
Abstract
The top of the Querales Formation is well exposed at the Quebrada Corralito section, 17 m thick, in northern Venezuela. The section, dominated by siliciclastic accumulations of fine-grained sediments, preserves one cycle of transgressive–regressive phases of the deltaic environments facing the marine platform. An X-ray analysis revealed kaolinite as the main clay mineral. The age ranges from late Early Miocene to early Middle Miocene—zones N8 to N9, Late Burdigalian to Langhian. The section shows low biodiversity, including eleven macroinvertebrate taxa and two ichnotaxa; at its base, it contains mostly plant debris and amber from the continent, thus revealing a continental source near the littoral zone. Its middle portion preserves macroinvertebrates from the littoral zone, but also including the cephalopod Aturia sp., transported by a hydraulic flow to an area of greater depth; these macroinvertebrates are absent from overlying sections, at a time in which the basin was flooded most. The X-ray diffraction on six rock samples revealed mainly the presence of quartz with calcite, subordinated plagioclase and traces of potassium feldspar. Other minerals that compose this stratigraphic section are pyrite, dolomite and siderite, but they are disseminated in a siltstone. The section is dominated by mudstone sediments. The succession is kaolinite in the most pure fraction, and in lesser proportion smectite and illite; the stratified illite–smectite, chlorite and chlorite–smectite are present in low quantity. The mineralogical components of a molluscan valve were also examined, showing abundant pyrite, and moderate amounts of siderite and dolomite. In a part of the section, we recorded a rich assemblage of calcareous microfossils with foraminifers, nannofossils and marine ostracods. In the foraminiferal assemblage, 99 % correspond to planktonic forms. Most macroinvertebrates from benthonic assemblages suggest that they may have inhabited a near-shore marine environment, and sporadically experienced storm transport to deeper, shelf waters.
Similar content being viewed by others

Introduction
The Falcón Basin in northwestern Venezuela is an area of importance for the oil industry (e.g. González de Juana 1937, 1938; Wheeler 1963; González de Juana et al. 1980; Díaz de Gamero 1989) and is also known for a sedimentary sequence extending for most of the Neogene that preserves a diverse extinct fauna of evolutionary and biogeographic significance (e.g. Jung 1965; Sánchez-Villagra et al. 2010; Smith et al. 2010). The study of marine microfossils and macroinvertebrates in this area is potentially relevant to address several issues, including changes of marine communities in the neotropics (Hendy 2013), the evolution of the Caribbean Sea (Sánchez-Villagra et al. 2010), and changes of the fluvial systems in the northern portion of the continent (e.g. Díaz de Gamero 1996; Johnson et al. 2009; Aguilera et al. 2013). The ‘Urumaco Trough’, in western Falcon State, is an area of accumulation of thousands of metres of sediments which document changes in continental and marine deposits going back to the Eocene. Changes in the sedimentary succession mainly occurred in connection with the uplift of the Andes and the interactions of the southern boundary of the Caribbean plate with the South American plate. Among the Neogene units is the Querales Formation, towards the base of the Urumaco sequence (Quiroz and Jaramillo 2010).
The Querales Formation (Hodson 1926), with the type locality in the southwest of Sabaneta, represents a geological unit from the “surco de Urumaco” which consists mainly of large accumulations of laminated black clay–shales and silty shales, interbedded siltstone and to a lesser extent, by thin layers of fine sandstones (Díaz de Gamero 1989; Hambalek et al. 1994). A few microfossils from the Querales Formation have been reported (Gamero and Díaz de Gamero 1963; Díaz de Gamero 1977; Lorente 1986; Wozniak and Wozniak 1987), but the formation has not been studied in depth thus far. Here, we present a detailed palaeontological, sedimentological, and biostratigraphical study from the uppermost Querales Formation.
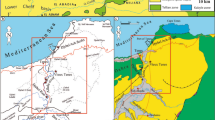
The studied site of the Querales Formation is located close to the road between Urumaco and Pedregal towns (Fig. 1). It is a succession of 17 m of a siliciclastic sequence with fine sandstones, laminated siltstones and massive to laminated dark shales. The site preserves a fossil assemblage with a low diversity of microfossils, macroinvertebrates, and other plant materials and ichnofossils. Most fossils are found in facies of the lower segment of the section. This new locality of 100 m of exposure along the quarry is named “Quebrada Corralito”, abbreviated ‘QC’ (Fig. 1). The complete Querales formation outcropping nearby along the Caduce river has a total thickness of 320 m (Quiroz and Jaramillo 2010). Here, we study the uppermost 17 m of the formation and we characterize sedimentologically and palaeontologically the deposits exposed at QC, and provide a biostratigraphic and palaeoenvironmental interpretation. These results are then discussed in the context of geological, palaeoenvironmental and palaeogeographical change in the Neogene Falcón Basin.

a Geographical setting of “Quebrada Corralito” (QC) quarry of the Querales Formation, Falcon Basin, northwestern Venezuela. Examined site is indicated with a star; GPS 11°07.180 N–70°13.092 W. b General view of the outcrop analysed. Geologic map after Quiroz and Jaramillo (2010)
Geological framework
Querales Formation
The Querales Formation was originally defined by Hodson (1926, p. 174) as “Querales Shales”, in Quebrada Querales, western tributary of Mitare River, Southwest of Sabaneta town, Falcón State. There have been nomenclatural and stratigraphical disagreements about the Querales Formation. Several authors include this unit as a lower portion of the Socorro Formation (Senn 1935; Liddle 1946; Gamero and Díaz de Gamero 1963). The top of the Querales Formation presents a transitional change into the Socorro Formation. Stainforth (1962) sub-divided the Querales Formation into two members: the lower Las Pilas Member, mainly composed of grey shales that crop-out in western Falcón Basin; and the upper Las Lomas Member, mainly composed by sandstones that crop-out in north eastern Falcón Basin. According to Díaz de Gamero et al. (1988) and Díaz de Gamero (1989), the Querales Formation extends all over the northern part of Falcón State and it is equivalent to the Las Pilas Member of Stainforth (1962). It is composed of more than 90 % dark-coloured shales with intercalations of fine-grained sandstones in packages up to 4 m thick, intensely burrowed, sparse and thin marlstone and limestone levels with mollusc shells. The shales become thicker upwards in and they yield concretions, ferruginous nodules and micro- and macrofossils. The one to four-metre-thick, fine-grained sandstones are locally silty and occasionally calcareous. They also show high degree of bioturbation that obliterates all previous sedimentary structures.
