
Abstract
The material for the present paper is based on 50 kg of processed sediment from the Early Kimmeridgian of Geisingen. The most frequent crinoids comprise cups and brachials of the roveacrinid Saccocoma quenstedti while Crassicoma feifeli is represented by a few cups only; these remains are not described. Cups and primibrachials of Eugeniacrinites cariophilites are the commonest cyrtocrinids, while cups of Pilocrinus moussoni are rather rare and not further treated, as are columnals of the two species. The ontogenetic series of cups and axillary primibrachials of E. cariophilites starts from ossicles of about 0.5 mm in height; and growth is allometric in both types. Postlarval cups resemble phyllocrinids; the distinctive shape with lanceolate interradial extensions and wide articular facets appears at a later stage. The spearhead-shaped axillary primibrachials are synapomorphic. The youngest of these ossicles with a muscular facet proximally were able to form a flat roof over the radial cavity. First primibrachials with synostosial distal facet appeared at a slightly later stage with a width of 1 mm; growth is isometric. A significant number of cups are mutilated by bite marks. The predatory attacks were sublethal in most cases as shown by secondary stereom growth and regenerated facets. First primibrachials are not deformed, but deformations occur in a small number of second primibrachials. The results suggest that the predators, presumably fishes, targeted the soft parts in the cup. Enigmatic brachials with a large wing to one side and a facet to an arm are thought to belong to E. cariophilites and presumably attached to deformed cups; the wings may have been an antipredatory adaptation providing some shelter to the cup. The various antipredator morphologies in cyrtocrinid history are discussed. The presently available growth series of Eugeniacrinites invites discussion of early cyrtocrinid phylogeny. Eugeniacrinites seems to have been derived from Phyllocrinidae; and Sclerocrinidae with Cyrtocrinus as the first representative in the Middle Jurassic is thought to have originated from Early Jurassic Tetracrinidae such as Bilecicrinus.
Similar content being viewed by others

Introduction
The cyrtocrinid Eugeniacrinites cariophilites (von Schlotheim 1813); Parkinson’s (1808) “clove encrinite”, is perhaps the best known of its kind and one of the oldest fossils on record. Wagner (1684) recorded the species under the name Caryophyllos aromaticos, the petrified fruit of the clove tree (Eugenia caryophyllata). Lhwyd (1699) was the first to refer it to the crinoids. Pre-Linnaean names include Caryophyllitarum Rosinus 1718, Caryophyllis Scheuchzer 1752, Caryophyllite Knorr 1755, Caryophyllitea Walch 1762, and Caryophyllites Knorr and Walch (1769). Rosinus (1718, pl. 3A, B) already figured quite a number of cups showing their variety, mostly specimens with narrow base, but also thickset ones and even a tetramerous cup. This peculiar and distinctive cyrtocrinid is known to most collectors as Eugeniacrinus caryophyllatus (Goldfuss 1829), and innumerable specimens are preserved in collections, both public and private. Despite the wealth of material and numerous descriptions, among them Quenstedt’s (1876) extensive record on the variety of cups and axillary primibrachials and Jaekel’s (1891) classic paper, the species has still not revealed all its secrets. For example, the early ontogenetic stages have not been described in detail to date. The present paper is based on material collected and processed by A. S. Gale from Kimmeridgian mudstones of the Geisingen (Danube) quarry (Fig. 1). The larger elements of the crinoid fauna mostly belong to E. cariophilites, represented by cups, primibrachials, attachment discs and columnals. Cups of Pilocrinus moussoni are uncommon. However, the cyrtocrinids are outnumbered by broken cups and brachials of the delicate roveacrinid Saccocoma quenstedti Sieverts-Doreck and Hess 2002 (in Hess 2002), while Crassicoma feifeli Sieverts-Doreck and Hess 2002 is represented by only a few cups. The fine fraction of the residue includes many cups and brachials of E. cariophilites that represent early ontogenetic stages. Some of the cups are preserved with primibrachials attached, a feature hitherto undescribed. A considerable number of cups are deformed or crippled, and these may explain the most peculiar wing plates of a type so far unknown in articulate crinoids.

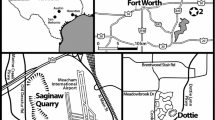
Location of the crinoid bed at the Geisingen quarry. Photo A. S. Gale
E. cariophilites ranges from the Middle Oxfordian sponge facies of Germany, France and Switzerland to the Lower Kimmeridgian of Germany. The assignment by Rasmussen (1961) of cups from the Early Cretaceous of France to this species is questionable. The remains have been assigned to a single species, although the stratigraphic range seems rather extensive. However, the large variability common to cyrtocrinids makes the definition of specific boundaries difficult (see Hess 2012). Major sites are in the Birmenstorf Member (Transversarium Zone) of Switzerland, the Marnes à spongiares (Transversarium Zone) of France, and the Lochen facies (Bimammatum to Divisum Zones) of southern Germany.
Locality and material
The material was collected by A. S. Gale in the Geisingen (Danube) quarry. The now mostly disused quarry is about 2 km northeast of the town; it was formerly run by Holcim and now belongs to Geisinger Kalkstein Schotterwerk (GKS). About 50 kg of sediment were collected from a 1 m thick greenish mudstone with common rhynchonellids, several metres above the base of the Kimmeridgian in the Lacunosamergel Formation (Fig. 1). According to A. S. Gale the bed is in the Platynota Zone, but it should rather be placed at the base of the Divisum Zone (G. Schweigert, personal communication 2013). The processed part of the residue with the crinoids contains mainly roveacrinids and cyrtocrinids. The majority of the latter belongs to E. cariophilites and includes 470 cups, 299 first primibrachials and 767 second primibrachials. Tentatively assigned to this species are 136 “wing plates”. Pilocrinus moussoni (Desor) is represented by 36 cups, three of them deformed and revealing bite marks. In addition are numerous brachials, columnals and some attachment discs. Columnals and discs cannot easily be assigned to one of the cyrtocrinid species in many cases and are not further discussed. Only half a dozen of columnals show superficial lesions or bite marks and none have significant growth of stereom to repair the damage. A few isocrinid columnals and some brachials that include a comatulid species are also present. Preservation of all ossicles is excellent, with no signs of abrasion or weathering.
The material is housed in the Natural History Museum Basel (M followed by registration numbers).
Ontogeny
The majority of specimens figured in the literature (cups, first and second primibrachials, columnals) correspond in size and shape to those in the reconstruction in Fig. 2a, based on Jaekel’s (1907) work, which also illustrates the unique find of first secundibrachials still attached to a second primibrachial (Fig. 2b). The material includes many long columnals, suggesting a fairly long column. The longest columnals have a length of 11 mm at a diameter of 1.5 mm; and the facets correspond to a pluricolumnal of four attached to the cup figured by Jaekel (1891, pl. 40, fig. 1), the longest preserved column in the literature. Figure 3 illustrates the size range of selected examples of cups and primibrachials discussed in the present study.

