
Abstract
Cyrtocrinids are morphologically diverse crinoids of the Mesozoic, yet their origin and early evolution are still poorly understood. Here, we attempt to disentangle the early evolutionary history of the cyrtocrinids taking into account phylogenetic, palaeobiogeographic, and palaeoecological evidences. In addition, we describe new finds of cyrtocrinids from an upper Pliensbachian to lower Aalenian rock reef section of Feuguerolles, Calvados, France. The material includes the new species Tetracrinus solidus n. sp. and Sacariacrinus amadei n. sp. The common Cotylederma docens, Eudesicrinus mayalis, and Praetetracrinus inornatus are represented by ossicles of all growth stages, complementing previous descriptions. A single ossicle is ascribed to Cotylederma miliaris. Columnals of P. inornatus and primibrachials of C. docens bear yet unknown ichnological traces. In terms of faunal spectrum, the Feuguerolles assemblage is more closely related to Lower Jurassic faunas of northeastern Europe than to Tethyan faunas from Switzerland, Italy and Turkey. Finally, several lines of evidence suggest that the cyrtocrinids originated in the deep sea and invaded the shelf seas in the course of the Early Jurassic.
Similar content being viewed by others

Introduction
Starting from near zero at the end of the Permian crinoids diversified during the Triassic into several orders, creating the base of the Subclass Articulata (Hagdorn 2011). According to Simms (1990), “crinoid faunas between the early Carnian and Early Jurassic experienced a considerable change, but the actual record for the intervening interval is very poor, being distorted by both geographic and stratigraphic preservational and collecting bias”. This is particularly true for the cyrtocrinid crinoids. In fact, cyrtocrinids suddenly appeared in the fossil record as an already highly diverse group in the Tethys Ocean of the Mediterranean area during the Early Jurassic (Hess 2006). These first forms included odd-shaped species, small flower-like phyllocrinids with tiny arms, taller forms with radials prolonged stalk-like, species with well-developed column, crown symmetries varying from four to six rays and forms with cups comprised of differently sized radials (Hess in Hess and Messing 2011). The exceptionally diverse morphological spectrum of the early cyrtocrinids together with their apparently sudden emergence makes them an intriguing group to study.
An extraordinarily rich collection of previously undescribed cyrtocrinid remains from the Early and Middle Jurassic of Normandy, France, provides an excellent opportunity to re-assess the early evolution of the cyrtocrinids. The new material, although largely disarticulated, offers additional information about the species previously described from the nearby outcrop of May-sur-Orne (Loriol 1882–1889), which Hess (2006) compared with the slightly older fauna from Arzo, Switzerland.
In the present paper, we describe the new cyrtocrinid material, introducing two new species. Furthermore, we conduct a phylogenetic analysis of cyrtocrinids using morphological characters and reconstruct palaeobiogeographic patterns of their initial diversification during the Early Jurassic. Finally, based on the currently available lines of evidence, we attempt to reconstruct the origin and early evolution of the cyrtocrinids.
Geological context
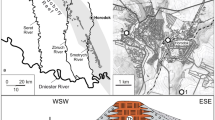
The material described herein was collected in the now abandoned and largely re-filled Guérin quarry on the left bank of the river Orne near Feuguerolles, southwest of Caen, Calvados Department in Normandy, France (Hess and Thuy 2016, fig. 1). The Ordovician Armorican quartzites, which were the target of the quarrying activities, are unconformally overlain by a highly condensed succession of Lower to Middle Jurassic marls and limestones. The oldest Jurassic sediments represent transgressive infillings of the eroded surface of the tilted quartzites which, at the time of deposition, formed a near-shore rock reefs with numerous decimetre- to metre-scale cavities and depressions (Rioult 1968). This particular depositional setting is informally referred to as “rock reef”, because the relief and internal structural complexity set it apart from other hardgrounds, and because it was not sufficiently close to the shoreline to be considered a rocky shore. These infillings are overlain by a succession of thin marl and limestone beds, which had considerable, small-scale spatial variation in thickness and lateral extent as a result of the complex palaeo-topography.
In spite of their obvious interest as deposits of an exceptional palaeo-environmental setting, regrettably few studies have dealt with the Lower and Middle Jurassic sediments of Feuguerolles (Rioult 1968; Dugué et al. 1998; Vernhet et al. 2002; Weis et al. 2015; Hess and Thuy 2016). As outlined by Hess and Thuy (2016), the chronozone-level (sensu Page 2003) stratigraphic subdivision of the Jurassic Feuguerolles succession is reasonably robust. The transgressive infillings within and on top of the Paleozoic quartzite cavities are mostly dated to the late Pliensbachian Margaritatus Chronozone, although some are slightly older (early Pliensbachian Ibex and Davoei Chronozones). Evidence for the Spinatum Chronozone is sparse and, as yet, insufficiently documented. The overlying succession of marls and limestones yielded ammonite and thecid evidence for the early Toarcian Tenuicostatum Chronozone (Semicelatum Subchronozone), Serpentinum Chronozone (Exaratum and Falciferum Subchronozones), and Bifrons, Variabilis, Thouarsense, and Aalensis Chronozones (no data on Subchronozones yet). The youngest crinoid material described herein was recovered from sediments dated to the Aalenian Opalinum Chronozone.
Materials and methods
The present paper deals with the cyrtocrinids of a crinoid fauna collected by Marc Chesnier at the Guérin quarry of Feuguerolles, southwest of Caen, Calvados Department (Normandy). A previous paper was devoted to the rare but diverse comatulids and pentacrinines (Hess and Thuy 2016). Together with the isocrinid Isocrinus (Chladocrinus) basaltiformis (Miller) and the millericrinid Amaltheocrinus amalthei (Quenstedt), the cyrtocrinids are the dominant crinoids. A. amalthei is represented at Feuguerolles by attachment structures, columnals, cup plates (radials, primibrachials), and secundibrachials, not treated in the present paper (for descriptions, see Quenstedt 1876; Jäger 1993). Remains of I. basaltiformis are not described either (for descriptions see Loriol 1886 in 1882–1889; Quenstedt 1876; Simms Simms 1989; Jäger 1985).
Among cyrtocrinids, cup plates and brachials of a new species of Sacariacrinus and an intact cup of a new species of Tetracrinus are of particular interest. Cotylederma docens Deslongchamps is well-represented by all ossicle types, with many primibrachials bearing ichnological traces. The remains of Eudesicrinus mayalis (Deslongchamps) comprise basal elements, partly with attached radial circlet including cups of different sizes, isolated radials, and secundibrachials. Also noteworthy are numerous ossicles, in particular columnals, basal circlets, isolated radials and primibrachials, of Praetetracrinus inornatus (Simms), previously known only from England and Germany; attachment disks assignable to this species are rare. The new species of Sacariacrinus is known from only three samples (357, 425, and 540) by basal circlets, radial circlets, isolated radials, first and second primibrachials, and secundibrachials. The new Tetracrinus species is represented by a single cup.
For the stratigraphic range of the different species at Feuguerolles including the cyrtocrinids, see Hess and Thuy (2016).
Crinoid remains were picked from the residues of 220 individual washed bulk samples taken bed by bed in various sections of the Guérin quarry in Feuguerolles. The samples vary widely in number of crinoid plates, and the remains are poorly sorted in most cases as demonstrated by the presence of all sizes, including juvenile specimens. Crinoids are highly disarticulated but mostly well preserved. Selected specimens were mounted on aluminium stubs and gold-coated for scanning electron microscopy using a Jeol Neoscope JCM-5000. All specimens figured and/or described herein are housed at the Natural History Museum of Le Mans (France) (collection acronym MHNLM).
Cyrtocrinid origins—state of knowledge (Fig. 1)
Unequivocal Cyrtocrinida have not been identified in Triassic sediments. One might expect remains of these crinoids in platform sediments of the Late Triassic (upper Ladinian/lower Carnian) reefs of the Cassian Beds, but none have been identified despite extensive collecting for nearly 180 years (e.g., Zardini 1976, 1988). Salamon et al. (2009) ascribed small, probably juvenile, columnals from the Rhaetian of the Tatra Mountains to cyrtocrinids. The specimens have facets with thick, radiating crenulae and a rather wide lumen. Similar facets are found among encrinids, e.g., Dadocrinus kunischi Wachsmuth and Springer [see Hagdorn (1996, pl. 2, fig. d, e, g) and Hess in Hess and Messing (2011, fig. 14)]. Therefore, assignment of these Rhaetian ossicles to cyrtocrinids is doubtful. This is particularly relevant as Cohen and Pisera (2015) used the putative first appearance of cyrtocrinid ossicles in Rhaetian strata described by Salamon et al. (2009) to calibrate the timing of living crinoid diversification, which they estimate at 258 Mya.

First appearance and range of selected genera of Cyrtocrinida. Species of a given genus are linked by solid lines; dashed lines indicate inferred generic relationships. Data according to Hess (2006), Hess in Hess and Messing (2011), and Hess (2012). Castaneacrinus and Fusicrinus are known from brachials only. Occurrence of Cyathidium in the Tithonian according to Sieverts-Doreck (1951)
Thus, the origin of the Cyrtocrinida remains unresolved. Based on cylindrical columns and attachment by a disk, possible ancestors have been sought among the Millericrinida or Encrinida. Molecular phylogenetic data indicate that the extant Cyrtocrinida and Hyocrinida are sister-group clades (Rouse et al. 2013; Hemery et al. 2013). Hyocrinida were treated by Hess (in Hess and Messing 2011) as a separate order without a known fossil record. In contrast, Roux (1980), followed by Manni and Nicosia (1990) and Améziane et al. (1999), placed the Hyocrinidae in the Cyrtocrinida based on a comparison of columnal facets and their microstructure with those of Cyclocrinus amalthei and Cyclocrinus hausmanni. However, the latter species are now placed in Amaltheocrinus and, thus, in Millericrinida (Hess in Hess and Messing 2011). Hyocrinids attach to rocky substrates by an encrusting disk and mostly live in cold, deep water, where food supply is limited. They have a delicate skeleton with heavily pinnulate arms exposed to the currents. This is in sharp contrast to cyrtocrinids that have developed extensive and at times intricate skeletons during the course of their history, perhaps as an antipredatory strategy (Hess 2014b).