An earliest Middle Miocene age has been suggested for the Querales Formation (Wheeler 1963; González de Juana et al. 1980; perhaps, even latest Early Miocene, Díaz de Gamero 1989). Wheeler (1963) stated that the unit preserves the transgressive phase where deep marine sediments deposited in the western Falcón Basin. Díaz de Gamero (1989) hypothesized fluvio-deltaic to deltaic–marine conditions, while Quiroz and Jaramillo (2010) suggest shallow-marine, open sea conditions, typical of a prodeltaic/offshore-shelf environment. The deposits of the Querales Formation and those of the Cantaure Formation (exposed in the Paraguaná Peninsula, Jung 1965), may be partly correlatable and/or laterally equivalent (Hunter and Bartok 1974; Díaz de Gamero 1989).
Palaeontology of Querales Formation
Lorente (1986) studied palynomorphs from the Querales Formation and reported abundant Polypodiisporites usmensis, Polypodiisporites spp., and Mauritiidites franciscoi, and sparse occurrences of Malvacipolloides maristellae, Retitricolporites? irregularis, and Psilamonocolpites medius. Díaz de Gamero et al. (1988) recovered the planktonic foraminifers Globorotalia praemenardii, G. foshi peripheroronda, G. continuosa, Orbulina spp. among an abundant and highly diverse assemblage, bearing also Praeorbulina sp., Globigerinoides sp., Globigerina sp. and Globoquadrina sp. They also recorded some benthonic foraminifers and the calcareous nannofossils Coccolithus pelagicus, Discoaster spp., Helicosphaera ampliaperta, Reticulofenestra spp. and Sphenolithus heteromorphus.
There is extensive illustrated bibliographical information on Early Neogene molluscs in northwestern Venezuela and adjacent areas (e.g. Rutsch 1930, 1934; Senn 1935; González de Juana 1937; Miller and Thompson 1937; Ingram 1947; Weisbord 1962, 1964a, b; Jung 1965, 1966a, b; Gibson-Smith and Gibson-Smith 1974, 1983; Wesselingh et al. 2006; Landau and Marques da Silva 2010; Hendy et al. 2015), but hardly any for the Querales Formation, in which only the presence of molluscs has been reported. Hodson (1926) reported turritelids from this stratigraphic unit (cf. also Hodson et al. 1927; Hodson and Hodson 1931). Quiroz and Jaramillo (2010) recognized the presence of ichnofossils in the sandstone beds at the top of the sequences, including Thalassinoides and Planolites.
Materials and methods
We conducted sedimentological and palaeontological analyses of a detailed profile scale 1:25 at the “Quebrada Corralito” (QC) section, GPS: 11°07.180°N–70°13.092 W. We collected six fresh-rock samples for X-ray and microfossil analysis (Fig. 2).

Section logged in (QC) and stratigraphic location of the samples used for analysis of x-ray and microfossils, the levels with macrofossils are indicated with drawings in the column, and overall transgressive/regressive trend, denoted by arrows
For the sedimentological analysis, X-ray diffraction (XRD) was carried out on fine screen-washed sample material (<20 µm), measured with a PANalytical X’Pert PRO diffractometer, with Cu lamp (k = 1.5403 Å) operated at 40 mÅ and 40 kV at the Centro de Investigaciones Geológicas (CIG). The samples were measured from 2° to 40° 2, with a scan speed of 0.04°/s and a time per step of 0.50 s.
For calcareous microfossil analysis (foraminifera and ostracods), 0.2 kg of rock was smashed on an iron mortar, set to boiling point in a jar with tap water and 20 ml of hydrogen peroxide at 100 % volume, and then washed on a 64 µm sieve. Almost 99 % of the original rock was lost after washing it, and the remaining was picked under a stereo microscope. The recovered foraminifers are plotted in a Range Chart (Table 1). Planktonic foraminifers were photographed under the SEM of the Electron Microscopy Service of the Museo de La Plata, Argentina.
Calcareous nannofossil samples were prepared for study in a petrographic microscope with polarized light at a magnification of 1200×, using the standard gravity settling technique of Bramlette and Sullivan (1961). Some drops of Sodium-hexametaphosphate were added to the jar, so the clays could be deflocculated. Different time intervals (2, 10 and 45 min) of sediment deposition in the jar were controlled to concentrate the nannofossil content. Their relative abundances are plotted in a Range Chart (Table 1).
Macroinvertebrates were studied under light microscope and photographed using a digital camera. Measurements were taken according to Stanley (1970) using 0–150 mm digital calliper. Abbreviations used are L = length; H = height; W = width.
Abbreviations used in the text are: AMU-PI/MF/PB (AMU = Alcaldía del Municipio de Urumaco, PI = Colección de Paleontología Invertebrados; MF = Colección de Micropaleontología and PB = Colección Paleobotánica) Museo Paleontológico de Urumaco, Falcón State, República Bolivariana de Venezuela. NMB (Naturhistorisches Museum Basel), Switzerland.
Results
Sedimentology
The sediments of the QC are dominated by mudstones and blue–black shales, with low levels of highly bioturbated sandstones arranged in the base and the top of the section. These beds show massive to laminated levels with macroinvertebrates, microfossils and plant debris, with a layer containing small nodules of amber (Fig. 2). The QC section has a thickness of 17 m, and it is dominated by siliciclastic materials.