Eugeniacrinites cariophilites; a reconstruction (after Ubaghs 1953); b second primibrachial with attached first secundibrachial, Oxfordian, Germany, X1 (after Quenstedt 1876, pl. 106, fig. 71; and Jaekel 1891, fig. 25). Cup shaded dark-grey, first primibrachial (IBr1) white, second primibrachial (IBr2) light-grey, IIBr1 = first secundibrachial

Growth stages of Eugeniacrinites cariophilites at same magnification; a–f cups in lateral view (f with attached first columnal); g–k first primibrachials (IBr1), g–i proximal facet, j distal facet, k adoral view with large muscle facets to radial; l–m adoral view of juvenile, axillary primibrachials (IBr) with muscular proximal facet; n–o adoral view of second primibrachials (IBr2) with synostosial proximal facet. Reference to ossicles: a Fig. 4a2, b Fig. 4f, c Fig. 4g, d Fig. 4i, e Fig. 5c2, f Fig. 5e; g Fig. 7a, h Fig. 7d, i Fig. 7e, j Fig. 7f, k Fig. 7g; l Fig. 8a, m Fig. 8b, n Fig. 9b, o Fig. 9e. Scale bar 1 mm
Cups
Among the total number of cups are 31 (6.6 %) with a diameter lesser than 1 mm; the height of such cups is 70–80 % of maximum diameter (Fig. 4). The smallest cup has a diameter of 0.6 mm (Fig. 4a). Such cups are bowl shaped to conical; with radial ribs and a blunt apex that lacks a distinct symplectial facet for the column (Fig. 4a–e). In distal view the profile is rounded pentagonal. The side view is predominated by the radial bulges or ribs that widen at the upper end for the reception of small, circular facets for the primibrachials. They are the highest points in these small cups. The interradial areas are concave; the radial cavity is wide and unsculptured. Later in development the cups assume a pentalobate profile, and the lobes with their facets are separated in the cavity by interradial buttresses (Figs. 5d, 6). On the outside weak ribs may occur at the suture between the radials (Fig. 4d, g–k) but these interradial ribs disappear with growth (Fig. 5). With increasing size of the animal the interradial areas extend upwards (Fig. 4e, f–i) and rise above the now sunken articular facets. At diameters of >2 mm the cups achieve the distinctive shape of adults (Fig. 5). Adult cups vary in shape, as described in the literature (Quenstedt 1876). Most cups in the present material have a slender lower part, especially in juvenile individuals. Among larger specimens a considerable number (68 of the total or 14.4 %) are thickset (Fig. 5f). One cup is low, disc-like and lacks a shaft; the facet to the column is partly deformed; the unusual shape seems to result from an attack by a predator (Fig. 5g). de Loriol (1882, p. 130; pl. 13, fig. 7) described a very low cup as “monstrueux, ayant une base énorme”. However, the facet to the column is symplectial and intact in this case. The largest cup in the material is 8 mm in height, at a diameter of 7 mm distally. Thickset cups may be as wide distally as at the base (e. g. Hess 1975, pl. 23, fig. 2); the slightly smaller cup in Fig. 5f is as wide as high, but has a narrower, rounded base.

Prejuvenile (a–e) and juvenile cups (f–k) of Eugeniacrinites cariophilites. a Cup with narrow radial ridges and concave interradial areas, upper margin nearly straight, facets to primibrachials small and circular; M11417; a1 distal view, a2 lateral. b Lateral view of cup with slightly larger facets to primibrachials and concave upper interradial margins, M11418. c Aboral view of cup similar to a, facet to column synostosial; M11419. d Aboral (proximal) view of cup with wider facets to primibrachials, interradial areas with faint ridge; M11420. e Lateral view of cup with facets to primibrachials sunken relative to interradial areas, interradial processes blunt; M11421. f Lateral-distal view of small juvenile cup with facets to primibrachials distinctly sunken between lanceolate interradial processes; M11422. g Lateral view of cup with strongly developed radial ridges and deep concave interradial areas, facet to column wide; M11423. h Lateral view of cup with sharp radial ridges, facet to column narrow; M11424. i Lateral-distal view of cup with narrow lower part, strongly raised interradial processes with adjoining ribs near sutures; M11425. j Aboral view of cup with interradial ribs along sutures, facet to column synostosial, M11426. k Aboral view of cup with irregular interradial ribs, facet to column weakly symplectial, M11427. Scale bar 1 mm, upper bar a–e, lower bar f–k

Juvenile (a–d) and adult cups (e–g) of Eugeniacrinites cariophilites. a Lateral view of cup with wide articular facets, bulging radial areas separated by interradial notches, interradial processes high; M11428. b Lateral-distal view of cup with narrow articular facets, strongly bulging radial areas separated by interradial notches, interradial processes broad and high; M11429. c Cup with wide articular facets, strongly bulging radial areas separated by interradial notches; M11430; c1 aboral with symplectial facet to column; c2 lateral, note sculptured interradial processes along sutures in radial cavity. d Adoral view of cup, note interradial buttresses along sutures; M11431. e Cup with attached first columnal, such morphology with wide articular facets and flat column-like lower part corresponds to most of the finds; M11432. f Thickset cup, M11433. g Lateral view of low cup with damaged passage to column, presumably caused by predator, M11434. Scale bar 1 mm

Distal view of juvenile cups of Eugeniacrinites cariophilites with attached axillary primibrachials. The primibrachials are of the type figured in Fig. 8a, with proximal muscular facet; and they resemble Fig. 8d in lateral view. The primibrachial in Fig. 6f is slightly displaced and shows a lateral notch for the attachment of a first secundibrachial (arrow). Specimens c and d are slightly asymmetrical. Note interradial buttresses in the radial cavity. a M11435, b M11436, c M11437, d M11438, e M11439, f M11440. Scale bar 1 mm
During growth the articular facets widen while the interradial concavity narrows to a deep suture separating bulging radial areas (Fig. 5a–c). Eventually, there are only weakly expressed sutures between the radials, and the surface of the cup is even (Fig. 5e, f). At all stages the articular facets have deep muscle fossae; these are circular in small cups and become oval in larger ones, in line with the widening facet (Fig. 5e, f). Relative proportions of aboral ligament pit, transverse ridge and axial canal remain roughly constant during growth. Interarticular ligament areas are indistinct in juvenile cups. Adult cups have a deep median notch that separates narrow bands on each side. These were erroneously considered to be muscle fossae in both the Treatise of 1978 (Rasmussen 1978, p. T833) and 2011: “Radial articular facets steep, low, and wide, with small, aboral ligament fossa, deeply excavated interarticular fossae and low adoral muscle fossae separated by ridge parallel to fulcral ridge” (Hess & Messing, p. 181). In fact, the interarticular ligament fossae remain indistinct during growth while the muscle fossae are deep from the early stages onwards (Figs. 4i, 5d). The narrow bands or ledges may be more or less excavated (Fig. 5e, f); such morphology is not seen at the earlier stages. In all first primibrachials the proximal facet has flat ledges above the muscle fossae for articulation with the bands (Fig. 7e, g). As a consequence, the Treatise diagnosis the should be modified as follows: “Radial articular facets steep and wide in adults, with small aboral ligament fossa and deeply excavated adoral muscle fossae, separated from radial cavity by narrow ledges extending from interradial processes to median notch; interarticular ligaments indistinct”. The presence of deep muscle fossae and a small aboral ligament fossa or pit in combination with indistinct (or missing) interarticular ligament fossae is common in cyrtocrinids, especially in phyllocrinids (Hess 2012, fig. 13).