In Sinemurian–Pliensbachian (Thuy et al. 2014), Late Pliensbachian (Hess 2006) and Early Toarcian (Manni and Nicosia 1990) sites of the western Tethys, diverse cyrtocrinids appeared rather suddenly. The Late Pliensbachian site of Arzo is the earliest with a diverse cyrtocrinid fauna (Hess 2006). The morphological spectrum is remarkable and includes (1) comparatively large forms attached by a basal element (Cotylederma inaequalis, Eudesicrinus mayalis), (2) forms with stalk-like elongated cups (Dinardocrinus), (3) stalked forms with a high cup and primibrachials of variable height (Sacariacrinus), (4) stalked forms with cryptic basals and a funnel shaped cup (Quenstedticrinus), and (5) stalked forms with ornamented columnals, cups and brachials (Bilecicrinus), similar to later Cyrtocrinus. These forms are accompanied by tiny cups of stalked Phyllocrinidae (Nerocrinus petri, Ticinocrinus coronatus) that still have basals.
Manni and Nicosia (1999) suggested that Nerocrinus (for which they created a separate family) was a “quite primitive forefather of the cyrtocrinids”. We consider Nerocrinus with three basals and Ticinocrinus with a circlet of five basals to be members of the Phyllocrinidae. The two genera share small articular facets, sunken between interradial processes; a synapomorphy common to Phyllocrinus species (Hess 2012).
Manni and Nicosia (1996) assumed the ancestor of cyrtocrinids to lie among the Dadocrinidae, comprising the genera Dadocrinus and Neodadocrinus that share high basals. In their classification, cyrtocrinids are members of the order Dadocrinida, subdivided into several suborders. However, Dadocrinus is considered to belong to the Encrinida (Hagdorn 1996), and Neodadocrinus was assigned to the Millericrinida, Family Neodadocrinidae, in the Revised Treatise (Hess in Hess and Messing 2011). Neodadocrinus is pentamerous; it has discrete, high basals, and thick radials with distinct facets to basals, as well as thick primibrachials. This species co-occurs at Arzo with numerous cyrtocrinids, including the superficially similar Sacariacrinus. It seems possible that ancestors of stalked cyrtocrinids are to be found among Neodadocrinidae. In fact, Kristan-Tollmann (1975; fig. 26, 3–5) described brachials remarkably similar to those of Sacariacrinus altineri from Arzo from the pelagic Hallstatt Limestones (upper Ladinian to lower Carnian) of the Taurus Mountains (Turkey) and assigned them to Entrochus tenuispinosus. Did Kristan-Tollmann discover the cradle of a precursor to cyrtocrinids?
Cyrtocrinid phylogeny (Fig. 2)
So far, the only comprehensive phylogenetic analysis of the cyrtocrinids was performed by Romano et al. (2016) who suggested a complete revision of cyrtocrinid taxonomy thus imparting a sweeping blow to existing classifications, including the one in the latest Treatise (Hess and Messing 2011), which is rooted in the previous work of several authors with intimate knowledge of the group (e.g., Loriol, Jaekel, Sieverts-Doreck, Arendt, Žítt; see references). The parsimony-based analysis by Romano et al. (2016) produced well-resolved trees, but suffers from a number of significant flaws: first, it includes data on recognised genera as well as informal based on a preliminary and insufficiently substantiated subdivision of Eugeniacrinites; second, the analysis includes 33 taxa and 39 characters, resulting in a very low character/taxon ratio; and third, and most important, the study fails to show support values for the trees, thus precluding any meaningful interpretation of the results.

Evolutionary tree of the cyrtocrinid crinoids based on Bayesian inference with taxa positioned with respect to their stratigraphic distribution (thick black lines: range of the actual species; thick grey lines: range of the genus). Numbers at nodes indicate posterior probabilities
We have undertaken a new cladistics analysis with an ingroup of 24 fossil and 3 extant species (Table 1). We focused on fossil forms encompassing a wide spectrum of morphologies and those with the most completely known morphologies. We preferred using nominal species, not genera, because of species-level differences in a given genus. We conducted analyses using a different outgroup taxon (Holocrinus beyrichi versus Dadocrinus kunischi) and either with or without Neodadocrinus brevis in line with the comments above on the potential cyrtocrinid ancestry.
We compiled a character matrix consisting of 51 characters (see “ESM Appendix”). Analyses were run with characters unweighted and unordered in the absence of evidence for a clear ontogenetic or size-related progression between character states. When character states could not be assessed due to a lack of data, the character was scored with a “?”. Inapplicable characters were scored with a “–”.
Bayesian inference analysis was performed using MrBayes (Huelsenbeck and Ronquist 2001) as an MCMC simulation with default parameters for morphological data following the approach of Thuy and Stöhr (2016). MrBayes uses a modified version of the Juke–Cantor model for morphological data as outlined by Lewis (2001) with variable character states from 2 to 10 (Wright and Hillis 2014). All character states were considered to have equal frequency, and prior probabilities were equal for all trees. We assumed that evolutionary rates varied between sites according to a discrete gamma distribution. Branch lengths were unconstrained. Average standard deviations of split frequencies stabilized at about 0.008 after 3 million generations (mgen), sampled every 1000 generations. The first 25% of the trees were discarded as burnin.
The resulting tree from Bayesian inference has a moderate amount of structure, although many relationships remain unresolved and/or poorly supported (Fig. 2). Tree topology has a well-supported and resolved a phyllocrinid–psalidocrinid clade and a eugeniacrinitid clade that surprisingly includes the sclerocrinid Pilocrinus moussoni. Thus, our phylogeny endorses grouping of these families on a higher systematic level. Another reasonably well-supported but poorly resolved clade unites the plicatocrinids and tetracrinids, again endorsing the superfamily-level grouping of these families. The sclerocrinids (except P. moussoni) and the holopodine families, however, seem to be more problematic. Most of the analysed members of these groups form a poorly supported clade with an internal topology failing to follow currently accepted superfamily- and family-level classification concepts (Hess and Messing 2011). Curiously, the sclerocrinids Neogymnocrinus richeri and Cyrtocrinus nutans have an unresolved position with respect to the above-mentioned clades.
The resulting phylogeny helps disentangle some of the cyrtocrinid morphologies, yet fails to fully reveal the synapomorphies needed to clarify the evolutionary history of the group. Possible main evolutionary driving forces in cyrtocrinids are the reduction of skeletal features and paedomorphosis as inferred for other echinoderm groups (e.g., Stöhr and Martynov 2016). Both factors can mask true synapomorphies and thus hamper morphology-based phylogenetic estimates (e.g., Thuy and Stöhr 2016; Thuy, personal observations), especially when the concerned skeletal features are scored at face value rather than against the background of their origin. In spite of these reservations, our analysis clearly indicates the basal split of main cyrtocrinid morphologies, and tip dating of the tree suggests that divergence of these morphologies must have occurred well before the late Sinemurian (i.e., the age of the oldest known unequivocal cyrtocrinids).
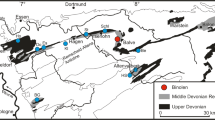
Palaeogeography of Lower Jurassic cyrtocrinid locations (Fig. 3)
The oldest known unequivocal cyrtocrinids are from the Sinemurian–Pliensbachian of northwestern Turkey and the Glasenbach section in Austria (Delogu and Nicosia 1986; Thuy et al. 2014). The cyrtocrinid localities in northwestern Turkey are situated on the Sakarya microcontinent (Sengör and Yilmaz 1981), a Mesozoic ‘island’ in a maze of younger zones of subduction–accretion formed between the deep-sea basins of the closing Palaeotethys to the north and the opening Neotethys to the south (Sengör 1979). The Glasenbach section, situated in the Austroalpine nappes of the Northern Calcareous (Eastern) Alps, is interpreted as a succession of hemipelagic/pelagic limestones and marls with numerous intercalations of slump deposits and calcareous turbidites derived from the bathyal slope of a submarine high on the former southern continental margin of the Alpine Tethys (Bernoulli and Jenkyns 1970; Böhm 2003; Thuy et al. 2014). Interestingly, both settings were within or adjacent to deep-sea basins during the Early Jurassic.

Early Jurassic palaeogeography with discussed sites (see Table 2)
In the course of the Pliensbachian, cyrtocrinids started spreading to various parts of the shelves around the northwestern tips of the Neotethys deep-sea basins, as documented by occurrences from the Apennines (IT), southern Germany, Normandy and southern England (see references in Table 2). This was probably facilitated by tectonic events in northwestern Europe during late Pliensbachian times that created connections with the Tethys as documented by ammonites (Rioult and Chirat 1999). All currently known Lower Jurassic cyrtocrinid localities are interpreted as current-swept settings with at least partially indurated substrate. Localities situated on the subsiding continental margins, farther away from the oceanic basin of the Neotethys, include non-volcanic seamounts and their slopes within the deep carbonate plateaus of the Umbrian Apennines (Tivoli, Gorgo a Cerbara) on the Adriatic microplate. These environments were characterised by low sedimentation rates and, in case of carbonate seamounts, by early submarine limestone cementation, local hardground formation and stratigraphic condensation (Manni and Nicosia 1996). Other Lower Jurassic cyrtocrinid shallow-sea localities at some distance from the deep basins include open shelf areas with low sedimentation rates (Sulzkirchen in southern Germany, West Bay in southern England) and near-shore rock reefs (Feuguerolles in Normandy).
Of particular, relevance to the question of cyrtocrinid origins is the locality of Arzo in southern Switzerland (Hess 2006). During the Early Jurassic, Arzo was situated on the Adriatic microplate that was to become the southern margin of the Alpine Tethys after rifting in late Norian and early Liassic times and after continental break-up during the Toarcian–Bajocian interval (Bernoulli and Jenkyns 2009, fig. 30). The Broccatello limestone underlying the cyrtocrinid-yielding strata in Arzo is interpreted as an ancient carbonate mudmound that developed at subphotic depths (Neuweiler and Bernoulli 2005; Hess 2006). It was lithified at an early stage and provided sheltered cavities at 100 or more metres water depth suitable for colonization by crinoids, whose remains accumulated in depressions of the seafloor. Strikingly, the Arzo cyrtocrinid assemblage is not only one of the deepest from the Early Jurassic but also by far the most diverse, and shares only a few species with coeval or slightly younger cyrtocrinid assemblages from the surrounding shelf settings (Hess 2006). Generally speaking, while many cyrtocrinid genera were widespread during the Early Jurassic, cyrtocrinid species seem to have been endemic to certain localities or regions within the same part of the shelf seas (e.g., Nerocrinus petri and Dinardocrinus tiburtinus in southern Switzerland and central Italy, Praetetracrinus inornatus in northwestern Europe), suggesting only limited dispersal across the shelf seas. In summary, the palaeogeographic evidence available to date converges to a pre-Sinemurian Tethyan deep-sea origin of the cyrtocrinids with a subsequent rise to shallow circum-Tethyan shelf seas in several regions.
