
Abstract
Ariidae catfish are described from the early Miocene Pirabas Formation in northeastern Brazil, Cantaure and Castillo formations in northwestern Venezuela, and Castilletes Formation in North Colombia. A skull from the Pirabas Formation is described as Cathorops goeldii, n. sp. and an otolith is identified as Cathorops sp. The otoliths from Cantaure, Castillo and Castilletes formations are described as Cantarius nolfi n. sp. and Bagre protocaribbeanus n. sp. These coeval lithostratigraphic units reflect the Proto-Caribbean conditions and the shallow water tropical palaeoenvironment during the Miocene, particularly associated with marine coastal lagoon and estuarine influence.
Similar content being viewed by others


Introduction
Ariidae (sea catfish) comprises 30 living genera and 138 species (Eschmeyer 2013; Betancur-R. et al. 2007; Marceniuk and Menezes 2007), including 16 fossil species described from Africa (six), the Americas (three), Asia (one), and Europe (six) ranging in age from the upper Cretacous to the Pliocene (Frizzell 1965; Huddleston and Savoie 1983; Ferraris 2007). This high diversity seems to be the product of a rapid evolution and high diversification. The taxonomy of the fossil species from the Proto-Caribbean region has been difficult and mostly remains in open nomenclature due to fragmentary or poor preservation (Nolf 1976; Nolf and Stringer 1992; Nolf and Aguilera 1998; Aguilera and Rodrigues de Aguilera 1999, 2001, 2004; Landini et al. 2002). Recently, Bogan and Agnolin (2011) and Aguilera and Marceniuk (2012) described new extinct species from South America. Ariid catfishes form a well-diagnosed monophyletic group of primarily marine fishes (Gayet and Meunier 2003; Betancur-R 2009; Marceniuk et al. 2012a, b) and numerous characters preserved in the fossil skull are useful for identification of genera and species (Arratia 2003). In addition, ariids are one of the few siluriform groups that can be recognized by isolated otoliths.
The fossils studied here were collected from the early Miocene Pirabas (Maury 1925), Cantaure (Hunter and Bartok 1974), Castillo (Wheeler 1960), and Castilletes (Renz 1960) formations. These formations belong to the palaeobiogeographic Gatunian Province (Landau et al. 2008; Aguilera et al. 2011), and the corresponding fossil records are the product of evolutionary events in the Proto-Caribbean.
The objective of this paper was to describe a fossil ariid skull from the early Miocene Pirabas Formation (Brazil) and otoliths (lapilli) from early Miocene Pirabas (Brazil), Cantaure, Castillo (Venezuela), and Castilletes (Colombia) formations.
Materials and methods
A single catfish skull of Cathorops goeldi n. sp. was collected from the Pirabas Formation, Atalaia beach (0°35′37″S, 47°18′54.4″W), Salinópolis Prefecture in the northeastern Pará State, Brazil (Figs. 1a, 2a), from the marls. An otolith identified as Cathorops sp. was obtained from 10 kg bulk sample of the same location from the gray laminated mudstone (Fig. 2a).

Location map. a Atalaia beach in the Bragantina region, Pará state, Brazil; b San José de Cocodite in the Paraguaná Peninsula, Falcón state, Venezuela; c Cerro La Cruz, southern flank of the Serranía La Baragua, Lara state, Venezuela; d Cocinetas, Alta Guajira Peninsula, Colombia (map A was modified from Távora et al. 2010; and Aguilera and Páes 2012)

Stratigraphic section. a Upper succession of the Pirabas Formation, Atalaia section (from unpublished data of Afonso Nogueira); b Cantaure Formation, outcrop and artisan well section (from unpublished data of Anthony Coates and modified from Aguilera et al. 2011)
Ten otoliths (lapilli) of Bagre protocaribbeanus n. sp. and 50 otoliths (lapilli) of Cantarius nolfi n. sp. were collected from Cantaure Formation outcrops and from subsurface locations in the San José de Cocodite section from the Cantaure Formation on the Paraguaná Peninsula (11°56′32″S, 70°01′07″W), Falcón State, Venezuela (Figs. 1b, 2b).
Ten otoliths (lapilli) of Bagre protocaribbeanus n. sp. and ten otoliths (lapilli) of Cantarius nolfi n. sp. were collected from the Castillo Formation outcrops in Cerro La Cruz, near La Mesa Town (10°23′N, 70°03′W), Lara State, Venezuela (Figs. 1c, 3a).

Stratigraphic section. a Lower succession of the Castillo Formation, Cerro La Cruz section (modified from Sánchez-Villagra et al. 2001); b lower succession of the Castilletes Formation, Cocinetas (from unpublished data of Carlos Jaramillo)
Eight otoliths (lapilli) of Bagre protocaribbeanus n. sp. and nine otoliths (lapilli) of Cantarius nolfi n. sp. were collected from an outcrop in the east of the Cocinetas Basin, Alta Guajira Peninsula (11°56′44.8″N, 71°21′51.7″W), Colombia (Figs. 1d, 3b).
The bulk samples from Pirabas (Brazil) and Cantaure (Venezuela) formations were washed and sieved using a 2-mm mesh. The specimens from the Castillo and Castilletes formation were collected directly on the surface. The otoliths were studied and illustrated using light stereomicroscopy and a camera lucida. Photographs of otolith were made with a stereomicroscope Leica M205A multifocal. Photos were taken remotely controlled from a computer. Multiple sequential photos were taken in regular field of depth steps of each view. The focused parts of each photograph representing a specific field of depth range were then automatically fused with the multifocal system to produce a continuous field of depth. The resulting photographs from the stereomicroscope were digitally edited to remove the background. The skull collected from the Pirabas Formation associated with the pectoral girdles is embedded in a marly matrix. The consolidated marly matrix surrounding the dorsal and lateral surface of the specimen was removed with an electrical driller and by manual cleaning with fine-tipped dental tools. The ventral surface of the skull was studied by the use of an aquilion tomograph with 64 channels to acquire images of 0.5 mm calibration with KV: 120, mAs: 200, Tilt: 0°. The reconstructions are based on the use of coronal, sagittal and axial slides, volume rendering, and maximal intesity progretion. The comparative material includes dry skeletons and otoliths of extant ariids from the western Atlantic and eastern Pacific. The ariid otolith nomenclature is based on Ohe (2006). The skull nomenclature is based on Mo (1991) except “parieto-supraoccipital” for “supraoccipital” (Arratia and Gayet 1995). The term “nuchal plate” is referring to the fusion of the anterior with the posterior plate (Royero 1987). We use the utricular otolith (= lapilli) is referred to as the “otolith”. The comparative otolith morphology (Figs. 7c–d, 9a–e, 13a–p) and the otolith measurements, features, and remarks are summarized in Table 1. However, the ratio otolith length/thickness in the fossil specimens can be altered by diagenesis or erosion so that the unusual wide range of values needs to be interpreted with caution.
Geological setting
Pirabas Formation (Brazil)
The Pirabas Formation (Maury 1925), early Miocene, extends along the Pará, Maranhão, and Piauí States of northern Brazil, and consists of carbonate rocks of an offshore platform environment (grainstone and consolidated packstone, stratified wackestone packstone and laminated mudstone). In addition, littoral facies (shoreface/foreshore), marginal lagoons, restricted platform environments (gray to olive mudstone and conglomeratic sandstone), and mangrove estuarine lagoons (dark mudstone, massif or laminated) have been recorded (Góes et al. 1990; Rossetti 2001; Rossetti and Góes 2004; Costa 2011). The formation is rich in fossils and was studied for the first time by Ferreira-Penna (1876). The planktonic foraminifera association from the Pirabas Formation (Petri 1957; Fernandes 1984, 1988; Ferreira et al. 1978; Fernandes and Távora 1990; Távora and Fernandes 1999) correlates with the early Miocene N4–N5 global biozones (Blow 1969). The early contributions of White (1887) and Maury (1925) are the basal references in respect to mollusk, bryozoan, and coral research, followed by the contributions of Petri (1954, 1957) on foraminifera, Beurlen (1958a, b) on crustaceans, Santos (1958, 1967) on echinoids, Santos and Travassos (1960) on fishes, and Paula-Couto (1967) on sirenians. Many other references, summarized in Rossetti and Góes (2004) and Távora et al. (2010) provide additional descriptions, records, and/or reviews of different fossil taxa. The Atalaia shale and mudstone section (Fig. 2a) from where the fossils were obtained represents the upper-most segment of this formation, just below the early to middle Miocene Barreira Formation. The palaeoenvironment along the Atalaia section is interpreted as a transition tidal platform, with infaunal and epifaunal invertebrate richness associated with high algal and seagrass productivity (photic zone) on the biocalcirudite deposit, where the mollusk assemblage was fossilized in “live” position. Biocalcilutite, also rich in fossils, overlays the biocalcirudite rock and represents a sublittoral palaeoenvironment that was exposed to waves and tidal influence. Then, gray laminated mudstones occur abruptly over the biocalcilutite, where the tidal platform is in this section replaced by very shallow coastal lagoon sediments associated with a rapid burial rate of siliciclastic deposits from the continental drainage. Marly overlayer the mudstone and the palaeoenvironment can be interpreted as brackish. The green laminated mudstone with thalassinoid bioturbations and leaves indicate an inter-tidal palaeoenvironment, and the black mudstone with leaves and trunks represents a marginal lagoon with mangrove vegetation. The top of the section consists of biocalcirudite rocks deposited in a tidal platform (photic zone), with abundant broken shells and echinoids as a consequence of the marine tropical storm effects in the shallow sublittoral.