First primibrachials of Eugeniacrinites cariophilites with muscular proximal facets (a–e) and synostosial distal facet (f). a–e Proximal view; a M11442, b M11443, c M11444, d M11445, e M11446. f Aboral-distal view with synostosial facet to second primibrachial, M11447. g Adoral view with muscle fossae below, separated by radial furrow, and synostosial facet to second primibrachial above, M11442. Scale bar 1 mm
The significance of excavated bands above the muscle fossae is unclear, although one might speculate that they served as attachment for ligaments. However, ligaments positioned adorally to muscles are not observed in crinoids; because arm movement occurs by muscle contraction and antagonistic recoil of aboral ligament during muscle relaxation. In E. cariophilites the aboral ligament fossa is unusually small in comparison with the conspicuous muscle fossae. Such discrepancy may indicate that muscle relaxation leading to separation of the second primibrachials at their tip and unfolding of the arms (Fig. 2a) was sluggish in relation to closure by muscle contraction. Cautious opening and rapid closure would have made sense to a crinoid in danger of predators.
Primibrachials
The first primibrachials are wide and low (Fig. 7); they are mostly symmetrical but some are more or less asymmetrical, with unequal muscle fossae (Fig. 7d). The proximal facet is muscular, with wide muscle fossae that deepen towards the adradial notch; the interarticular ligament areas are indistinct and separated by a sharp transverse ridge from the small, deep aboral ligament pit. The axial canal is circular in small specimens and oval in larger ones. The distal facet is synostosial; the aboral side is smooth, and the adoral side shows a deep food groove (Fig. 7g). No first primibrachial with muscular articulation at both facets has been found, all have a synostosial distal facet.
The second primibrachials are axillary; the smallest have a total width of 1–1.5 mm and a width of 0.5 mm at the proximal facet, these facets are muscular (Fig. 8a–g; not figured are eight ossicles, the largest is 1.7 mm in height at a total width of 1.6 mm). Slightly larger ossicles have an indistinct muscular to synostosial proximal facet (Figs. 8h–j, 9a). Thus, the change of a muscular to a synostosial facet seems gradual. The presence of muscular proximal facets in the smallest axillary primibrachials means that differentiation into first and second primibrachials joined by synostosis occurred later in development. In fact, the smallest first primibrachials in the material have a width of approximately 1 mm. Such development is confirmed by the occurrence of juvenile cups with attached axillary primibrachials that seal the radial cavity (Fig. 6). These primibrachials have a muscular facet proximally. In the smallest cups the muscle fossae are displaced to some extent towards the radial cavity. This allowed the primibrachials to rise from a horizontal position (Fig. 6a) to a vertical one, exposing the food grooves to the cavity and allowing the arms to uncoil for food collection. Small axillary primibrachials are arrowhead-shaped and only slightly bent (Fig. 8a, d). The facets to the secundibrachials are not seen from above in cups with primibrachials preserved in situ, but they do show up when an ossicle is slightly turned as in Fig. 6f (arrow). The small cups with attached primibrachials resemble the cups of Cyathidium pourtalesi Améziane et al. (1999, fig. 6a, b). The coiled arms with only a few secundibrachials are illustrative of juvenile E. cariophilites.

Juvenile primibrachials of Eugeniacrinites cariophilites. a–g Primibrachials with muscular proximal facet; a–c adoral, d lateral view of nearly straight specimen; e–g adoral-proximal view to show muscle fossae more clearly. h–i adoral-proximal view of first primibrachials with synostosial facet, j lateral view of curved specimen. a M11449, b M11450, c M11451, d M11452, e M11453, f M11454, g M11455, h M11456, i M11457, j M11458. Scale bar 1 mm

Adoral view of primibrachials of Eugeniacrinites cariophilites of various shapes, with muscular proximal facet (a) and synostosial proximal facet (b–f). a M11459, b M11461, c M11505, d M11460, e M11462, f M11463. Scale bar 1 mm
Second primibrachials of postjuvenile to adult individuals are much more variable than the rather uniform first primibrachials. To maintain the possibility of sealing the radial cavity the ossicles are more strongly bent and are joined by their thickened extensions (Fig. 10g, h). While adjoining ossicles are symplectially articulated at the lower part, articulation between the spearhead-shaped extensions may vary from symplectial to nearly synostosial. Some of the odd shapes of these ossicles are discussed in the section dealing with crippled ossicles.