To better characterize the sedimentology and palaeontology of the studied section, we defined six lithofacies within it (Fig. 3; Table 2). The top of the section is covered by vegetation.

a Massive mudstone (Fm), showing the conchoidal and irregular fracture of the massive material; b Laminated and bioclastic mudstone (Fl), the arrow indicates the observed parallel lamination; c Laminated mudstone with plant debris (Ll), showing lamination and fragmentation; d Heterolithic lutite (pelite) (Htp); e Heterolithic facies (Hta), sandstones with trough cross-bedding, and thin levels of mudstones interbedded in strata thickening upwards; f Sandstone (Sm), amalgamated sandstone levels with signs of bioturbation (arrowed)
Rx analysis of the sediments
The X-ray diffraction provided information on lithological composition. This analysis was conducted on six rock samples (S) from the stratigraphic column representing the six facies described above (Fig. 2). The diffractogram of total rock (Table 3a) revealed mainly the presence of quartz (Qz) with calcite (Ca), subordinated plagioclase (Pl) and traces of potassium feldspar (Fsp). Other minerals that compose this stratigraphic section are the pyrite (Py), dolomite (D) and siderite (S), but they are disseminated in a siltstone.
The section is dominated by mudstone sediments; the X-ray analysis indicated that the main clay mineral is kaolinite (Table 3b). The succession is kaolinite (K) in the most pure fraction, and in lesser proportion smectite (Sm) and illite (I), the stratified illite–smectite (IS), chlorite (C) and chlorite–smectite (CS) are present in low quantity. The mineralogical component of a molluscan valve was also examined, showing abundant pyrite, and moderate amounts of siderite and dolomite.
Calcareous microfossils
Samples S2 and S6 are barren of microfossils. In samples S1, S4 and S5 only rare to few, poorly preserved, agglutinated foraminifera of the genus Ammobaculites were recovered. Sample S3, in contrast, yielded a rich assemblage of calcareous microfossils where foraminifers, calcareous nannofossils and marine ostracods were recovered.
Regarding the very abundant foraminifera assemblage, 99 % correspond to planktonic forms. The preservation varies from very good to poor, with frequent pyritization. This assemblage [AMU-MF-002] is composed of the planktonic species Globigerina praebulloides, Globigerinoides obliquus obliquus, Globigerinoides trilobus immaturus, Globigerinoides trilobus trilobus, Globoquadrina dehiscens, Globorotalia archaeomenardi, Praeorbulina glomerosa, and the benthonic Rosalina californica (Fig. 4; Table 1). The marine ostracods are represented by five species of the Cytherellidae and Cytheridae [AMU-MF-004].

Foraminifers recovered at Quebrada Corralito. (a–k, sample S3; l, sample S4). a Globorotalia archaeomenardi Bolli; b, Globigerina praebullooides Blow; c, Praeorbulina glomerosa (Blow); d, e Globoquadrina dehiscens (Chapman, Parr & Collins), apertural and side view; f Globigerinoides obliquus obliquus Bolli; g, h Globigerinoides trilobus trilobus (Reuss), apertural and side view; i, j Globigerinoides trilobus immaturus Bolli, spiral and side view; k Rosalina californica (Cushman); l Ammobaculites sp. Scale bar a, b, e, k = 50 µm; c, d, f, g, h, i, j = 100 µm; l = 200 µm
The calcareous nannofossil assemblage [AMU-MF-001] is of very low abundance (0.026 specimens per field of view) and the specimens have strong signs of etching and overgrowth. In some cases, it was impossible to determine them at species level. However, the assemblage has a high biodiversity index, due to the recognition of 19 species that are represented in almost the same relative abundance (Fig. 5; Table 1). These are the heterococcoliths Calcidiscus leptoporus, Coccolithus miopelagicus, C. pelagicus, Helicosphaera ampliaperta, H. obliqua, Reticulofenestra haqii, R. pseudoumbilicus, the nannoliths Discoaster deflandrei, D. druggii, D. exilis, D. formosus, D. sanmiguelensis, Discoaster sp. indet., cf. Catinaster calyculus, Scyphosphaera apsteinii, Sphenolithus heteromorphus, S. moriformis, Triquetrorhabdulus rugosus, and the thoracospherid Thoracosphaera heimii (Electronic Supplementary Material 1).

Some calcareous nannofossils recovered at Quebrada Corralito. All specimens from sample S3. a, b Coccolithus miopelagicus Bukry, 1971, emend. Wise, 1973; c Coccolithus pelagicus (Wallich, 1877) Schiller, 1930; d Reticulofenestra pseudoumbilicus (Gartner, 1967) Gartner, 1969; e Helicosphaera obliqua Bramlette & Wilcoxon, 1967; f–h Discoaster exilis Martini & Bramlette, 1963; i, j Discoaster formosus Martini & Worsley, 1971; k, l, Discoaster sanmiguelensis Bukry, 1981; m, n Discoaster druggii Bramlette & Wilcoxon, 1967; o, p cf. Catinaster calyculus Martini & Bramlette, 1963; q–s Sphenolithus heteromorphus Deflandre, 1953; t, u Sphenolithus moriformis (Brönnimann & Stradner, 1960) Bramlette & Wilcoxon, 1967; v Thoracosphaera heimii (Lohmann, 1919) Kamptner, 1941; w Watznaueria barnesae (Black in Black & Barnes, 1959) Perch-Nielsen, 1968, Cretaceous reworked heterococcolith; x Micrantholithus flos Deflandre in Deflandre & Fert, 1954, Paleogene reworked nannolith. Scale bar 2 µm
Fossil macroinvertebrates
Three layers containing macroinvertebrates were recognized at the QC (Fig. 2). In the lower one (S2), a low-diversity association of molluscs, crab fragments and one brachiopod was recovered; whereas in the upper layers, the association was composed only by sparse bivalves and gastropods. In the three layers, specimens are enclosed in a massive matrix, with complete valves and in some cases in butterfly position. In almost all cases, the fossil macroinvertebrates are represented by complete specimens that lack signals of interactions with epibiotic organisms (bioeroders and/or encrusters) and without any marked taphonomical alteration. The specimens are usually well preserved and articulated, being mostly casts and moulds, and occasionally presenting pyrite-coated surfaces (S4).