Subsequent evolutionary history of cyrtocrinids
Early Jurassic crinoids without a stalk were attached to the substrate by an expanded aboral element (Cotyledermatidae: Cotylederma, Paracotylederma; Eudesicrinidae: Eudesicrinus); and such barnacle-like morphology with reduced arms covered by primibrachials in the non-feeding stage proved is also found in Cretaceous (Rasmussen 1961), Paleocene (Donovan and Jakobsen 2004) and extant successors (Wisshak et al. 2009; Syverson et al. 2015). Living Holopodina include two families, the Holopodidae (Cyathidium, Holopus) and the Eudesicrinidae (Proeudesicrinus). Flat trapezoidal secundibrachials without pinnule sockets apparently sealed the radial cavity in Cotylederma docens (see Fig. 6b–e). This is in contrast to fossil and extant species of Cyathidium that have reduced but pinnulate arms. Both at Arzo and at Feuguerolles, Cotylederma is an important part of the cyrtocrinid fauna. These sites offered habitats such as hardground cliffs and under overhangs or in caves, as in extant representatives of the group living in deep-shelf environments (Wisshak et al. 2009; Syverson et al. 2015). Donovan and Jakobsen (2004) described a Cyathidium–barnacle association from the Paleocene (Danian) of Denmark and questioned whether the tight closure of the crinoid was the result of antipredator behaviour only. In shallow water or intertidal environments, a watertight seal would have served crinoids such as C. docens. In deeper water, such as at Arzo, tight closure would have protected C. inaequalis from predators and unfavourable environmental conditions.
Thus, holopodine cyrtocrinids living at the lowest tier could seal their radial cavity against benthic predators. Nerocrinus and Ticinocrinus are stalked and very small, and their deep body cavity prevented easy access. Larger stalked cyrtocrinids collected food at higher tiers, away from benthic predators. Early Jurassic forms such as Sacariacrinus had cups and arms offering little protection against predators. Subsequent forms show various antipredator measures. Bilecicrinus and Arzocrinus had sturdy arms that may have carried spike-like pinnules, unfortunately not preserved. However, two non-cyrtocrinid forms from Arzo have quite spectacular antipredator morphology. They include Pustulocrinus iguana with crested pinnulars and Serracrinus planus with spiked brachials (Hess 2006).
Antipredator measures increased during the Middle Jurassic (Hess 2012). Some members of the Phyllocrinidae (Scutellacrinus, Phyllocrinus colloti, and P. voultensis) carried interradial extensions similar to Nerocrinus. Phyllocrinus fenestratus developed spike-like interradial extensions protecting the radial cavity that housed small arms. Lonchocrinus had cup and columnals comparable to the Oxfordian Eugeniacrinites, and soft parts were protected by spike-like primibrachials pointing outward and upward. The first species of the widespread genus Cyrtocrinus, C. praenutans, had arms suitable for inward-coiling, similar to those of the later Cyrtocrinus nutans and the living Neogymnocrinus richeri (Hess 2012, fig. 16). Presumably non-lethal epizoic pits on ossicles of C. praenutans indicate increased ecological interactions (Hess 2012, figs. 18–20).
During Oxfordian times sponge–algal reefs offered ideal habitats to cyrtocrinids. Eugeniacrinites cariophilites carried modified second primibrachials with the ability to form a roof over soft parts and inward-curled small arms. Nevertheless, this spectacular autapomorphy did not prevent predators, presumably fishes, from attacking the vital parts, leaving bite marks and causing malformations on the skeleton (Hess 2014b). In Plicatocrinus and Tetracrinus, arms were unsuitable for curling or folding together, but spike-like fused pinnules may have deterred predators (Hess in Hess and Messing, 2011, Fig. 95, 1f).
The Early Cretaceous (Valanginian) includes the famous Štramberk fauna with innumerable spoon-like and odd-shaped cups (e.g., Arendt 1974; Žítt 1974a, b, 1975, 1978a, b, 1979a, b, 1983). Well-known forms are stalked Phyllocrinidae (Phyllocrinus, Apsidocrinus), Psalidocrinidae (Psalidocrinus) and Sclerocrinidae (Sclerocrinus, Hemicrinus, Strambergocrinus, and Torynocrinus). In contrast, Hemibrachiocrinidae were fused to the substrate by an expanded base, similar to Cyathidium, and the small arms were folded together over the radial cavity (Hess in Hess and Messing 2011). Cyrtocrinids declined in disparity and numbers during the Cretaceous. The Cenozoic fossil record is exceedingly sparse; extant cyrtocrinids are of low diversity, environmentally, and bathymetrically restricted.
Ichnological traces (Figs. 4, 5)
A well-preserved parabolic pit on a radial of Cotylederma docens is assigned to Ooichnus paraboloides Bromley (Fig. 4; see Donovan 2017). The moderately deep pit is on the aboral, granular surface of the ossicle. Granules extend into the pit and are partly enlarged at the rim, indicating that infestation by the indeterminate epibiont was non-lethal. This radial is the only occurrence of such a pit in the examined material. Donovan and Tenny (2015) figured a pluricolumnal from the Mississippian with Ooichnus paraboloides pits. That specimen has concave pits flush with the surface on one side and raised swellings with pits on the other side. The authors assumed that the column was recumbent in life; pits atop cyst-like swellings indicate infestation on the crinoid during life. Flush pits on the other side show no growth reactions and were made after death. The pit on the C. docens specimen is flush with the surface, but granules in the pit suggest reaction growth either during or shortly after infestation. Ooichnus parabolides occur throughout the Phanerozoic and were caused by unknown invertebrates. Spectacular examples were described from the middle Jurassic of Ardèche (Hess 2012), where they found on 7.5% of cups and 3.6% of columnals of Cyrtocrinus praenutans. Some of these pits are associated with skeletal growth and swelling, but an elevated rim around the pit is exceptional.

Cotylederma docens, aboral view of radial with concave pit of Ooichnus paraboloides Bromley. The pit was probably caused by an unknown epibiont; MHNLM 2015.1.39, sample 350-1, early Toarcian (Serpentinum Chronozone), Feuguerolles (Normandy)

Cotylederma docens primibrachials with traces. a–e Cotylederma docens Deslongchamps. a Aboral (exterior) view of asymmetric second primibrachial with concentric folds; MHNLM 2015.1.40, sample 350-2, early Toarcian (Serpentinum Chronozone), Feuguerolles (Normandy). b Second primibrachial aboral, with folds and scratch marks; MHNLM 2015.1.41, sample 350-1, early Toarcian (Serpentinum Chronozone), Feuguerolles (Normandy). c First primibrachial in proximal–aboral view, note circular folds and scratch marks; MHNLM 2015.1.42, sample 368-1, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). d First primibrachial aboral, note concentric scratch marks; MHNLM 2015.1.43, sample 349-1, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy), photo Hess. e First primibrachial aboral, note scratch marks on hump; MHNLM 2015.1.44, sample 358-1, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy), photo Hess
Our material also includes nearly concentric marks on the aboral surface of primibrachials of Cotylederma docens, which to our knowledge have not been reported in the literature (S. K. Donovan, personal communication). On first primibrachials, the marks are mostly around the smooth central bulge and consist of smooth folds accompanied by scratch marks (Fig. 5c). Similar marks are found on a second primibrachial (Fig. 5b). A smaller ossicle of this type (Fig. 5a) has concentric folds around the centre, where some deeper scratch marks also occur. Deep scratches are on the bulge of first primibrachials (Fig. 5e) and on the sides of the bulge (Fig. 5d). Raised, smooth folds, suggest some repair growth. The origin of the concentric marks and the scratches is unknown. Multiple scratch marks are made by grazing chitons, as illustrated in, e.g., Kázmér and Tabaroši (2012). Another scenario could involve a parasitic or commensal gastropod, responsible for the concentric marks, subsequently targeted by fish leaving the scratch marks as collateral damage to the crinoid.
Scratch marks on columnals of Praetetracrinus inornatus are discussed and illustrated in the systematic section.
Conclusions
Cyrtocrinids appeared suddenly in the Tethys Ocean in the Sinemurian. Even the oldest known assemblages already include the highly distinctive variety of morphologies, ranging from stalkless forms attached by basal element (Holopodina) to stalked forms attached by a disk (Cyrtocrinina). By the end of the Early Jurassic, cyrtocrinids had spread to various shelf seas, especially in the northwestern Peritethys. The early cyrtocrinid fossil record is remarkable in several ways: (1) phylogenetic estimates suggest that the early radiation of the group took place well before the Sinemurian, predating the oldest known unequivocal cyrtocrinid fossils; (2) the putative ancestor of cyrtocrinids appears to be a neodadocrinid from Triassic pelagic deep-water settings of Turkey (Kristan-Tollmann 1975); (3) the oldest unequivocal cyrtocrinids are from bathyal slope settings (Glasenbach) and shelf settings near deep-sea slopes (northwestern Turkey); (4) the most diverse Lower Jurassic cyrtocrinid assemblage was from a bathyal setting (Arzo); and (5) from the late Pliensbachian onward, cyrtocrinids occur in various shelf settings but with fewer species than the Arzo assemblage and generally with species endemic to individual localities or regions.
These various lines of evidence suggest a Tethyan deep-sea origin of the cyrtocrinids followed by a successive rise to shallower depths from the Sinemurian onward and a spreading to suitable environments (current-swept hard substrates) in the shelf seas of the northern and northwestern Peritethys. This scenario is strikingly similar to the evolutionary history of the ophiacanthid ophiuroids that originated and diversified in the deep sea and temporarily invaded shelf environments in the course of the Jurassic as suggested by the fossil record of the group (Thuy 2013) and recently by molecular evidence (Bribiesca-Contreras et al. 2017). Thus, the cyrtocrinids add to the emerging evidence that deep-sea origination and range extension to shallower depths played a major role in shaping marine biodiversity and that the Early Jurassic was a time of enhanced faunal exchange along the bathymetric gradient.