Second primibrachials of Eugeniacrinites cariophilites. a Adoral view of specimen missing interradial projection, M11464. b Adoral view of specimen with wide interradial projection displaced to one side, M11465. c Adoral view of specimen without interradial projection, area between articular facets is damaged, presumably by predator; M11466. d Adoral view of specimen with forked interradial projection, M11467. e Adoral view of specimen with strongly bent interradial projection, the specimen is slightly turned to the left show the facet for the first secundibrachial; M11468. f Adoral view of pathological ossicle with articular facet for only one arm on extension at right, narrow symplectial facet at left for articulation with adjoining ossicle; a triangular, small first primibrachial (arrow) is fused to the second primibrachial; M11469. g Lateral view of specimen with interradial projection at low angle to body axis, M11470. h Lateral view of specimen with interradial projection at high angle to body axis, M11471. Scale bar 1 mm
Discussion
Early ontogenetic stages of cyrtocrinids described in the literature are scarce. Manni and Nicosia (2004) reported on a growth series of the sclerocrinid Gammarocrinites bakonicus Manni et al. (1992) from the Late Jurassic of Hungary. From the smallest specimen with a cup diameter of 1 mm up to specimens of about 4 mm overall morphology is similar. Of more interest is a sequence of Phyllocrinus sabaudianus Pictet and de Loriol (1858) from the Early Cretaceous (see de Loriol 1879, p. 240, pl. 19, fig. 31–32) described by Arendt (1974, p. 122, fig. 22). Adult specimens have the interradial processes distinctive for the genus. The processes are developed already in a specimen with a height of 1 mm and increase allometrically with size of the specimens. Only the smallest specimen with a cup height of 0.75 mm lacks the processes and is highest near the facets. But for the lack of radial ribs it is remarkably similar to the cups in Fig. 4a–b. In both forms the interradial areas grow upwards along the radial sutures, although to a much larger degree in P. sabaudianus. In contrast to E. cariophilites growth of the facets is isometric in P. sabaudianus. In a growth series of Phyllocrinus sinuatus (Remeš 1912) from the Early Cretaceous reported by Žítt (1978) the smallest cup with a height of 3 mm has interradial processes of similar relative dimensions as the largest cup of 8.5 mm height, growth is thus largely isometric.
In their apprisal of cyrtocrinid phylogeny Pisera and Dzik (1979, fig. 14, p. 832) considered Phyllocrinidae and Eugeniacrinitidae to be “very closely related and descended from primitive sclerocrinids” although this is not obvious in their phylogenetic tree. They considered the Early Jurassic Sclerocrinus deslongchampsi (de Loriol 1882) a basic form. This species, whose secundibrachials and pinnules are unknown, is now assigned to Quenstedticrinus and classified with Plicatocrinoidea (Hess and Messing 2011). According to the present material Eugeniacrinites and its kind may have been derived from Phyllocrinidae that include the earliest members of the Suborder Eugeniacrinitoidea, Nerocrinus Manni and Nicosia 1999 and Ticinocrinus Hess 2006. These genera possess well-developed basals, another indication of the primitive character of the family. Phyllocrinid cups have small, sunken articular facets between interradial processes of various lengths that shelter the small arms. The bulk of the known species belongs to the genus Phyllocrinus with mostly small cups on thin columns. The apogee is reached in the large Apsidocrinus with particularly high interradial processes that could shelter longer arms. The earliest stages of E. cariophilites have small articular facets on top of radial ribs, and the muscle fossae are close to the margin (Fig. 4a1); this indicates that the primibrachials, whatever their shape, were too narrow to seal the radial cavity. The distinctive, arrowhead-shaped primibrachials are a synapomorphy of a slightly later, but still juvenile stage (Fig. 6).
Cripples
Among the 470 cups nearly 10 % are mutilated in one way or another. The majority of such cups belong to the thickset variety; juvenile specimens with cup diameters below 1 mm are hardly ever crippled. However, in a small cup one of the articular facets is poorly developed (Fig. 6d, facet at lower left), and in the small cup in Fig. 12b the two upper lobes are swollen and carry no articular facet. Such swollen ossicles that lack a pit may occur in deformed columns (see Hess 2013, fig. 7f, for a specimen of Balanocrinus brachiospina) but are rare in cups; the causing agent is unknown. A number of cups have bite marks (Figs. 11, 12, 13), mostly causing deformation and/or skeletal overgrowth in neighbouring areas. In the specimen in Fig. 13a three well-defined bite marks occur on opposite sides of the shaft, and this suggest that the predator, presumably a fish, grasped the cup with its teeth.

Pathological and mutilated cups of Eugeniacrinites cariophilites. a Cup with bite marks on lower part, facet in front smaller than the others, radials granulated adorally along sutures, M11471. b Lateral-distal view of partly mutilated cup with bite marks in lower part, facets in front and to the left small, facet to the right partly damaged, facets at the back intact; M11473. c Mutilated cup with one radial articular facet intact and one damaged, the other three bitten off; c1 lateral, c2 distal, the partly damaged facet is at left; M11475. d Cup mutilated in upper part, without articular facets, M11474. e Mutilated cup with most of the upper part bitten off, an intact facet is in the back, the facet at left has only the muscle fossa near the interradial process intact, the other is obliterated; no bite marks on shaft; M11476. Scale bar 1 mm

Pathological cups of Eugeniacrinites cariophilites. a Distal view of cup with one intact articular facet (upper right), adjoining facets only half intact; the three facets to the left are obliterated, and the shaft has bite marks on the left side; M11477. b Distal view of prejuvenile cup with three intact facets and two swollen radial extensions without facet, shaft corresponds to specimens in Figs. 4g and 5 and is intact; M11478. c Distal view of cup with three large and two small articular facets that are underlain by irregular growth of the rim, M11479. d Lateral-distal view of cup with intact articular facets, shaft shows bite marks and twisted radial suture, M11480. e Distal-lateral view of cup, the two small articular facets at left are above a part of the shaft showing bite marks; the two adjoining facets are damaged, and only the facet between the processes at upper right is intact; M11481. f Distal view of cup with one intact articular facet; the four small but otherwise regular facets are on part of the shaft with deep bite marks; M11482. Scale bar 1 mm