We identified eleven molluscan taxa, after comparative work at the NMB collection: Bivalves, Anadara sp. [AMU-PI-001], Trachycardium sp. [AMU-PI-002], Cyclinella cf. cyclica [AMU-PI-003]; Psammacoma cf. falconensis [AMU-PI-004], Tellina sp. [AMU-PI-005], Veneridae indet.1 [AMU-PI-006], Veneridae indet.2 [AMU-PI-007], the gastropods ‘Turritella’ cf. matarucana [AMU-PI-008], Oliva sp. [AMU-PI-009], Natica sp. [AMU-PI-010] and one nautiloid fragment of Aturia sp. (see Klug and Etter 2012, Fig. 4.12; cf. also Miller and Thompson 1937, pls. 8–10; Jung 1966b, pls. 1–2). The other macroinvertebrates recovered correspond to small and fragmentary valves of the brachiopod Glottidia sp. [AMU-PI-011] (Fig. 6) and carapace and pincer fragments of crabs (Fig. 7) referred to the Plagusiidae, Callianassidae of the genus Neocallichirus and Calappidae (Electronic Supplementary Material).

Examples of the macroinvertebrate record and of amber from Quebrada Corralito, Querales Formation. Mollusc bivalves; a Anadara sp. [AMU-PI-001] (left valve); b Trachycardium sp. [AMU-PI-002] (right valve, internal mould); c Cyclinella cf. cyclica (Guppy, 1866) [AMU-PI-003] (left valve, internal mould); d Psammacoma cf. falconensis (H.Hodson, 1931) [AMU-PI-004] (right valve, steinkern); e Tellina sp. [AMU-PI-005] (right valve, steinkern); f Veneridae indet.1 [AMU-PI-006] (left valve, internal mould); g Veneridae indet.2 [AMU-PI-007] (right valve). Gastropods; h ‘Turritella’ cf. matarucana F.Hodson, 1926 [AMU-PI-008] (steinkern with shell remains); i Oliva sp. [AMU-PI-009] (steinkern); j Natica sp. [AMU-PI-010] (steinkern with shell remains). Cephalopod; k Aturia sp (lateral view of broken phragmocone, suture partly visible). Linguliform brachiopod; l Glottidia sp. [AMU-PI-011] (pair of divergent internal septa just discernible through semi-translucent shell); m Amber nodule [AMU-PB-1]. Scale bar l = 0.5 cm; a, e, i, j, k, m = 1 cm; b, c, d, f, g, h = 2 cm

Crustaceans from Quebrada Corralito, Querales Formation. Decapods; a–f ?Plagusiidae (a dorsal, b ventral, c in situ mould of carapace, d dorsal, e ventral, f in situ mould of carapace), g Calappidae in situ; h–i Neocallichirus (outer and inner lateral views of cheliped). Scale bar 1 cm
Fossil plants
One layer (S2) containing plant debris was recorded in the lower section of the stratigraphic column (Fig. 2). In this layer, corresponding with facies Ll, there is a dense accumulation of non-identifiable and charcoalized plant debris. In this bed, we also found small nodules of amber of dark brown colouration, translucid and lacking visible organisms inside (Fig. 6). This record of amber from Venezuela is one of the few from this part of the world together with the reports from Colombia (only copal), Peru, and Brazil and the well-known Dominican Republic one (Antoine et al. 2006; Feist et al. 2007; Hinojosa-Díaz and Engel 2007; Pereira et al. 2009; Judson 2010). Karsten (1851) mentioned the finding of amber in the Falcón Basin, from an imprecise area and unknown Formation. Karsten (1851, p. 445) reported amber in a geologic formation with coal (‘Zur tertiären Formation gehören Paraguaná, der grösste Theil des nördlichen Coro, soweit es das Gebiet zwischen den älteren Inselbergen ausfüllt (eine Kohlenformation in der sich Bernstein findet’)). The second reference to amber, from the same publication, concerns La Mesa, close to Carora, Lara State, south of the Baragua River. He mentioned abundant fragments of amber, in different sizes, underlying a coal layer and also isolated in the sand of the river, and stated that there are excellent and well-preserved fragments of yellow and light-brown amber with wasps and bees embedded inside.
Discussion
Sedimentology
In accordance to previous characterizations of the Querales Formation (Hodson 1926), the type section is dominated by siliciclastic accumulations of fine-grained sediments originated from a transgressive–regressive event. The Quebrada Corralito (QC) section represents a complete relative sea-level cycle, located at the top of the unit. The geographic and stratigraphic position of the QC section indicates that this is the last phase of accumulation of the Querales Formation in the area.
The QC section starts at the base with a prominent bed of massive sandstones (F-Sm) which were probably deposited in coastal areas with influence from waves (lower to middle shoreface). The ichnofossils Thalassinoides and Planolites indicate an environment of high productivity and with well-oxygenated sediments (Buatois and Mángano 2011). In these facies, we identified Thalassinoides isp. and Planolites isp. Above this psammitic layer, in a transitional passage, the laminated mudstone (F-Ll) preserves plant debris and nodules of amber; the latter having been transported by currents that originated in the continent and which entered a low-mid energy environment (shoreface). In this distal setting of low energy, the environment was little affected by water currents, as in deltaic fronts in which plant remains and resin accumulate (Grimaldi 1996).