Systematic palaeontology
Order Cyrtocrinida Sieverts-Doreck 1952.
Suborder Holopodina Arendt 1974.
Family Cotyledermatidae Wright 1876.
Cotylederma Quenstedt 1852.
Cotylederma docens Deslongchamps in Deslongchamps and Deslongchamps 1858, Figs. 4, 5, 6, 7.

Cotylederma docens aboral element, first primibrachials; Cotylederma miliaris, first primibrachial. a–e Cotylederma docens Deslongchamps. a Aboral element with attached radial, a1 lateral view, a2 adoral view with radial facet on top and first primibrachial in radial–aboral cavity; MHNLM 2015.1.45, sample 357-12, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). b Low radial, b1 adoral (interior), b2 distal; MHNLM 2015.1.46, sample 279-1, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). c High radial, adoral (interior) view; MHNLM 2015.1.47, sample 357-18, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). d First primibrachial, d1 aboral, d2 proximal with facet to radial; MHNLM 2015.1.48, sample 279-2, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). e Thick radial distal, note scratch marks; MHNLM 2015.1.49, sample 350-3, early Toarcian (Serpentinum Chronozone), Feuguerolles (Normandy). f Cotylederma miliaris Deslongchamps; first primibrachial, f1 adoral (interior), f2 proximal/aboral with granules, note small facet to adjoining ossicle and scratch marks, photo Hess; MHNLM 2015.1.50, sample 297, early Toarcian (Tenuicostatum/Serpentinum Chronozones), Feuguerolles (Normandy)

Cotylederma docens second primibrachial, secundibrachials. a–e Cotylederma docens Deslongchamps. a Asymmetric second sprimibrachial, a1 adoral (interior), a2 aboral (exterior) with concentric traces, magnified in Fig. 5a; MHNLM 2015.1.40, sample 350-2, early Toarcian (Serpentinum Chronozone), Feuguerolles (Normandy). b Thick secundibrachial adoral; MHNLM 2015.1.51, sample 357-23, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). c Thin, weakly trapezoidal secundibrachial, aboral with crenulated margin; MHNLM 2015.1.52, sample 259, early Toarcian (Serpentinum Chronozone), Feuguerolles (Normandy). d Trapezoidal secundibrachial adoral; MHNLM 2015.1.53, sample 357-22, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). e Trapezoidal secundibrachial, e1 adoral, e2 aboral; MHNLM 2015.1.54, sample 357-22, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy)
Material MHNLM 2015.1.39–2015.1.49 and 2015.1.51–2015.1.54. This species ranges from the upper Pliensbachian to upper Toarcian (Hess and Thuy 2016) and is represented by numerous disarticulated basal elements, radials, first and second primibrachials, and secundibrachials. Only in two specimens are radial circlets still attached to the basal element. Remains are especially common in the upper Pliensbachian (sample 355) and lower Toarcian (Tenuicostatum Chronozone, samples 267, 268, 354, 356–358, 425; and Serpentinum Chronozone, sample 350). Height of elements with similar diameter varies, and the largest basal elements reach a diameter of nearly 15 mm (samples 355–357). The species was thoroughly described by Loriol (1882) who noted the distinctive swollen first primibrachials. The present material includes primibrachials with peculiar concentric traces.
Diagnosis Aboral element thin-walled, hollow, of variable height and shape; upper margin with five weak depressions for radial circlet. Radial cavity wide and deep, extending into basal element. Radials symmetric, widening upward from narrow lower margin, joined by synostosis; articular facet with pair of small, deep, inward-sloping muscle fossae separated by a notch; ligament areas indistinct. First primibrachials trapezoidal in outline, often somewhat asymmetric; bulge on aboral side, leading to triangular profile in proximal or distal view; bulbous apex smooth and directed proximally; proximal and distal facets muscular, similar. Second primibrachials thick, trapezoidal. Secundibrachials thin, outline rectangular to skewed trapezoidal, food groove deep, articular facets muscular, no pinnule sockets.
Description The present material includes an aboral element with misformed cup of just one radial; the specimen’s first primibrachial (Fig. 6a) is in the body cavity, and on one side two budding extensions are visible that seem to be juvenile basal elements. Such elements were described by Rasmussen (1961, pl. 35, figs. 3, 4; as thecae) and Donovan and Jakobsen (2004, fig. 3a, c) from cups growing inside older cups of Cyathidium holopus from the Danian. Radials are narrow and vary in height (Fig. 5b, c, e); articular facets to first primibrachials have small aboral ligament fossae, flat interarticular ligament areas and small, circular and deep muscle fossae separated by a notch; and muscle fossae extend onto the adoral side and are rimmed. This morphology is mirrored in the proximal facet of the first primibrachials (Fig. 6d) that have muscular facets proximally and distally. First primibrachials are trapezoidal in outline, and food groove is narrow (Figs. 5c–e, 6d). A considerable number of first primibrachials have concentric furrows and folds around the bulge (Fig. 5c–e), and some have also scratch marks (Fig. 5d, e), see above. Second primibrachials are trapezoidal (Fig. 5b) to nearly triangular (Figs. 5a, 7a) and are slightly convex; some carry ichnological traces as discussed above. Second primibrachials are thin and rectangular (Fig. 7c) to skewed trapezoidal in outline (Fig. 7b, d, e) with muscular facets at both ends. The aboral surface is finely granular; an ossicle has distinct marginal crenulae (Fig. 7c).
Remarks The upper Pliensbachian C. inaequalis Hess (2006) differs from C. docens by having mostly asymmetric radials; primibrachials are highly variable in outline, and first primibrachials lack a bulge. Cotylederma ambiguum Manni and Nicosia (1990) from the lower Toarcian (Serpentinum Chronozone) of Italy seems to be conspecific with C. docens. C. docens is morphologically similar to upper Cretaceous to extant Cyathidium (see Heinzeller et al. 1996; Donovan and Jakobsen 2004; Hess in Hess and Messing 2011). In contrast to Cyathidium species, Cotylederma docens has unfused primibrachials, and the brachials lack cirri. Nevertheless, it appears to be a precursor to Cyathidium. Early growth stages of Cyathidium foresti have two primibrachials that fuse in later stages into a single element, and early stages lack pinnules that develop on first secundibrachials only in juveniles larger than 6 mm in diameter (Heinzeller et al. 1997). In the extant form, brachials close the gap left by the fused primibrachials and, thus, seal the body cavity (Heinzeller et al. 1997, fig. 2e); such sealing can be assumed for fossil Cotylederma. According to the diagnosis of Améziane et al. (1999), extant Holopodidae (including fossil Cyathidium) have a cup comprised of five fused radials attached directly to the substrate. In contrast, Cotylederma has a hollow, variable aboral element attached to the substrate; its narrow upper edge articulates to the radial circlet, forming a thin-walled bowl.
Cotederma miliaris Deslongchamps, 1858, Fig. 6f.
Material MHNLM 2015.1.50. One first primibrachial.
Description The ossicle is wide and narrow, and food groove is strongly displaced to one side making this part only half as wide as the other; the narrow part is raised distally at an angle of 10° and is indented at the outer margin, presumably for articulation with adjoining ossicle. Aboral surface is covered by coarse granules. Aboral ligament pit and axial canal are elliptical and separated by thin ridge. Muscle fossae are small and deep, and they are separated from the adoral notch by a small peak. Interarticular ligament areas are large and flat and on the proximal side are with scratch marks.
Remarks The ossicle differs from first primibrachials of C. docens in lacking an aboral bulge and in having a surface with coarse granules. Loriol (1882, pl. 19, fig. 18) described a similar ossicle from the site of May-sur-Orne and ascribed it to C. miliaris Deslongchamps. Loriol’s material included two such ossicles assumed to be first primibrachials; his material contained neither radials nor axillary second primibrachials. The present ossicle differs from the one described by Loriol by its asymmetric profile and wider food groove. It is possible that Loriol’s specimen represents a secundibrachial. Granular basal elements were also described by Loriol; they differ not only by their granulation but also by their vase-like profile from those of C. docens. In summary, ossicles of C. miliaris fall outside the range of C. docens, so that two species of the genus are represented at May-sur-Orne and Feuguerolles.
Family Eudesicrinidae Bather 1899.
Eudesicrinus Loriol, 1882 in Loriol 1882–1889.
Eudesicrinus mayalis Deslongchamps in Deslongchamps and Deslongchamps, 1858, Fig. 8.

Eudesicrinus mayalis, cups with basal elements. a–f Eudesicrinus mayalis (Deslongchamps). a Symmetric juvenile cup fused to low basal element (arrows); MHNLM 2015.1.55, sample 357-13 early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). b Asymmetric juvenile cup fused to basal element, b1 lower side, attachment to substrate by two circular facets at left, b2 distal, two low radials lack articular facet; MHNLM 2015.1.56, sample 357-11, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). c Oblique distal view of post-juvenile cup; MHNLM 2015.1.57, sample 358-1, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). d Radial circlet fused to skew basal element, d1 distal view showing large radial, d2 lateral; MHNLM 2015.1.58, sample 358-5, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). e Cup (radial circlet) separated from basal element, e1 distal facet with large radial, e2 proximal facet connecting to basal element by ridges; MHNLM 2015.1.59, sample 358-2, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). f Large cup (radial circlet separated from basal element), f1 distal, f2 proximal with concave facet to basal element; MHNLM 2015.1.60, sample 425-5, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy)
Material MHNLM 2015.1.55–2015.1.60. Remains of this species include basal elements, mostly with attached radial circlet, cups (radial circlets), isolated radials, and secundibrachials. The best and most complete specimens are from the Tenuicostatum Chronozone (samples 357, 358, 425).