Cantaure Formation (Venezuela)
The Cantaure Formation (Hunter and Bartok 1974), early Miocene, can be found approximately 10 km west of Pueblo Nuevo Town on the Paraguaná Peninsula, Falcón State, Venezuela. Outcrops of the formation are exposed west of Casa Cantaure and consist of silty shales interbedded with thin algal limestones and shell beds (Hunter and Bartok 1974). There is an unexposed unit of Cantaure Formation that is 48 m thick and can be accessed by a local artisan well (Fig. 2b). The section consists mainly of silty to medium sandstone, intercalated with massive mudstone. The formation has been assigned to the late early Miocene Burdigalian based on planktonic foraminifera and calcareous nannofossil biostratigraphy (Díaz de Gamero 1974; Rey 1996). A diverse fossil fauna, particularly rich in mollusks, decapods, and fishes, has been identified along the Cantaure section (Jung 1965; Thomas and MacDonald 1970; Nolf and Aguilera 1998; Aguilera 2010; Aguilera and Lundberg 2010; Aguilera et al. 2010). These fossils are indicative of a tropical-marine, clear shallow-water coastal environment of normal salinity (Jung 1965; Díaz de Gamero 1974; Nolf and Aguilera 1998; Aguilera and Rodrigues de Aguilera 2001).
Castillo Formation (Venezuela)
The Castillo Formation (Wheeler 1960, 1963) dates as late Oligocene to early Miocene (Lorente 1997) and is situated in the western Falcón Basin, northwestern Lara State, Venezuela. The outcrop of the formation from where the fossils were obtained is exposed at Cerro La Cruz near La Mesa Town, on the southern flank of the Serranía La Baragua, and consists of marls, sandstones, and claystones (Fig. 3a). Wheeler (1960) suggested an early Miocene age for the exposures of the Castillo Formation in the area studied in this paper. The palaeoenvironment is interpreted as near-shore marine and includes diverse assemblages of mollusks, crustaceans, fishes, turtles, crocodiles, and terrestrial, aquatic and marine mammals (Sánchez-Villagra et al. 2000; Sánchez-Villagra and Clack 2004; Johnson et al. 2009).
Castilletes Formation (Colombia)
The Castilletes Formation (Renz 1960; Rollins 1965) outcrops in the Alta Guajira Peninsula (Irving 1972), northern Colombia. Its deposits are characterized by marly limestones, clays, calcareous and non-calcareous sandstones, and conglomerates. Toward the base of the formation, the limestones are coarsely textured, marly, argillaceous, sandy fossiliferous, and fairly indurated (Fig. 3b). The clays are silty, brown to buff, gray, greenish gray, and reddish, with some sandy intercalations (Rollins 1965). Toward the top, sandstones and conglomeratic sandstones are more common, often poorly sorted, and matrix-supported in channel lenses. The unit rests conformably on the Jimol Formation while the upper contact is not exposed. The Castilletes Formation is ~340 m thick in the study area. The unit was deposited in a very shallow marine environment (Rollins 1965) intermixed with continental facies produced by progradation of a fan delta. The Castilletes Formation is rich both in marine and terrestrial fossils including plants, mammals, crocodiles, turtles, bivalves, gastropods, crabs, and fishes. The otoliths were found near the base of the Castilletes Formation (stratigraphic meter 40) in marly limestone beds. Dating of Castilletes Formation is still controversial. Renz (1960) suggests an early Miocene age for the basal sediments based on Miogypsinaantillea (Cushman), Bürgl (1960) estimated middle Miocene age at the unit base and Rollins (1965) proposes a Miocene to early Pliocene age. Based on fish otolith occurrence and the close relationship between teleostean assemblage faunas with Cantaure and Castillo formations we suggest an early Miocene age in agreement with Renz (1960).
Institutional and collection abbreviations DU: Duke University, USA; FURG: Universidade Federal de Rio Grande, Brazil; INVEMAR: Instituto de Investigaciones Marinas de Punta de Betín, Colombia; MPEG: Museu Paraense Emilio Goeldi, Brazil; OPC: Otolith collection-Fumio Ohe, Japan; STRI: Smithsonian Tropical Research Institute, Panamá; UNEFM PR: Universidad Nacional Experimental Francisco de Miranda, Venezuela; USNM: National Museum of Natural History, Smithsonian Institution, USA.
Comparative material examined Amphiarius rugispinis (Valenciennes), UNEFM PR-04, UNEFM PR-073, Orinoco Delta, Atlantic; Aspistor quadriscutis (Valenciennes), UNEFM PR-012, UNEFM PR-019, UNEFM PR-068, Orinoco Delta, Atlantic; Bagre bagre (Linnaeus), UNEFM PR-09, Venezuelan Gulf, Caribbean; B. marinus (Mitchill), UNEFM PR-011, Gulf of Venezuela, Caribbean; B. panamensis (Gill), STRI-5.3.1.15, DU-P1018, Panamá Bay, Pacific; B. pinnimaculatus (Steindachner), STRI-5.3.2.6, Panamá Bay, Pacific; Cathorops agassizii (Eigenmann and Eigenmann), MPEG-1256, off coast of Pará, Brazil; C. aguadulce (Meek), UNEFM PR-020, Términos Lagoon, Mexican rivers; C. arenatus (Valenciennes), MPEG-4665, off the coasts of Pará state, Brazil; C. dasycephalus (Günther), STRI 5.2.1.7, Panamá Bay, Pacific; C. hypophthalmus (Steindachner), STRI-5.4.2.2, Panamá Bay, Pacific; C. spixii (Spix and Agassiz), MPEG-3415, off the coasts of Pará state, Brazil; C. wayuu, (Betancur-R., Acero and Marceniuk), UNEFM PR-010, UNEFM PR-020, Orinoco Delta, Atlantic; Galeichthys afer (Castelnau), OPC0220305-2, Fish River, South Africa; G. feliceps Valenciennes, OPC020305-3, Fish River, South Africa; Genidens barbus (Lacépède), FURG-uncatalog, off the coasts of Rio Grande, Brazil, Atlantic; G. genidens (Cuvier), FURG- uncatalog, off the coasts of Rio Grande, Brazil, Atlantic; G. planifrons (Higuchi, Reis and Araújo), FURG- uncatalog, off the coasts of Rio Grande, Brazil, Atlantic; Notarius cookei (Acero and Betancur-R.), STRI-5.2.10.9, Panamá Bay, Pacific; N. grandicassis (Valenciennes), UNEFM PR-012, Gulf of Venezuela, Caribbean; N. kessleri (Steindachner), STRI-5.2.4.21, UNEFM PR-081, Panamá Bay, Pacific; N. lentiginosus (Eigenmann and Eigenmann), STRI-5.2.12.1, Panamá Bay, Pacific; N. osculus (Jordan and Gilbert), STRI-5.5.5.8, Panamá Bay, Pacific; N. planiceps (Steindachner), STRI-5.2.6.8, UNEFM PR-08, Panamá Bay, Pacific; N. troschelli (Gill), STRI-ST, Panamá Bay, Pacific; Occidentarius platypogon (Günther), STRI-5.2.7.3, Panamá Bay, Pacific; Sciades couma (Valenciennes), UNEFM PR-018, Orinoco Delta, Atlantic; S. dowii (Gill), STRI-5.2.7, UNEFM PR-079, Panamá Bay, Pacific; S. felis (Linnaeus), UNEFM PR-013, Términos Lagoon, México; S. guatemalensis (Günther), STRI-5.1.2.2, Panamá Bay, Pacific; S. herzbergii (Bloch), UNEFM PR-017, Orinoco Delta, Atlantic, UNEFM PR-054, Gulf of Venezuela, Caribbean; S. parkeri (Traill), UNEFM PR-070, Orinoco Delta, Atlantic; S. passany (Valenciennes), UNEFM PR-015, UNEFM PR-071, Orinoco Delta, Atlantic; S. proops (Valenciennes), UNEFM PR-014, Orinoco Delta, Atlantic; S. seemani (Günther), STRI-5.1.1.10, DU-1017, Panamá Bay, Pacific.