This gradual change from a coastal environment to a relatively deeper one (shoreface to offshore) is reflected in the overlying facies (Fl and Fm), respectively. We recorded the largest concentration of macroinvertebrates of the section in Fl, that is characteristic of littoral environments, and in the fine-grained matrix of the massive layer we recorded few fossils of pelagic organisms. The fossils found in the upper levels of the section, together with the sedimentological and mineralogical features recorded, suggest that the studied section represents times of maximal flooding of the basin. This middle segment of the stratigraphic column is mostly non-fossiliferous, yielding only a fragmentary specimen of the nautiloid Aturia sp. These homogenous levels with little clastic components have been deposited mainly via decantation in the inner and outer platform (offshore). The geochemical analysis supports the hypothesis of accumulation via decantation in an inner platform, with the presence of pyrite in facies Fl and Fm (S3 and S4) and the replacement of the original shells of invertebrates. These facies were formed in a reductive environment, which facilitated the formation of ion sulphur characteristic of anoxic conditions in the sediment found below the line of compensation of calcium carbonate.
In the upper part of the section, the facies change gradually from (Fm) to (Htp) and (Hta). This gradual change is characterized by an increase in the grain size towards the top, all lacking organic remains. In this portion of the section, the depositional conditions suggest that the process of lowering of the sea level (regression) was coupled with an increase of clastic materials and the presence of thin sandstone layers with trough cross-bedding, instead of laminated shales. The increase of transported material together with the decrease in deposition of sediments by decantation suggests the change from an offshore transition to low shoreface environment.
Calcareous microfossils
According to the recovered calcareous microfossils in the QC, it is possible to identify two different marine environments along the succession. The lower and upper levels (samples S1–S2 and S4–S6) represent shallow, marginal-marine environments, where planktonic foraminifera and calcareous nannofossils are absent, and only rare agglutinated benthonic foraminifera of the genus Ammobaculites were recorded (Fig. 4). The species of the genus Ammobaculites are infaunal detritivores that frequently dominate brackish marshes, lagoons and other marginal-marine temperate to tropical environments (Murray 1991, 2006).
The middle part of the succession (sample S3) is interpreted as an outer shelf marine environment (over 200 m deep) on the basis of the high proportion of planktonic foraminifera against benthonic ones, and the occurrence of a poorly preserved but diverse calcareous nannofossil assemblage. Pyrite granules and pyritization observed in some calcareous microfossil specimens, together with the strong dissolution that affected the poorly preserved calcareous nannofossil assemblage, suggest deep marine waters with very low energy and anoxic/dysoxic conditions near the sea floor. The ostracods, as the molluscs, were probably transported from coastal areas.
The calcareous microfossils also allowed to establish a reliable age for this locality on the basis of the assemblage recovered in sample S3. Both planktonic foraminifera and the calcareous nannofossils suggest a late Early Miocene to early Middle Miocene age. The most abundant species of planktonic foraminifera are those of the genus Globigerinoides, a widely observed characteristic of the planktonic foraminiferal assemblages that occur after the Oligocene/Miocene boundary (Bolli and Saunders 1985). On the basis of the biostratigraphic ranges of the planktonic foraminifers found in this section (Electronic Supplementary Material, Planktonic Foraminiferal Zones), the assemblage is interpreted to fall into the Zones N8–N9 of Berggren and van Couvering (1974). The recovered calcareous nannofossil assemblage is consistent with the planktonic foraminifer’s age interpretation. The marker species Helicosphaera ampliaperta, Discoaster exilis, Sphenolithus moriformis and Triquetrorhabdulus rugosus suggest an assignation to the Zones CN3–CN4 of Okada and Bukry (1980) for the assemblage (Electronic Supplementary Material, Calcareous Nannofossil Zones). Díaz de Gamero et al. (1988) also interpreted the same age for the Querales Formation in its type area, based on a more diverse assemblage of planktonic foraminifera.
Macroinvertebrates
There is a low-diversity association of macroinvertebrates that yielded only eleven taxa and two ichnotaxa, corresponding mainly to a littoral marine fauna. The assemblage found in this site corresponds to a heterogeneous palaeoenvironment, with a mixture of species characterized by different ecological features, occurring in a chaotic arrangement in a small segment of the Quebrada Corralito section. Based on comparison with extant close relatives of the recorded taxa (Stanley 1970), the taxa represent different guilds (Aberhan 1994; Brenchley and Harper 1998 del Río et al. 2001; Lazo et al. 2005).
Endobenthonic: with shallow infaunal forms, including shallow-burrowing suspension feeders (Glottidia sp., Trachycardium sp., Anadara sp., Cyclinella cf. cyclica, Veneridae indet.1 and Veneridae indet.2); deep-burrowing suspension/detritus feeders (Psammacoma cf. falconensis, Tellina sp.); and deep-burrowing deposit feeders (Callianassidae).
Epibenthonic: including mobile carnivores (Oliva sp., Natica sp., Calappidae, and ?Plagusiidae), and a mobile epi-to semi-infaunal suspension/detritus feeder or browser (‘Turritella’ cf. matarucana; Allmon 1988).
Nekto-benthonic: with a single representative of scavengers (Aturia sp.).
The great majority of the macroinvertebrate taxa sampled were deposited in an environment untypical for the palaeoecology of the groups they represent. The sampled bivalves inhabit in near-shore (littoral) zones of warm seas with normal salinity, in areas with sandy and granular sediments, especially the species capable of burrowing themselves relatively deep and which feed by filtering using their syphons. All the bivalves were preserved with their shells in external view, and in some cases, with both valves joined by the ligament in “butterfly” pattern, typical of a post-mortem deposition after a rapid and short-distance transportation from the original living area. The gastropods are also organisms that inhabit granular and soft sediments on the sea floor. The same is valid for the sampled crustaceans, especially the callianasid, which build deep gallery systems in sandy sediments, lining them with mudstone pellets to avoid the collapse of the galleries due to movement and erosion produced by waves. Finding these fossils in a mudstone matrix suggests that they have been transported, with a change in the abundance and preservation of remains, which decreases from the Fl facies to the Fm facies, in which almost no macroinvertebrates have been recorded. Only the occurrence of Aturia sp. represents an organism which inhabited the area where the sediments were deposited, as this taxon had nekto-benthonic habits.
