonian Institution, United States National Museum Bulletin, 100(5), 1–486.
Liao, Y., & Clark, A. M. (1995). The echinoderms of southern China (p. 614). Beijing: Science.
Lütken, C. F. (1855). Bidrag til kundskab om Slangestjernerne. II. Oversigt over de vestindiske Ophiurer. Videnskabelige Meddelelser fra det Dansk Naturhistorisk Förening i Kjøbenhavn, 1856(7), 1–19.
Lütken, C. F. (1857). Oversigt over Grönlands Echinodermata samt over denne Dyreklasses geographiske og bathymetriske Udbredningsforhold. Fakultet ved Kjöbenhavns Universitet (unpubl. doctoral thesis), pp. 1–109.
Lyman, T. (1878). Ophiuridae and Astrophytidae of the “Challenger”Expedition. Part I. Bulletin of the Museum of Comparative Zoology at Harvard College, 5(7), 65–168. (10 pls).
Manso, C. L. C. (2010). Deep-water Ophiuroidea (Echinodermata) from off Chile in the eastern South Pacific. Biota Neotropica, 10(2), 185–199.
Matsumoto, H. (1915). A new classification of the Ophiuroidea, with description of new genera and species. Proceedings of the Academy of Natural Sciences of Philadelphia, 68, 43–92.
Matsumoto, T. (1939). Geology of Mihune District, Kumamoto Prefecture, Kyushu (with special reference to the Cretaceous System). The Journal of the Geological Society of Japan, 46(544), 1–11. (in Japanese, with English abstract).
Matsumoto, T., Noda, M., & Maiya, S. (1991). Towards an integrated ammonoid-, inoceramid- and foraminiferal biostratigraphy of the Cenomanian and Turonian (Cretaceous) in Hokkaido. Journal of Geography, 100(3), 378–398. (in Japanese, with English abstract).
Matsumoto, T., Obata, I., Tashiro, M., Ohta, Y., Tamura, H., Matsukawa, M., et al. (1982). Correlation of marine and non-marine formations in the Cretaceous of Japan. Fossils, 31, 1–26. (in Japanese, with English abstract).
McKnight, D. G. (1975). Some echinoderms from the northern Tasman Sea. New Zealand Oceanographic Institute Records, 2(5), 49–76.
Müller, J., & Troschel, F. H. (1840). Über die Gattungen der Ophiuren. Archiv für Naturgeschichte, 6, 327–330.
Murakami, S. (1942). Ophiurans of Izu, Japan. Journal of the Department of Agriculture, Kyushu Imperial University, 7(1), 1–36.
O’Hara, T. D., Hugall, A. F., Thuy, B., & Moussalli, A. (2014). Phylogenomic resolution of the Class Ophiuroidea unlocks a global microfossil record. Current Biology, 24, 1874–1879.
O’Hara, T. D., Hugall, A. F., Thuy, B., Stöhr, S., & Martynov, A. V. (2017). Restructuring higher taxonomy using broad-scale phylogenomics: The living Ophiuroidea. Molecular Phylogenetics and Evolution, 107, 415–430.
O’Hara, T. D., Stöhr, S., Hugall, A. F., Thuy, B., & Martynov, A. V. (2018). Morphological diagnoses of higher taxa in Ophiuroidea (Echinodermata) in support of a new classification. European Journal of Taxonomy, 416, 1–35.
Perrier, E. (1893). Traité de Zoologie, 1(2) (p. 864). Paris: Masson.
Peters, W. (1851). Übersicht der an der Küste von Mossambique eingesammelten Ophiuren, unter denen sich zwei neue Gattungen befinden. Bericht über die zur Bekanntmachung geeigneten Verhandlungen der Königlich Preussischen Akademie der Wissenschaften zu Berlin, 1851, 463–466.
Rasmussen, H. W. (1950). Cretaceous Asteroidea and Ophiuroidea with special reference to the species found in Denmark. Danmarks Geologiske Undersøgelse, 77, 1–134.
Saito, M., Miyazaki, K., Toshimitsu, S., & Hoshizumi, H. (2005). Geology of the Tomochi district. Quadrangle Series, 1:50,000, Geological Survey of Japan (p. 218). Tokyo: AIST.
Stöhr, S., O’Hara, T. D., & Thuy, B. (2012). Global diversity of brittle stars (Echinodermata: Ophiuroidea). PLoS One, 7(3), e31940.
Stöhr, S., O’Hara, T. D., & Thuy, B. (eds) (2018). World Ophiuroidea database. http://www.marinespecies.org/ophiuroidea. Accessed 20 Feb 2018
Štorc, R., & Žítt, J. (2008). Late Turonian ophiuroids (Echinodermata) from the Bohemian Cretaceous Basin, Czech Republic. Bulletin of Geosciences, 83(2), 123–140.
Tamura, M. (1979). Cenomanian bivalves from the Mifune Group, Japan, Part 3. Memoirs of the Faculty of Education, Kumamoto University, Section 1, Natural Science, 28, 59–74.
Tamura, M., & Matsumura, M. (1974). On the age of the Mifune Group, central Kyushu, Japan. Memoirs of the Faculty of Education, Kumamoto University, Section 1, Natural Science, 23, 47–56.
Tamura, M., Okazaki, Y., & Ikegami, N. (1991). Occurrence of Cenomanian and herbivorous dinosaurs from Upper Formation of Mifune Group, Japan. Memoirs of the Faculty of Education, Kumamoto University, Natural Science, 40, 31–45. (in Japanese, with English abstract).
Thuy, B. (2013). Temporary expansion to shelf depths rather than an onshore-offshore trend: The shallow-water rise and demise of the modern deep-sea brittle star family Ophiacanthidae (Echinodermata: Ophiuroidea). European Journal of Taxonomy, 48, 1–242.
Thuy, B., & Stöhr, S. (2016). A new morphological phylogeny of the Ophiuroidea (Echinodermata) accords with molecular evidence and renders microfossils accessible for cladistics. PLoS One, 11(5), e0156140.
Thuy, B., & Stöhr, S. (2018). Unravelling the origin of the basket stars and their allies (Echinodermata, Ophiuroidea, Euryalida). Scientific Reports, 8, 8493.
Acknowledgements
We express deep gratitude to the late Hans Hess (Natural History Museum, Basel, Switzerland) for providing many references and useful comments on the identification of extinct ophiuroids and, more generally, for his outstanding contributions to ophiuroid palaeontology. We also thank Kyoichi Iwanaga for donating the fossils. Comments by John Jagt and an anonymous reviewer greatly improved an earlier version of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial Handling: C. A. Meyer.
Rights and permissions
About this article

Cite this article
Ishida, Y., Thuy, B., Fujita, T. et al. A new species of Stegophiura (Ophiuroidea, Ophiopyrgidae) from the mid-Cretaceous of southern Japan. Swiss J Palaeontol 137, 319–325 (2018). https://doi.org/10.1007/s13358-018-0168-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-018-0168-7