Description Our material of this well-known species is of interest, because it includes a juvenile cup less than 1 mm in diameter and height (Fig. 8a). In this specimen, radials are comparatively high and have identical size and facets; the basal element to which the radial circlet is fused is low. Cups with a diameter exceeding 1 mm have one radial enlarged (Fig. 8b, c), and such differentiation continues with growth to create the distinctive, inclined profile of the cup (Fig. 8d, e). The smallest specimen of the species from the Pliensbachian of Sulzkirchen (Jäger 1991, fig. 6) has a diameter of approximately 1 mm, with one radial enlarged and cup inclined, demonstrating that tilting of the cup may start in juveniles of similar ages. Articulation with the basal element is by radial ridges and depressions (Fig. 8, e2; see also Jäger 1991, figs. 1–4). However, the lower side of a large cup is concave and lacks ridges (Fig. 8, f2). Thus, the facet is similar to specimens figured by Loriol from the site of May-sur-Orne (1882; pl. 8, figs. 1d and 5b). However, the radial in pl.8, fig. 2b seems to have ridges on the lower side. The lower surface of basal elements commonly is concave and wider than the upper part that connects to the radial circlet (Fig. 8, d2). A number of radial circlets are fused with the basal element. Primibrachials and secundibrachials correspond to those figured by Loriol (1882) and Hess (2006, pl. 29, figs. 5–7), and their surface is granulated.
Remarks Radial circlets are fused to basal element or are provided with ridges and depressions, indicating rather strong articulation. This is in contrast to the specimens of Cotylederma docens, where the aboral element is thin-walled and often preserved isolated.
Suborder Cyrtocrinina Sieverts-Doreck 1952.
Superfamily Plicatocrinoidea Zittel 1879.
Diagnosis Cup consisting of fused basal circlet and circlet of 3–8 radials, commonly 4–6; radials may be partly or completely fused. Primibrachials 1 and 2 fused to axillary that carries unbranched arms, or joined by synostosis in lower Jurassic Plicatocrinus sulzkirchenensis, Sacariacrinus amadei n. sp., Praetetracrinus inornatus and P. doreckae, or muscular in Praetetracrinus kutscheri. Pinnules comprised of pinnulars that may be fused to form long spines or slightly curved rods. Column never fused to cup, columnals cylindrical or barrel-shaped to lenticular, facets with radiating marginal crenulae commonly arranged in groups or with irregular granules. Attachment by disk. (Modified from Hess in Hess and Messing 2011.)
Family Plicatocrinidae Zittel 1879.
Sacariacrinus Nicosia 1991.
Diagnosis Cup circular in section, comprised of 5–6-thick radials of variable height, with distinct sutures or fused, aboral surface smooth or weakly granulated, interradial embayments weak or absent; basal circlet compact, of variable height, may be fused with radial circlet. Radial articular facet rounded trapezoidal to elliptical, occupying full width of radial plate, inward sloping, and muscle fossae small and deep, surrounded by lip. Synostosis between primibrachials 1 and 2. First primibrachials of variable height. Secundibrachials joined by muscular articulation, proximal brachials bearing distinct pinnule sockets; distal brachials high, without pinnule sockets. Column unknown. (Modified from Hess in Hess and Messing 2011.)
Sacariacrinus amadei n. sp., Figs. 9 and 10.

Sacariacrinus amadei n. sp., cups and radials. a–h Sacariacrinus amadei n. sp. a Large cup, a1 proximal, a2 distal, a3 lateral; MHNLM 2015.1.61; holotype, sample 357-1, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). b Incomplete cup contained of three fused radials, b1 proximal, b2 distal; MHNLM 2015.1.62 paratype, sample 357-2, early Toarcian (Tenuicostatum Zone), Feuguerolles (Normandy). c Basal circlet, c1 proximal (lower), c2 distal (upper, with facets to 5 radials); MHNLM 2015.1.63, sample 425-3, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). d Radial, d1 aboral, d2 adoral, d3 facet; MHNLM 2015.1.64, sample 375-6, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). e Aboral/proximal view of radial, facet to basal circlet with weak ridge; MHNLM 2015.1.65, sample 425-1, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). f Smooth radial, f1 aboral, f2 distal note strong relief; MHNLM 2015.1.66, sample 357-7, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). g Small radial, aboral view, note profile of ridge on lower facet; MHNLM 2015.1.67, sample 357-9, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). h Radial, h1 aboral with granular surface, h2 facet; MHNLM 2015.1.68, sample 357-3, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy)

Sacariacrinus amadei n. sp., cups, radials, first and second primibrachials, secundibrachials. a–l Sacariacrinus amadei n. sp. a Radial with slightly angular facet to basal circlet, a1 aboral, a2 adoral; MHNLM 2015.1.69, sample 540-3, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). b Sculptured radial with slightly angular facet to basal circlet, b1 aboral, b2 adoral; MHNLM 2015.1.70, sample 540-4, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). c Incomplete cup with fused radials, one radial missing, c1 proximal, c2 distal; MHNLM 2015.1.71, sample 540-6, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). d Second primibrachial adoral; MHNLM 2015.1.72, sample 357-4, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). e Second primibrachial adoral; MHNLM 2015.1.73, sample 540-5, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). f Small second primibrachial, aboral/proximal view showing synostosial facet with marginal crenulae; MHNLM 2015.1.74, sample 425-5, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). g Aboral/distal view of first primibrachial; MHNLM 2015.1.75, sample 540-2, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). h First primibrachial, h1 aboral, h2 adoral, h3 oblique proximal; paratype, MHNLM 2015.1.76, sample 357-8, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). i Proximal secundibrachial adoral, note large pinnule socket; MHNLM 2015.1.77, sample 357-24, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). j Secundibrachial with pinnule socket, adoral view; MHNLM 2015.1.78, sample 357-25, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). k High first primibrachial adoral; MHNLM 2015.1.79, sample 357-27, early Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy). l Adoral view of distal secundibrachial lacking pinnule socket; MHNLM 2015.1.80, sample 357-26, Toarcian (Tenuicostatum Chronozone), Feuguerolles (Normandy)
Material MHNLM 2015.1.61–2015.1.80. Three basal circlets, two with five facets to radials, one with six; three radial circlets, two incomplete and attached to basal circlet, isolated radials, first and second primibrachials, secundibrachials. Samples 357, 358, 425, 540; early Toarcian (Tenuicostatum Zone).
Holotype MHNLM 2015.1.61. Radial circlet, Fig. 9a.
Paratypes MHNLM 2015.1.62 Incomplete radial circlet attached to intact basal circlet, Fig. 9b; MHNLM 2015.1.76, first primibrachial, Fig. 10h.
Etymology Species named in honour of both Wolfgang Amadeus Mozart, arguably the rock star among the composers of the Classical Era, and of Falco who paid tribute to Mozart’s subliminal rock’n’roll attitude with his song “Rock me Amadeus”.
Type locality and horizon Feuguerolles, early Toarcian (Tenuicostatum Zone).
Diagnosis Radial circlet comprised of 5–6 low compact plates that may be completely or partly fused; basal circlet low. Width of radial cavity about 30% of cup diameter. First and second primibrachials joined by synostosis, rarely weakly muscular. Proximal secundibrachials with pinnule sockets.
Description The holotype (Fig. 9a) is a rather low circlet comprised of five smooth radials with distinct sutures; the profile is circular with weak interradial embayments. Articulation to the basal circlet is by five synostosial facets that have scattered granules near the outer margin. The aboral surface is smooth. Radial articular facets are rounded trapezoidal and framed by a rim toward the radial cavity that narrows to the basal circlet. Aboral ligament fossa is wide and slightly outward-sloping; it occupies up to half the facet, and the ligament pit is elliptical. The adoral part of the facet with triangular interarticular ligament fossae is inward-sloping and separated from the aboral part by a distinct transverse ridge pierced by the axial canal. Muscle fossae are small, rimmed, and separated by a narrow notch. A strong lip separated by an interradial notch borders the facets from the radial cavity. The paratype (Fig. 9b) is an incomplete radial circlet of three tightly joined smooth pieces on top of an intact basal circlet. Radials originally numbered six, leading to hexagonal profile. Outline of the basal circlet is rounded hexagonal. A third, partly intact radial circlet (Fig. 10c) misses one radial out of five; it is rounded pentagonal in outline; and the radials are tightly fused, without interradial embayments. An isolated basal circlet (Fig. 9c) has the upper (distal) side with five facets to radials; the lower (proximal) side is concave and smooth. This ossicle demonstrates that the radial cavity is prolonged basally into the uppermost columnal. Most radials are sturdy and smooth aborally, but some are more or less granular (Figs. 9d, h, 10b). Whereas their upper facets are all similar, the lower facets to the basal circlet are somewhat variable, ranging from more or less angular (Figs. 9e, g, 10a, b) to nearly straight (Fig. 9d, f, h). Such a difference seems to indicate that articulation between basal and radial circlets varied in strength, and this is also demonstrated by preservation of radials still attached to the basal circlet. Radials are joined by smooth synostosial facets; the inner, adoral part is concave and of somewhat variable width. However, all radials are low and lack interradial extensions. A number of first primibrachials are assigned to the species. They vary considerably in height (Fig. 10g, h, k), but all have narrow food grooves, a proximal facet matching the radial facet, and a synostosial distal facet (Fig. 10g). The ossicles are weakly convex in profile, and the aboral side is smooth. Second primibrachials are lower (Fig. 10d, e); they are smooth aborally and have synostosial proximal and muscular distal facets (Fig. 10d). These extend adorally with rimmed muscle fossae, similar to those of the radials. A small second primibrachial has a weakly cryptosyzygial proximal facet (Fig. 10f). Secundibrachials vary widely in height (Fig. 10i, j, l). Their facets are muscular, and proximal brachials carry a large pinnule socket (Fig. 10i).
Remarks The present species is distinguished from S. altineri Nicosia (1991) by lower radials and basal circlet. Nicosia mentioned the occurrence of two morphotypes with thicker and thinner radials at the late Sinemurian–early Pliensbachian-type locality, his figures relate to the thick-walled type. Thick radials, primibrachials and a basal element from the upper Pliensbachian (Domerian) of Arzo, were ascribed by Hess (2006) to Sacariacrinus altineri, whereas thinner elements were designated S. cf. altineri. The thinner variety is similar to Praetetracrinus inornatus, although the Arzo radials are taller and the adoral incision with the muscle fossae extends more strongly downward (see Hess 2006, pl. 3, fig. 1b).
Family Tetracrinidae Nicosia 1991.
Diagnosis see Hess in Hess and Messing 2011.
Tetracrinus Münster, 1839.
Tetracrinus solidus n. sp., Fig. 11.

