
Abstract
A new, marine osteichthyan (bony fish) fauna from the Early Triassic of northern India is presented. The material was collected in situ at localities within Pin Valley (Lahaul and Spiti District, Himachal Pradesh, India) and is dated as middle-late Dienerian (one specimen possibly earliest Smithian). The new ichthyofauna includes a lower jaw of the predatory basal ray-finned fish Saurichthys, a nearly complete specimen of a parasemionotid neopterygian (cf. Watsonulus cf. eugnathoides), as well as further articulated and disarticulated remains (Actinopterygii indet., Actinistia indet.), and thus comprises the most complete Triassic fish fossils known from the Indian subcontinent. Saurichthys is known from many Triassic localities and reached a global distribution rapidly after the Late Permian mass extinction event. Parasemionotidae, a species-rich family restricted to the Early Triassic, also achieved widespread distribution during this epoch. Comparison of the Spiti material with other parasemionotid species reveals similarities with Watsonulus eugnathoides from Madagascar. However, taxonomic ambiguities within Parasemionotidae prevent a specific attribution of the Spiti specimen. The new material also includes an isolated actinistian urohyal exhibiting morphology distinct from any previously described urohyal. Marine Dienerian black shale deposited on continental shelves are common not only in the Himalayas but also in other geographic regions. Anoxic depositional settings provide ideal preservational conditions for vertebrate fossils, suggesting that additional ichthyofaunas could still be discovered in marine Dienerian strata of other localities. The study of Early Triassic fish assemblages, including the presented one, is fundamental for our understanding of the great osteichthyan diversification after the Late Permian mass extinction event.
Similar content being viewed by others


Introduction
Bony fishes flourished in the wake of the Late Permian mass extinction event, reaching a distinct diversity peak in the Middle Triassic (e.g. Romano et al. 2014a; Scheyer et al. 2014; Tintori et al. 2014; Friedman 2015). While research on Middle Triassic fishes recently experienced a renaissance following the discovery of new fossiliferous assemblages (e.g. Luoping biota, South China; Hu et al. 2010), knowledge of Early Triassic ichthyofaunas has been improving at a much slower pace. One reason for this discrepancy is the remote location of classical Early Triassic fish Lagerstätten, i.e. Greenland, Madagascar, Spitsbergen (Svalbard, Norway), and British Columbia (Canada), implicating logistically difficult and often expensive fieldwork (Scheyer et al. 2014). Moreover, for most of these Lagerstätten the pinnacle of research took place several decades ago (e.g. Stensiö 1921, 1925, 1932; Piveteau 1934, 1939–1940; Nielsen 1936; Lehman 1952; Lehman et al. 1959; Beltan 1968) and many of the described taxa are in need of revision and/or lack detailed biostratigraphic information. Even though other localities yielding marine and freshwater bony fishes of Early Triassic age are frequent (e.g. López-Arbarello 2004; Brinkmann et al. 2010; Romano et al. 2014a), most of them have not proven to be rich in well-preserved fossils. A geographic province that has a particularly poor record of Triassic fishes in spite of the widespread occurrences of suitable strata is the Himalayan Range (Deecke 1927; Chang and Miao 2004).
Here we present a previously unknown Early Triassic marine bony fish fauna from Pin Valley (Lahaul and Spiti District, Himachal Pradesh, India), a collection area which has the potential for further discoveries. Within Gondwana, the present-day Indian subcontinent includes some of the largest numbers of sites producing Early Triassic chondrichthyan and osteichthyan remains, but most localities yield only microfossils or rare and incomplete larger specimens, often with very limited biochronostratigraphic age constraints (see “Discussion”). The new ichthyofauna from Spiti not only includes the most complete macrofossils ever described from the Triassic of India (both marine and freshwater), it is also complemented with a detailed biostratigraphic framework based on ammonoids (Brühwiler et al. 2010; Ware et al. 2015, in production a, b) and conodonts (e.g. Goel 1977; Krystyn et al. 2004; Goudemand 2010, 2014). Future excavations in Spiti and neighbouring areas may thus help widen our understanding of Gondwanan fish faunas at the time of recovery after the end-Permian mass extinction event. The aim of this paper is to describe the first finds of Early Triassic fishes from Spiti, which have been recovered during two short field trips.
Material and geological setting
The specimens described below were recovered from different sites within Pin Valley, Spiti area, northern India (Fig. 1). The Spiti Subdistrict of Himachal Pradesh is famous for its well-exposed Early Triassic ammonoid- and conodont-rich sections (e.g. Diener 1897, 1912; Hayden 1904; Krafft and Diener 1909; Goel 1977; Krystyn et al. 2004; Goudemand 2010; Brühwiler et al. 2010, 2012; Ware et al. in production a), but the occurrence of fish macrofossils within these strata was unknown until 2009, when two articulated specimens were discovered near the village of Mud (cf. Fig. 1) in sediments of Dienerian age (late Induan, Early Triassic; we herein use the Early Triassic subdivision of Tozer 1965). In 2010, additional fieldwork was conducted by members of the PIMUZ to explore the fish richness of Dienerian strata at three selected localities near the villages of Guling, Mud, and Tilling (Pin Valley, Fig. 1). The excursion was successful and yielded an additional articulated individual as well as numerous fragmentary remains. The material recovered during the 2009 and 2010 expeditions is curated by the PIMUZ.

a Map of Early Triassic Pangaea (modified from PALEOMAP project, www.scotese.com), with indication of the approximate location of Spiti (white star) on the Northern Indian Margin (Gondwana, Neotethys realm), b map of present-day South Asia, with Himachal Pradesh of India indicated as dark grey area, c map of Himachal Pradesh (enlarged from b), with Lahaul and Spiti District highlighted; d locality map of Pin Valley and surrounding areas within the Spiti Subdistrict (indicated as black frame in c). White stars in d indicate the three finding sites near the villages of Guling, Mud, and Tilling, where the described fish material was found. Exposures of the Early Triassic and Anisian Mikin Formation are indicated in dark grey and main roads of Spiti and Pin Valley by dashed lines in d
All specimens were found in situ within the lower part of the Limestone and Shale Member of the Mikin Formation, which is dated as middle Dienerian to earliest Smithian on the basis of ammonoids (e.g. Brühwiler et al. 2010; Ware et al. 2015, in production a; Fig. 2). The stratigraphic interval from which the fishes come from is often referred to as the “Meekoceras beds” (e.g. Krafft and Diener 1909), the “Gyronites beds” (e.g. Krystyn et al. 2004) or the “Ambites beds” (Brühwiler et al. 2010). It consists of black shale layers rich in organic matter and pyrite with a few intercalated tempestitic limestone beds and large, early diagenetic, calcareous concretions (for references and further details concerning the lithostratigraphy of the Dienerian of Spiti see Ware et al. in production a). Articulated fish fossils usually occur in calcareous concretions, whereas the intercalated black shale layers frequently yield isolated remains, such as bones or incomplete fins. Black shale is typically deposited in an anoxic, reducing environment on the outer platform. During the Early Triassic, the study area was situated on the northern margin of Gondwana (Neotethys realm) at a palaeolatitude of ca. 30°S (Fig. 1a).

Stratigraphic sections of the lower part of the Limestone and Shale Member (Mikin Formation) of the studied sections near the villages of Mud, Tilling, and Guling (Pin Valley, Spiti Subdistrict, Himachal Pradesh, India; cf. Fig. 1), with occurrences of the osteichthyan material described herein. 1 Unlabelled specimen (Actinopterygii indet.), 2 PIMUZ A/I 4361 (Actinopterygii indet.), 3 PIMUZ A/I 4145 (Saurichthys sp.), 4 PIMUZ A/I 4360 (cf. Watsonulus cf. eugnathoides), 5 PIMUZ A/I 4362 (Actinistia indet.). For more details on the stratigraphy see text and Ware et al. (2015, in production a, b)
Within the Spiti Subdistrict, outcrops of the Lower Triassic and Anisian (lower Middle Triassic) Mikin Formation (cf. Bhargava et al. 2004) are widespread (Fig. 1). The prominent, cliff-forming Niti Limestone Member (Spathian, upper Olenekian, upper Lower Triassic) of the Mikin Formation is easily recognizable in the field. The Mikin Formation partly extends into remote areas and some sections may be difficult to access. Moreover, the fish-bearing beds of the Limestone and Shale Member are often covered by firn, rubble and/or vegetation. Some of the most accessible outcrops are found within the valleys of the Pin River and its tributary streams at roughly 3800 m above sea level (Fig. 1). There, sections near the villages of Mud (sometimes spelled Muth or Mudh) and Gulling have been intensively exploited for ammonoids and conodonts (e.g. Diener 1897, 1908; Krafft and Diener 1909; Krystyn and Orchard 1996; Bhargava et al. 2004; Krystyn et al. 2004, 2007a, b; Brühwiler et al. 2010, 2012; Goudemand 2010, 2014; Ware et al. in production a).
Early Triassic ammonoids from Spiti are known since the late 19th century (Diener 1897). The taxonomy and biostratigraphy of Dienerian and Smithian ammonoids from the Spiti Subdistrict were recently revised by Ware et al. (2015, in production a) and Brühwiler et al. (2010, 2012), respectively. Brühwiler et al. (2010) demonstrated the occurrence of typically Smithian ammonoid faunas lower than previously described in a section near Mud, which has recently been proposed as a GSSP for the base of the Olenekian stage by Krystyn et al. (2007a, b). We herein follow the definition of the Dienerian-Smithian substage boundary of Brühwiler et al. (2010), which is also supported by conodont data (Goudemand 2010, 2014). Within the Dienerian of Spiti, Ware et al. (in production a) recognised ten ammonoid zones, confirming the results obtained for faunas from the Salt Range in Pakistan (Ware et al. in production b) and the subdivision of the Dienerian into three parts (early, middle, and late; Ware et al. 2015).
Actinopterygian bone terminology used herein is adopted from Lehman (1952). One fossil recovered from the Early Triassic of Spiti is identified as an actinistian urohyal. Comparative studies have shown that the sarcopterygian ‘urohyal’ is not homologous with the teleostean urohyal and consequently some authors (e.g. Arratia and Schultze 1990) used inverted commas when referring to this element in lobe-finned fishes. We here follow most recent authors (e.g. Cavin et al. 2013; Dutel et al. 2015a, b) and refer to the actinistian urohyal without inverted commas, but no homology with the teleostean urohyal is implied. The use of open nomenclature in the present paper adheres to the recommendations of Bengtson (1988).
Systematic palaeontology and description
Class Osteichthyes Huxley, 1880
Subclass Actinopterygii Cope, 1887 (sensu Rosen et al. 1981)
Infraclass Actinopteri Cope, 1872, (sensu Patterson 1982)
Family Saurichthyidae Owen, 1860 (sensu Stensiö 1925)
Genus Saurichthys Agassiz, 1834 (sensu Mutter et al. 2008)
Type species. Saurichthys apicalis Agassiz, 1834 from the Middle Triassic of Bayreuth, Germany.