Systematic palaeontology
Siluriformes sensu Berg 1940
Ariidae Bleeker 1862
Cathorops Jordan and Gilbert 1882
Cathorops goeldii new species

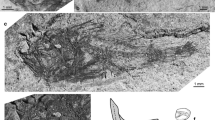
Skull and pectoral girdle of Cathorops goeldii, n. sp., Holotype, MPEG-1526-V, from the Pirabas Formation, Atalaia beach, Brazil. a Oblique posterodorsal view; b left side view; c right side view; d external view of the cleithrum; e internal mold of the cleithrum; f schematic reconstruction in dorsal view; g schematic reconstruction in left lateral view. adp-cl anterior dorsal process of cleithrum, cl cleithrum, pmg posterior medial groove, dspe dorsal spine, ls lateral sulcus, np nuchal plate, pa-soc parieto-supraoccipital, pp-cl posterior process of cleithrum, pt pterotic, spt spinelet, tf temporal fossa. Schematic draw: solid black lines show artificial bones sutures; Unpreserved bones in light gray color. Fossil specimen in dark gray color. Scale bar 10 mm

Tomography sections of Cathorops goeldi n. sp., Holotype, MPEG-1526-V, from the Pirabas Formation, Atalaia beach, Brazil. a–c Progressive coronal sections; d–f progressive sagittal sections; g–l progressive horizontal sections. bo basioccipital, cl cleithrum, dspe dorsal spine, mr müllerian ramus, np nuchal plate, oc otic capsule, pa-soc parieto-supraoccipital, pr prootic, sco scapulocoracoid, svo superficial ventral ossification of the weberian apparatus, spt spinelet, svp subvertebral process, tp transcapular process. Scale bar 10 mm
Holotype
Skull with associated pectoral girdle, MPEV-1526-V.
Locality and geological age
Cathorops goeldii n. sp. is only known from the early Miocene Pirabas Formation, Atalaia beach (0° 35′31.9″S, 47°19′00″W), Salinópolis Prefecture, Pará State, Brazil.
Etymology
The species is named in honor of Emilio Goeldi, Swiss zoologist and early Amazonas natural science researcher after whom the Museu Paraense in Belém, Brazil, has been named.
Diagnosis
The new species is placed in the genus Cathorops based on the combination of the following characters: the skull bones forming a granulated cephalic shield [vs. smooth or grooved in Bagre (except in B. marinus) and Galeichthys, Marceniuk and Menezes 2007, and Fig. 6]; the posterior cranial fontanelle is reduced to a small opening, posteriorly continued in a deep medial groove along the parieto-supraoccipital (vs. without posterior deep medial groove in Amphiarius, Aspistor, Bagre bagre, Galeichthys, Notarius, and Sciades, Marceniuk and Menezes 2007, and Fig. 6); the parieto-supraoccipital bone is prominent; its process is long, almost as narrow at the base as its posterior portion (vs. base distinctly wider than posterior portion in Aspistor and Sciades, almost as narrow as posterior portion in Galeichthys and base conspicuously narrower than posterior portion in Notarius grandicassis, Marceniuk and Menezes 2007, and Fig. 6); the nuchal plate is half-moon shaped (vs. butterfly shaped in Aspistor, shield-like in Sciades, except S. herzbergi and Notarius trosheli, Marceniuk and Menezes 2007, and Fig. 6); the anterior margin of the nuchal plate is convex (vs. concave in Aspistor, Marceniuk and Menezes 2007, and Fig. 6); the posterior cleithral process is short and acute, and the surface coarsely ornamented with reticulating ridges and subcircular pits (vs. laminar and triangular with the tip dorsally oriented in Notarius, slender and acute with the tip dorsally oriented in Bagre; surface smooth in Bagre, striate and granulate in Aspistor, Notarius and Sciades, Marceniuk and Menezes 2007, and Fig. 6); the posterior dorsal cleithral process is not preserved.

Comparative skull and pectoral girdle of living Ariidae. a, b Amphiarius rugispinis (Valenciennes), UNEFM PR-04; c, d Aspistor quadriscutis (Valenciennes), UNEFM PR-012; e, f Bagre marinus (Mitchill), UNEFM PR-011; g, h Cathorops wayuu, (Betancur-R., Acero and Marceniuk), UNEFM PR-010; i, j Genidens planifrons (Higuchi, Reis and Araújo), FURG- uncatalog; k, l Notarius grandicassis (Valenciennes), UNEFM PR-012; m, n Sciades parkeri (Traill), UNEFM PR-070; o C. hypophtalmus (Steindachner), STRI 5.4.2.2; p C. dasycephalus (Günther), STRI 5.2.1.7; q C. multiradiatus (Günther), INVEMAR-PEC 6798; r C. arenatus (Valenciennes), USNM 233482. adp-cl anterior dorsal process of cleithrum, cl cleithrum, dg dorsal groove, dspe dorsal spine, ex extrascapula, fr frontal, me mesethmoid, np nuchal plate, pdp-cl posterior dorsal process of cleithrum, pp-cl posterior process of cleithrum, pt pterotic, pttsc posttemporal-supracleitrum, psoc parieto-supraoccipital, psocp parieto-supraoccipital process, sp sphenotic, spt spinelet. Scale bar 10 mm (scale unavailable in figures q and r) (plate and photos modified from Aguilera and Rodrigues de Aguilera 2004; and Aguilera and Luckenbill, http://catfishbone.acnatsci.org/Ariidae/index.html; photo q courtesy of R. Betancur-R; photo r courtesy of A. Marceniuk)
Cathorops goeldii n. sp. can be differentiated from its extant subcongeners from Central and South Western Atlantic by the combination of the following characters: The cephalic shield is strongly granulate, parallel to the medial groove; the parieto-supraoccipital posterior process is short to medium in size, almost as narrow at its base as at its posterior portion; the medial groove is deep along the parieto-supraoccipital process reaching the level of the temporal fossa; each side of the medial groove has a pair of divergent, short and shallow lateral sulci; both transcapular processes form an arch, which is anteriorly oriented relative to the body axis; the posterior process of the cleithrum is short and acute, its tip posteriorly oriented; the cleithrum has an expanded bulbous external area around the pocket structure to receive the articular head of the pectoral spine.
Description
Neurocranium (Figs. 4a–c, f–g, 5a–i). The bones of the cephalic shield are strongly granulated and parallel to the medial groove. The posterior cranial fontanelle is reduced to a small opening and posteriorly continued in a deep medial groove along the parieto-supraoccipital, reaching the level of the temporal fossa. Each side of the medial groove has a pair of divergent, short, and shallow lateral sulci. The frontals contact the parieto-supraoccipital posteriorly and the sphenotics postero-laterally. The sphenotic is flat with a straight lateral margin and articulates with the parieto-supraoccipital and pterotic. The extrascapular is oval to subquadrangular in shape and is located on the posterodorsal region of the neurocranium and sutured to the parieto-supraoccipital, the pterotic, and the supracleithrum. The parieto-supraoccipital is the largest bone of the skull; its posterior process is short to medium in size, almost as narrow at its base as at its posterior portion; the posterior margin contacts the nuchal plate through a concave-convex articulation. The upper branch of the supracleithrum is flat, subtriangular in shape, and articulates lateroposteriorly with the cephalic shield. The inferior limb of the relatively short and thick supracleithrum, or transcapular process, is cylindrical to columnar in shape. Both transcapular processes are oriented posteriorly and together form an arch anteriorly oriented relative to the body axis. A thin sheet of the Müllerian ramus can be observed just behind the transcapular process. The very large otic capsules are well differentiated and limited by the prootic, pterotic, and exoccipital. The basioccipital is dagger-shaped. It is posteriorly joined with the first vertebra, forming the distinct, long subvertebral process. The superficial ventral ossification of the Weberian apparatus forms a carina.
Dorsal fin (Figs. 4a–c, f–g, 5a–c). The nuchal plate is halfmoon-shaped and coarsely ornamented.
Pectoral girdle (Figs. 4a–e, g, 5j–l). The anterior dorsal process is narrow and acute, the posterior dorsal process is not preserved, and the posterior process of the cleithrum is short and acute. The scapulocoracoid forms a very broad surface with well-developed crests ending in an interdigitized projection sutured with the opposite pectoral girdle. The cleithrum has an expanded bulbous external area around the pocket structure to receive the articular head of pectoral spine. Both scapulocoracoids form an arch, which is anteriorly oriented relative to the body axis. The cleithrum surface is coarsely ornamented with reticulate ridges and subcircular pits.