The fossil macroinvertebrates consist only of individual specimens, with no records of interactions with epibionts and bioeroders, most of the preserved forms being burrowers. This may be explained by sporadic and sudden storms, which resulted in scour and transport of bioclastic materials from the littoral zone to deeper areas of the marine platform. The evidence provided by scattered, disarticulated, very fragile, single valves of linguloid brachiopods is also compatible with a brief, storm-derived, transport from a shallower, subtropical, environmental setting although some extant species may inhabit up to greater, bathyal, depths (Emig et al. 1978; Emig 1986, 1997).
Interpretation of depositional environments
The fine-grained sediments of the Quebrada Corralito section resulted from a transgressive–regressive event in a deltaic system which came into contact with a carbonate platform and generated the resulting deposits. This combination of the two systems helps to explain the recorded mixture of fossil organisms and the differences between the upper and lower facies of the upper Querales Formation. The kind of preservation of both macroinvertebrates and microinvertebrates suggests that transport over long distances was not involved, as is clear, for example, in the case of the microfossils from S3. The deposition of sediments started in a deltaic system which faced a marine front, with the subsequent mixed accumulation of organisms. Plant debris and amber were transported from the continent and mixed with marine elements. The palaeontological and sedimentological changes recorded in the section relate to transgressive–regressive phases along it.
Conclusions
-
1.
The studied section shares many features and overall the same kind of depositional environment with the type section of the Querales Formation.
-
2.
Low faunal diversity is present in the section, the base of which mostly contains plant debris and amber from the continent, indicating a continental source near the littoral zone. Whereas the middle portion preserves macroinvertebrates from the littoral zone, transported by a hydraulic flow to an area of greater depth, macroinvertebrates are absent from overlying sections, at a time in which the basin was flooded most.
-
3.
The age the of upper Querales Formation can range from late Early Miocene to early Middle Miocene (zones N8 to N9, late Burdigalian to Langhian).
-
4.
The Quebrada Corralito section preserves one cycle of transgressive–regressive phases of the deltaic environments facing the marine platform.
References
Aberhan, M. (1994). Guild-structure and evolution of Mesozoic benthic shelf communities. Palaios, 9, 516–545.
Aguilera, O. A., Lundberg, J., Birindelli, J., Sabaj Pérez, M., Jaramillo, C., & Sánchez-Villagra, M. R. (2013). Palaeontological evidence for the last temporal occurrence of the ancient western Amazonian river inflow into the Caribbean. PLoS One, 8(9), e76202. doi:10.1371/journal.pone.0076202.
Allmon, W. D. (1988). Ecology of Recent Turritelline gastropods (Prosobranchia, Turritellidae): current knowledge and paleontological implications. Palaios, 3, 259–284.
Antoine, P.-O., De Franceschi, D., Flynn, J. J., Nel, A., Baby, P., Benammi, M., et al. (2006). Amber from western Amazonian reveals Neotropical diversity during the middle Miocene. Proceedings of the National Academy of Science USA, 103, 13595–13600.
Berggren, W. A., & Van Couvering, J. A. (1974). The late Neogene biostratigraphy, geochronology and paleoclimatology of the last 15 million years in marine and continental sequences. Palaeogeography, Palaeoclimatology, Palaeoecology, 16, 1–216.
Bolli, H. M., & Saunders, J. B. (1985). Oligocene to Holocene low latitude planktic foraminifera. In H. M. Bolli, J. B. Saunders & K. Perch-Nielsen (Eds.), Plankton Stratigraphy (pp. 155–262). Cambridge: Cambridge University Press.
Bramlette, M. N., & Sullivan, F. R. (1961). Coccolithophorids and related nannoplankton of the early Tertiary in California. Micropaleontology, 2, 129–188.
Brenchley, P. J., & Harper, D.A.T. (1998). Palaeoecology: ecosystems, environments and evolution (402 pp.). London: Chapman & Hall.
Buatois, L. A., & Mángano M. G. (2011). Ichnology organism-substrate interactions in space and time (366 pp.). Cambridge: Cambridge University Press.
del Río, C., Martínez, S. A., & Scasso, R. A. (2001). Nature and origin of spectacular marine Miocene shell beds of northeastern Patagonia (Argentina): Paleoecological and bathymetric significance. Palaios, 16, 3–25.
Díaz de Gamero, M. L. (1977). Revisión de las unidades litoestratigráficas en Falcón central, en base a su contenido de foraminíferos planctónicos. Congreso Geológico de Venezuela, Memorias, I, 81–86.
Díaz de Gamero, M. L. (1989). El Mioceno Temprano y Medio de Falcón Septentrional. GEOS, 29, 25–35.
Díaz de Gamero, M. L. (1996). The changing course of the Orinoco River during the Neogene: a review. Palaeogeography, Palaeoclimatology, Palaeoecology, 123, 385–402.
Díaz de Gamero, M. L., Mitacchione, V., & Ruíz, M. (1988). La Formación Querales en su área tipo, Falcón Noroccidental, Venezuela. Boletín de la Sociedad Venezolana de Geólogos, 34, 34–46.
Emig, C. C. (1986). Conditions de fossilisation du genre Lingula (Brachiopoda) et implications paléoécologiques. Palaeogeography, Palaeoclimatology, Palaeoecology, 53, 245–253.
Emig, C. C. (1997). Ecology of inarticulated brachiopods. In R. L. Kaesler (Ed.), Treatise on invertebrate paleontology. Part H (revised), vol. 1 (pp. 473–495). Lawrence: Geological Society of America & University of Kansas.
Emig, C. C., Gall, J. C., Pajaud, D., & Plaziat, J. C. (1978). Réflexions critiques sur l’ écologie et la systématique des lingules actuelles et fossiles. Géobios, 11(5), 573–609.
Feist, M., Lamprecht, I., & Müller, F. (2007). Thermal investigations of amber and copal. Thermochimica Acta, 458(1–2), 162–170.