Saurichthys sp. (Fig. 3)

Saurichthys sp. (PIMUZ A/I 4145): left lower jaw with teeth from the late Dienerian (lower part of the Limestone and Shale Member, Mikin Formation) of Mud Bottom Section (Pin Valley, Spiti Subdistrict, Himachal Pradesh, India). a Part a of A/I 4145 (scale bar measures 50 mm); b part b of A/I 4145 showing details of the distinct lateral ornamentation pattern (scale bar measures 20 mm)
Material and stratigraphy. A single specimen, PIMUZ A/I 4145 (Fig. 3), which is an almost complete left lower jaw preserved on two slabs of black shale (parts a and b). Part a of A/I 4145 (Fig. 3a) is a body fossil and part b (Fig. 3b) a cast of the middle portion of part a, with impressions of the external ornamentation. A/I 4145 was found above the village Mud (“Bottom Section” of Ware et al. in production a) in Pin Valley (Lahaul and Spiti District, Himachal Pradesh, India; Fig. 1). The specimen originates from late Dienerian strata between the Vavilovites meridialis Zone and the Kingites davidsonianus Zone of Ware et al. (2015, in production a, b) within the lower part of the Limestone and Shale Member, Mikin Formation (Fig. 2).
Description. The hemimandible is anteriorly complete and bears several large teeth, which are best-seen on part a of PIMUZ A/I 4145 (Fig. 3a). Posteriorly, the left lower jaw is incomplete and it is unclear whether the angular and surangular bones are present. The length of the preserved part of A/I 4145a adds up to ca. 130 mm. The maximum height in the hind region of A/I 4145a measures ca. 15 mm. The straight superior margin and the gently S-curved inferior rim converge towards rostrally, giving the lower jaw a gracile, pointed front end.
On the lateral surface, the upper part of the dentary bone is covered with parallel running, subvertical striae (A/I 4145b, Fig. 3b). In closer view, these fine striae are made up of oblong tubercles, some of which coalesce. The lower part of the external face of the mandible is ornamented with striae that run parallel to the ventral margin of the bone. These striae are much coarser than those in the upper part (Fig. 3b).
Several conical teeth of two size classes are developed in the anterior and middle portion of the dentary. There are about nine widely spaced larger teeth and at least thirteen smaller ones in between. The teeth possess a vertically striated basal portion and a smooth apical cap that, as far as can be seen on the larger teeth, measures a bit more than one third of the total tooth height.
Remarks. The overall morphology and the peculiar lateral ornamentation pattern of PIMUZ A/I 4145 are well in agreement with those of the Triassic predatory actinopterygian Saurichthys (e.g. Stensiö 1925; Lehman 1952; Griffith 1959; Lehman et al. 1959), but the incomplete preservation of the Spiti specimen precludes a determination to species level (see "Discussion"). The dimensions of A/I 4145 suggest an estimated total length range of ca. 400–600 mm for the animal (cf. Griffith 1959; Romano et al. 2012). A/I 4145 was already mentioned in Romano et al. (2012).
Series Neopterygii Regan, 1923
Superdivision Holostei Müller, 1845 (sensu Grande 2010)
Division Halecomorphi Cope, 1972 (sensu Grande and Bemis 1998)
Family Parasemionotidae Stensiö, 1932
Genus Watsonulus Brough, 1939
Remarks. Watsonulus is the replacement name for Watsonia Piveteau, 1934, which is preoccupied (Brough 1939: p. 91).
Type and only species. Watsonia eugnathoides Piveteau, 1934 from the Early Triassic of northwest Madagascar.
cf. Watsonulus cf. eugnathoides (Figs. 4, 5, 6)

cf. Watsonulus cf. eugnathoides (PIMUZ A/I 4360) from the late Dienerian (lower part of the Limestone and Shale Member, Mikin Formation) of Tilling Section (Pin Valley, Spiti Subdistrict, Himachal Pradesh, India). a Photo of the eastern slope of Pin Valley between the villages of Khar and Tilling (cf. Fig. 1), with indication of the approximate location where A/I 4360 was found (white arrow), b part a of A/I 4360, c part b of A/I 4360. Scale bar in b and c measures 50 mm

cf. Watsonulus cf. eugnathoides (PIMUZ A/I 4360) from the late Dienerian (lower part of the Limestone and Shale Member, Mikin Formation) of Tilling Section (Pin Valley, Spiti Subdistrict, Himachal Pradesh, India). a Close-up view of the skull and anterior body portion of part a of A/I 4360, b drawing of the bones of the cranium and pectoral girdle of A/I 4360a, with indication of the bones and sensory canals (dashed grey lines). An angular, Ao antorbital bone, Br branchiostegal ray, Cl cleithrum, Cv clavicula, De dentary, Dp dermopterotic, Ds dermosphenotic, Ex extrascapular, F frontal, Io infraorbital bone, Iop interoperculum, lat.ca. lateral line sensory canal, Mx maxilla, Na nasal, Op operculum, P parietal, Pcf pectoral fin, Pcl postcleithrum, Pm praemaxilla, Pop praeoperculum, pop.ca. praeopercular sensory canal, Pt posttemporal, Scl supracleithrum, Sm supramaxilla, So supraorbital, so.ca. supraorbital sensory canal, Sop suboperculum, st.ca. supratemporal sensory canal

cf. Watsonulus cf. eugnathoides (PIMUZ A/I 4360) from the late Dienerian (lower part of the Limestone and Shale Member, Mikin Formation) of Tilling Section (Pin Valley, Spiti Subdistrict, Himachal Pradesh, India). a Close-up view of the dorsal fin of A/I 4360a, b magnified detail of the pelvic and anal fin of A/I 4360b. Scale bar for a and b measures 50 mm. Af anal fin, Pvf pelvic fin
Material and stratigraphy. A single specimen, PIMUZ A/I 4360 (Figs. 4, 5, 6), which is a complete fish preserved within a calcareous nodule as part (A/I 4360a; Figs. 4b, 5, 6a) and counterpart (A/I 4360b; Figs. 4c, 6b). The specimen is exposed in left lateral view in part a, whereas part b mostly contains the impressions of the bones preserved in part a. A latex peel cast of A/I 4360b was prepared for this study. A/I 4360 was found about halfway between the villages of Khar and Tilling (Pin Valley, Lahaul and Spiti District, Himachal Pradesh, India), high above the valley floor at ca. 4030 m altitude (“Tilling Section” of Ware et al. in production a, N32°00′46.2′′/E78°05′04.5′′; Figs. 1, 4a). It was recovered from the late Dienerian Kingites davidsonianus Zone of Ware et al. (2015, in production a, b), lower part of the Limestone and Shale Member, Mikin Formation (Fig. 2).
Description. PIMUZ A/I 4360 is flattened, but the three-dimensional shape of the skull and the body are still discernible. For the most part, the bones are weathered away and usually only their medial impression is preserved. For this reason, no comments can be made about the external ornamentation. A/I 4360 has a standard length of ca. 210 mm, whereas the head (including gill cover) measures ca. 60 mm.
The dermal skull roof of A/I 4360 (Fig. 5) is preserved as one unit in dorsal view (Fig. 5). Its postorbital portion is better preserved than the anterior one. The posterior part of the dermal skull roof is shaped by two paired elements: the parietals medially and the large dermopterotics laterally. Extrascapulars are not preserved in situ, but a small fragment situated posterior to the dermal skull roof may belong to the left extrascapular bone (Fig. 5). The parietals are antero-posteriorly relatively short and of subquadrangular shape. These bones are bounded by fairly straight caudal and lateral margins. Their medial borders are distinctly undulating, whereas their anterior confinements are gently oblique, running from antero-medial to postero-lateral. The dermopterotics are nearly twice as long as the parietals. Each dermopterotic is bounded by four margins: a very short and gently oblique posterior one, a convex medial border, a short, concave antero-lateral margin, and a relatively straight postero-lateral border.
The central portion of the dermal skull roof is occupied by the large, paired frontal bones and the very small, paired dermosphenotics (Fig. 5). The rhomboidal dermosphenotics are firmly incorporated into the dermal skull roof and postero-laterally adjoin the dermopterotics. The frontal is the largest bone of the dermal skull roof. It is distinctly longer than wide and possesses a notch in the lateral margin, which houses the neighbouring dermosphenotic. The medial margin of the frontal is sinuous. Rostrally, the frontals border the paired nasal bones. The confinement between these elements is seemingly mildly curved. As far as can be seen, the anterior ends of the left and right frontal are medially separated and the nasal bones are medially not in contact with each other. Antero-laterally to the right frontal and postero-laterally to the right nasal lie about two serially arranged supraorbital bones (Fig. 5).
The dermal skull roof is distinctly three-dimensionally bent (Fig. 5). Within its posterior portion, up to the level of the orbit, the dorsal surface of the dermal skull roof is concave along its median axis but convex along its lateral flanks. At the level of the anterior supraorbital, the dermal skull roof is relatively flat. Further rostrally, however, the external surface of the dermal skull roof becomes increasingly convex anteriorly and laterally. The curvature of the dermal skull roof of A/I 4360 is bilaterally symmetrical, suggesting that it reflects the original shape, but some taphonomic deformation cannot be excluded.
The cheek region is mainly occupied by the large, slightly anteriorly inclined praeoperculum. This bone is higher than long and bounded by four margins: a long, convex posterior one, straight antero-dorsal and antero-ventral borders, and a short, concave anterior confinement. The lateral surface of the praeoperculum is gently convex. The circumorbital series is incompletely preserved and only represented by three elements. Next to the left dermosphenotic lies a poorly preserved bone that likely corresponds to an infraorbital. A further, teardrop-shaped infraorbital bone is found in situ between the praeoperculum and the supramaxilla. In addition, two fragmentary elements that are antero-laterally bordering the nasal bones may belong to the antorbitals. A sclerotic ring is not preserved.
The bones of the upper and lower jaw are located on either side of the dermal skull roof (Fig. 4). On the left side of the cranium (Fig. 5), the jaw bones are preserved in situ. The upper jaw consists of the long and slender maxilla, the supramaxilla, and possibly a pair of praemaxillae. The latter may be preserved anterior to the nasals (Fig. 5), but the poor state of preservation of these bones precludes a description. The left maxilla is preserved in two separate segments in A/I 4360a (Fig. 5). The rostral portion and most of the ventral margin of the maxillary bone are missing and teeth have not been observed. The left mandibular branch is largely complete and only its rostral end is lacking. The boundaries between the dentary, the angular, and surangular bones are barely visible. The dentition of the lower jaw is not preserved.
Of the bones of the operculogular series only the operculum, suboperculum, and a few branchiostegal rays are visible (Fig. 5). Gulars are not preserved. The operculum and suboperculum are both large, plate-like bones that shape most of the lateral gill cover. The operculum is a roughly five-sided bone bounded by a gently concave antero-ventral border, a straight antero-dorsal margin, a long, deeply convex postero-dorsal border, a straight postero-ventral margin, and a short, convex ventral confinement. The postero-dorsal border is damaged and mostly preserved as an impression. The suboperculum adjoins the operculum ventrally. This bone is longer than high and confined by five margins: a slightly concave anterior, a deeply concave dorsal, a distinctly convex posterior, a nearly straight postero-ventral, and a short, concave antero-ventral. The suboperculum borders the interoperculum antero-ventrally and a branchiostegal ray ventrally, but both of these elements are only preserved fragmentarily. Two additional radii branchiostegii are located further ventrally (Fig. 5).