Measurements The holotype, MPEG-1526-V, preserved skull length: 52.0 mm; preserved skull width: 39.0 mm.
Cathorops sp.
Fig. 7a–b

Otoliths (lapilli) of Cathorops sp., a–b fossil, MPEG-1523-V, from the Pirabas Formation, Atalaia beach, Brazil; c extant Cathorops wayuu Betancur-R., Acero and Marceniuk, UNEFM PR-020, from the Orinoco Delta, Venezuela; d C. spixii (Spix and Agassiz), uncatalog-18 (from A. Marceniuk), off the coasts of São Paulo, Brazil. add anterodistal ditch, amp anterior antero-mesial projection, df dorsal face, dis distal, msd mesial shallow depression, post posterior, vf ventral face. Dorsal and ventral face, lateral view, draw and photo, a–d are the left otolith
Examined specimen
Otolith (lapilli), MPEG-1523-V.
Locality and geological age
Early Miocene Pirabas Formation, Atalaia beach (0°35′31.9″S, 47°19′00″W), Salinópolis Prefecture, Pará State, Brazil.
Description
The otolith is macula-shaped to almost rectangular. The anterior margin is incurved. The dorsal surface is smooth and convex, the ventral surface is concave. The anterodistal ditch is deep, continuing to the anterior margin. The pseudocauda of the mesial shallow depression is expanded posteriorly. No mesial inward curvature is developed. Ratio lapillus length/width: 1.4. Ratio lapillus length/thickness: 2.8.
Cantarius new genus
Fig. 8a–e

Otoliths (lapilli) of Cantarius nolfi n. sp. a Holotype, UNEFM-PF-3223, from the Cantaure Formation, artisan well; b paratypes, UNEFM-PF-3224-1 from the Cantaure Formation, artisan well; c–e paratypes, UNEFM-PF-3225-1 to UNEFM-PF-3225-3, from the Cantaure Formation outcrops. add anterodistal ditch, amp anterior antero-mesial projection, df dorsal face, dis distal, mic mesial inward curve, msd mesial shallow depression, post posterior, vf ventral face. Dorsal and ventral face, lateral view, draw and photo, d and e are the right otolith (lapillus) others are left ones. Scale bar 1 mm
Etymology
The first part of the generic name relates to the Cantaure Formation and the second part “arius” to the genus name Arius.
Type species
Cantarius nolfi n. sp.
Diagnosis
Cantarius n. gen. is distinguished from morphologically similar otoliths of Genidens (Fig. 9a–c), an extant subtropical Atlantic genus (South Brazil, Uruguay and Argentina) and Galeichthys (Fig. 9d–e), an extant subtropical Pacific genus (Peru and South Africa) by the strong convexity of the dorsal and ventral surface (vs. feeble convexity of the dorsal surface in Genidens and Galeichthys); the otolith is ovoid to elongate in shape (vs. subquadrangular in Genidens and Galeichthys), the mesial inward curvature is well developed (vs. absent or poorly developed in Galeichthys and Genidens), and the anterior margin is slightly concave (vs. almost straight in Galeichthys and Genidens).

Otolith (lapilli) of extant a Genidens barbus (Lacépède), FURG-uncatalogued; b G. genidens (Cuvier), FURG-uncatalogued; c G. planifrom (Higuchi, Reis and Araújo), FURG-uncatalogued; d Galeichthys ater Castelnau, OPC020305-2; e G. feliceps Valenciennes, OPC020305-3. Dorsal and ventral face, lateral view, a–d are right otolith, e is the left one
Cantarius nolfi new species

Otolith (lapilli) of Cantarius nolfi n. sp. Non-Type. a–d UNEFM PF-3229-1 to 3229-4, from Castillo Formation, Cerro La Cruz (10°23′N, 30°03′W), Venezuela; e STRI 16555 (1), from the Castilletes Formation, La Guajira Peninsula (11°51′08.9″N, 71°21′53.0″ W), Colombia; f–h STRI 16677 (3) from the Castilletes Formation, La Guajira Peninsula (11°55′34.1″N, 71°21′33.8″W), Colombia, b, d, e, g and h are right otolith, others are the left ones. Scale bar 2 mm
Genidens sp. Nolf and Aguilera 1998, p. 251, pl. 4, figs. 1–4, tab. 3
Arius sp. Monsch 1998, p. 43, pl. 4, figs. 19, 20
Ariidae ind. Monsch 1998, p. 43, pl. 4, fig. 19
Aguilera and Rodrigues de Aguilera 2001, p. 733, fig. 1
Aguilera 2010, p. 115
Etymology
Named in honor of Dirk Nolf, Institut Royal des Sciences Naturelles de Belgique, for contributions to knowledge of fossil teleostean otoliths.
Holotype
Otolith (lapilli), UNEFM PF-3223
Paratypes
Otolith (lapilli), UNEFM PF-3224-1 to UNEFM PF-3224-6, UNEFM PF-3225-1 to UNEFM PF-3225-6, MPEG-1524-V-1 to MPEG-1524-V-6.
Non-type specimens
Otolith (lapilli), UNEFM PF-3229-1 to 3229-4, from Castillo Formation, Cerro La Cruz (10°23′N, 70°03′W), Lara State, Venezuela; STRI 16555 (1) and STRI 16677 (3) from the Castilletes Formation, La Guajira Peninsula, Colombia.
Locality and geological age
Cantarius nolfi n. sp. is known from the early Miocene Cantaure Formation, San José de Cocodite (11°56′32″S, 70°01′07″ W), Paraguaná Peninsula, Falcón State, Northwestern Venezuela. Also from the early Miocene Castillo and Castilletes formations (Colombia and Venezuela, respectively), and middle Miocene Pebas Formation in Colombia and Peru (Monsch 1998).
Diagnosis
Same as genus (monospecific genus).
Description
The otolith is very regularly ovoid and extremely thick. The anterior margin is slightly concave. The caudal margin is rounded. The antero-mesial projection is small and rounded. The umbo is located on the anterior part of the dorsal surface. The dorsal surface is irregular and very convex. The ventral surface is convex, and both convexities result in a globular aspect of the otolith. The pseudocauda of the mesial shallow depression forms an arch and is expanded posteriorly. The mesial inward curvature forms a well-developed wedge shape oriented posteriorly. Ratio lapillus length/width: 1.1–1.5 (n: 28). Ratio lapillus length/thickness: 1.8–2.6 (n: 28).
Bagre Cloquet 1816
Bagre protocaribbeanus new species

Otolith (lapilli) of Bagre protocaribbeanus n. sp. a Holotype, UNEFM-PF-3226. b–e Paratypes, UNEFM-PF-3227-1 to UNEFM-PF-3227-4, from Cantaure Formation, San José de Cocodite, Falcón State, Venezuela. mic mesial inward curve, mn mesial notch. Dorsal and ventral face, lateral view, draw and photo. c and e are the right otolith (lapillus), others are the left ones. Scale bar 2 mm

Otolith (lapilli) of Bagre protocaribbeanus n. sp. Non-Type. a–d UNEFM PF-3228-1 to 3228-4, from the Castillo Formation, Cerro La Cruz (10°23′N, 70°03′W), Venezuela; e STRI 16710, from the Castilletes Formation, La Guajira Peninsula (11°55′34.1″N, 71°21′33.8″W), Colombia; f. STRI 16677 (1), from the Castilletes Formation, La Guajira Peninsula (11°55′34.1″N, 71°21′33.8″W), Colombia; g STRI 16555 (1), from the Castilletes Formation, La Guajira Peninsula (11°51′08.9″ N, 71°21′53.0″W), Colombia; h STRI 16519 (1), from the Castilletes Formation, La Guajira Peninsula (11°51′04.2″N, 71°19′26.6″W), Colombia, b, d and f are right otolith, others are the left ones. Scale bar 1 mm
Arius sp. Nolf and Aguilera 1998, p. 250, pl. 3, figs. 1–5, tab. 3
Arius sp. Aguilera and Rodrigues de Aguilera 2001, p. 733, fig. 1
Holotype
Otolith (lapilli), UNEFM PF-3226
Paratypes
Otolith (lapilli), UNEFM PF-3227-01 to UNEFM PF-3227-09, MPEG-1525-V-01 to MPEG-1525-V-06.
Locality and geological age
Early Miocene Cantaure Formation, San José de Cocodite (11°56′32″S, 70°01′07″W), Paraguaná Peninsula, Falcón State, Northwestern Venezuela.