Pathological cups of Eugeniacrinites cariophilites. a Asymmetrical cup with three more or less intact articular facets, two facets are not developed, shaft with bite marks; a1, 2 two lateral-distal views, asterisk indicates position in distal view (a3), arrows indicate three bite marks also illustrated in a4; a3 distal view; a4 lateral view with bite marks (arrows); M11483. b Tetramerous cup with regular facets, the shaft has bite marks and skeletal overgrowth; b1 lateral, b2 distal; M11484. Scale bar 1 mm
No crippled first primibrachial has been found. Second primibrachials are more variable in shape. Variability includes overall shape and length of the spearhead-shaped median prolongation, but also the angle between prolongation and base with the proximal facet (Fig. 10g, h, e). Not surprisingly, crippled ossicles occur, but their number is much lower compared to cups. A dozen second primibrachials lack the spearhead-shaped median prolongation (Fig. 10a, c). In the ossicle in Fig. 10c this may be due to sublethal predation as indicated by bite marks on the adoral side, yet largely intact facets for the arms. In the specimen in Fig. 10a only two weak lesions are seen on the adoral side, and the distal edge is concave but intact. An unfigured ossicle has one half with its articular facet bitten off while the other half is intact; because the edge is sealed by skeletal growth this seems to be another case of sublethal predation. In ten ossicles the prolongation is widened at the top (Fig. 10b), or bifurcates (Fig. 10d). Some lesions in these ossicles indicate attack by a predator and subsequent healing. Two ossicles are strongly deformed. The second primibrachial in Fig. 10f is hardly recognizable as such; it is prolonged to one side, with a wide muscular facet at the end. This ossicle is still connected to a small first primibrachial (arrow). Crippling may have been caused by lethal attack in this case. Finally, two isolated median prolongations without any indication of repair suggest that they were bitten off and discarded by the predator.
Discussion
The rather large number of deformed or crippled cups in comparison with columnals and second primibrachials suggests that cups with their soft parts were the preferred target for attack by predators. Among cyrtocrinids E. cariophilites has one of the most elaborate morphologies to protect the soft parts in the radial cavity, and this is developed early in ontogeny. Despite such protection many attacks seem to have been successful, although not necessarily lethal. This is in stark contrast to the Bathonian Cyrtocrinus praenutans Hess (2012) for which bite marks are unknown; in contrast, 3.7 percent of the cups have pits caused by an unknown epizoan. Such pits, assigned to Oichnus paraboloides Bromley 1981, are commonly accompanied by increased skeletal growth causing swelling, but this did not lead to the animal’s death (see Hess 2012). Pits of this type are not observed in the present material, although two cases have been figured by Quenstedt (1876, pl. 105, fig. 73 and 91), both from the Oxfordian of Germany; the pit extends from the topmost columnal to the cup. The percentage of deformed second primibrachials is much lower, and this suggests that they did not offer great resistance to an attack. The majority of crippled cups have bite marks on the shaft, with part of the articular facets intact or reduced in size. Such cases indicate sublethal attacks and subsequent regeneration of parts of the cup, although the regenerated parts are reduced in size. Cups with their upper part completely or largely bitten off appear to have been victims of lethal attacks that precluded subsequent regeneration. Among the 36 cups of Pilocrinus moussoni three are deformed and have bite marks, and this is a similar percentage as in E. cariophilites. None of the brachials and columnals is pitted in P. moussoni. Thus, a high degree of predator attacks and a low degree of epizoic infestation is common to both species.
Enigmatic wing plates
The material includes 136 brachials of a type never described before nor seen by the author. The brachials have a spectacular wing that extends from one side of the food groove and is attached to a thicker “head” part bearing the articular facets. The presumed proximal facets are mostly synostosial, flat or weakly concave; because they slant inwards in most cases they are exposed in the adoral view of the ossicles. The largest ossicle (not figured) has a width of 3.3 mm with a synostosial proximal facet. Some small specimens (Fig. 14d, h, i) have a muscular proximal facet, but in contrast to the distal facets with their strong relief and large, deep muscle fossae the relief is flat and the muscle fossae are small, with a diameter matching that of the axial canal. The distal facets are sunken and muscular. The deep muscle fossae may differ in size, and they are separated by a ridge that is connected to the transverse ridge. The transverse ridge is pierced by the axial canal, and it may widen outwards for indistinct interarticular ligaments. The aboral ligament fossae are quite variable, depending on the shape of the head part, and they have a deep pit near the transverse ridge. As demonstrated in Fig. 14 shape and profile of wing part and thicker head part are quite variable. The thick part is mostly rounded but may be triangular (Fig. 14d1) or even spine-like (Fig. 14a). The exterior of the wings is invariably smooth. However, in one case the thick part has a rope-like sculpture on the surface (Fig. 14j). The interior side has food grooves along the head part that extend from a small notch proximally at the synostosial facet to the embayed distal facet. From this paraxial groove a branch extends at a right angle to the rim, terminating in a notch of variable depth. This transverse groove is faint and mostly open, but it may be enclosed in part (Fig. 14b, g). In one case the groove branches a second time, so that the rim has three lappets (Fig. 14h)