Most elements of the shoulder girdle are present, but the majority of them are poorly preserved. A fragmentary bone attached to the putative left extrascapular may correspond to the left posttemporal (Fig. 5). The supracleithrum is found in situ and is best visible in part b of A/I 4360 (Fig. 4c). It is an elongate, four-sided bone that in vivo was partially overlapped by the adjacent operculum. The cleithrum is probably preserved in place but heavily damaged. The endoskeletal shoulder girdle cannot be traced. A fragmentary, plate-like bone neighbouring the branchiostegal rays could correspond to the clavicula. There are two postcleithra, an elongate upper one that is anteriorly in contact with the supracleithrum and cleithrum, and a smaller, teardrop-shaped lower one (Fig. 5).
All fins are present but in different preservational conditions. The pectoral fin is largely disarticulated, but several isolated lepidotrichial segments are found ventrally to the skull and shoulder girdle (Figs. 4b, c, 5). The length of some fin rays (up to ca. 50 mm) and the distribution pattern of the lepidotrichial segments suggests that the pectoral fin of A/I 4360 was of rather large size. The pelvic fins, even though poorly preserved, lie closer to the anal fin than to the pectoral girdle (Figs. 4b, c, 6b). At least four fin rays are observed in situ. The first lepidotrichium of the pelvic fin is thicker than the subsequent ones and distally segmented into very short units (Fig. 6b). Furthermore, its leading edge is equipped with small fringing fulcra (A/I 4360a). Several isolated fin ray segments in close proximity to the pelvics may also belong to these fins. The pelvic fins were seemingly short-based and generally small. The virtually opposed dorsal and anal fins are placed closer to the caudal fin than to the cranium (Fig. 4). The dorsal fin begins a very short distance ahead of (but a few scale rows behind) the anal fin and both fins contain about twelve lepidotrichia each. Each fin ray of the dorsal and anal fin consists of a relatively long basal segment and numerous, very short distal ones (Fig. 6). At least those lepidotrichia terminating at the posterior fin margin are once or twice distally bifurcated. The anterior margin of the first lepidotrichium of the dorsal and anal fin is edged with fringing fulcra. A few basal fulcra are probably developed in front of the dorsal fin (Fig. 6a). The caudal fin is relatively poorly preserved and the distal parts of its upper and lower lobes are missing (Fig. 4c). The number of lepidotrichia in the caudal fin cannot be determined. As far as can be seen, the tail fin is only weakly heterocercal and at least the leading edge of the dorsal caudal lobe is armed with fulcra.
The entire body of A/I 4360 (Fig. 4) is covered with rhombic scales. Hence, the endoskeletal support of the fins and the elements of the axial skeleton cannot be studied. The squamation consists of oblique scale rows. At least 35 scale rows are counted along the lateral line. Scales on the side of the body are usually higher than long, whereas those situated more dorsally and ventrally are usually longer than high. The largest scales are found behind the upper postcleithrum (Fig. 5a). The ornamentation of the scales is not preserved.
The lateral line sensory canal can be traced across multiple scales along the flank of the body (best seen in the cast of A/I 4360b). Anteriorly, the lateral line canal pierces the supracleithrum through its posterior margin and traverses this bone just above the area covered by the neighbouring operculum (Fig. 5). Sensory canals can also be spotted on the dermal skull roof. The supratemporal section of the lateral line runs through the dermopterotic before it enters the dermosphenotic. Due to limited preservation the canal cannot be traced within the infraorbital series and the antorbital bones. The left and right branches of the supraorbital sensory canal are well visible in the parietal, frontal, and nasal bones. The praeopercular sensory canal is seen near the posterior margin of the praeoperculum. It closely follows the caudal margin of the praeoperculum in its upper part, whereas the distance to the hind border increases towards the ventral portion of the bone. The mandibular canal, which usually runs along the posterior and ventral margins of the lower jaw, could not be detected in the studied specimen.
Remarks. PIMUZ A/I 4360 shows many similarities with Watsonulus eugnathoides (Piveteau 1934) and can, thus, be referred to Parasemionotidae Stensiö (1932). However, taxonomic problems within Parasemionotidae presently impede a definite determination of the Spiti specimen (see “Discussion”).
A/I 4360 is associated with several internal moulds of indeterminable bivalves (Fig. 4b–c). A dense accumulation of clams is found just ventrally to the skull and partially also on the lower jaw of the fish (cf. A/I 4360b). The bivalves seem disarticulated but otherwise undamaged, which suggests that this accumulation is neither a regurgitalite of the fish nor a coprolite of a different animal.
Actinopterygii indet. (Fig. 7)

Actinopterygii indet. (PIMUZ A/I 4361) from the late Dienerian (lower part of the Limestone and Shale Member, Mikin Formation) of Mud Bottom Section (Pin Valley, Spiti Subdistrict, Himachal Pradesh, India). a Part a of A/I 4361 representing a nearly complete fish lacking the skull and most fins (scale bar measures 50 mm), b close-up view of the squamation of the caudal peduncle with preserved ornamentation (scale bar measures 20 mm). Cf caudal fin, Pcf pectoral fin
Material and stratigraphy. Two articulated specimens preserved in limestone nodules from the lower part of the Limestone and Shale Member of the Mikin Formation. Both individuals were recovered in 2009 at a site above the village of Mud (“Bottom Section” of Ware et al. in production a), Pin Valley, Lahaul and Spiti District, Himachal Pradesh, India (Fig. 1). The first specimen, PIMUZ A/I 4361 (Fig. 7), is preserved on two complementary plates (a and b) and originates from the late Dienerian Vavilovites meridialis Zone (Ware et al. in production a, b) (Fig. 2). The second specimen, which is from the middle Dienerian Ambites lilangensis Zone (Ware et al. 2015, in production a, b; Fig. 2), requires preparation and is thus not taken further into account herein. A/I 4361 is described below but is treated in open nomenclature due to the absence of diagnostic features.
Description. PIMUZ A/I 4361 (Fig. 7) is a relatively large, slender actinopterygian with only its body preserved, including the pectoral fin and a few lepidotrichia of the caudal fin. The skull and shoulder girdle are entirely missing and the remaining fins cannot be traced. The length of the preserved body portion of A/I 4361 (without caudal fin rays) measures ca. 275 mm. The postero-ventrally directed pectoral fin (Fig. 7a) is peculiarly long (at least 55 mm) and slender. The number of fin rays and the segmentation pattern are difficult to determine, but the count of lepidotrichia seems to be low. Fringing fulcra are seemingly absent on the pectoral fin. The endoskeletal part of the pectoral fin is not preserved.
The trunk is covered with relatively small, rhombic scales. Most scales are poorly preserved but the oblique scale rows can still be traced. About 67 scale rows are counted in total. Some scales still have their lateral ornamentation preserved, especially those on the flanks of the caudal peduncle. The ornamentation consists of thick horizontal striae (Fig. 7b).
Remarks. PIMUZ A/I 4361 and the second, unlabelled specimen are both assigned to Actinopterygii based on the presence of oblique rows of rhombic scales.
Subclass Sarcopterygii Romer, 1955 (sensu Rosen et al. 1981)
Infraclass Actinistia Cope, 1871 (sensu Forey 1998)
Actinistia indet. (Fig. 8)

Actinistia indet. (PIMUZ A/I 4362): Urohyal of a ‘coelacanth’ fish from the late Dienerian (possibly earliest Smithian) of Guling River section (lower part of the Limestone and Shale Member, Mikin Formation, Spiti Subdistrict, Himachal Pradesh, India), seen in dorsal view. a Part a of A/I 4362, b counterpart (A/I 4362b), c drawing of A/I 4362b. Arrow points anteriorly. Scale bar measures 20 mm
Material and stratigraphy. A single, largely complete actinistian urohyal, PIMUZ A/I 4362, preserved in dorsal aspect on two rock hand samples of black shale, part a (A/I 4362a; Fig. 8a) and its counterpart b (A/I 4362b; Fig. 8b), with the fracture plane going through the element. The most posterior part of the bone is incomplete. A/I 4362 was collected near Guling on the opposite side of the Pin River Valley (“Guling River” section in Ware et al. in production a), Lahaul and Spiti District, Himachal Pradesh, India (Fig. 1). It is derived from the lower part of the Limestone and Shale Member of the Mikin Formation (Fig. 2) and was recovered in situ from black shale layers between the late Dienerian Kingites davidsonianus Zone and the earliest Smithian Flemingites bhargavai Zone of Brühwiler et al. (2010) and Ware et al. (2015, in production a, b).
Description. PIMUZ A/I 4362 (Fig. 8) is a thin, elongate element with a pronounced slender shape. It is dorso-ventrally flat and horizontally broad. The form of the urohyal in the horizontal plane roughly resembles that of an hourglass possessing unequally sized lobes: a short anterior one (ca. 7 mm long) and a longer posterior one (at least 16 mm). A distinct constriction analogous to the neck of an hourglass is developed within the rostral portion of this bone.
The short anterior segment of the urohyal is rostrally broad, reaching a maximum width of ca. 3 mm. The front end is blunt and the terminal margin is marked by a small central notch (Fig. 8). The long caudal segment of the urohyal is posteriorly forked and thereby tapers off as two wings, each of which probably had an acute hind end. The caudal wings of the urohyal reach their greatest horizontal width (ca. 7 mm) about halfway between the constricted portion of the bone and its hind end. Caudally, the two posterior rami are separated by a distinct interspace. Anteriorly, the wings are connected via a wedge-shaped median portion. The hind part of this medially placed segment is incompletely preserved but it seems to have extended halfway between the level of greatest width of the urohyal and the posterior end of the bone (Fig. 8c). The preserved total length of the urohyal is about 23 mm.
For the most part, the urohyal is plate-like, especially in its elongate posterior portion. Two fine ridge-like structures are detected along the lateral confinements of the anterior segment (A/I 4362; Fig. 8). Just behind the constricted portion, the bone is distinctly dorsally arched. This curvature of the dorsal surface continues along the lateral margins within the rostral part of the posterior wings of the urohyal. The flat, triangular median portion between the caudal wings is separated from the latter by two distinct ridges. These ridges continue posteriorly along the medial margins of the caudal wings of the urohyal. In dorsal view, the triangular median portion appears slightly lowered relative to the posterior wings (A/I 4362b; Fig. 8b).
Remarks. The distinct shape and morphology of PIMUZ A/I 4362 allows a confident identification of this element as an actinistian urohyal. However, due to limited knowledge of the urohyal of fossil ‘coelacanth’ fishes, including most Early Triassic forms, we currently refrain from a determination at low taxonomic rank (see “Discussion” section).