Gamero, G. A., & Díaz de Gamero, M. L. (1963). Estudio de una sección de referencia de las formaciones Cerro Pelado y Socorro en la región de El Saladillo, Estado Falcón. GEOS, 9, 7–44.
Gibson-Smith, J., & Gibson-Smith, W. (1974). The genus Strombina in Venezuela, with description of a new Recent and some fossil species. Boletín Informativo de la Asociación Venezolana de Geología, Minería y Petróleo, 17(4–6), 49–85.
Gibson-Smith, J., & Gibson-Smith, W. (1983). Neogene melongenid gastropods from the Paraguaná Peninsula, Venezuela. Eclogae geologicae Helvetiae, 76(3), 719–728.
González de Juana, C. (1937). General Geology and Stratigraphy of the Cumarebo Area, State of Falcón. Boletín de Geología y Minería, 1, 187–205.
González de Juana, C. (1938). Contribución al estudio de la cuenca sedimentaria Zulia-Falcón. Boletín de Geología y Minería, 2, 123–138.
González de Juana, C., Iturralde de Arozena, J. M., & Picard Cadillat, X. (1980). Geología de Venezuela y de sus cuencas petrolíferas. Tomo II. (pp. 415–1031). Caracas.
Grimaldi, D. A. (1996). Amber: windows to the past. New York: Harry N. Abrams. 216 pp.
Hambalek, N., Rull, V., de Digiacomo, E., & Díaz de Gamero, M. L. (1994). Evolución paleoecológica y paleoambiental de la secuencia del Neógeno en el Surco de Urumaco, estado Falcón. Estudio palinológico y litológico. Boletín de la Sociedad Venezolana de Geólogos, 19, 7–19.
Hendy, A. (2013). Spatial and stratigraphic variation of marine paleoenvironments in the Upper Miocene Gatun Formation, Isthmus of Panama. Palaios, 28, 210–227.
Hendy, A. J. W., Jones, D. S., Moreno, F., Zapata, V., & Jaramillo, C. (2015). Neogene molluscs, shallow marine paleoenvironments, and chronostratigraphy of the Guajira Peninsula, Colombia. Swiss Journal of Palaeontology, 134, 45–75. doi:10.1007/s13358-015-0074-1.
Hinojosa-Díaz, I. A., & Engel, M. S. (2007). A new fossil orchid bee in Colombian copal (Hymenoptera: Apidae). American Museum Novitates, 3589, 1–7.
Hodson, F. (1926). Venezuelan and Caribbean turritellas; with a list of Venezuelan type stratigraphic localities. Bulletins of American Paleontology, 11(45), 171–220.
Hodson, F., & Hodson, H. K. (1931). Some Venezuelan mollusks. Bulletins of American Paleontology, 16(59–60), 1–132.
Hodson, F., Hodson, H. K., & Harris, G. D. (1927). Some Venezuelan and Caribbean mollusks. Bulletins of American Paleontology, 13(49), 1–160.
Hunter, V. F., & Bartok, P. (1974). The age and correlation of the Tertiary sediments of the Paraguaná Peninsula, Venezuela. Boletín Informativo de la Asociación Venezolana de Geología, Minería y Petróleo, 17, 143–154.
Ingram, W. M. (1947). New fossil Cypraeidae from Venezuela and Colombia. Bulletins of American Paleontology, 31(121), 1–12.
Johnson, K. G., Sánchez-Villagra, M. R., & Aguilera, O. (2009). The Oligocene–Miocene transition on coral reef in the Falcón Basin (NW Venezuela). Palaios, 24, 59–69.
Judson, M. L. I. (2010). Redescription of Chelifer eucarpus DALMAN (Arachnida, Chelonethi, Withiidae) and first records of pseudoscorpions in copal from Madagascar and Colombia. Palaeodiversity, 3, 33–42.
Jung, P. (1965). Miocene mollusca of the Paraguana Peninsula, Venezuela. Bulletins of American Paleontology, 49(223), 385–652.
Jung, P. (1966a). Miocaene Mollusken von der Halbinsel Paraguaná, Venezuela. Verkürzte Fassung der Inauguraldissertation zur Erlangung der Würde eines Doktors der Philosophie vorgelegt der Philosophisch-Naturwissenschaftlichen Fakultät der Universität Basel, 16 pp.
Jung, P. (1966b). Zwei miocaene Arten von Aturia (Nautilaceae). Eclogae Geologicae Helvetiae, 59(1), 485–492.
Karsten, H. (1851). Über die geognostischen Verhältnisse des nördlichen Venezuela. Archiv Mineralogie, Geognosie, Bergbau und Hüttenkunde, 24, 440–479.
Klug, C., & Etter, W. (2012). Los cefalópodos: Los ammonites y sus parientes. In M. R. Sánchez-Villagra (Ed.), Venezuela Paleontológica, evolución de la diversidad en el pasado geológico (378 pp.). Printworkart Press, Zurich.
Landau, B., & Marques da Silva, C. (2010). Early Pliocene gastropods of Cubagua, Venezuela: taxonomy, paleobiogeography and ecostratigraphy. Palaeontos, 19, 1–221.
Lazo, D. G., Cichowolski, M., Rodríguez, D. L., & Aguirre-Urreta M. B. (2005). Lithofacies, palaeoecology and palaeoenvironments of the Agrio Formation, Lower Cretaceous of the Neuquen Basin, Argentina. In G. D. Veiga, L. A. Spalletti, J.A. Howell., & Schwarz. E (Eds.), The Neuquén Basin, Argentina: A case study in sequence stratigraphy and basin dynamics. Geological Society, London, Special Publications, 252, 295–315.
Liddle, R. A. (1946). The Geology of Venezuela and Trinidad (2nd ed.). Ithaca: Paleontological Research Institute. 890 p.
Lorente, M. A. (1986). Palynology and Palynofacies of the upper Tertiary in Venezuela. Dissertationes Botanicae, 99 (p. 224). Berlin: J. Cramer.