Winged cyrtocrinid brachials tentatively assigned to Eugeniacrinites cariophilites; the brachials are figured with the presumed proximal, mostly synostosial facet below and the distal muscular facet directed upwards. a Brachial with spine-like process, M11485; a1 distal facet; a2 adoral-proximal view. b Adoral view of brachial with pit below muscle fossae that continues in hollow, mostly closed tube to the margin; M11486. c Brachial with wide wing, M11487; c1 distal with muscular facet; c2 adoral-proximal with synostosial facet. d Brachial, M11488; d1 distal; d2 adoral, proximal facet is weakly muscular. e Brachial, M11489; e1 distal; e2 adoral with synostosial facet. f Brachial, M11490; f1 distal, f2 adoral-proximal with synostosial facet. g Adoral view of brachial with synostosial proximal facet, M11491. h Adoral view of brachial with weakly muscular proximal facet and branched food groove, M11493. i Adoral view of brachial with weakly muscular proximal facet, M11492. j Aboral-proximal view of brachial with knobby thicker part, M11494. Scale bar 1 mm
The winged brachials are tentatively interpreted as second, non-axillary primibrachials of E. cariophilites with a single facet for an arm. In larger ossicles the presumed proximal facet is synostosial and may thus be combined with the distal facet of the first primibrachial. However, these ossicles are wide and low and have symmetrical synostosial facets in most cases while the corresponding facets of the wing plates tend to be asymmetrical (Fig. 14a2, c2). The distal facets with their deep muscle fossae resemble those of the axillary second primibrachials (Figs. 9, 10). A few small wing plates have flat, indistinct muscular facets proximally (Fig. 14d2, h, i), and these may have articulated directly to the cup, similar to the small axillary primibrachials with muscular proximal facet in Fig. 8. Possible attachment sites for the wing plates are pathological cups in which the number of facets is reduced or the interradial processes poorly developed, leaving sufficient space for the wings. Such cups may also have borne asymmetrical first primibrachials.
Discussion
The purpose of the plates is open to discussion. To some extent they resemble the conspicuous wing plates of the camerate Pterotocrinus from the Mississippian; these plates are hypertrophied tegminal ambulacrals and may develop into an extraordinary variety of forms (Ubaghs 1978). Meyer and Ausich (1983) suggested that they provided shelter to the arms. In contrast, Baumiller and Plotnick (1989) assumed that they rather served as stabilizing rudders, allowing the passive maintenance of an efficient feeding posture in moving water. Such a function is obviously out of question in the present case, as demonstrated by the muscular articular facet for the attachment of an arm. One might hypothesize that the wings on the plates were an antipredatory substitute for the spearhead-shaped second primibrachials on undeformed cups.
Antipredator morphology and cyrtocrinid evolution
The cyrtocrinids developed various antipredator morphologies. For these mostly small stalked crinoids occupying a low tier protection of the soft parts within the cup was essential. During evolution protection was achieved through several strategies. (1) In the Phyllocrinidae the radial articular facets are sunken between interradial processes that shelter small, seemingly unbranched arms. In the more evolved Apsidocrinus the processes are triangular in section to make room for the adpressed arms. Protective clefts for the arms is a common defensive feature (Meyer and Ausich 1983). (2) In the Eugeniacrinitidae the cups also developed interradial processes between sunken articular facets during early growth. However, protection of the soft tissues in cup and arms was assured by the primibrachials that increase in size and acquire their distinctive shape. The arms remained small and hidden from predators under the closed roof formed by the second primibrachials in Eugeniacrinites, or between the spear-shaped prolongations of these ossicles in Lonchocrinus; in both forms the cups are similar, and the column is rather long. (3) The Sclerocrinidae is the largest family, and it includes the living Neogymnocrinus known from many specimens. The cup is compact, the radials mostly lack interradial processes, and the radial cavity is shallow. The crown is well developed, the brachials are robust and the arms branch at least once at the second primibrachials, and more often in Neogymnocrinus. Protection of food grooves and viscera is achieved by inward-coiling of the arms; in addition proximal brachials are adorally united enclosing the food groove. Neogymnocrinus has first primibrachials with closely fitting extensions that seal the adoral side of the cup; this is reminiscent of the juvenile cups of E. cariophilites in Fig. 6. Variability, asymmetry and fusion of ossicles are common in this group; these are truly the “crooked crinoids” from which the order derives its name. The family includes the fossil Cyrtocrinus, Hemicrinus, Pilocrinus, Sclerocrinus, Strambergocrinus, and Torynocrinus; and it reached its apogee in the Lower Cretaceous. The stalkless Holopodina with Holopus, Cyathidium, Cotylederma, Eudesicrinus, and Hemibrachiocrinus adopted a similar strategy, as a result of their shorter arms and lack of a column they resemble barnacles rather than an arm topped by a fist as seen in Neogymnocrinus. (4) The Plicatocrinoidea include forms with spike-like pinnules, beautifully displayed in Plicatocrinus fraasi von Zittel 1882 (see Hess and Messing 2011, fig. 95, 1f). Unfortunately, such spinosity has to be inferred in other forms of the group because most of these small crinoids are only known by their cups and columnals.
To what extent does the mode of protection mirror phylogeny? The earliest cyrtocrinids are Sinemurian to Pliensbachian in age. Prominent representatives belong to cotyledermatids, eudesicrinids, tetracrinids, quenstedticrinids, and phyllocrinids (Hess 2006). The first two groups have cups attached directly to the substrate and are barnacle-like; such a strategy is also found in Cyathidium (Tithonian-Recent) and the extant Holopus. The tetracrinids are mostly sturdy, some having crowns with dagger-shaped pinnules, but most are insufficiently known. The same is true of the quenstedticrinids. The phyllocrinids are thought to be ancestral to the eugeniacrinitids, as discussed previously. Could they be ancestral to the sclerocrinids as well? The cup of Nerocrinus has three basals, but the radial circlet is suggestive of later phyllocrinids. The cup of Ticinocrinus has five basals; the radial circlet is composed of high ossicles surrounding a rather narrow, cylindrical radial cavity, and this feature makes the form unsuitable as ancestor to sclerocrinids. No satisfactory explanation of the origin of the Sclerocrinidae has been given so far. The first member is Cyrtocrinus praenutans Hess 2012 from the Middle Jurassic; it is known from numerous cups, brachials, columnals and attachment discs. The material includes many juvenile cups that resemble the adult ones in overall morphology. In fact, characters of this early cyrtocrinid may be found in the extant Neogymnocrinus (see Bourseau et al. 1991; Hess and Messing 2011). It seems unlikely that sclerocrinids derived from phyllocrinids with their sunken radial facets and small arms. In contrast, some Early Jurassic tetracrinids, such as Bilecicrinus, have sturdy cups, arms and columnals (see Hess 2006; Hess and Messing 2011) and are promising candidates. The holotype of Bilecicrinus arenosus Hess 2006 is an asymmetrical cup of five radials with a small radial cavity; cups of four radials also occur, and in such cases one of the radials has two facets. The proximal side of the radial circlet is synostosial; and it was probably connected to a wedge-shaped basal element. The