Discussion
Record of Triassic fishes from the Indian subcontinent
Despite the long history of palaeontological and geological research in Spiti (e.g. Diener 1897, 1908, 1912; Hayden 1904; Krafft and Diener 1909; Krystyn and Orchard 1996; Krystyn et al. 2004; Bhargava et al. 2004; Brühwiler et al. 2010), fish macrofossils have hitherto not been reported from this region (see also below). Merely Goel (1977) mentioned the occurrence of microscopic fish remains (teeth and scales) in a section of the Mikin Formation near the village of Khar (cf. Fig. 1), but this material was neither described nor figured. The fact that fish macrofossils have not yet been reported is surprising given our own experience during fieldwork in Spiti. Within only a couple of days we were able to find several isolated remains and even some articulated specimens. Considering the vast exposures of the Mikin Formation (cf. Fig. 1) compared with the small number of localities so far explored for fish remains, it is likely that further explorations in Spiti will yield more material.
The Indian subcontinent bears numerous sites producing Early Triassic fish fossils, as a matter of fact more than most other former Gondwanan provinces (cf. López-Arbarello 2004; Brinkmann et al. 2010). Outcrops within the peninsular part of India are all freshwater settings and yield only ichthyoliths (Panchet Formation, e.g. Chatterjee and Roy-Chowdhury 1974; Bandyopadhyay 1999; Bandyopadhyay et al. 2002; Gupta 2009), whereas those from the extrapeninsular part (Northern Indian Margin) all represent marine deposits (e.g. Chang and Miao 2004; Brinkmann et al. 2010). Younger Triassic fish assemblages from the Indian subcontinent are mainly freshwater (e.g. the Middle Triassic Yerrapalli Formation or the Upper Triassic Tiki and Maleri formations), rarely marine (e.g. Misra et al. 1973; Chhabra and Mishra 2002), and usually typified by isolated skeletal remains (e.g. Agarwal and Singh 1981; Bandyopadhyay 1999; Bandyopadhyay et al. 2002; Chang and Miao 2004; Prasad et al. 2008).
Early Triassic marine chondrichthyan and osteichthyan fishes from the Northern Indian Margin (Neotethys realm, Fig. 1a) have been described from the Salt Range in northwest Pakistan (e.g. De Koninck 1863a, b; Waagen 1895; Romano et al. 2014b) as well as from the Himalayas of Jammu and Kashmir (e.g. Sahni and Chhabra 1976; Agarwal and Singh 1981; Srivastava and Mehrotra 1986), India (e.g. Sahni and Chhabra 1976; Mehrotra et al. 1983a, b; Mishra et al. 1990), Nepal (Beltan and Janvier 1978), and South Tibet (Zhang 1976; Brinkmann et al. 2010). However, most of this material consists of isolated remains, such as teeth or scales, which are valuable for chondrichthyan parataxonomy but only of limited use for osteichthyan taxonomy. Merely two finds from the Northern Indian Margin are more complete, i.e. a tooth whorl fragment of the edestid chondrichthyan Sinohelicoprion qomolangma Zhang, 1976 from South Tibet and a partial skull of Saurichthys nepalensis Beltan and Janvier, 1978 from the Nepalese Annapurna (Chang and Jin 1996; Chang and Miao 2004; Jin 2006; Brinkmann et al. 2010). Precise biostratigraphic data for the aforementioned fish fossils is, in many cases, wanting (e.g. Beltan and Janvier 1978). The new ichthyofauna from Spiti includes the most complete fish macrofossils ever described from the Triassic of India and benefits from a detailed biostratigraphic framework (e.g. Ware et al. 2015, in production a).
Taxonomic discussion of the Spiti ichthyofauna
The presented Early Triassic fish assemblage from Spiti comprises material of Saurichthys Agassiz, 1834, a parasemionotid (cf. Watsonulus cf. eugnathoides), as well as further actinopterygian and actinistian remains. The Spiti ichthyofauna thus includes taxa also known from several other coeval assemblages (see below), highlighting the similarities of fish biocoenoses during the Early Triassic (see e.g. Romano et al. 2014a for a recent summary and references). Cosmopolitanism at that time may have been the combined result of the devastating effects of the end-Permian mass extinction event, of poorly differentiated habitats, and of the simple Pangaean supercontinent configuration, facilitating faunal exchange of the surviving and newly evolved taxa.
Saurichthys is a common component of Triassic fish assemblages and usually very easily identifiable due to its peculiar anatomy (e.g. Romano et al. 2012). During the Early Triassic, Saurichthys had a worldwide distribution and this taxon has previously also been described from the Northern Indian Margin (Beltan and Janvier 1978), even though with poor age control. The presence of Saurichthys in the Dienerian of Spiti underlines the successful circumpangaean dispersal of this highly specialised predator within only a couple of hundred thousand years (Galfetti et al. 2007; Burgess et al. 2014; Ovtcharova et al. 2015) after the end-Permian mass extinction event (Mutter et al. 2008; Romano et al. 2012; Scheyer et al. 2014).
Watsonulus Brough, 1939 was first described from the Early Triassic of Ambilobe Basin in northwest Madagascar (Piveteau 1934, 1939–1940; Lehman 1952; Beltan 1968; Olsen 1984; Grande and Bemis 1998), but was later also discovered in strata of similar age in southwest Madagascar (Morondava Basin) studied by Lehman et al. (1959). The material from both northwest and southwest Madagascar is referred to a single species, Watsonulus eugnathoides (Piveteau, 1934). However, based on minor morphological differences several types of W. eugnathoides have been distinguished (A, B, and C of Piveteau 1934; 1, 2, and 3 of Lehman 1952). Furthermore, from the earliest Triassic (Griesbachian substage) of East Greenland, Nielsen (1936) also mentioned specimens referable to Watsonulus, but a description of this material was never published.
Watsonulus belongs to Parasemionotidae Stensiö, 1932 (=Ospiidae Stensiö, 1932; see e.g. Nielsen 1936), a very speciose and cosmopolitan family known only from the marine Early Triassic (Grande and Bemis 1998). Morphological and cladistic analyses suggest that these fishes are basal halecomorph neopterygians (e.g. Patterson 1973; Grande and Bemis 1998; Xu et al. 2014; Friedman 2015). Besides India, parasemionotids are known from Greenland, Madagascar, China, and Canada, and at least 17 species have been described, most of which belong to monotypic genera (Table 1). In addition, Patterson (1973) included several other Triassic taxa within Parasemionotidae, amongst others the Early Triassic genera Tungusichthys Berg, 1941 (Siberia, Russia) and Paracentrophorus Piveteau, 1939–1940 (northwest Madagascar), but his proposition was later rejected by Grande and Bemis (1998). Grande and Bemis (1998) list the Early Triassic Helmolepis Stensiö, 1932 as a parasemionotid, which is erroneous (see e.g. Nielsen 1936; Neuman and Mutter 2005). A parasemionotid-like fish, Peia Li, 2009, was described from the Early Triassic of Jiangsu Province, China. Interestingly, parasemionotids are absent at the Spitsbergen Lagerstätte (Svalbard, Arctic Norway), in spite of the otherwise similar ichthyofaunal composition to other Early Triassic sites (e.g. Nielsen 1936; Brinkmann et al. 2010).
Although a few authors have expressed confidence regarding the validity of several parasemionotid genera (e.g. Nielsen 1936), many taxa are distinguished by relatively small morphological differences, some of which could reflect ontogenetic or individual variation (Patterson 1973, 1975; Grande and Bemis 1998). Parasemionotidae is in need of thorough taxonomic revision, but such a task is beyond the scope of this study. The high number of parasemionotid species as well as current uncertainties regarding the validity of several of them renders the taxonomic interpretation of PIMUZ A/I 4360 (Figs. 4, 5, 6) difficult. A/I 4360 is herein referred to cf. Watsonulus cf. eugnathoides due to similarities with the type specimens of W. eugnathoides (MNHN.F.MAE 11, MNHN.F.MAE 33, MNHN.F.MAE 34, MNHN.F.MAE 42, MNHN.F.MAE 108) figured by Piveteau (1934). In the following, A/I 4360 is compared with other parasemionotids, where the taxa are listed respectively in the sequence of publication of their names (cf. Table 1).
Based on the literature and personal observations (CR), the most notable differences between parasemionotid species concern (1) the anatomy of the brain case, (2) the proportions of the skull and body, (3) the shape and fragmentation pattern of the praeoperculum, (4) the number of branchiostegal rays, (5) the size of the pectoral fins, (6) the relative position of the pelvic fins, and (7) the number of lepidotrichia as well as presence or absence of fringing fulcra. Regarding point (1) no comparisons can be made because the endocranium is not preserved in A/I 4360. This also precludes comparisons with Devillersia Beltan, 1968 and Piveteaunotus Beltan, 1968, both of which are primarily known from brain cases. The proportions between the head and body (point 2) of A/I 4360 mostly resemble those of Watsonulus, Lehmanotus Beltan, 1968, Jurongia Liu in Liu et al., 2002, and Suius Liu in Liu et al., 2002. In these taxa and A/I 4360, the length of the head (tip of snout to hind margin of gill cover) usually measures about one-fourth or more of the standard length (Fig. 4; Piveteau 1934; Beltan 1968; Liu et al. 2002; Tong et al. 2006). By contrast, A/I 4360 differs from taxa such as Parasemionotus or Qingshania Liu in Liu et al., 2002, which have a smaller head to body length ratio (e.g. Priem 1924; Piveteau 1934). In addition, specimen A/I 4360, as well as Ospia, Watsonulus (especially type C of Piveteau 1934) and Jurongia, are characterized by a long lateral gill cover (operculum plus suboperculum) relative to the length of the anterior part of the skull (Fig. 5; Stensiö 1932; Piveteau 1934; Liu et al. 2002). However, the lateral gill cover is distinctly shorter with respect to the rest of the skull in Stensionotus Lehman, 1952, Jacobulus Lehman, 1952, Thomasinotus Lehman, 1952, Albertonia, Lehmanotus, and Suius (e.g. Beltan 1968; Schaeffer and Mangus 1976; Liu et al. 2002).
The praeoperculum (point 3) is a single, relatively large and subvertically arranged bone in A/I 4360, much like in Parasemionotus, Ospia, Broughia, Watsonulus, and Jurongia (Fig. 5; Priem 1924; Stensiö 1932; Piveteau 1934; Liu et al. 2002). By contrast, the antero-dorsal part of the praeoperculum is subdivided into smaller plates in Stensionotus, Jacobulus, Thomasinotus, and Lehmanotus (Lehman 1952; Beltan 1968). However, Lehman (1952) referred specimens to Parasemionotus and Watsonulus that showed a few very small “praeopercular plates” (anamestic bones) dorsally to the main praeoperculum. There also seems to be variation of the antero-posterior extension of the praeoperculum between taxa (e.g. Stensiö 1932; Piveteau 1934; Lehman 1952). The number of branchiostegal rays (point 4) seems to vary between parasemionotid species (e.g. Piveteau 1934; Lehman 1952; Lehman et al. 1959; Olsen 1984; Li 2009), although this character is often not well known. Also, differences in counts of radii branchiostegii could be due to ontogenetic variation as the number of these elements usually increases during growth (Arratia and Schultze 1990).