Non-type specimens
Otolith (lapilli), UNEFM PF-3228-1 to 3228-5, from the Castillo Formation, Cerro La Cruz (10°23′N, 70°03′W), Lara State, Venezuela, and STRI 16519 (1), 16555 (1), 16677 (1), 16710 (1), from the Castilletes Formation, La Guajira Peninsula, Colombia.
Etymology
The species name refers to the Miocene Proto-Caribbean bioprovince.
Diagnosis
Bagre protocaribbeanus n. sp. is distinguished from the extant Bagre species by the long, narrow expansion of the mesial inward curvature and deep mesial notch (vs. reduced in B. bagre, B. marinus and B. panamensis). The otolith is circular (vs. slightly ovoid in B. bagre and B. panamensis).
Description
The broad otolith is clam-shaped with a circular outline, and the anteromesial projection is sharp. The umbo is located on the anterior part of the irregular and strongly convex dorsal surface. The ventral surface is slightly convex. The pseudocauda of the mesial shallow depression forms an arch. The mesial inward curvature forms a long, narrow expansion that is oriented posteriorly. The mesial notch is deep and wide. The ratio lapillus length/width is 1.1–1.3 (number of measured specimens n: 16), and the ratio lapillus length/thickness is 1.3–2.9 (n: 16).
Palaeoenvironment, palaeoceanography and occurrences of fossil ariids
The early Miocene Proto-Caribbean fossil record is a reflection of the drastic macroevolutionary events that occurred in the Neogene as a consequence of the strong palaeoenvironmental changes in the Caribbean, which in turn were a consequence of the closure of the seaway between the Atlantic and Pacific oceans (e.g., Jackson et al. 1993; O′Dea et al. 2007; Johnson et al. 2008; Aguilera et al. 2011; Klaus et al. 2011) during the uplift of the Panamá Isthmus (Coates et al. 2005). The dynamic tectonic of the Caribbean Plate in combination with the North and South American Plates, and the Nazca and Cocos Plates (Bachmann 2001, Pindell et al. 2005) produced island formation and displacement (Greater, Lesser and Leeward Antilleans). As a consequence the littoral and sea palaeoenvironments and the palaeoceanography rapidly changed in the different sedimentary basins in the Proto-Caribbean (Iturralde-Vinet 2004–2005; Iturralde-Vinet and McPhee 1999). The passive margin of Brazil and the coastal platform (Mohriak 2003; Rossetti 2006) were mainly influenced by global eustatic sea-level changes (Carter 1998; Rossetti 2001; Kominz et al. 2008), while the equatorial Atlantic oceanic current (Johns et al. 1998, 2002), and high wave sequence, and tidal influence (Beardsley et al. 1995; Diretoria de Hidrografia e Navegação-DHN 2011) produced dynamic changes in the depositional conditions in the Proto-Caribbean region called Gatunian Province (Landau et al. 2008; Aguilera et al. 2011) during the Miocene to early Pleistocene. In addition, the Andes uplift (Gregory-Wodzicki 2000; Martinod et al. 2010) and particularly the rise of the eastern Andean Cordillera in Colombia isolated the water inflow of the northern drainage system in South America to the Caribbean through the modern Amazonian effluent, resulting in changes of the course of the Proto-Orinoco river (Díaz de Gamero 1996), the establishment of the Amazon drainage to the Atlantic coast (Hoorn et al. 1995, 2010; Figueiredo et al. 2009), and today′s northwestern South American estuarine conditions (e.g., Johnson et al. 2008; Smith et al. 2010). These factors have a strong influence on the development of the coastal marine and estuarine environments which are inhabited by catfishes and influence the biogeographic distribution of these fishes.
In tropical South America, the genus Cathorops is nowadays represented by at least 19 species (Ferraris 2007; Marceniuk and Menezes 2007; Marceniuk 2007; Marceniuk and Betancur-R. 2008; Marceniuk et al. 2012a, b). According to Marceniuk et al. (2012a, b) Cathorops spixii exhibits a wide distribution along the Brazilian coast, from the Amazon delta to Rio de Janeiro State, overlapping only with the distribution of C. agassizii (Amazon delta to near the coast of Sergipe State) and C. arenatus (near the coast of Pará and Maranhão States). The presence of a fossil Cathorops in the Pirabas Formation (Pará State, northern Brazil) and the previous fossil records from Venezuela of the time interval from late Miocene to early Pliocene (Aguilera and Rodrigues de Aguilera 2001; Aguilera 2010; Aguilera et al. 2011), and in middle Miocene strata of Trinidad (Nolf 1976; Aguilera et al. 2011) is associated with a tropical origin. Cathorops goeldii n. sp. from the Pirabas Formation (early Miocene) represents the earliest Cathorops fossil record and possibly predates the New World Cathorops diversification observed today, which is thought to have originated during the early Pliocene (Betancur-R. et al. 2012). Therefore, it is argued here that C. goeldi may be regarded as an offshoot from the main Cathorops stem lineage.
Admittedly, however, the lack of knowledge of otoliths of all extant Cathorops species and the restricted inter-specific diagnostic characters observed in their otolith morphology limits the interpretation of the fossil Cathorops otoliths from the Pirabas Formation (Figs. 7a–d).

Comparative otoliths (lapilli) of living Ariidae. a Aspistor quadriscutis (Valenciennes); b Amphiarius rugispinis (Valenciennes); c Bagre bagre (Linnaeus); d Bagre marinus (Mitchill); e Bagre panamensis (Gill); f Notarius grandicassis (Valenciennes); g Notarius kessleri (Steindachner); h Notarius planiceps (Steindachner); i Sciades dowii (Gill); j Sciades felis (Linnaeus); k Sciades guatemalemsis (Günther); l Sciades herzbergii (Bloch); m Sciades parkeri (Traill); n Sciades passany (Valenciennes); o Sciades proops (Valenciennes); p Sciades seemani (Günther). add anterodistal ditch, amp anterior antero-mesial projection, df dorsal face, dis distal, md mesial dent, mic mesial inward curve, mn mesial notch, msd mesial shallow depression, post posterior, vf ventral face. Left to right size, dorsal and ventral face, lateral view, g, h and i are left otolith, others are the right ones. Scale bar 2 mm
The newly described Cantaurius nolfi shows a wide geographic distribution along the northwestern coast of South America of the Proto-Caribbean region during the early Miocene. The locations and strata, from where C. nolfi was obtained, are characterized by marine shallow water palaeoenvironments. However, some strata were deposited under brackish conditions and environments with estuarine influence (Castillo and Castilletes formations). Cantaurius nolfi also was recorded (as Ariidae indeterminate, Arius sp. and Arius cf. gigas) in Colombia (Los Chorros, Mocagua, Buenos Aires), middle Pebas Formation (Monsch 1998), and in Perú (Santa Julia), Solimões/Pebas formations of northwestern Amazonia in conditions of the so-called “marine incursion” as concluded from the mixture of euryhaline and freshwater taxa (Monsch 1998; Latrubesse et al. 2010). The specimens studied by Monsch (1998) were not accessible for this review, but his illustrations and description support the diagnostic characters of Cantaurius nolfi.
The extant Central and South American ariid species along the Caribbean and the Atlantic coasts are mainly exclusive marine species. However, some species like Cathorops cf. festae (cited by Betancur-R. et al. 2012), Notarius bonillae (Miles), and N. cookie (Acero and Betancur-R.) inhabit freshwater environments and Amphiarius phrygiatus (Valenciennes), Cathorops hypophthalmus (Steindachner), C. tuyra (Meek and Hildebrand), and C. agassizii (Eigenmann and Eigenmann) euryhalyne environments. For all of them (except for A. phrygiatus that lacks sufficient detail information) an origin and diversification age of 4 m.y. was estimated (Betancur-R. et al. 2012). Therefore, the presence of Cantaurius nolfi in the Pebas Formation is interpreted as the result of early independent and unsuccessful invasions of freshwater, and does not necessarily support a marine incursion in the Amazonian region.
Fossil skulls of B. marinus Mitchill, 1815 were previously collected from the late Miocene Urumaco Formation in Venezuela in a wetland deltaic plain palaeoenvironment (Aguilera and Rodrigues de Aguilera 2001; Aguilera 2010). The genus Bagre is represented by one fossil species described here from the early Miocene (Bagre protocaribbeanus) and four valid extant species (Ferraris 2007; Marceniuk and Menezes 2007), two of them from the Pacific coast [(B. panamensis (Gill, 1863) and B. pinnimaculatus (Steindachner, 1876)] and two from the Atlantic coast (B. marinus Mitchill, 1815 and B. bagre Linnaeus, 1766). The age of diversification of these ariids was estimated between 20 and 15 m.y. by Betancur-R. et al. (2012) and is confirmed well in agreement with the age of fossil records treated here.