lower facet of this element and the intercolumnal facets are symplectial. The two primibrachials are united by synostosis, and further arm articulations are muscular. The muscle fossae are small and deep; the pinnule sockets are pronounced and at a right angle to the distal face. Accordingly, the pinnules could be stretched out and seem to have been rather long. The secundibrachials are wedge-shaped so that the arms assumed a zigzag appearance. Incidentally, such an appearance is also shown by the reconstructed arm of a plicatocrinoid figured by Jaekel (1892, fig. 2). The surface of all ossicle types is densely granulated. The large number of secundibrachials found together with cup plates and columnals suggests that the arms were relatively long. The form was assigned to Tetracrinidae on the basis of characters of the cup, but the similarities with Sclerocrinidae such as Cyrtocrinus are obvious. We may thus assume that Sclerocrinidae were derived from primitive Tetracrinidae. Arm structure of Bilecicrinus arenosus indicates that the arms did not coil inwards for defence but rather used spiked pinnules to ward off predators. Asymmetry is weakly developed in Bilecicrinus and C. praenutans. In contrast, Late Jurassic cyrtocrinids commonly are very asymmetrical; and this is also true of the extant Neogymnocrinus richeri. Although asymmetry of cup and crown probably was induced by unilateral currents it may also have added to protection, with the shorter arms and the disc partly hidden by the longer arms coiled inwards (see Bourseau et al. 1991, pls. 8–9).
The most likely predators of Recent crinoids are lobsters, but especially fishes (Meyer and Ausich 1983). Predation pressure is low on living crinoids such as comatulids, and this seems to be due in part to their distastefulness or toxicity but also to the development of rigid and spike-like proximal pinnules that form a palisade over the disc (Meyer and Ausich 1983). In addition, comatulids may escape predators by swimming away or hiding. Stalked isocrinids have been observed to crawl away from cidaroid echinoids when attacked (Baumiller et al. 2008). The nature of the bite marks on the cups of E. cariophilites excludes echinoids as possible predators. However, such attacks are seen in cups and columnals of Apiocrinites roissyanus d’Orbigny 1841 from the Middle Oxfordian of Raedersdorf (Haut-Rhin, France). The remains show superficial scratch marks apparently caused by Pelanechinus corallinus (Wright 1858) from the same outcrop; this sea-urchin has particularly strong teeth that match the marks (Hess 1984, 2014). The few surviving cyrtocrinids are restricted to deeper water (Hess and Messing 2011); Cyathidium has been found by submersibles even on cave ceilings (Heinzeller et al. 1996) or concealed underneath bedrock overhangs (Wisshak et al. 2009). No case of a predator attack on a living cyrtocrinid has been recorded so far. Predation pressure by lobsters and fishes presumably forced the cyrtocrinids, as all the other stalked crinoids, into deeper water during the Cretaceous (Meyer and Ausich 1983). The significant number of crippled cups described here may be an indication that pressure started in the Late Jurassic.
Conclusions
The described material is remarkable for the presence of all growth stages and of deformed cups and primibrachials in E. cariophilites, but also for peculiar wing plates that seem to belong to this species. The excellent preservation may be ascribed to the vicinity of sponge-algal mounds that furnished the hardgrounds on which the cyrtocrinids thrived and from which their remains were washed into the adjacent basin where also the remains of the pelagic saccocomids accumulated. The number of cups deformed by predators is unusually high. The mostly sublethal nature of the damage is inferred by regeneration in various ways to restore at least some of the animal’s functions. Restoration may have been supported by wing plates that carried an arm but also shielded the cup. Predation pressure is thought to have accounted for the survival of stalked crinoids only in waters deeper than about 100 m after the Mesozoic (Meyer and Ausich 1983). Interestingly, such migration also includes the cyrtocrinids despite their elaborate antipredatory measures. Most of the antipredatory morphologies evolved during the first cyrtocrinid radiation in the Pliensbachian. Barnacle-like, compact forms are represented today by the rather common Cyathidium and Holopus with several species; the rare Proeudesicrinus Améziane-Cominardi and Bourseau (in Améziane-Cominardi et al. 1990) is similar to the Early Jurassic Eudesicrinus, making it a true living fossil. Most conspicuous among the survivors is Neogymnocrinus richeri (Bourseau et al. 1987) that combines the protective features of arm coiling, proximal brachials with adorally enclosed food grooves and closely fitting first primibrachials sealing the radial cavity. Despite some progress in the evaluation of cyrtocrinid phylogeny presented here the possibly polyphyletic origin of this fascinating group of crinoids remains in the dark and can probably only be resolved by finds from the Upper Triassic.
References
Améziane, N., Bourseau, J.-P., Heinzeller, T., & Roux, M. (1999). Les genres Cyathidium et Holopus au sein des Cyrtocrinida (Crinoidea; Echinodermata). Journal of Natural History, 33, 439–470.
Améziane-Cominardi, N., Bourseau, J-P., Avocat, R., & Roux, M. (1990). Les Crinoïdes pédonculés de Nouvelle-Calédonie: inventaire et réflexions sur les taxons archaïques. In C. de Ridder, P. Dubois, M-C. Lahaye M. Jangoux (Eds.), Echinoderm Research, Proceedings Second European Conference Echinoderms Brussels/Belgium, 18–21 September 1989 (pp. 117–124). Rotterdam: Balkema.
Arendt, Y.A. (1974). The Sea lilies Cyrtocrinids. Academy of Sciences of the USSR, Transactions of the Palaeontological Institute, 144, p. 251 [in Russian].
Baumiller, T. K., & Plotnick, R. E. (1989). Rotational stability in stalked crinoids and the function of wing plates in Pterotocrinus depressus. Lethaia, 22, 317–326.
Baumiller, T. K., Mooi, R., & Messing, C. G. (2008). Urchins in the meadow: paleobiological and evolutionary implications of cidarid predation on crinoids. Paleobiology, 14, 22–34.
Bourseau, J-P., Améziane-Cominardi, N., & Roux, M. (1987). Un crinoïde pédonculé nouveau (Echinodermes), représentant actuel de la famille jurassique des Hemicrinidae: Gymnocrinus richeri nov. sp. des fonds bathyaux de Nouvelle-Calédonie (S.W. Pacifique). Comptes Rendus de l’Académie des Sciences Paris, 305, série. III, pp. 595–599.
Bourseau, J-P., Améziane-Cominardi, N., Avocat R., & Roux M. (1991). Echinodermata: Les Crinoïdes pédonculés de Nouvelle-Calédonie. In A. Crosnier (Ed.), Résultats des Campagnes MUSORSTOM 8. Mémoires du Muséum national d’Histoire naturelle (A) 151, pp. 229–333.
Bromley, R. G. (1981). Concepts in ichnotaxonomy illustrated by small round holes in shells. Acta Geológica Hispánica, 16, 55–64.
Goldfuss, G. A. (1826–44). Petrefacta Germaniae, tam ea, Quae in Museo Universitatis Regiae Borussicae Fridericiae Wilhelmiae Rhenanae, serventur, quam alia quaecunque in Museis Hoeninghusiano Muensteriano aliisque, extant, iconibus et descriptiones illustrata.-Abbildungen und Beschreibungen der Petrefacten Deutschlands und der Angränzenden Länder, unter Mitwirkung des Herrn Grafen Georg zu Münster, herausgegeben von August Goldfuss. 1 (1826–33), Divisio prima. Zoophytorum reliquiae, 1–114; Divisio secunda. Radiariorum reliquiae, pp. 115–221 [Echinodermata]; Divisio tertia. Annulatorium reliquiae, pp. 222–242. Arnz & Co.: Düsseldorf.
Heinzeller, T., Fricke, H., Bourseau, J.-P., Améziane-Cominardi, N., & Welsch, U. (1996). Cyathidium plantei sp. n., an extant cyrtocrinid (Echinodermata, Crinoidea)—morphologically identical to the fossil Cyathidium depressum (Cretaceous, Cenomanian). Zoologica Scripta, 25, 77–84.