The size of the pectoral fin (point 5) appears to be a distinctive feature that separates Watsonulus, Albertonia Gardiner, 1966, and Icarealcyon Beltan, 1980 from other parasemionotids. Whereas the pectoral fins of most parasemionotids are usually small or only slightly enlarged (e.g. Parasemionotus, Ospia, Jacobulus, Lehmanotus, Jurongia; Stensiö 1932; Piveteau 1934; Lehman 1952; Beltan 1968; Liu et al. 2002), these fins possess a considerable size in Watsonulus (MNHN.F.MAE 11, Piveteau 1934: pl. 9 Figs. 1, 1a), Albertonia (CMNFV 757, Lambe 1916: pl. 2; CMNFV 12330; Schaeffer and Mangus 1976: Fig. 18a), and Icarealcyon (MNHN.F.MAE 576, Beltan 1984: pl. 3a, b). Although incompletely preserved, the pectoral fin of A/I 4360 appears to have been of fairly large size (Figs. 4, 5), hence being similar to that of Watsonulus, Albertonia, or Icarealcyon. Nevertheless, whether the size of the pectoral fin has taxonomic value or whether alternative explanations such as preservational bias, ontogenetic variation or sexual dimorphism can account for it requires detailed comparative analysis.
The pelvic fins (point 6) are small and short-based in all parasemionotids and they are normally placed approximately midway between the pectoral girdle and the anal fin. In some taxa, such as Watsonulus or Albertonia, the pelvic fins seem to be slightly nearer to the anal fin than to the shoulder girdle (e.g. MNHN.F.MAE 11, Piveteau 1934: pl. 9 Figs. 1, 1a; CMNFV 757, Lambe 1916: pl. 2; CMNFV 12330, Schaeffer and Mangus 1976: Fig. 18a), whereas in others like Parasemionotus (e.g. MNHN.F.MAE 122; Priem 1924; Piveteau 1934) the pelvic fins appear to be slightly closer to the pectoral girdle than to the anal fin. In this respect, A/I 4360 shows more resemblance to Watsonulus or Albertonia than with most other taxa.
Some variation has been noted in the distribution of fringing fulcra and the count of lepidotrichia (point 7). For instance, whereas fringing fulcra have been detected on the leading margin of the fins of most parasemionotids (e.g. Parasemionotus, Watsonulus, Albertonia; Lambe 1916; Piveteau 1934; Lehman 1952), such elements are reportedly absent on the leading edge of the pectoral and pelvic fins of Icarealcyon (Beltan 1984). Differences in the number of lepidotrichia are small in some cases but more distinct in others. For example, Beltan (1984) counted in Icarealcyon about 35 pectoral fin rays, four to six pelvic fin rays, approximately 20 dorsal fin rays, roughly 12 anal fin rays, and 26 caudal fin rays. By contrast, Albertonia has about 21 pectoral fin rays, nine or ten pelvic fin rays, about 17 dorsal fin rays, roughly 18 anal fin rays, and about 30 caudal fin rays (Schaeffer and Mangus 1976). Although differences in fin ray counts are potentially useful to separate taxa, the precise number of lepidotrichia is sometimes difficult to determine in fossil actinopterygians.
Additionally, with a standard length of ca. 210 mm, A/I 4360 has a comparable body size to specimens of Ospia or Watsonulus (cf. Stensiö 1932; Piveteau 1934), but differs, for instance, from Parasemionotus, Lehmanotus, Qingshania, and Suius, which seemingly only achieved maximum standard lengths of ~120 mm or less (Priem 1924; Lehman 1952; Lehman et al. 1959; Beltan 1968; Liu et al. 2002; Tong et al. 2006). The body of A/I 4360 is slender as in most parasemionotids, with the exception of the more hypsisomatic Albertonia (Schaeffer and Mangus 1976; Davies et al. 1997; Neuman 2015). Based on the preceding comparisons with other parasemionotids, we conclude that A/I 4360 from Spiti shows the most similarities with Watsonulus.
Among the new osteichthyan material from Spiti is a bone (PIMUZ A/I 4362; Fig. 8) that without doubt can be identified as an actinistian urohyal. The urohyal of actinistians is usually dorso-ventrally flattened and has the shape of an inverted Y when seen from above or below, with a narrow anterior end and a broad, bifid posterior portion. In addition, the centre part of the actinistian urohyal is bilaterally constricted (Millot and Anthony 1958; Lehman 1966; Schultze 1993; Forey 1998). The morphology of A/I 4362 is, on the other hand, well-distinguished from that of the actinopterygian urohyal. In ray-finned fishes, this element is normally either plate-like and vertically expanded (e.g. Birgeria nielseni; Lehman 1952; Arratia and Schultze 1990) or rod-like in both dorso-ventral and lateral view (teleosts; Arratia and Schultze 1990). Additionally, it may, in some cases, exhibit a repeatedly notched hind margin, which gives it a fringe-like appearance (Kusaka 1974). The urohyal of dipnoans is poorly researched (Arratia and Schultze 1990) but seemingly different from that of actinistians (see e.g. Miles 1977; Bemis et al. 1987). The urohyal occurs in various shapes and its very distinct morphology is highly diagnostic not only at higher taxonomic levels but also at the species rank (Kusaka 1974; Forey 1998).
Below, A/I 4362 (Figs. 8, 9a) is compared with the urohyal of several actinistians. Concerning the validity as well as the chronological and spatial occurrence of the included taxa we mostly adhere to Cloutier and Forey (1991) and Forey (1998). Within Actinistia, the urohyal is well-documented in the following taxa: Diplocercides kayseri (Koenen, 1895) (Fig. 9b), Rhabdoderma elegans (Newberry, 1856) (Fig. 9c), Rhabdoderma (Synaptotylus?) newelli (Hibbard, 1933) (Fig. 9d), Whiteia woodwardi Moy-Thomas, 1935 (Fig. 9e), Wimania sinuosa Stensiö, 1921 (Fig. 9f), Coelacanthus granulatus Agassiz, 1833–1843 (Fig. 9g), Trachymetopon liassicum Hennig, 1951 (Fig. 9h), Ticinepomis cf. T. peyeri (Fig. 9i), Megalocoelacanthus dobiei Schwimmer, Stewart and Williams, 1994 (Fig. 9j), Macropoma lewesiensis (Mantell, 1822) (Fig. 9k), and the recent Latimeria chalumnae Smith, 1939 (Fig. 9l). Additionally to the aforementioned species, incompletely preserved urohyalia have been described in several other fossil taxa, including (in alphabetical order) Diplurus newarki (Bryant, 1934) from the Late Triassic of North America (Schaeffer 1952: pl. 6 Fig. 4; presumed urohyal), Dobrogeria aegyssensis Cavin and Grădinaru, 2014 from the late Early Triassic of Romania (Cavin and Grădinaru 2014: Fig. 14; fragmentary preservation), Laugia groenlandica Stensiö, 1932 from the Early Triassic of East Greenland (Stensiö 1932: pl. 3; urohyal not well visible), and Undina penicillata Münster 1834 from the Late Jurassic of Germany (Reis 1888: pl. 1 Fig. 57; Reis 1892: pl. 2 Fig. 10). However, the available data on the urohyalia of most of the aforementioned taxa is not suitable for comparison.

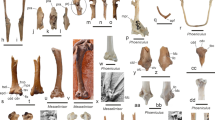
Morphological comparison of actinistian urohyalia (bone outlines in dorso-ventral aspect redrawn from the literature, anterior is on top) in a phylogenetic context (cladogram modified from Dutel et al. 2015a: Fig. 6b). a PIMUZ A/I 4362 (Actinistia indet., this study) from the Early Triassic of India, b Diplocercides kayseri (Koenen, 1895) from the Devonian of Germany (after Stensiö 1922: Fig. 8, pl. 3 Fig. 3, pl. 4 Figs. 1, 2; Stensiö 1937: pl. 1, pl. 8 Fig. 2); c Rhabdoderma elegans (Newberry, 1856) from the Carboniferous of Europe (after Aldinger 1931: Fig. 5, pl. 6; Forey 1998), d Rhabdoderma (Synaptotylus?) newelli (Hibbard, 1933) from the Carboniferous of North America (after Echols 1963: Fig. 5; see also Forey 1998); e Whiteia woodwardi Moy-Thomas, 1935 from the Early Triassic of northwest Madagascar (after Lehman 1952: Fig. 12b; Lehman 1966: Fig. 24b), f Wimania sinuosa Stensiö, 1921 from the Early Triassic of Spitsbergen (after Stensiö 1921: Fig. 29, pl. 4 Fig. 2, pl. 7 Fig. 3); g Coelacanthus granulatus Agassiz, 1833–1843 from the Lopingian of Germany and England (after Moy-Thomas and Westoll 1935: Fig. 5; Brandt 2007: Fig. 10), h Trachymetopon liassicum Hennig, 1951 from the Early Jurassic of Germany (after Hennig 1951: pl. 8 Fig. 5; Dutel et al. 2015a: Figs. 1, 4–5); i Ticinepomis cf. T. peyeri from the Middle Triassic of Switzerland (after Cavin et al. 2013: Fig. 5); j Megalocoelacanthus dobiei Schwimmer, Stewart and Williams, 1994 from the Late Cretaceous of the USA (after Dutel et al. 2012: Fig. 16b), k Macropoma lewesiensis (Mantell, 1822) from the Late Cretaceous of England (after Forey 1998: Fig. 7.7), l Latimeria chalumnae Smith, 1939 from present-day western Indian Ocean (after Millot and Anthony 1958: Figs. 15, 17, pl. 47; Adamicka and Ahnelt 1976: Fig. 3; Forey 1998: Fig. 7.6b)
Comparison of the Spiti specimen with the urohyal of other taxa reveals notable differences. Most strikingly, A/I 4362 has a generally more slender shape than the urohyal of most actinistians (Fig. 9). Additionally, the hind segment of the urohyal of Coelacanthus, Latimeria, Megalocoelacanthus, Undina, and Wimania is relatively shorter than in A/I 4362 (Reis 1888, 1892; Stensiö 1921; Moy-Thomas and Westoll 1935; Millot and Anthony 1958; Adamicka and Ahnelt 1976; Forey 1998; Brandt 2007; Dutel et al. 2012; Fig. 9). In Rhabdoderma (including Synaptotylus), Trachymetopon, and Whiteia the posterior portion of the urohyal is also distinctly wider than in A/I 4362 (Aldinger 1931; Hennig 1951; Lehman 1952; Dutel et al. 2015a). In all other taxa the constriction between the front and the hind segment of the urohyal is less pronounced compared to A/I 4362 (e.g. Stensiö 1921; Moy-Thomas and Westoll 1935; Lehman 1952, 1966; Millot and Anthony 1958; Adamicka and Ahnelt 1976; Forey 1998; Dutel et al. 2012; Cavin et al. 2013; Cavin and Grădinaru 2014; Dutel et al. 2015a; Fig. 9). In Coelacanthus, Diplocercides, Macropoma, Megalocoelacanthus, Rhabdoderma (including Synaptotylus), and Undina the tips of the posterior wings are broad (Reis 1888, 1892; Stensiö 1922, 1937; Aldinger 1931; Moy-Thomas and Westoll 1935; Forey 1998; Brandt 2007; Dutel et al. 2012), whereas in A/I 4362 and some other taxa they are acute (Fig. 9). A clear phylogenetic pattern in the outline of the actinistian urohyal (using the cladogram of Dutel et al. 2015a: Fig. 6b) is not apparent based on the available data (Fig. 9).