Miller, A. K., & Thompson, M. I. (1937). Beiträge zur Kenntnis tropisch-amerikanischer Tertiärmollusken. Teil VI. Some tertiary Nautiloids from Venezuela and Trinidad. Eclogae Geologicae Helvetiae, 30(1), 59–73.
Murray, J. W. (1991). Ecology and paleoecology of benthic foraminifera (p. 397). Avon: Longman Scientific & Technical.
Murray, J. W. (2006). Ecology and applications of benthic foraminifera (p. 426). Cambridge: Cambridge University Press.
Okada, H., & Bukry, D. (1980). Supplementary modification and introduction of code numbers to the low-latitude coccolith biostratigraphic zonation (Bukry, 1973; 1975). Marine Micropaleontology, 5, 321–325.
Pereira, R., de Souza Carvalho, I., Simoneit, B. R. T., & de Almeida Azevedo, D. (2009). Molecular composition and chemisystematic aspects of Cretaceous amber from Amazonas, Araripe and Recôncavo basins, Brazil. Organic Geochemistry, 40, 863–875.
Quiroz, L., & Jaramillo, C. (2010). Stratigraphy and sedimentary environments of Miocene shallow to marginal marine deposits in the Urumaco trough, Falcon basin, western Venezuela. In M. R. Sánchez-Villagra, O. Aguilera, & A. Carlini (Eds.), Urumaco and Venezuelan paleontology, the fossil record of the northern neotropics (304 p.). Bloomington: Indiana University Press.
Rutsch, R. F. (1930). Einige interessante Gastropoden aus dem Tertiär der Staaten Falcón und Lara (Venezuela). Eclogae Geologicae Helvetiae, 23(2), 604–614.
Rutsch, R. F. (1934). Die Gastropoden aus dem Neogen der Punta Gavilan in Nord-Venezuela. I–II. Abhandlungen der schweizerischen paläontologischen Gesellschaft, 54–55, 1–169.
Sánchez-Villagra, M. R., Aguilera, O. A., & Carlini, A. A. (Eds.). (2010). Urumaco and Venezuelan paleontology – the fossil record of the northern neotropics (304 p.). Bloomington: Indiana University Press.
Senn, A. (1935). Die stratigraphische Verbreitung der Tertiären Orbitoiden, mit spezieller Berücksichtigung ihres Vorkommens in Nord-Venezuela und Nord-Marokko. Eclogae Geologicae Helvetiae, 28(1), 51–113, 369–373.
Smith, C., Collins, L., Jaramillo, C., & Quiroz, L. (2010). Marine paleoenvironments of Miocene–Pliocene formations of north-central Falcón State, Venezuela. Journal of Foraminiferal Research, 40, 266–282.
Stainforth, R. M. (1962). Definitions of some new stratigraphic units in western Venezuela: Las Pilas, Cocuiza, Vergel, EL Jebe, Tres Esquinas and Nazaret. Asociación Venezolana de Geología, Mineralogía y Petrología, Boletín Informativo, 5(10), 279–282.
Stanley, S. M. (1970). Relation of shell form to life habits of the bivalvia (Mollusca). The Geological Society of America, Memoir, 125, 1–296.
Weisbord, N. E. (1962). Late Cenozoic gastropods from northern Venezuela. Bulletins of American Paleontology, 42(193), 1–672.
Weisbord, N. E. (1964a). Late Cenozoic pelecypods from northern Venezuela. Bulletins of American Paleontology, 45(204), 1–564.
Weisbord, N. E. (1964b). Late Cenozoic scaphopods and serpulid polychaetes from northern Venezuela. Bulletins of American Paleontology, 47(214), 111–199.
Wesselingh, F. P., Hoorn, M. C., Guerrero, J., Räsänen, M., Romero, Pittmann L., & Salo, J. (2006). The stratigraphy and regional structure of Miocene deposits in western Amazonia (Peru, Colombia and Brazil), with implications for Late Neogene landscape evolution. Scripta Geologica, 133, 291–322.
Wheeler, C. B. (1963). Oligocene and Lower Miocene stratigraphy of Western and Northeastern Falcón Basin, Venezuela. Bulletin of the American Association of Petroleum Geologists, 47, 35–68.
Wozniak, J., & Wozniak, M. H. (1987). Bioestratigrafía de la región nor-central de la Serranía de Falcón, Venezuela nor-occidental. Boletín de Geología, 16, 101–139.
Acknowledgments
We thank the Rodolfo Sánchez family for their hospitality during fieldwork, Jorge González and Jorge Carrillo-Briceño for their collaboration in the design of the figures, Carlos Jaramillo and Luis Quiroz for informative discussions on the geology of the Urumaco region, Francisco Vega-Vera for kindly providing comparative material of crustaceans, and both anonymous reviewers for their valuable comments which helped to improve this manuscript. The SEM Microscope operators of the Electron Microscopy Service of the Museo de La Plata, Dr. D. Poire and X-ray services at CIG-CONICET, for assistance in the analysis of rock samples collected in the field, and the Alcaldía of Urumaco and the Instituto del Patrimonio Cultural for the permission to work in Urumaco. We thank the students of the University of Zurich of the 2011 and 2013 excursions for their assistance in the fieldwork, Dr. Loïc Costeur from the Naturhistorisches Museum of Basel Museum for access to collections deposited there and Jorge Carrillo Briceño for assistance in the field and in curatorial tasks in Urumaco. This work was supported by the Swiss NF grant 31003A-149605 to MRS-V.
Author information
Authors and Affiliations
Corresponding authors
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Pérez, L.M., Pérez Panera, J.P., Aguilera, O.A. et al. Palaeontology, sedimentology, and biostratigraphy of a fossiliferous outcrop of the Early Miocene Querales Formation, Falcón Basin, Venezuela. Swiss J Palaeontol 135, 187–203 (2016). https://doi.org/10.1007/s13358-015-0105-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-015-0105-y