References
Aguilera, O. (2010). Peces fósiles del Caribe de Venezuela. In GeoBio Consultores (Ed.), Washington, DC: Gorham Printing.
Aguilera, O., & Lundberg, J. (2010). Venezuelan Caribbean and Orinocoan Neogene fish. In M. Sánchez-Villagra, O. Aguilera & F. Carlini (Eds.), Urumaco and Venezuelan paleontology (pp. 129–152). Indiana: Indiana Press University.
Aguilera, O., & Marceniuk, A. (2012). Aspistor verumquadriscutis, a new fossil species of sea catfishes (Siluriformes; Ariidae) from the upper Miocene of Venezuela. Swiss Journal of Palaeontology, 131(2), 265–274.
Aguilera, O., & Páes, E. (2012). The Pirabas Formation (Early Miocene from Brazil) and the Tropical Western Central Atlantic subprovince. Boletim do Museu Paraense Emilio Goeldi—Ciências Naturais, 7(1), 29–45.
Aguilera, O., Ramos, M. I., Paes, E., Costa, S., & Sánchez-Villagra, M. (2011). The Neogene Tropical America fish assemblage and the palaeobiogeography of the Caribbean region. Swiss Journal of Palaeontology, 130(2), 217–240.
Aguilera, O., & Rodrigues de Aguilera, D. (1999). Bathymetric of Neogene and Quaternary Teleostean fishes from the coast of Panamá and Costa Rica. Bulletins of American Paleontology, 357, 251–269.
Aguilera, O., & Rodrigues de Aguilera, D. (2001). An exceptional coastal upwelling fish assemblage in the Caribbean Neogene. Journal of Paleontology, 75(3), 732–742.
Aguilera, O., & Rodrigues de Aguilera, D. (2004). Amphi-American neogene sea catfishes (Siluriformes, Ariidae) from northern South America. Special Paper in Palaeontology, 71, 29–48.
Aguilera, O., Rodrigues de Aguilera, D., Vega, F. J., & Sánchez-Villagra, M. (2010). Mesozoic and Cenozoic decapod crustaceans from Venezuela and related ichnofossils. In M. Sánchez-Villagra, Aguilera, O., & Carlini, F. (Eds.), Urumaco and Venzuelan Paleontology: The fossil record of the Northern Neotropics (pp. 103–128). Indiana: Indiana Press University.
Arratia, G. (2003). Catfish head skeleton: an overview. In G. Arratia, B. G. Kapoor, M. Chardon & R. Diogo (Eds.), Catfishes (pp. 490–522). New Hampshire: Science Publishers, Inc. Enfield.
Arratia, G., & Gayet, M. (1995). Sensory canals and related bones of tertiary siluriform crania from Bolivia and North America and comparison with recent forms. Journal of Vertebrate Paleontology, 15, 482–505.
Bachmann, R. (2001). The Caribbean plate and the question of its formation. Institute of Geology, University of Mining and Technology Freiberg, Department of Tectonophysics (17 p). http://www.fiu/orgs/caribgeol.
Beardsley, R., Candela, J., Limeburner, R., Gleyer, W., Lentz, S. J., Castro, B. M., et al. (1995). The Me tide on the Amazon shelf. Journal of Geophysical Research, 100, 2283–2319.
Berg, L. S. (1940). Classification of fishes, both recent and fossil. Travaux de l’Institut Zoologique de l’Academie des Sciences de l’URSS (Vol. 5, pp. 87–517) (English translation, Ann Arbor, Michigan, 1947).
Betancur-R., R. (2009). Systematics and evolutionary history of sea catfishes (Siluriformes: Ariidae). Thesis PhD. Auburn University, Alabama.
Betancur-R., R., Acero P. A., Bermingham, E., & Cooke, R. (2007). Systematics and biogeography of New World sea catfishes (Siluriformes: Ariidae) as inferred from mitochondrial, nuclear and morphological evidence. Molecular Phylogenetics and Evolution, 45, 339–357.
Betancur-R., R., Orti, G., Stein, A., Marceniuk, A., & Pyron, R. (2012). Apparent signal of competition diversification after ecological transitions from marine to freshwater habits. Ecology Letters. 10.1111/j.1461-0248.2012.01802-x
Beurlen, K. (1958a). Contribuição á paleontología do estado do Pará. Crustáceos decápodes da Formação Pirabas. Boletim do Museu Paraense Emilio Goeldi, Nova Série Geológica, 5, 1–48.
Beurlen, K. (1958b). Contribuição á paleontología do estado do Pará. Um balanomorfo da Formação Pirabas. Boletim do Museu Paraense Emilio Goeldi, Nova Série Geológica, 5, 1–48.
Bleeker, P. (1862–1863). Atlas Ichthyologique des Indes Orientales Néêrlandaises, publié sous les auspices du Gouvernement Colonial Néêrlandais. Tome II. Siluroïdes, Chacoïdes et Hétérobranchoïdes, Amsterdam (pls. 49–101).
Blow, W. H. (1969). Late middle Eocene to Recent planktonic foraminiferal biostratigraphy. In P. Bronnimann & H. H. Renz (Eds.), Proceedings of the First International Conference on Planktonic Microfossils, Geneva 1967 (Vol. 1, pp. 199–422). Leiden: E.J. Brill.
Bogan, S., & Agnolin, F. L. (2011). Descripción de una nueva especie de bagre marino fóssil (Teleostei, Siluriformes, Ariidae) del Mioceno de la Provincia de Río Negro. Argentina. Papéis Avulsos de Zoología, 51(25), 373–382.
Bürgl, H. (1960). Geología de la Península de La Guajira: Servicio Geológico Nacional de Colombia. Boletín de Geología, 6(1–3), 129–168.
Carter, R. M. (1998). Two models: global sea-level change and sequence stratigraphic architecture. Sedimentary Geology, 122, 23–36.
Cloquet, H. (1816). Dictionnaire des Sciences Naturelles, Pisces accounts (Vol. 4, pp. 52–53). Paris: Imprint Le Normant.
Coates, A., Mcneill, D. F., Aaubry, M.-P., Berggren, W., & Collins, A. L. S. (2005). An introduction to the geology of the Bocas del Toro Archipelago, Panamá. Caribbean Journal of Science, 41(3), 374–391.
Costa, S. (2011). Ictiólitos da Formação Pirabas, Mioceno do Pará, Brasil, e suas implicações paleoecológicas. Tese de Doutorado. Universidade Federal do Pará, Instituto de Geociências.
Díaz de Gamero, M. L. (1974). Microfauna y edad de la Formación Cantaure, Península de Paraguaná, Venezuela. Boletín Informativo de la Asociación Venezolana de Geología, Minería y Petróleo, 17, 41–47.
Díaz de Gamero, M. L. (1996). The changing course of the Orinoco river during the Neogene: a review. Palaeogeography, Palaeoclimatology, Palaeoecology, 123, 385–402.
Diretoria de Hidrografia e Navegação-DHN. (2011). Centro de Hidrografia da Marinha do Brasil, Banco Nacional de dados Oceanográficos. http://www.mar.mil.br/dhn/chm/tabuas/index.htm. Accessed 24 Aug 2011.
Eschmeyer, W. N. (Ed.). (2013). Catalog of Fishes. San Francisco: California Academy of Sciences (http://research.calacademy.org/research/ichthyology/catalog/fishcatmain.asp). Electronic version accessed 03 February 2013.
Fernandes, J. M. G. (1984). Paleoecology of Pirabas Formation, Pará state. In 33° Congresso Brasileiro de Geologia. Anais da Academia Brasileira de Ciências SBG, 1, 330–340.
Fernandes, J. M. G. (1988). Biostratigraphy of Pirabas Formation, Pará state. In 35° Congresso Brasileiro de Geologia. Anais da Academia Brasileira de Ciências, SBG, 1, 2376–2382.
Fernandes, M. J. G., & Távora, V. A. (1990). Estudo dos Foraminíferos da Formação Pirabas procedentes do furo CB-UFPa-P1 (85), município de Capanema, estado do Pará. In 36° Congresso Brasileiro de Geologia. Anais da Academia Brasileira de Ciências, SBG, 1, 470–475.
Ferraris, C. J. (2007). Checklist of catfishes, recent and fossil (Osteichthyes: Siluriformes), and catalogue of siluriform primary types. Zootaxa, 1418, 1–628.
Ferreira, C. S., Macedo, A. C. M., & Assis, J. F. P. (1978). A Formação Pirabas no estado do Pará. Novo registro de subsuperfície: Belém (Furo 4be-01-Pa, Cprm). Anais da Academia Brasileira de Ciências, 50(3), 427–427.