Tetracrinus solidus n. sp., cup.Tetracrinus solidus n. sp. a Holotype proximal, b distal, c lateral; MHNLM 2015.1.81, sample 358, early Toarcian (Tenuicostatum Zone), Feuguerolles (Normandy)
Material Only the holotype.
Holotype MHNLM 2015.1.81. Cup, Fig. 11.
Etymology L, solidus, for the firmly joined plates making up the cup.
Type locality and horizon Feuguerolles, sample 358, early Toarcian (Tenuicostatum Zone).
Diagnosis Cup conical, squarish, comprised of four radials fused with basal circlet to single piece with smooth latus. Radial articular facets nearly horizontal, occupying most of the distal part of the cup, no interradial sutures; aboral ligament fossa narrow with elliptical pit; interarticular ligament fossae large, separated by faint ridge from muscle fossae; and muscle fossae with distinct lip to radial cavity. Width of radial cavity 20% of cup diameter. Facet to column shallow concave and smooth, width of axial canal similar to that of radial cavity.
Description See diagnosis.
Remarks At first glance, the specimen seems to be comprised of only fused radials; but the lower, slightly concave facet suggests attachment to a column. Thus, it must be the lower (proximal) facet of a basal circlet. In the upper Jurassic Tetracrinus moniliformis (Münster), the cup articulates with the topmost columnal by symplexy with grouped radial crenulae (see Hess in Hess and Messing 2011). In the early Oxfordian, T. galei Hess (2014a) articulation is symplectial with reduced crenulae. The species co-occurs at sample 358 with Sacariacrinus amadei n. sp., whose radials are superficially similar. However, the cup of S. amadei is comprised of five somewhat bulging radials with a rather narrow base; radial articular facet of S. amadei is inward-sloping and bears small deep muscle fossae surrounded by a distinct rim. Tetracrinus solidus n. sp. is distinguished from the Upper Jurassic T. moniliformis by the higher, smooth cup, and narrow radial cavity; correspondingly, facets are wider. The Oxfordian T. galei differs from the present one by a wider radial cavity. The holotype of T. galei is a smooth conical cup with basal element and radial circlet separated by a distinct constriction; interradial sutures are indistinct, and thus, it approaches the new lower Jurassic species. Tetracrinus kocyigiti Nicosia (1991) from the upper Sinemurian/lower Pliensbachian of Turkey has 4–5 distinct basals carrying an inclined circlet of five radials.
Praetetracrinus Jäger 1995.
Remarks Jäger (1995) proposed the genus Praetetracrinus for Lower Jurassic cyrtocrinoids with tetramerous symmetry, fused basal circlet, high triangular and thin-walled radials and primibrachials connected by synostosis. Included species are P. doreckae Jäger 1995 (type species, lower Aalenian), P. inornatus (Simms 1989, Domerian–Toarcian), and P. kutscheri Jäger (1995, upper Toarcian–lower Aalenian). On a visit to the type locality of inornatus (Watton Cliff, Dorset), Jäger (1995, p. 10) noted the occurrence of two separate species, one being inornatus and the other, smaller species unnamed. According to Jäger, P. doreckae is closely related to P. inornatus, the two forms being parts of a phylogenetic series characterised by an increase in size.
Diagnosis See Hess in Hess and Messing 2011.
Type species Praetetracrinus doreckae Jäger, 1995.
Praetetracrinus inornatus (Simms, 1989), Figs. 12 and 13.

Praetetracrinus inornatus, basal circlets and columnals. a–m Praeteracrinus inornatus (Simms). a Basal circlet lateral; MHNLM 2015.1.82, sample 082-3, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). b Basal circlet, b1 distal (upper), b2 proximal (lower); MHNLM 2015.1.83, sample 082-2, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). c Basal circlet distal; MHNLM 2015.1.84, sample 490-3, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). d Basal circlet, d1 distal (upper), d2 proximal (lower); MHNLM 2015.1.85, sample 325-5, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). e Facet of proximal columnal; MHNLM 2015.1.86, sample 457-2, late Toarcian (Variabilis Chronozone), Feuguerolles (Normandy). f Topmost columnal, f1 upper facet with extension of radial cavity, f2 lower facet to column; MHNLM 2015.1.87, sample 082-1, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). g Cylindrical columnal, g1 lateral with furrows, g2 facet with three groups of crenulae; MHNLM 2015.1.88, sample 308-1, late Toarcian (Variabilis Chronozone), Feuguerolles (Normandy). h Cylindrical columnal, h1 lateral with furrows, h2 facet; MHNLM 2015.1.89, sample 325-6, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). i Cylindrical columnal, i1 lateral with epizoans, i2 facet; MHNLM 2015.1.90, sample 325-3, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). j Barrel-shaped columnal with weakly expressed facet, j1 lateral, j2 facet; MHNLM 2015.1.91, sample 370-3, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). k Barrel-shaped columnal, k1 lateral, k2 facet with perilumen, areola and crenulated rim; MHNLM 2015.1.92, sample 370-5, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). l Cylindrical columnal, l1 lateral, l2 tetramerous facet; MHNLM 2015.1.93, sample 389-1, late Toarcian (Thouarsense Chronozone), Feuguerolles (Normandy). m cylindrical columnal, m1 lateral, m2 facet with weakly expressed crenulae; MHNLM 2015.1.94, sample 400, early Aalenian (Opalinum Chronozone, last occurrence of species), Feuguerolles (Normandy)

Praetetracrinus inornatus, radials and primibrachials. a–j Praetetracrinus inornatus (Simms). a radial without interradial processes, a1 adoral, a2 aboral; MHNLM 2015.1.95, sample 540-1, early Toarcian (Tenuicostatum Chronozone, first occurrence of species), Feuguerolles (Normandy). b Radial with interradial processes, b1 adoral, b2 aboral; MHNLM 2015.1.96, sample 375-7, early Toarcian (Tenuicostatum/Serpentinum Chronozones), Feuguerolles (Normandy). c Adoral view of radial with interradial process on one side; MHNLM 2015.1.97, sample 293-2, late Toarcian (Variabilis Chronozone), Feuguerolles (Normandy). d Radial with weak interradial processes, d1 adoral, d2 aboral, d3 distal (facet); MHNLM 2015.1.98, sample 082-9 middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). e Wide radial, aboral view with scratch marks; MHNLM 2015.1.99, sample 293-1, late Toarcian (Variabilis Chronozone), Feuguerolles (Normandy). f Thick radial, f1 adoral, note constriction, f2 distal, f3 aboral, f4 proximal, f5 proximal view of radial circlet reconstituted from f4; MHNLM 2015.1.100, sample 370-b, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). g First primibrachial, g1 adoral (interior), g2 aboral, g3 proximal facet, g4 distal facet; MHNLM 2015.1.101, sample 082-5, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). h Aboral view of high second primibrachial; MHNLM 2015.1.102, sample 211-4, early Toarcian (Serpentinum Chronozone), Feuguerolles (Normandy). i Second primibrachial, i1 aboral/proximal, i2 adoral; MHNLM 2015.1.103, sample 082-7, middle Toarcian (Bifrons Chronozone), Feuguerolles (Normandy). j low second primibrachial, j1 aboral, j2 adoral; MHNLM 2015.1.104, sample 418-4, early Toarcian (Serpentinum Chronozone), Feuguerolles (Normandy)
Plicatocrinus inornatus Simms, 1989, p. 86.
Praetetracrinus inornatus (Simms), Jäger 1995, p. 21.
Sacariacrinus cf. altineri Nicosia, Hess 2006, pl. 5, figs. 1, 2, 12
Material MHNLM 2015.1.82–2015.1.104. Numerous columnals, basal circlets, radials, and first and second primibrachials. The remains are common from the lower Toarcian Serpentinum Zone to the late Toarcian Variabilis Chronozone. A single radial is from the Tenuicostatum Chronozone (Fig. 13a), and a columnal is from the Aalenian (Fig. 12m).
Diagnosis Basal circlet fairly low, slightly concave, proximal facet with four groups of a few marginal crenulae, distal facet with four ridges separating slightly concave synostosial facets for articulation with the radials. Radial cavity wide; radials high, thin-walled, with interradial extensions; radial articular facet narrow, muscle fossae hardly extending downward. Columnals high to low, cylindrical or barrel-shaped; facets with crenulae in four groups or scattered, crenulae weakly developed in barrel-shaped columnals. (The diagnosis is adapted from Simms’ original description and includes the present material.)
Description The basal circlet is smooth and circular in outline, but may be rounded tetragonal in small specimens (Fig. 12c). The lower (proximal) facet is concave and has four groups of crenulae that do not reach the narrow axial canal. The upper (distal) facet has a central depression of somewhat variable width and four outward-sloping facets to radials; paired nerve canals are around the central depression (Fig. 12b, c). Thin topmost columnals have four groups of 2–3 crenulae (Fig. 12e, f); in one ossicle, the other facet is concave, suggesting connection to the basal circlet (Fig. 12f). Columnals are mostly smooth and cylindrical; facets are symplectial with short crenulae in four groups (Fig. 12g, h); and in some specimens, crenulae are reduced or scattered (Fig. 12i, m). Barrel-shaped columnals are less common, and their facets variable; some have weakly crenulated rims with areola and raised perilumen (Fig. 12k); and others have flat facets with a few scattered granules and a weak perilumen (Fig. 12j). However, facets such as those in Fig. 12j, k seem to match. Many cylindrical columnals have furrows or bite marks, discussed below. Radials are thin, aborally smooth and trapezoidal in profile; most have short interradial extensions. Facet to the basal circlet is straight in profile (Fig. 13a–f). Interradial facets are mostly narrow (Fig. 13a, b), but they may widen in part (Fig. 13c) or all of their height (Fig. 13d, f). The adoral side is concave and may have irregular sculpturing (Fig. 13d). Radial articular facet is a narrow ellipse. Aboral ligament is narrow and visible from the outside; muscle fossae are rimmed and separated by a narrow notch. They extend adorally pouch-like into the radial cavity but less so than in Sacariacrinus amadei n. sp. First primibrachials are rounded rectangular in outline and moderately high, and their surface is smooth. The food groove is narrow; it separates rimmed muscle fossae that slope inward to nearly half of ossicle height (Fig. 13g, 1). Distal facet is synostosial, with some marginal crenulae (Figs. 4, 13g). Axillary second primibrachials vary in height (Fig. 13i, h). Their proximal facets are synostosial; the distal muscular facets are separated by a process.