In summary, the morphology of A/I 4362 (Figs. 8, 9a) is distinct from the urohyal of the taxa listed above. A/I 4362 most prominently differs from other known actinistian urohyalia in its pronounced slender, elongate habitus and the extreme narrowness of its constricted portion. We therefore conclude that the specimen from Spiti does not belong to any actinistian species of which the urohyal is known.
The sarcopterygian urohyal, which is formed from cartilage, is not homologous to the teleostean urohyal, which is an ossification of the tendon of the sternohyoideus muscle (e.g. Millot and Anthony 1958; Arratia and Schultze 1990; Forey 1998). The urohyal of actinistians is a relatively large, bilaterally symmetrical median element placed postero-ventrally to the unpaired basibranchial of the hyoid arch. The urohyal articulates with the basibranchial via its broad, flat anterior end (Forey 1998). Despite its name, the urohyal does not form part of the hyoid arch. In Actinistia, the urohyal plays an important role as the point of origin of several muscles involved with mandibular depression, lowering of the buccal floor, and retraction of the shoulder girdle (Millot and Anthony 1958; Adamicka and Ahnelt 1976; Dutel et al. 2015b).
Concluding remarks
Deecke (1927) already noticed the paucity of the Himalayan record of Triassic fishes and, regrettably, our knowledge about these faunas has not significantly improved since that time. The herein presented, new Early Triassic (mainly middle-late Dienerian) marine osteichthyan fauna from Pin Valley of Spiti Subdistrict (Himachal Pradesh, India) includes the most complete fish specimens so far described from the marine and continental Triassic of India (cf. Jain and Roychowdhury 1987; Bandyopadhyay 1999; Chang and Miao 2004; Prasad et al. 2008; Gupta 2009). Regarding the wide extent of exposures and how little of it has been previously explored specifically for fish fossils there is great potential that future fieldwork in Spiti will yield more specimens. The quality of preservation is good and reveals fine morphological details (e.g. bone ornamentation and fin ray segmentation patterns). Additionally, the fish remains co-occur with abundant index fossils (ammonoids, conodonts) for precise biochronological dating. In this perspective, research on the Spiti fishes could considerably enhance our understanding of Himalayan Triassic ichthyofaunas.
Widespread marine anoxia on continental shelves during the middle and late Dienerian (Hermann et al. 2011; Ware et al. 2011) provided excellent preservational conditions for fishes and other vertebrates (e.g. Tintori 1992) and complete specimens could still be concealed in Dienerian strata of other localities that have not yet been specifically explored for such fossils (see e.g. Brinkmann et al. 2010; Ware et al. 2011). Evidently, the study of new fish faunas is vital for the advancement of palaeoichthyology and new Early Triassic assemblages can provide further insight into the evolutionary dynamics of fishes in the aftermath of the end-Permian mass extinction, a time of major osteichthyan radiation (Cavin et al. 2013; Romano et al. 2014a; Scheyer et al. 2014; Tintori et al. 2014).
Abbreviations
- CMNFV:
-
Canadian Museum of Nature (Fossil Vertebrate), Ottawa, Canada
- MNHN.F:
-
Muséum National d’Histoire Naturelle, Paris, France
- PIMUZ:
-
Paläontologisches Institut und Museum, Universität Zürich, Zürich, Schweiz
References
Adamicka, P., & Ahnelt, H. (1976). Beiträge zur funktionellen Analyse und zur Morphologie des Kopfes von Latimeria chalumnae Smith. Annalen des Naturhistorischen Museum in Wien, 80, 251–271.
Agarwal, P. N., & Singh, S. N. (1981). Recent advances in micropalaeontological investigations of the marine Triassic rocks of India. Journal of the Palaeontological Society of India, 25, 110–129.
Agassiz, L. (1833–1843). Recherches sur les poissons fossiles, II. Neuchâtel: Imprimerie de Petitpierre.
Agassiz, L. (1834). Abgerissene Bemerkungen über fossile Fische. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde, 1834, 379–390.
Aldinger, H. (1931). Über karbonische Fische aus Westfalen. Paläontologische Zeitschrift, 13(3), 186–201.
Arratia, G., & Schultze, H.-P. (1990). The urohyal: development and homology within osteichthyans. Journal of Morphology, 203(3), 247–282.
Bandyopadhyay, S. (1999). Gondwana vertebrate faunas of India. Proceedings of the Indian National Science Academy United States of America, 65(3), 285–313.
Bandyopadhyay, S., RoyChowdhury, T. K., & Sengupta, D. P. (2002). Taphonomy of some Gondwana vertebrate assemblages of India. Sedimentary Geology, 147, 219–245.
Beltan, L. (1968). La faune ichthyologique de l’Eotrias du N.W. de Madagascar: le neurocrâne. Paris: Cahiers de Paléontologie CNRS.
Beltan, L. (1980). Sur la présence d’un poisson volant, Icarealcyon malagasium, n.g. n.sp. dans l’Eotrias malgache. 26e Congrès Géologique International, 1, 155.
Beltan, L. (1984). A propos d’un poisson volant biplane de l’Eotrias du NW de Madagascar: Icarealcyon malagasium Beltan. Annales de la Société Géologique du Nord, 103, 75–82.
Beltan, L., & Janvier, P. (1978). Un nouveau Saurichthyidae (Pisces, Actinopterygii), Saurichthys nepalensis n. sp. du Trias inférieur des Annapurnas (Thakkola, Nepal) et sa signification paléobiogéographique. Cybium, 3(4), 17–24.
Bemis, W. E., Burggren, W. W., & Kemp, N. E. (1987). The biology and evolution of lungfishes. New York: Alan R. Liss Inc.
Bengtson, P. (1988). Open nomenclature. Palaeontology, 31(1), 223–227.
Berg, L. S. (1941). Lower Triassic fishes of the Tunguska coal basin, Yenisei, Siberia. Bulletin of the Academy of Sciences USSR (Biological Sciences), 1941, 414–419.
Bhargava, O. N., Krystyn, L., Balini, M., Lein, R., & Nicora, A. (2004). Revised litho- and sequence stratigraphy of the Spiti Triassic. Albertiana, 30, 21–39.
Brandt, S. (2007). Über isolierte Knochenfunde von Quastenflossern aus dem Ober-Perm von Thüringen und Hessen - Ein Beitrag zur Anatomie von Coelacanthus granulatus Agassiz. Veröffentlichungen Naturhistorisches Museum Schleusingen, 22, 69–78.
Brinkmann, W., Romano, C., Bucher, H., Ware, D., & Jenks, J. (2010). Palaeobiogeography and stratigraphy of advanced gnathostomian fishes (Chondrichthyes and Osteichthyes) in the Early Triassic and from selected Anisian localities (Report 1863–2009). Zentralblatt für Geologie und Paläontologie II, 2009(5/6), 765–812.
Brough, J. (1939). The Triassic fishes of Besano, Lombardy. London: British Museum (Natural History).
Brühwiler, T., Bucher, H., & Krystyn, L. (2012). Middle and late Smithian (Early Triassic) ammonoids from Spiti, India. Special Papers in Palaeontology, 88, 115–174.
Brühwiler, T., Ware, D., Bucher, H., Krystyn, L., & Goudemand, N. (2010). New early Triassic ammonoid faunas from the Dienerian/Smithian boundary beds at the Induan/Olenekian GSSP candidate at Mud (Spiti, Northern India). Journal of Asian Earth Sciences, 39(6), 724–739.
Bryant, W. L. (1934). New fishes from the Triassic of Pennsylvania. Proceedings of the American Philosophical Society, 73(5), 319–326.
Burgess, S. D., Bowring, S., & Shen, S. (2014). High-precision timeline for Earth’s most severe extinction. Proceedings of the National Academy of Sciences of the United States of America, 111(9), 3316–3321.
Cavin, L., Furrer, H., & Obrist, C. (2013). New coelacanth material from the Middle Triassic of eastern Switzerland, and comments on the taxic diversity of actinistans. Swiss Journal of Geoscience, 106(2), 161–177.
Cavin, L., & Grădinaru, E. (2014). Dobrogeria aegyssensis, a new early Spathian (Early Triassic) coelacanth from North Dobrogea (Romania). Acta Geologica Polonica, 64(2), 161–187.
Chang, M.-M., & Jin, F. (1996). Mesozoic fish faunas of China. In G. Arratia & G. Viohl (Eds.), Mesozoic fishes, systematics and paleoecology (pp. 461–478). München: Dr. Friedrich Pfeil.
Chang, M.-M., & Miao, D. (2004). An overview of Mesozoic fishes in Asia. In G. Arratia & A. Tintori (Eds.), Mesozoic fishes 3. Systematics, paleoenvironments and biodiversity (pp. 535–563). München: Dr. Friedrich Pfeil.
Chatterjee, S., & Roy-Chowdhury, T. (1974). Triassic Gondwana vertebrates from India. Indian Journal of Earth Sciences, 1(1), 96–112.
Chhabra, N. L., & Mishra, V. P. (2002). Middle Triassic fish teeth from the Kalapani Limestone of Malla Johar, Chamoli District (Uttaranchal). Journal of the Palaeontological Society of India, 47, 151–155.
Cloutier, R., & Forey, P. L. (1991). Diversity of extinct and living actinistian fishes (Sarcopterygii). Environmental Biology of Fishes, 32, 59–74.
Cope, E. D. (1871). Contribution to the ichthyology of the lesser Antilles. Transactions of the American Philosophical Society, 14, 445–483.
Cope, E. D. (1872). Observations on the systematic relations of the fishes. Proceedings of the American Association for the Advancement of Science, 20, 317–343.
Cope, E. D. (1887). Geology and palaeontology. Zittel’s manual on palaeontology. American Naturalist, 22(11), 1014–1019.
Davies, G. R., Moslow, T. F., & Sherwin, M. D. (1997). Ganoid fish Albertonia sp. from the Lower Triassic Montney Formation, Western Canada Sedimentary Basin. Bulletin of Canadian Petroleum Geology, 45(4), 715–718.
De Koninck, L. (1863a). Descriptions of some fossils from India, discovered by Dr. A. Fleming, of Edinburgh. The Quarterly Journal of the Geological Society of London, 19(1), 1–19.
De Koninck, L. (1863b). Notice sur les fossiles de l’Inde découverts par M. le Dr Fleming, d’Edimbourg. Mémoires de la Société Royale des Sciences de Liége, 18, 553–579.
Deecke, W. (1927). Über die Triasfische. Paläontologische Zeitschrift, 8(3), 184–198.