Hess, H. (1975). Die fossilen Echinodermen des Schweizer Juras. Veröffentlichungen aus dem Naturhistorischen Museum Basel, 8, p. 130.
Hess, H. (1984). Ein seltener Seeigel aus dem Malm des Juras. Schweizer Strahler, 6(12), 505–510.
Hess, H. (2002). Remains of Saccocomids (Crinoidea: Echinodermata) from the Upper Jurassic of southern Germany. Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und Paläontologie), 329, p. 57.
Hess, H. (2006). Crinoids from the Lower Jurassic (Pliensbachian) of Arzo, southern Switzerland. Schweizerische Paläontologische Abhandlungen, 126, p. 143.
Hess, H. (2012). Crinoids from the Middle Jurassic (Bajocian—Lower Callovian) of Ardèche, France. Swiss Journal of Palaeontology, 131, 211–253.
Hess, H. (2013). Balanocrinus and other crinoids from Late Jurassic mudstones of France and Switzerland. Swiss Journal of Palaeontology,. doi:10.1007/s13358-013-0059-x.
Hess, H. (2014). Seeigel auf der Jagd nach Seelilien. Fossilien, 31, Heft 2, 17–19. Quelle & Meyer: Wiebelsheim (in press).
Hess, H., & Messing, C.G. (2011). Treatise on Invertebrate Paleontology, Part T, Echinodermata 2, Revised, Crinoidea Volume 3 In W. I. Ausich (Ed.), p. 261. The University of Kansas Paleontological Institute: Lawrence, Kansas.
Jaekel, O. (1891). Ueber Holopocriniden mit besonderer Berücksichtigung der Stramberger Formen. Zeitschrift der Deutschen geologischen Gesellschaft, 43, 557–670.
Jaekel, O. (1892). Ueber Plicatocriniden, Hyocrinus und Saccocoma. Zeitschrift der Deutschen geologischen Gesellschaft, 44, 619–696.
Jaekel, O. (1907). Ueber die Körperform der Holopocriniten. Neues Jahrbuch für Mineralogie, Geologie, Paläontologie, Festband, 1907, 272–309.
Knorr, G.W. (1755). Sammlung von Merkwürdigkeiten der Natur und Alterthümern des Erdbodens: welcher petrificirte Körper enthält. p. 187+36. A. Bieling: Nürnberg.
Knorr, G.W., & Walch, J.E.E. (1768–73). Sammlung von Merkwürdigkeiten der Natur und Alterthümern des Erdbodens, 2 (1: 1768) (2: 1769), p. 303. 3 (1771), p. 235.; 4 (1773), p. 130. P. J. Felssecker: Nürnberg. [vol. 1, see Knorr, 1755].
Lhwyd, E. [Luidius, Eduardus] (1699). Lithophylacii britannici Ichnographia. p. 139. Ex officina M. C.: London.
Loriol, P. de (1877–1879). Monographie des crinoïdes fossiles de la Suisse. Mémoires de la Société Paléontologique Suisse, 4, 1–52; 5, 3–124; 6, 125–300.
Loriol, P. de (1882–1889). Paléontologie française, Série 1, Animaux invertébrés. Terrain jurassique, vol. 11, Crinoïdes, pt. 1 (1882–84), p. 627.; pt. 2 (1884–89), p. 580. G. Masson: Paris.
Manni, R., & Nicosia, U. (1999). The oldest cyrtocrinid, Nerocrinus petri n. gen. n. sp., and the Nerocrinidae, n. fam. In M. D. Candia Carnevali, F. Bonasero (Eds.), Echinoderm Research 1998, (pp. 327–331). Balkema: Rotterdam.
Manni, R., & Nicosia, U. (2004). The ontogeny sequence of Gammarocrinites bakonicus Manni, Nicosia & Szabό 1992. In Heinzeller, Nebelsieck (Eds.) Echinoderms München, Proceedings of the 11th International Echinoderm Conference, 6–10 October 2003, Munich, Germany, (pp. 467–470). Francis & Taylor Group: London.
Manni, R., Nicosia, U., & Szabό, J. (1992). Late Jurassic crinoids from the Eperkés-hegy (Bakony Mts., Hungary). Fragmenta Mineralogica et Palaeontologica, 15, 115–137.
Meyer, D. L., & Ausich, W. I. (1983). Biotic Interactions among Recent and among Fossil Crinoids. In M. J. S. Tevesz & P. L. McCall (Eds.), Biotic Interactions in Recent and Fossil Benthic Communities (pp. 277–427). New York: Plenum.
Orbigny, A. d’ (1840–41). Histoire naturelle, générale et particulière, des Crinoïdes, vivants et fossiles, comprenant la description géologique et zoologique de ces animaux. Livr. 1, 1–32 (1840); livr. 2–3, 33–98 (1841) (republished 1858). Published by the author: Paris.
Parkinson, J. (1808). Organic remains of a former world. An examination of the mineralized remains of the vegetables and animals of the antediluvian world; generally termed extraneous fossils. The second volume containing the fossil zoophytes, pp. 18 and 228. London. (Letter 22, p. 225, The Clove Encrinite; pl. 13, fig. 78).
Pictet, E. J., & Loriol, P. de (1858). Description des fossiles dans le Terrain néocomien des Voirons. Matériaux pour la Paléontologie suisse, sér. 2, p. 64.
Pisera, A., & Dzik, J. (1979). Tithonian crinoids from Rogoznik (Pieniny Klippen Belt, Poland) and their evolutionary relationships. Eclogae Geologicae Helvetiae, 72, 805–849.
Quenstedt, F. A. (1876). Petrefactenkunde Deutschlands. 1. Abt., 4. Band, Echinodermen (Asteriden und Encriniden), viii+742 pp. Fues’s Verlag: Leipzig.
Rasmussen, H.W. (1961). A Monograph on the Cretaceous Crinoidea. Kongelige Danske Videnskaberne Selskab, Biologiske Skrifter 12 (1), p. 428.
Rasmussen, H.W. (1978). Articulata. In R. C. Moore, & C. Teichert (Eds.), Treatise on Invertebrate Paleontology. Pt. T, Echinodermata 2, vol. 3, (pp. T813–T928), The Geological Society of America and The University of Kansas: Boulder, Colorado and Lawrence, Kansas.
Remeš, M. (1912). Nové zprávy o lilijiciích z moravského tithonu. Časopis Moravského Zemského Muzeum v Brně, 12, (pp. 157-169), Brno.
Rosinus, M.R. (1718). Tentaminis de lithozois ac lithophytis olim marinis jam vero subterraneis, Prodomus sive, de Stellis Marinis quondam nunc fossilibus, p. 92. N. Sauer: Hamburg.
Scheuchzer, J.J. (1752). Natur-Historie des Schwizerlandes, second edition, 3, p. 336. Heydegger: Zürich.
Schlotheim, E.F. von (1813). Beiträge zur Naturgeschichte der Versteinerungen. Leonard Taschenbuch für die gesammte Mineralogie, Jahrgang 7(1), (pp. 3–134). Frankfurt.
Ubaghs, G. (1953). Classe des Crinoïdes. In J. Piveteau (Ed.), Traité de Paléontologie, 3 (pp. 658–773). Paris: Masson.
Ubaghs, G. (1978). Skeletal morphology of fossil crinoids. In R. C. Moore & C. Teichert (Eds.), Treatise on Invertebrate Paleontology, Part T, Echinodermata 2 (Vol. 1, pp. T58–T216). Boulder: The Geological Society of America and The University of Kansas.
Wagner, J.J. (1684). Lapilli albi Caryophyllos aromaticos referentes. Miscellania curiosa sive ephemeridum medico-physicarum Germanicarum Academiae Caesareo-Leopoldinae Naturae Curiosorum dec. 2, anno 3, (pp. 370–372). Nürnberg.
Walch, J.E.I. (1762). Das Steinreich. p. 172. J.J. Gebauer: Halle.
Wisshak, M., Neumann, C., Jakobsen, J., & Freiwald, A. (2009). The ‘living-fossil’ community of the cyrtocrinid Cyathidium foresti and the deep-sea oyster Neopycnodonte zibrowii (Azores Archipelago). Palaeogeography, Palaeoclimatology, Palaeoecology, 271, 77–83.
Wright, T. (1857–78). Monograph of the British Fossil Echinodermata of the Oolitic Formations, vol. 1, The Echinoidea. Palaeontographical Society London, p. 468.
Žítt, J. (1974). Sclerocrinus Jaekel, 1891 and Proholopus Jaekel, 1907 (Crinoidea, Cyrtocrinida) from the Lower Cretaceous of Štramberk (Czechoslovakia). Sbornik geologickŷ věd, paleontologie, 16, 7–34.
Žítt, J. (1978). Phyllocrinus d’Orbigny, 1850 (Crinodea, Cyrtocrinida) from the Lower Cretaceous of Štramberk (Czechoslovakia). Časopis pro mineralogii a geologii, roč, 23(1), 39–51.
Zittel, K.A. von (1882). Über Plicatocrinus. Sitzungsberichte der mathematisch-physikalischen Klasse der Akademie der Wissenschaften München, 12, pp. 105–113.
Acknowledgments
I am most grateful to Andy Gale for the material described in the present paper and the photograph of the location. Günter Schweigert, Ben Thuy and Manfred Jäger provided data on the stratigraphy of the location. I wish to thank John W. M. Jagt for his thorough review of the manuscript. I am greatly indebted to Evi Bieler, Zentrum für Mikroskopie of the University of Basel, for the SEM photographs.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Hess, H. The crinoid Eugeniacrinites cariophilites from the Late Jurassic of southern Germany: babies, cripples and enigmatic wing plates. Swiss J Palaeontol 133, 121–140 (2014). https://doi.org/10.1007/s13358-014-0064-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-014-0064-8