Remarks Nicosia (1991) compared Sacriacrinus altineri with Plicatocrinus inornatus Simms, which he ascribed to Sacariacrinus. As described by Simms, P. inornatus has thin-walled radials with interradial extensions. In our material, P. inornatus is represented in the Tenuicostatum Zone, the type horizon of Sacariacrinus amadei n. sp., by a single, thin radial (Fig. 13a) with straight lower suture and without interradial extensions; Sacariacrinus radials have more or less angled sutures for articulation to the basal circlet (Fig. 9e; see also Nicosia 1991, Fig. 13b). P. inornatus is remarkably similar to P. bathonicus Hess (2012). However, in the middle Jurassic species, the basal circlet is higher; radials are slightly keeled aborally and have a narrower base. At Feuguerolles, secundibrachials could not be assigned to this species with confidence. Such ossicles were described by Simms (1989, pl. 15, figs. 17, 20), and Jäger (1995, pl. 5) described secundibrachials of Praetetracrinus doreckae. In both cases, articulations are muscular, and the ossicles carry pinnule sockets.
Jäger reported deep furrows (“Rillen”) on columnals of Praetetracrinus doreckae (1995, pl. 1, figs. 17–20) and P. kutscheri or inornatus (pl. 6, fig. 8). In the present material, shallow scratch marks on columnals are rather common (Fig. 12g, h, k, m). On a Praetetracrinus radial (Fig. 13e) are scratch marks; similar traces also occur on columnals of Amaltheocrinus. The absence of skeletal growth around furrows or scratches suggests that the traces were made on dead ossicles lying on the sea floor.
References
Améziane, N., Bourseau, J.-P., Heinzeller, T., & Roux, M. (1999). Les genres Cyathidium et Holopus au sein des Cyrtocrinida (Crinoidea; Echinodermata). Journal of Natural History, 33, 439–470.
Arendt, Y. A. (1974). The sea lilies cyrtocrinids. Trudy Paleontologiceskogo Instituta Akademia Nauka SSSR, 144, 1–251. (in Russian).
Bather, F. A. (1899). A phylogenetic classification of the Pelmatozoa. Report of the British Association for the Advancement of Science, 1898, 916–923.
Bernoulli, D., & Jenkyns, H. C. (1970). A Jurassic Basin: The Glasenbach Gorge, Salzburg, Austria. Verhandlungen der Geologischen Bundesanstalt, 1970(4), 504–531.
Bernoulli, D., & Jenkyns, H. C. (2009). Ancient oceans and continental margins of the Alpine-Mediterranean Tethys: Deciphering clues from Mesozoic pelagic sediments and ophiolites. Sedimentology, 56, 149–190.
Böhm, F. (2003). Lithostratigraphy of the Adnet Group (Early to Middle Jurassic, Salzburg, Austria). In W. E. Piller (Ed.), Stratigraphia Austriaca (Vol. 16, pp. 231–268). Graz: Österreichische Akademie der Wissenschaften, Schriftenreihe der Erdwissenschaftlichen Kommission.
Bribiesca-Contreras, G., Verbruggen, H., Hugall, A. F., & O’Hara, T. D. (2017). The importance of offshore origination revealed through ophiuroid phylogenomics. Proceedings of the Royal Society B, 284, 20170160.
Cohen, B. L., & Pisera, A. (2015). Crinoid phylogeny: a new interpretation of the main Permian-Triassic divergence, comparison with echinoids and brachiopods, and EvoDevo interpretations of major morphological variations. Biological Journal of the Linnean Society. https://doi.org/10.1111/bij.12868.
de Loriol, P. (1882). Paléontologie Française, Série 1, Animaux Invertébrés. Terrain Jurassique, 11, Crinoïdes (Vol. 1(1882–84)). Paris: G. Masson. (vol 2, 1884–89).
Delogu, D., & Nicosia, U. (1986). A new cyrtocrinid from the Lias of Turkey: Capsicocrinus souti n. gen., n. sp. Geologica Romana, 23, 157–162.
Deslongchamps, J. A., & Deslongchamps, E. E. (1858). Mémoires sur la couche à Leptaena. Bulletin de la Société Linnéenne de Normandie, 3(1857–58), 132–187.
Donovan, S. K. (2017). A plea not to ignore ichnotaxonomy: recognizing and recording Ooichnus Bromley. Swiss Journal of Palaeontology, 136, 369–372.
Donovan, S. K., & Jakobsen, S. L. (2004). An unsual crinoid-barnacle association in the type area of the Danian (Paleocene), Denmark. Lethaia, 37, 407–415.
Donovan, S. K., & Tenny, A. (2015). A peculiar bored crinoid from the Salthill Quarry, Clitheroe, Lancashire (Mississippian, Tournaisian), UK. Proceedings of the Yorkshire Geological Society, 60, 289–292. https://doi.org/10.1144/pygs2015-362.
Dugué, O., Fily, G., & Rioult, M. (1998). Le Jurassique des Côtes du Calvados. Biostratigraphie, sédimentologie, paléoécologie, paléogéographie et stratigraphie séquentielle. Bulletin Trimestriel de la Société Géologique de Normandie et des Amis du Muséum du Havre, 85, 1–132.
Hagdorn, H. (1996). Palökologie der Trias-Seelilie Dadocrinus. Geologisch-Paläontologische Mitteilungen Innsbruck, 21, 19–45.
Hagdorn, H. (2011). The Triassic—Crucial period of post-Palaeozoic crinoid diversification. Swiss Journal of Palaeontology, 130, 91–112.
Heinzeller, T., Fechter, H., Améziane-Cominardi, N., & Welsch, U. (1997). Development of Cyathidium foresti (Echinodermata: Crinoidea, Cyrtocrinida) from early attached larvae to adult-like juveniles. Journal of Zoological Systematics and Evolutionary Research, 35, 11–21.
Heinzeller, T., Fricke, H., Bourseau, J.-P., Améziane-Cominardi, N., & Welsch, U. (1996). Cyathidium plantei n. sp., an extant cyrtocrinid (Echinodermata, Crinoidea)—Morphologically identical to the fossil Cyathidium depressum (Cretaceous, Cenomanian). Zoologica Scripta, 25(1), 77–84.
Hemery, L. G., Roux, M., Améziane, N., & Eléaume, M. (2013). High-resolution crinoid phyletic inter-relationships derived from molecular data. Cahiers de Biologie Marine, 54, 511–523.
Hess, H. (2006). Crinoids from the Lower Jurassic (Pliensbachian) of Arzo, southern Switzerland. Schweizerische Paläontologische Abhandlungen, 126, 1–143.
Hess, H. (2012). Crinoids from the Middle Jurassic (Bajocian-Lower Callovian) of Ardèche, France. Swiss Journal of Palaeontology, 131, 211–253.
Hess, H. (2014a). Balanocrinus and other crinoids from Late Jurassic mudstones of France and Switzerland. Swiss Journal of Palaeontology, 133, 47–75.
Hess, H. (2014b). The crinoid Eugeniacrinites cariophilites from the Late Jurassic of southern Germany: Babies, cripples and enigmatic wing plates. Swiss Journal of Palaeontology, 133, 121–140.
Hess, H., & Messing, C. G. (2011). Treatise on invertebrate paleontology, Part T, Echinodermata 2, revised. In W. I. Ausich (Ed.), Crinoidea (Vol. 3, pp. 1–261). Lawrence: The University of Kansas Paleontological Institute.
Hess, H., & Thuy, B. (2016). Extraordinary diversity of feather stars (Echinodermata: Crinoidea: Comatulida) from a Lower Jurassic (Pliensbachian–Toarcian) rock reef of Feuguerolles (Normandy, France). Swiss Journal of Palaeontology, 136, 301–321. https://doi.org/10.1007/s13358-016-0122-5.
Huelsenbeck, J. P., & Ronquist, F. (2001). MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics (Oxford, England), 17, 754–755.
Jäger, M. (1985). Die Crinoiden aus dem Pliensbachium (mittlerer Lias) von Rottorf am Klei und Empelde (Niedersachsen). Berichte der Naturforschenden Gesellschaft Hannover, 128, 71–129.
Jäger, M. (1991). Die Crinoiden aus dem Pliensbachium vom Rhein-Main-Donau-Kanal bei Sulzkirchen. Teil 1: Cyrtocriniden mit reduziertem Stiel: Eudesicrinus und Cotylederma. Archaeopteryx, 9, 35–46. (Eichstätt).
Jäger, M. (1993). Die Crinoiden aus dem Pliensbachium vom Rhein-Main-Donau-Kanal bei Sulzkirchen. Teil 2: Gestielte Cyrtocrinina und Millericrinina: Amaltheocrinus, Shroshaecrinus?, Plicatocrinus und Nachtrag zu Teil 1 (Eudesicrinus). Archaeopteryx, 11, 73–104.
Jäger, M. (1995). Echinodermata aus dem Ober-Toarcium und Aalenium Deutschlands. I. Crinoidea: Cyrtocrinina und Millericrinina. Stuttgarter Beiträge zur Naturkunde B, 226, 1–51.
Kázmér, M., & Tabaroši, D. (2012). Bioerosion on the small scale—Examples from tropical and subtropical littoral. Hantkeniana, 7, 37–94. (Monostori jubilee volume, Budapest).
Kristan-Tollmann, E. (1975). 6. Stielcrinoiden. In E. Kristan-Tollmann & L. Krystyn (Eds.), Die Mikrofauna der ladinisch-karnischen Hallstätter Kalke von Saklibeli (Taurus-Gebirge, Türkei) (Vol. 184, pp. 259–340). Wien: Sitzungsberichte der Österreichischen Akademie der Wissenschaften, Mathematisch-Naturwissenschaftliche Klasse.
Lewis, P. O. (2001). A likelihood approach to estimating phylogeny from discrete morphological character data. Systematic Biology, 50, 913–925.
Manni, R., & Nicosia, U. (1990). New Crinoids from Liassic sediments nearby Tivoli (Central Italy); rearrangement in the systematics of eudesicrinids and cotyledermatids. Geologica Romana, 26, 81–92.
Manni, R., & Nicosia, U. (1996). A history of cyrtocrinid evolution. Palaeopelagos, 6, 3–17.