Diener, C. (1897). Part I: The Cephalopoda of the lower Trias. Palaeontologia Indica, series 15. Himalayan fossils, 2, 1–181.
Diener, C. (1908). Die Faunen der unteren Trias des Himalaya. Mitteilungen der Geologischen Gesellschaft in Wien, 1(1–2), 77–84.
Diener, C. (1912). The Trias of the Himálayas. Memoirs of the Geological Survey of India, 36(3), 1–176.
Dutel, H., Herbin, M., & Clément, G. (2015a). First occurrence of a mawsoniid coelacanth in the Early Jurassic of Europe. Journal of Vertebrate Paleontology, 35(3), e929581.
Dutel, H., Herrel, A., Clément, G., & Herbin, M. (2015b). Redescription of the hyoid apparatus and associated musculature in the extant coelacanth Latimeria chalumnae: functional implications for feeding. The Anatomical Record, 298(3), 579–601.
Dutel, H., Maisey, J. G., Schwimmer, D. R., Janvier, P., Herbin, M., & Clément, G. (2012). The giant Cretaceous coelacanth (Actinistia, Sarcopterygii) Megalocoelacanthus dobiei Schwimmer, Stewart & Williams, 1994, and its bearing on Latimerioidei interrelationships. PLoS ONE, 7(11), e49911.
Echols, J. (1963). A new genus of Pennsylvanian fish (Crossopterygii, Coelacanthiformes) from Kansas. Publications of the Museum of Natural History, University of Kansas, 12(10), 475–501.
Forey, P. L. (1998). History of the coelacanth fishes. London: Chapman & Hall.
Friedman, M. (2015). The early evolution of ray-finned fishes. Palaeontology, 58(2), 213–228.
Galfetti, T., Bucher, H., Ovtcharova, M., Schaltegger, U., Brayard, A., Brühwiler, T., et al. (2007). Timing of the Early Triassic carbon cycle perturbations inferred from new U–Pb ages and ammonoid biochronozones. Earth and Planetary Science Letters, 258(3–4), 593–604.
Gardiner, B. G. (1966). Catalogue of Canadian fossil fishes. Contribution/Royal Ontario Museum, Toronto, Life Sciences Division, 68, 1–154.
Goel, R. K. (1977). Triassic conodonts from Spiti (Himachal Pradesh), India. Journal of Paleontology, 51(6), 1085–1101.
Goudemand, N. (2010). Taxonomy and biochronology of Early Triassic conodonts. Ph.D. Thesis, University of Zurich.
Goudemand, N. (2014). Note on the conodonts from the Induan-Olenekian boundary. Albertiana, 42, 49–51.
Grande, L. (2010). An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy: The resurrection of Holostei. American Society of Ichthyology and Herpetology, Special Publication, 6(2A), 1–871.
Grande, L., & Bemis, W. E. (1998). A comprehensive phylogenetic study of amiid fishes (Amiidae) based on comparative skeletal anatomy. An empirical search for interconnected patterns of natural history. Society of Vertebrate Paleontology Memoir, 4, 1–690.
Griffith, J. (1959). On the anatomy of two saurichthyid fishes, Saurichthys striolatus (Bronn) and S. curionii (Bellotti). Proceedings of the Zoological Society of London, 132(4), 587–606.
Gupta, A. (2009). Ichthyofauna of the Lower Triassic Panchet Formation, Damodar valley basin, West Bengal, and its implications. Indian Journal of Geosciences, 63(3), 275–286.
Hayden, H. H. (1904). The geology of Spiti, with parts of Bashahr and Rupshu. Memoirs of the Geological Survey of India, 36(1), 1–129.
Hennig, E. (1951). Trachymetopon liassicum Ald., ein Riesen-Crossopterygier aus Schwäbischem Ober-Lias. Neues Jahrbuch für Geologie und Paläontologie Abhandlungen, 94(1), 67–79.
Hermann, E., Hochuli, P. A., Méhay, S., Bucher, H., Brühwiler, T., Ware, D., et al. (2011). Organic matter and palaeoenvironmental signals during the Early Triassic biotic recovery: The Salt Range and Surghar Range records. Sedimentary Geology, 234(1–4), 19–41.
Hibbard, C. W. (1933). Two new species of Coelacanthus from the Middle Pennsylvanian of Anderson County, Kansas. Kansas University Science Bulletin, 21(8), 279–287.
Hu, S., Zhang, Q., Chen, Z.-Q., Zhou, C., Lü, T., Xie, T., et al. (2010). The Luoping biota: exceptional preservation, and new evidence on the Triassic recovery from end-Permian mass extinction. Proceedings of the Royal Society B, 278, 2274–2282.
Huxley, T. H. (1880). On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proceedings of the scientific meetings of the Zoological Society of London, 1880, 649–662.
Jain, S. L., & Roychowdhury, T. (1987). Fossil vertebrates from the Pranhita-Godavari Valley (India) and their stratigraphic correlation. In G. D. McKenzie (Ed.), Gondwana six: Stratigraphy, sedimentology, and paleontology. Geophysical monograph series (Vol. 41, pp. 219–228). Washington D.C: American Geophysical Union.
Jin, F. (2006). An overview of Triassic fishes from China. Vertebrata PalAsiatica, 44(1), 28–42.
Krafft, A. V., & Diener, C. (1909). Lower Triassic Cephalopoda from Spiti, Malla Johar, and Byans. Palaeontologia Indica, Series 15, 6(1), 1–186.
Krystyn, L., Balini, M., & Nicora, A. (2004). Lower and Middle Triassic stage and substage boundaries in Spiti. Albertiana, 30, 40–53.
Krystyn, L., Bhargava, O. N., & Richoz, S. (2007a). A candidate GSSP for the base of the Olenekian Stage: Mud at Pin Valley; district Lahul & Spiti, Himachal Pradesh (Western Himalaya), India. Albertiana, 35, 5–29.
Krystyn, L., & Orchard, M. J. (1996). Lowermost Triassic ammonoid and conodont biostratigraphy of Spiti, India. Albertiana, 17, 10–21.
Krystyn, L., Richoz, S., & Bhargava, O. N. (2007b). The Induan-Olenekian Boundary (IOB) in Mud—An update of the candidate GSSP section M04. Albertiana, 36, 33–45.
Kusaka, T. (1974). The urohyal of fishes. Tokyo: University of Tokyo Press.
Lambe, L. M. (1916). Ganoid fishes from near Banff, Alberta. Proceedings and Transactions of the Royal Society of Canada, Series III, 10, 35–44.
Lehman, J.-P. (1952). Etude complémentaire des poissons de l’Eotrias de Madagascar. Kungliga Svenska Vetenskapsakademiens Handlingar, Fjärde Serien, 2(6), 1–201.
Lehman, J.-P. (1966). Crossopterygii. In J. Piveteau (Ed.), Traité de paléontologie. Tome IV (Vol. 3, pp. 301–412). Paris: Masson et Cie.
Lehman, J.-P., Château, C., Laurain, M., & Nauche, M. (1959). Paléontologie de Madagascar XXVII. Les poissons de la Sakamena moyenne. Annales de Paleontologie, 45, 175–219.
Li, Q. (2009). A new parasemionotid-like fish from the Lower Triassic of Jurong, Jiangsu Province, South China. Palaeontology, 52(2), 369–384.
Liu, G.-B., Feng, H.-Z., Wang, J.-X., Wu, T.-M., & Zhai, Z.-H. (2002). Early Triassic fishes from Jurong, Jiangsu. Acta Palaeontologica Sinica, 41(1), 27–52.
López-Arbarello, A. (2004). The record of Mesozoic fishes from Gondwana (excluding India and Madagascar). In G. Arratia & A. Tintori (Eds.), Mesozoic fishes 3. Systematics, paleoenvironments and biodiversity (pp. 597–624). München: Dr. Friedrich Pfeil.
Mantell, G. (1822). The fossils of the South Downs; or illustrations of the geology of Sussex. London: Lupton Relfe.
Mehrotra, D. K., Dass, S., & Sehgal, A. (1983a). Occurrence of fish microremains from the Kuti and Kalapani area, Kumaon Himalaya, Uttar Pradesh. Geoscience Journal, 4(2), 205–206.
Mehrotra, D. K., Sahgal, A., & Jangpangi, B. S. (1983b). On some Early Triassic fish microremains from the Shalshal area of Kumaun Himalaya, Uttar Pradesh. Himalayan Geology, 11, 433–437.
Miles, R. S. (1977). Dipnoan (lungfish) skulls and the relationships of the group: a study based on new species from the Devonian of Australia. Zoological Journal of the Linnean Society, 61(1–3), 1–328.
Millot, J., & Anthony, J. (1958). Anatomie de Latimeria chalumnae. Tome I. Squelette, muscles et formations de soutien. Paris: Éditions du Centre National de la Recherche Scientifique.
Mishra, V. P., Mehrotra, D. K., Pande, A. C., & Ali, Md A. (1990). A teleostean fish from the Lower Triassic of Chamba, Himachal Pradesh. Journal of the Palaeontological Society of India, 35, 73–76.
Misra, R. C., Sahni, A., & Chhabra, N. (1973). Triassic conodonts and fish remains from Niti Pass, Kumaun Himalaya. Himalayan Geology, 3, 148–161.
Moy-Thomas, J. A. (1935). The coelacanth fishes from Madagascar. Geological Magazine, 72(5), 213–227.
Moy-Thomas, J. A., & Westoll, T. S. (1935). On the Permian coelacanth, Coelacanthus granulatus, Ag. Geological Magazine, 72(10), 446–457.
Müller, J. (1845). Ueber den Bau und die Grenzen der Ganoiden, und über das natürliche System der Fische. Archiv für Naturgeschichte, 11(1–2), 91–141.
Mutter, R. J., Cartanyà, J., & Basaraba, S. A. U. (2008). New evidence of Saurichthys from the Lower Triassic with an evaluation of early saurichthyid diversity. In G. Arratria, H.-P. Schultze, & M. V. H. Wilson (Eds.), Mesozoic fishes 4, homology and phylogeny (pp. 103–127). München: Dr. Friedrich Pfeil.
Neuman, A. G. (2015). Fishes from the Lower Triassic portion of the Sulphur Mountain Formation in Alberta, Canada: geological context and taxonomic composition. Canadian Journal of Earth Sciences, 52(8), 557–568.
Neuman, A. G., & Mutter, R. J. (2005). Helmolepis cyphognathus, sp. nov., a new platysiagid actinopterygian from the Lower Triassic Sulphur Mountain Formation (British Columbia, Canada). Canadian Journal of Earth Sciences, 42, 25–36.
Newberry, J. S. (1856). Description of several new genera and species of fossil fishes, from the Carboniferous strata of Ohio. Proceedings of the Academy of Natural Sciences of Philadelphia, 8, 96–100.
Nielsen, E. (1936). Some few preliminary remarks on Triassic Fishes from East Greenland. Meddelelser om Grønland, 112(3), 1–55.