Ferreira-Penna, D. S. (1876). Breves notícias sobre os sambaquís do Pará. Arquivo do Museu Nacional, 1, 85–89.
Figueiredo, J., Hoorn, C., van der Ven, P., & Soares, E. (2009). Late Miocene onset of the Amazon River and the Amazon deep-sea fan: evidence from the Foz do Amazonas Basin. Geology, 37, 619–622.
Frizzell, D. L. (1965). Otolith of new fish (Vorhisia vulpes, n. gen., n. sp. Siluroidei ?) fom the upper Cretaceous of South Dakota. Copeia, 1965(2), 178–181.
Gayet, M. & Meunier, F. J. (2003). Palaeontology and Palaeogeography of catfishes. In G. Arratia, B. G. Kapoor, M. Chardon & R. Diogo (Eds.), Catfishes (pp. 490–522). New Hampshire: Science Publishers, Inc. Enfield.
Gill, T. N. (1863). Descriptive enumeration of a collection of fishes from the western coast of Central America, presented to the Smithsonian Institution by Captain John M. Dow. Proceedings of the Academy of Natural Sciences of Philadelphia, 15, 162–174.
Góes, A. M., Rossetti, D., Nogueira, A., & Toledo, P. M. (1990). Modelo deposicional preliminar da Formação Pirabas no Nordeste do Estado do Pará. Boletim do Museu Paraense Emilio Goeldi, Ciências da Terra, 2, 3–15.
Gregory-Wodzicki, K. M. (2000). Uplift history of the Central and Northern Andes: a review. Geological Society of America Bulletin, 112(7), 1091–1105.
Hoorn, C., Guerrero, J., Sarmiento, G. A., & Lorente, M. A. (1995). Andean tectonics as a cause for changing drainage patterns in Miocene northern South America. Geology, 23, 237–240.
Hoorn, C., Wesselingh, F. P., Ter Steege, H., Bermudez, M. A., Mora, A., Sevink, J., et al. (2010). Amazonia through time: Andean uplift, climate change, landscape evolution, and biodiversity. Science, 330, 927–931.
Huddleston, R. W., & Savoie, K. M. (1983). Teleostean otoliths from the Late Cretaceous (Maestrichtian age) Severn Formation of Maryland. Procceding of the Biological Society of Washington, 96(4), 658–663.
Hunter, V. F., & Bartok, P. (1974). The age and correlation of the tertiary sediments of the Paraguaná Peninsula, Venezuela. Boletín Informativo de la Asociación Venezolana de Geología, Minería y Petróleo, 17, 143–154.
Irving, E. (1972). Mapa Geológico de la Península de la Guajira (Compilación). Instituto Nacional de Investigaciones Geológicas-Mineras, U.S. Geological Survey. Mapa Geológico.
Iturralde-Vinent. M. A. (2004–2005). La Paleogeografía del Caribe y sus implicaciones para la biogeografía histórica. Revista del Jardín Botánico Nacional, 25–26, 49–78.
Iturralde-Vinent, M. A., & MacPhee, R. D. E. (1999). Paleogeography of the Caribbean region: Implications for Cenozoic biogeography. Bulletin of the American Museum of Natural History, 238, 1–95, 22 figs, 2 app.
Jackson, J., Jung, P., Coates, A., & Collins, L. (1993). Diversity and extinction of Tropical American mollusk and emergence of the Isthmus of Panamá. Science, 260, 1624–1626.
Johns, W. E., Lee, T. N., Beardsley, R. C., Candela, J., Limeburner, R., & Castro, B. (1998). Annual cycle and variability of the North Brazil Current. Journal of Physical Oceanography, 28(1), 103–128.
Johns, W. E., Tamara, L., Townsend, T. L., Fratantoni, D. M., & Wilson, W. D. (2002). On the Atlantic inflow to the Caribbean Sea. Deep-Sea Research, 1(49), 211–243.
Johnson, K. G., Budd, A. F., & Jackson, J. B. C. (2008). Coral reef development was independent of coral diversity in the Caribbean over 28 million years. Science, 319, 1521–1522.
Johnson, K., Sánchez-Villagra, M., & Aguilera, O. (2009). The Oligocene–Miocene transition on coral reefs in the Falcón basin (NW Venezuela). Palaios, 24, 59–69.
Jordan, D. S., & Gilbert, C. H. (1882). A review of the siluroid fishes found on the Pacific coast of tropical America, with descriptions of three new species. Bulletin of the United States Fish Commission, 2, 34–54.
Jung, P. (1965). Miocene mollusca from the Paraguaná Peninsula, Venezuela. Bulletins of American Paleontology, 49, 389–652.
Klaus, J. S., McNeill, D. F., Budd, A. F., & Coates, A. G. (2011). Neogene reef coral assemblages of the Bocas del Toro region, Panamá: the rise of Acropora palmata. Coral Reefs. doi:10.1007/s00338-011-0835-2
Kominz, M. A., Browning, J. V., Miller, K. G., Sugarman, P. J., Mizintseva, S., & Scotese, C. R. (2008). Late Cretaceous to Miocene sea-level estimates from the New Jersey and Delaware coastal plain coreholes: an error analysis. Basin Research, 20, 211–226.
Landau, B., Vermeij, G., & Marques da Silva, C. (2008). Southern Caribbean Neogene palaeobiogeography revisited. New data from the Pliocene of Cubagua, Venezuela. Palaeogeography, Palaeoclimatology, Palaeoecology, 257, 445–461.
Landini, W., Bianucci, G., Carnevale, G., Ragaini, L., Sorbini, C., Valleri, G., et al. (2002). Late Pliocene fossils of Ecuador and their role in the development of the Panamic bioprovince after the rising of Central American Isthmus. Canadian Journal of Earth Sciences, 39, 27–34.
Latrubesse, E. M., Cozzuol, M., Silva-Caminha, S. A. F., Rigsby, C. A., Absy, M. L., & Jaramillo, C. A. (2010). The Late Miocene paleogeography of the Amazon Basin and the evolution of the Amazon river system. Earth-Science Reviews, 99, 99–124.
Lorente, M. A. (1997). Castillo Formation. In Committee for stratigraphy and nomenclature. Stratigraphical Lexicon of Venezuela. http://www.pdvsa.com/lexico/lexicohi.htm.
Marceniuk, A. P. (2007). Description of Cathorops manglarensis, a new species from the Colombian Pacific, with redescription of Cathorops multiradiatus (Siluriformes; Ariidae). Zootaxa, 1529, 33–48.
Marceniuk, A. P., & Menezes, N. A. (2007). Systematics of the family Ariidae (Ostariophysi, Siluriformes), with a redefinition of the genera. Zootaxa, 1416, 1–126.
Marceniuk, A. P., & Betancur-R., R. (2008). Revision of the species of the genus Cathorops (Siluriformes: Ariidae) from Mesoamerica and the Central American Caribbean, with description of three new species. Neotropical Ichthyology, 6(1), 25–44.
Marceniuk, A. P., Betancur-R, R., Acero, A., & Muriel-Cunha, J. (2012a). Review of the genus Cathorops (Siluriformes: Ariidae) from the Caribbean and Atlantic South America, with description of a new species. Copeia, 1, 77–97.
Marceniuk, A. P., Menezes, N. A., & Britto, M. R. (2012b). Phylogenetic analysis of the family Ariidae (Ostariophysi, Siluriformes), with a hypothesis on the monophyly and relationships of the genera. Zoological Journal of the Linnean Society, 165, 534–669.
Martinod, J., Husson, L., Roperch, P., Guillaume, B., & Espurt, N. (2010). Horizontal subduction zones, convergence velocity and the building of the Andes. Earth and Planetary Science Letters, 299, 299–309.
Maury, C. J. (1925). Fósseis terciários do Brasil com descrição de novas formas Cretáceas. Serviço Geológico e Mineralógico do Brasil, 4, 1–665.
Mitchill, S. L. (1815). The fishes of New York described and arranged. Transactions of the Literary and Philosophical Society of New York, 1, 355–492. (pls. 1–6).
Mo, T. (1991). Anatomy, relationships and systematics of the Bagridae (Teleostei: Siluroidei) with a hypothesis of siluroid phylogeny. Theses Zoologicae (Vol. 17). Koenigstein: Koeltz. (63 figs).
Mohriak, W. V. (2003). Bacias sedimentares da margem continental Brasileira. In L. A. Bizzi, C. Schobbenhaus, R. M. Vidotti, & J. H. Gonçalves (Eds.), Geologia Tectônica e Recursos Minerais do Brasil. Brasília: CPRM.