Manni, R., & Nicosia, U. (1999). The oldest cyrtocrinid, Nerocrinus petri n. gen. n. sp., and the Nerocrinidae, n. fam. In M. D. Candia Carnevali & F. Bonasero (Eds.), Echinoderm Research (pp. 327–331). Amsterdam: Balkema.
Münster, G. G. Z. (1839). Beschreibung einiger neuer Crinoideen aus der Übergangsformation. Beiträge zur Petrefactenkunde, 1, 1–124.
Neuweiler, F., & Bernoulli, D. (2005). Mesozoic (Lower Jurassic) red stromatactis limestones from the southern Alps (Arzo, Switzerland): Calcite mineral authigenesis and syneresis-type deformation. International Journal of Earth Sciences, 94, 130–146.
Nicosia, U. (1991). Mesozoic crinoids from the north-western Turkey. Geologica Romana, 27, 389–436.
Page, K. N. (2003). The Lower Jurassic of Europe: Its subdivision and correlation. In J. R. Ineson & F. Surlyk (Eds.), The Jurassic of Denmark and Greenland (Vol. 1, pp. 23–59). Copenhagen: Geological Survey of Denmark and Greenland Bulletin.
Quenstedt, F. A. (1852). Handbuch der Petrefactenkunde (1st ed., p. 792). Tübingen: Laupp.
Quenstedt, F. A. (1876). Petrefactenkunde Deutschlands. 1. Abt., 4. Band, Echinodermen (Asteriden und Encriniden), Atlas zu den Asteriden und Encriniden (pp. 90–114). Leipzig: Fuess.
Rasmussen, H. W. (1961). A monograph on the cretaceous Crinoidea. Biologiske Skrifter Det Kongelige Danske Videnskabernes Selskab, 12(1), 1–428.
Rioult, M. (1968). Contribution à l’étude du Lias de la Bordure Occidentale du Bassin de Paris. PhD Thesis, Université de Caen (unpublished).
Rioult, M., & Chirat, R. (1999). Ammonites jurassiques sur la bordure occidentale du Bassin anglo-parisien. Contrôle et modalités de leur répartition. Bulletin de la Société Linnéenne de Normandie, 117, 49–60.
Romano, M., Manni, R., & Nicosia, U. (2016). Phylogenetic analysis of cyrtocrinid crinoids and the influence on traditional classifications. Journal of Mediterranean Earth Sciences, 8, 1–21.
Rouse, G. W., Jermin, L. S., Wilson, N. G., Eeckhaut, I., Lanterbecq, D., Oji, T., et al. (2013). Fixed, free, and fixed: the fickle phylogeny of extant Crinoidea (Echinodermata) and their Permian-Triassic origin. Molecular Phylogenetics and Evolution, 66, 161–181.
Roux, M. (1980). Les articulations du pédoncule des Hyocrinidae (Échinodermes, Crinoïdes pédonculés): Intérêt systématique et conséquences. Bulletin du Muséum d’Histoire Naturelle de Paris, 4e Sér., 2, Section A, 1, 31–57.
Salamon, M. A., Gorzelak, P., & Zaton, M. (2009). In quest of cyrtocrinids origins: evidence from Late Triassic ossicles from the Tatra Mountains. Acta Palaeontologica Polonica, 54, 171–174.
Sengör, A. M. C. (1979). Mid-mesozoic closure of Permo-Triassic Tethys and its implications. Nature, 279, 590–593.
Sengör, A. M. C., & Yilmaz, Y. (1981). Tethyan evolution of Turkey: A plate tectonic approach. Tectonophysics, 75, 181–241.
Sieverts-Doreck, H. (1951). Cyathidium im Tithon von Mähren? Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 94, 1–4.
Sieverts-Doreck, H. (1952). In: Moore, R.C., Lalicker, C.G. & Fischer, A.G. (eds) Invertebrate Fossils. New York: McGraw-Hill.
Simms, M. J. (1989). British Lower Jurassic crinoids. Monograph of the Palaeontographical Society (London, Publication), 581, 1–103.
Simms, M. J. (1990). Crinoid diversity and the Triassic/Jurassic Boundary. Cahiers de l’Université Catholique de Lyon, Série Sciences, 3, 67–77.
Stöhr, S., & Martynov, A. (2016). Paedomorphosis as an evolutionary driving force: Insights from deep-sea brittle stars. PLoS One, 11(11), e0164562.
Syverson, V. J., Messing, C. G., Stanley, K., & Baumiller, T. K. (2015). Growth, injury, and population density in the extant cyrtocrinid Holopus mikihe (Crinoidea, Echinodermata) near Rohatán, Honduras. Bulletion of Marine Sciences, 91(1), 46–61.
Thuy, B. (2013). Temporary expansion to shelf depths rather than an onshore–offshore trend: The shallow-water rise and demise of the modern deep-sea brittle star family Ophiacanthidae (Echinodermata: Ophiuroidea). European Journal of Taxonomy, 48, 1–242.
Thuy, B., Kiel, S., Dulai, A., Gale, A. S., Kroh, A., Lord, A. R., et al. (2014). First glimpse into Lower Jurassic deep-sea biodiversity: in situ diversification and resilience against extinction. Proceedings of the Royal Society B, 281, 20132624.
Thuy, B., & Stöhr, S. (2016). A new morphological phylogeny of the Ophiuroidea (Echinodermata) accords with molecular evidence and renders microfossils accessible for cladistics. PLoS One, 11(5), e0156140. https://doi.org/10.1371/journal.pone.0156140.
Vernhet, Y., Maurizot, P., Le Gall, J., Gigot, P., Dupret, L., Lerouge, G., et al. (2002). Notice explicative, Carte géologique de la France à 1/50 000, feuille Villiers-Bocage (145) (p. 229). Orléans: Bureau de Recherches Géologiques et Minières.
von Zittel, K. A. (1879). Handbuch der Palaeontologie (Vol. 1). Oldenburg: Palaeozoologie, Abt. 1. R.
Weis, R., Dzyuba, O. S., Mariotti, N., & Chesnier, M. (2015). Lissajousibelus nov. gen., an Early Jurassic “canaliculate” belemnite from Normandy, France. Swiss Journal of Palaeontology, 134(2), 289–300.
Wisshak, M., Neumann, C., Jakobsen, J., & Freiwald, A. (2009). The ‘living-fossil community’ of the cyrtocrinid Cyathidium foresti and the deep-sea oyster Neopycnodonte zibrowii (Azores Archipelago). Palaeogeography, Palaeoclimatology, Palaeoecology, 271, 77–83.
Wright, T. (1876). On the Cotyledermidae. Geological Magazine, 2(3), 94–95.
Wright, A. M., & Hillis, D. M. (2014). Bayesian analysis using a simple likelihood model outperforms parsimony for estimation of phylogeny from discrete morphological data. PLoS One, 2014(9), e109210. https://doi.org/10.1371/journal.pone.0109210.
Zardini, R. (1976). Fossili di Cortina. Cortina d’Ampezzo: Atlante degli echinodermi cassiani (Trias medio superior) della regione Dolomitica attorno a Cortina d’Ampezzo.
Zardini, R. (1988). Geologia e fossili delle Dolomiti di Cortina e dintorni. Cortina d’Ampezzo: Nuova edizione a cura di M. Spampani. (Edizioni Dolomiti).
Žítt, J. (1974a). Sclerocrinus Jaekel, 1891 and Proholopus Jaekel, 1907 (Crinoidea, Cyrtocrinida) from the Lower Cretaceous of Štramberk (Czechoslovakia). Sbornik Geologickŷ Věd, Paleontologie, 16, 7–34.
Žítt, J. (1974b). Eugeniacrinites Miller, 1821 from the Lower Cretaceous of Štramberk (Czechoslovakia). Věstnik Ústředního Ústavu Geologického, 49, 265–272.
Žítt, J. (1975). Sclerocrinus kotoucensis sp. n. (Crinoidea, Cyrtocrinida) from the Lower Cretaceous of Štramberk (Czechoslovakia). Věstnik Ústředního Ústavu Geologického, 50, 115–117.
Žítt, J. (1978a). Apsidocrinus Jaekel, 1907 and Psalidocrinus Remeš (Crinoidea, Cyrtocrinida) from the Lower Cretaceous of Štramberk (Czechoslovakia). Sbornik Geologickŷ Věd, Paleontologie, 21, 107–124.
Žítt, J. (1978b). Phyllocrinus d’Orbigny, 1850 (Cyrtocrinida) from Štramberk (Czechoslovakia). Časopius Pro Mineralogii a Geologii Roč. 23, č, 3, 39–51.
Žítt, J. (1979a). Hemibrachiocrinidae Arendt, 1968 (Crinoidea, Cyrtocrinida) from the Lower Cretaceous of Štramberk (Czechoslovakia). Věstnik Ústředního Ústavu Geologického, 54, 341–348.
Žítt, J. (1979b). Strambergocrinus gen. n. (Cyrtocrinida) from the the Lower Cretaceous of Štramberk (Czechoslovakia). Časopius Pro Mineralogii a Geologii, Roč. 24, č., 3, 237–247.
Žítt, J. (1983). Spoon-like crinoids from Štramberk (Lower Cretaceous, CSSR). Sborník Narodního Muzea V Praze (Acta Musei Nationalis Pragae), 39 B(2), 69–114.
Acknowledgements
We are greatly indebted to Marc Chesnier (Cresserons, Calvados, France) who skillfully collected the Feuguerolles crinoid material in the course of many years and generously made it available for study. He, furthermore, provided all relevant details on the stratigraphy of the material. We are grateful to Daniel Bernoulli (Basel, Switzerland) for information and discussion on the palaeogeography of the western Tethys and to Walter Etter (Basel, Switzerland) for Figs. 1 and 3. Steve Donovan provided information on ichnological traces. We also thank the reviewers Tom Baumiller and William Ausich, whose comments greatly improved the manuscript. This work was carried out as part of the research project ‘The Pliensbachian–Toarcian perturbations and their legacy in marine biodiversity of the Western Tethys’ by the Natural History Museum Luxembourg.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial Handling: Daniel Marty.
Deceased: Hans Hess.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Hess, H., Thuy, B. Emergence and early radiation of cyrtocrinids, with new species from a Lower to Middle Jurassic rock reef of Feuguerolles (Normandy, France). Swiss J Palaeontol 137, 133–158 (2018). https://doi.org/10.1007/s13358-018-0160-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-018-0160-2