Olsen, P. E. (1984). The skull and pectoral girdle of the parasemionotid fish Watsonulus eugnathoides from the Early Triassic Sakamena Group of Madagascar, with comments on the relationships of holostean fishes. Journal of Vertebrate Paleontology, 4(3), 481–499.
Ovtcharova, M., Goudemand, N., Hammer, Ø., Guodun, K., Cordey, F., Galfetti, T., et al. (2015). Developing a strategy for accurate definition of a geological boundary through radio-isotopic and biochronological dating: the Early-Middle Triassic boundary (South China). Earth Science Reviews, 146, 65–76.
Owen, R. (1860). Palaeontology or a systematic summary of extinct animals and their geological relations. Edinburgh: A & C Black.
Patterson, C. (1973). Interrelationships of holosteans. In P. H. Greenwood, R. S. Miles, & C. Patterson (Eds.), Interrelationships of fishes (pp. 233–305). London: Academic Press.
Patterson, C. (1975). The braincase of pholidophorid and leptolepid fishes, with a review of the actinopterygian braincase. Philosophical Transactions of the Royal Society of London B, 269(899), 275–579.
Patterson, C. (1982). Morphology and interrelationships of primitive actinopterygian fishes. American Zoologist, 22(2), 241–259.
Piveteau, J. (1929). Sur un type de poisson fossile provenant du nord de Madagascar. Comptes Rendus Hebdomadaires des Séances de l’Académie des Sciences, 189, 1017–1019.
Piveteau, J. (1934). Paléontologie de Madagascar XXI. — Les poissons du Trias inférieur. Contribution à l’étude des actinoptérygiens. Annales de Paleontologie, 23, 83–178.
Piveteau, J. (1939–1940). Paléontologie de Madagascar XXIV. Nouvelles recherches sur les poissons du Trias inférieur. Annales de Paleontologie, 28, 71–88
Prasad, G. V. R., Singh, K., Parmar, V., Goswami, A., & Sudan, C. S. (2008). Hybodont shark teeth from the continental Upper Triassic deposits of India. In G. Arratria, H.-P. Schultze, & M. V. H. Wilson (Eds.), Mesozoic fishes 4, homology and phylogeny (pp. 413–432). München: Dr. Friedrich Pfeil.
Priem, F. (1924). Paléontologie de Madagascar XII. Les poissons fossiles. Annales de Paléontologie, 13, 107–132.
Regan, C. T. (1923). The skeleton of Lepidosteus, with remarks on the origin and evolution of the lower neopterygian fishes. Proceedings of the Zoological Society of London, 93(2), 445–461.
Reis, O. M. (1888). Die Coelacanthinen, mit besonderer Berücksichtigung der im Weissen Jura Bayerns vorkommenden Gattungen. Palaeontographica, 35, l–94.
Reis, O. M. (1892). Zur Osteologie der Coelacanthinen. I. Theil (Rumpfskelet, Knochen des Schädels und der Wangen, Kiemenbogenskelet, Schultergürtel, Becken, Integument und innere Organe). Inaugural-Dissertation, K. Ludwigs-Maximilians-Universität zu München.
Romano, C., Kogan, I., Jenks, J., Jerjen, I., & Brinkmann, W. (2012). Saurichthys and other fossil fishes from the late Smithian (Early Triassic) of Bear Lake County (Idaho, USA), with a discussion of saurichthyid palaeogeography and evolution. Bulletin of Geosciences, 87(3), 543–570.
Romano, C., Koot, M. B., Kogan, I., Brayard, A., Minikh, A. V., Brinkmann, W., et al. (2014a). Permian-Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution. Biological Reviews. doi:10.1111/brv.12161.
Romano, C., Kriwet, J., Baal, C., & Brinkmann, W. (2014b). Recovery from the Late Permian mass extinction: new insights from a neglected Early Triassic fish fauna from the Salt Range (Pakistan, Neotethys realm). In: 74th annual meeting of the Society of Vertebrate Paleontology, Berlin, Meeting Program and Abstracts, 216.
Romer, A. S. (1955). Herpetichthyes, amphibioidei, choanichthyes or sarcopterygii? Nature, 176, 126.
Rosen, D. E., Forey, P. L., Gardiner, B. G., & Patterson, C. (1981). Lungfishes, tetrapods, paleontology, and plesiomorphy. Bulletin of the American Museum of Natural History, 167(4), 159–276.
Sahni, A., & Chhabra, N. L. (1976). Microfish remains from certain Triassic sections in the Kashmir and Kumaun Himalayas. In: Proceedings of the VI Indian colloquium on micropaleontology and stratigraphy (pp. 218–224).
Schaeffer, B. (1952). The Triassic coelacanth fish Diplurus, with observations on the evolution of the Coelacanthini. Bulletin of the American Museum of Natural History, 99(2), 25–78.
Schaeffer, B., & Mangus, M. (1976). An Early Triassic fish assemblage from British Columbia. Bulletin of the American Museum of Natural History, 156(5), 127–216.
Scheyer, T. M., Romano, C., Jenks, J., & Bucher, H. (2014). Early Triassic marine biotic recovery: the predators’ perspective. PLoS ONE, 9(3), e88987.
Schultze, H.-P. (1993). Patterns of diversity in the skulls of jawed fishes. In J. Hanken & B. K. Hall (Eds.), The skull 2, patterns of structural and systematic diversity (pp. 189–254). Chicago: University of Chicago Press.
Schwimmer, D. R., Stewart, J. D., & Williams, G. D. (1994). Giant fossil coelacanths of the Late Cretaceous in the eastern United States. Geology, 22(6), 503–506.
Smith, J. L. B. (1939). A living fish of Mesozoic type. Nature, 143, 455–456.
Srivastava, J. P., & Mehrotra, D. K. (1986). Microfish remains from the Triassics of Kashmir Himalaya. Records of the Geologcal Survey of India, 113(8), 91–99.
Stensiö, E. (1921). Triassic fishes from Spitzbergen 1. Wien: Adolf Holzhausen.
Stensiö, E. (1922). Über zwei Coelacanthiden aus dem Oberdevon von Wildungen. Paläontologische Zeitschrift, 4(2–3), 167–210.
Stensiö, E. (1925). Triassic fishes from Spitzbergen 2. Kungliga Svenska Vetenskapsakademiens Handlingar, Tredje Serien, 2(1), 1–261.
Stensiö, E. (1932). Triassic fishes from East Greenland 1–2. Meddelelser om Grønland, 83(3), 1–305.
Stensiö, E. (1937). On the Devonian coelacanthids of Germany with special reference to the dermal skeleton. Kungliga Svenska Vetenskapsakademiens Handlingar, Tredje Serien, 16(4), 1–56.
Tintori, A. (1992). Fish taphonomy and Triassic anoxic basins from the Alps: a case history. Rivista Italiana di Paleontologia e Stratigrafia, 97(2–3), 393–408.
Tintori, A., Hitij, T., Jiang, D.-Y., Lombardo, C., & Sun, Z.-Y. (2014). Triassic actinopterygian fishes: The recovery after the end-Permian crisis. Integrative Zoology, 9(4), 394–411.
Tong, J., Zhou, X., Erwin, D. H., Zuo, J., & Zhao, L. (2006). Fossil fishes from the Lower Triassic of Majiashan, Chaohu, Anhui Province, China. Journal of Paleontology, 80(1), 146–161.
Tozer, E. T. (1965). Lower Triassic stages and ammonoid zones of Arctic Canada. Paper of the Geological Survey of Canada, 65, 1–14.
Uyeno, T. (1978). On some Lower Triassic fishes from Ankilokara, Madagascar. Bulletin of the National Science Museum, Series C (Geology & Paleontology), 4(4), 193–198.
von Koenen, A. (1895). Über einige Fischreste des norddeutschen und böhmischen Devons. Abhandlungen der Königlichen Gesellschaft der Wissenschaften zu Gottingen, 40, 1–37.
Waagen, W. (1895). Salt-Range fossils 2. Fossils from the Ceratite Formation, Part I: Pisces—Ammonoidea. Memoir of the Geological Survey of India, Palaeontologia Indica, Series 13, 2, 1–323.
Ware, D., Jenks, J. F., Hautmann, M., & Bucher, H. (2011). Dienerian (Early Triassic) ammonoids from the Candelaria Hills (Nevada, USA) and their significance for palaeobiogeography and palaeoceanography. Swiss Journal of Geosciences, 104(1), 161–181.
Ware, D., Bucher, H., Brayard, A., Schneebeli-Hermann, E., & Brühwiler, T. (2015). High-resolution biochronology and diversity dynamics of the Early Triassic ammonoid recovery: the Dienerian faunas of the Northern Indian Margin. Palaeogeography, Palaeoclimatology, Palaeoecology. doi:10.1016/j.palaeo.2015.09.013.
Ware, D., Bucher, H., Brühwiler, T., & Krystyn, L. (in production a). Dienerian (Early Triassic) ammonoids from Spiti (Himachal Pradesh, India). Fossil & Strata.
Ware, D., Bucher, H., Brühwiler, T., Schneebeli-Hermann, E., Hochuli, P.A., Roohi, G., et al. (in production b). Griesbachian and Dienerian (Early Triassic) ammonoids from the Salt Range, Pakistan. Fossil & Strata.
Xu, G.-H., Zhao, L.-J., & Coates, M. I. (2014). The oldest ionoscopiform from China sheds new light on the early evolution of halecomorph fishes. Biology Letters, 10(5), 20140204.
Zhang, M. (1976). A new species of helicoprionid shark from Xizang. Scientia Geologica Sinica, 1976(4), 332–336.
zu Münster, G. (1834). Mittheilungen an Professor Bronn gerichtet. Neues Jahrbuch für Mineralogie, Geognosie, Geologie und Petrefaktenkunde, 1834, 538–542.
Acknowledgments
We greatly appreciate discussions with Ilja Kogan (Department of Palaeontology, Geological Institute, TU Bergakademie Freiberg, Germany), Piotr and Roksana Skrzycki (Krakow, Poland), as well as Thodoris Argyriou, Richard Hofmann, and Michael Hautmann (all PIMUZ) during preparation of the manuscript. Gaël Clément and Monette Véran (both MNHN.F) are thanked for their hospitality and help during visit of the Madagascar collection at the Paris museum in 2012. We thank Lui Unterrassner (Zürich) for assistance during field work in 2009. Rosi Roth (PIMUZ) is thanked for photographs and preparation of the cast of PIMUZ A/I 4360 and Ashley Latimer (PIMUZ) for correcting the English. We highly appreciate critical remarks by Adriana López-Arbarello (Bayerische Staatssammlung für Paläontologie, Munich, Germany) and a second, anonymous reviewer, which helped to improve the quality of this contribution. We deeply acknowledge support by the Swiss National Science Foundation (project numbers 120311/135075 and 144462 to W.B. and H.B, and 200021/135446 to H.B.).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Romano, C., Ware, D., Brühwiler, T. et al. Marine Early Triassic Osteichthyes from Spiti, Indian Himalayas. Swiss J Palaeontol 135, 275–294 (2016). https://doi.org/10.1007/s13358-015-0098-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-015-0098-6