Monsch, K. A. (1998). Miocene fish fauna from northwestern Amazonia basin (Colombia, Peru, Brazil) with evidence of marine incursions. Palaeogeography, Palaeoclimatology and Palaeoecology, 14, 31–50.
Nolf, D. (1976). Les otolithes dês Téléostéens néogénes de Trinidad. Eclogae Geologicae Helvetiae, 69, 703–742.
Nolf, D., & Aguilera, O. (1998). Fish otoliths from the Cantaure Formation (early Miocene of Venezuela). Bulletin de l’Institut Royal des Sciences Naturelles de Belgique, Sciences de la Terre, 68, 237–262.
Nolf, D., & Stringer, G. L. (1992). Neogene paleontology in the northern Dominican Republic: Otoliths of teleostean fishes. Bulletin of American Paleontology, 102, 41–81.
O′Dea, A., Jackson, J., Fortunato, H., Smith, T., D′Croz, L., Johnson, K., & Todd, J. A. (2007). Environmental change preceded Caribbean extinction by 2 million years. PNAS, 104, 5263–5704.
Ohe, F. (2006). Skulls and otoliths of eleven sea catfishes (Family Ariidae) from Malaysia and one species related to them from the East China Sea. Natural Environmental Science Research, 19, 11–28.
Paula-Couto, C. (1967). Contribuição à paleontologia do estado de Pará. Um sirênio na formação Pirabas. Atas do 1° Simpósio sobre a Biota Amazônica, 1, 345–357.
Petri, S. (1954). Foraminíferos fósseis da Bacia de Marajó. Boletim da Faculdade de Filosofia. Ciências e Letras da Universidade de São Paulo (Geologia), 134, 21–38.
Petri, S. (1957). Foraminíferos Miocênicos da Formação Pirabas. Boletim da Faculdade de Filosofia Ciências e Letras da Universidade de São Paulo (Geologia), 216, 1–172.
Pindell, J., Kennan, L., Maresch, W. V., Walter, V., Stanek, K.-P., Draper, G., et al. (2005). Plate-kinematics and crustal dynamics of circum-Caribbean arc continent interactions: tectonic controls on basin development in Proto-Caribbean margins. Geological Society of America Special Paper, 394, 7–52.
Renz, O. (1960). Geología de la parte sureste de la Península de la Guajira (República de Venezuela). III Congreso Geológico Venezolano, Caracas, 1959. Memorias del Boletín de Geología, 3(1), 317–347.
Rey, O. (1996). Estratigrafía de la Península de Paraguaná, Venezuela. Revista de la Facultad de Ingeniería, 11, 35–45.
Rollins, J. (1965). Stratigraphy and structure of the Guajira Peninsula, northwestern Venezuela and northeastern Colombia. University of Nebraska Studies, New Series, 30, 1–1102.
Rossetti, D. (2001). Late Cenozoic sedimentary evolution in northeastern Pará, Brazil, within the context of sea level changes. Journal of South American Earth Sciences, 14, 77–89.
Rossetti, D. (2006). Evolução sedimentar miocênica nos estados do Pará e Maranhão. Revista do Instituto de Geociências, USP Série Científica, 6(2), 7–18.
Rossetti, D., & Góes, A. (2004). Geologia. In D. Rossetti & A. Góes (Eds.), O Neogeno da Amazônia Oriental (pp. 13–52). Belém: Museu Paraense Emilio Goeldi.
Royero, R. (1987). Morfología de la aleta dorsal en los bagres (Teleostei: Siluriformes), con especial referencia a las familias americanas. Trabajo especial de grado. Caracas: Universidad Central de Venezuela, Facultad de Ciencias.
Sánchez-Villagra, M. R., & Clack, J. A. (Eds.) (2004). Fossils of the Miocene Castillo Formation, Venezuela: Contributions in Neotropical Palaeontology. Special Papers in Palaeontology (Vol. 71, 112 pp.).
Sánchez-Villagra, M. R., Burnham, R. J., Campbell, D. C., Feldmann, R. M., Gaffney, E. S., Kay, R. F., et al. (2000). A new near-shore marine fauna and flora from the early Neogene of Northwestern Venezuela. Journal of Paleontology, 74, 957–968.
Sánchez-Villagra, M. R., Gasparini, Z., Lozsán, R., Moody, J. M., & Uhen, M. D. (2001). New discoveries of vertebrates from a near-shore marine fauna from the early Miocene of northwestern Venezuela. Paläontologische Zeitschrift, 75(2), 227–232.
Santos, M. E. C. M. (1958). Equinóides miocênicos da Formação Pirabas. Boletim da Divisão de Geologia e Mineralogia, 179, 1–24.
Santos, M. E. C. M. (1967). Equinóides miocenicos da Formação Pirabas. Atas 1° Simpósio sobre a Biota Amazônica, 1, 407–410.
Santos, R. S., & Travassos, S. (1960). Contribuição a paleontologia do estado do Pará. Peixes fósseis da Formação Pirabas. Monografia da Divisão de Geologia e Mineralogia, 16, 1–35.
Smith, C. J., Collins, L. S., Jaramillo, C., & Quiroz, L. I. (2010). Marine paleoenvironments of Miocene-Pliocene formations of north-central Falcon state, Venezuela. Journal of Foraminiferal Research, 40(3), 266–282.
Steindachner, F. (1876). Ichthyologische Beiträge, IV. Sitzungsberichte der Kaiserlichen Akademie der Wissenschaften, Mathematisch-aturwissenschaftlichen Classe, Wien, Abt. 1. Botanik, Zoologie, Anatomie, Geologie und Paläontologie, 72, 551–616. (pls. 1–13).
Távora, V., & Fernandes, J. M. (1999). Estudio de los foraminíferos de la Formación Pirabas (Mioceno Inferior), estado de Pará, Brasil y su correlación con faunas del Caribe. Revista Geológica de America Central, 22, 63–74.
Távora, V., Santos, A. A., & Araujo, R. N. (2010). Localidades fossilíferas da Formação Pirabas (Mioceno Inferior). Boletim do Museu Paraense Emilio Goeldi, Ciências Naturais, 5(2), 207–224.
Thomas, D. J., & MacDonald, W. D. (1970). The Cantaure Formation of the Paraguaná Peninsula. Boletin Informativo de la Asociación Venezolana de Geología. Minería y Petróleo, 13, 177–179.
Wheeler, C. B. (1960). Estratigrafía del Oligoceno y Mioceno Inferior de Falcón occidental y nororiental. Memorias III Congreso Geológico Venezolano, 1, 407–465.
Wheeler, C. B. (1963). Oligocene and Lower Miocene stratigraphy of Western and Northeastern Falcon Basin, Venezuela. Bulletin of the American Association of Petroleum Geologists, 4, 35–68.
White, C. A. (1887). Contribuições à Paleontologia do Brasil. Arquivos do Museu Nacional, 7, 1–273.
Acknowledgments
This research was supported by the Museum Paraense Emilio Goeldi, the Conselho Nacional de Desenvolvimento Científico e Tecnológico (Process CNPq 311783/2011-2 to OA and CNPq 565046/2010-1 to HM-S) in Brazil, the Universidad Francisco de Miranda in Venezuela, and the Smithsonian Tropical Research Institute in Panamá. We thank Dione Rodrigues de Aguilera for preparation of the fossil and suggestions. Julio Reyes and Ana Cristina Oliveira helped us with the skeletal preparation. Vladimir Zapata, Federico Moreno and Jorge Moreno for the geological survey and for collecting the otolith during a field trip to the La Guajira Peninsula in Colombia. Marcelo Sánchez for collecting the otolith during a field trip to Cerro La Cruz in Venezuela. David Santana for the early review of the manuscript. Zoneibe Luz helped with otolith photographs. Wolmar Wosiacki helped us with skull photograph and supported us with the access to the fish collection in the MPEG. Guillermo Aguilera supported with the 3D and 2D platform, figures and plates design. Jean dos Santos-Lobato helped us with the tomography and image captions. We thank Alexander Marceniuk, Ricardo Betancur-R. and Marcelo Sánchez-Villagra for comments and suggestions. The authors would like to thank especially Werner Schwarzhans, Bettina Reichenbacher, Florian Witzmann, anonymous reviewers and editors for corrections, comments and suggestions that improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Aguilera, O.A., Moraes-Santos, H., Costa, S. et al. Ariid sea catfishes from the coeval Pirabas (Northeastern Brazil), Cantaure, Castillo (Northwestern Venezuela), and Castilletes (North Colombia) formations (early Miocene), with description of three new species. Swiss J Palaeontol 132, 45–68 (2013). https://doi.org/10.1007/s13358-013-0052-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13358-013-0052-4